
Effect of Enterococcus hirae GS22 Fermentation-Assisted Extraction on the Physicochemical and Bioactivities of Sea Cucumber Intestinal Polysaccharides

 , , and
, , and
Abstract
1. Introduction
2. Results and Discussion
2.1. Bacterial Growth and pH Variation During Fermentation
2.2. Extraction Yield and Chemical Compositions of the Polysaccharides
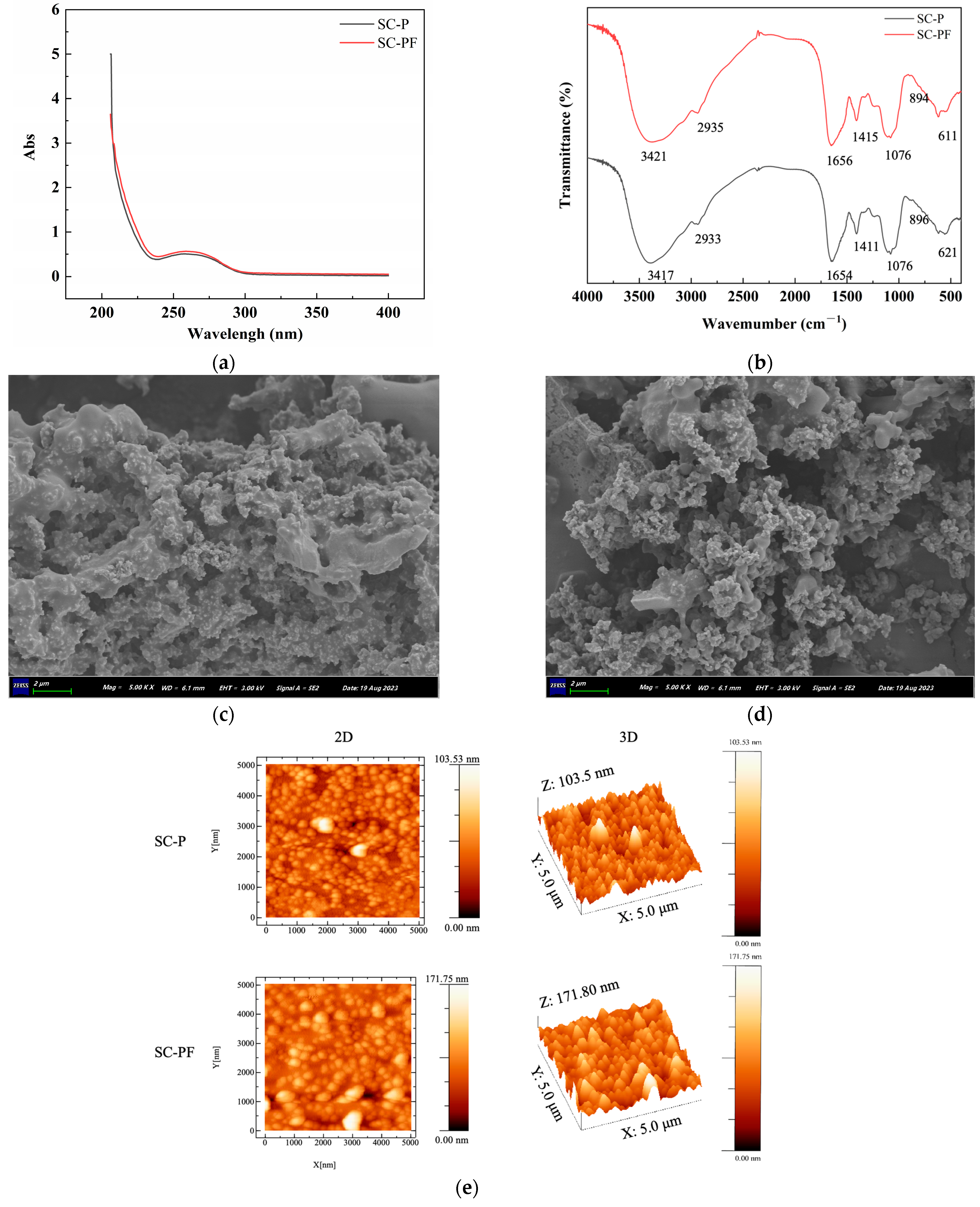
2.3. The Spectra and Morphology Analysis of the Polysaccharides
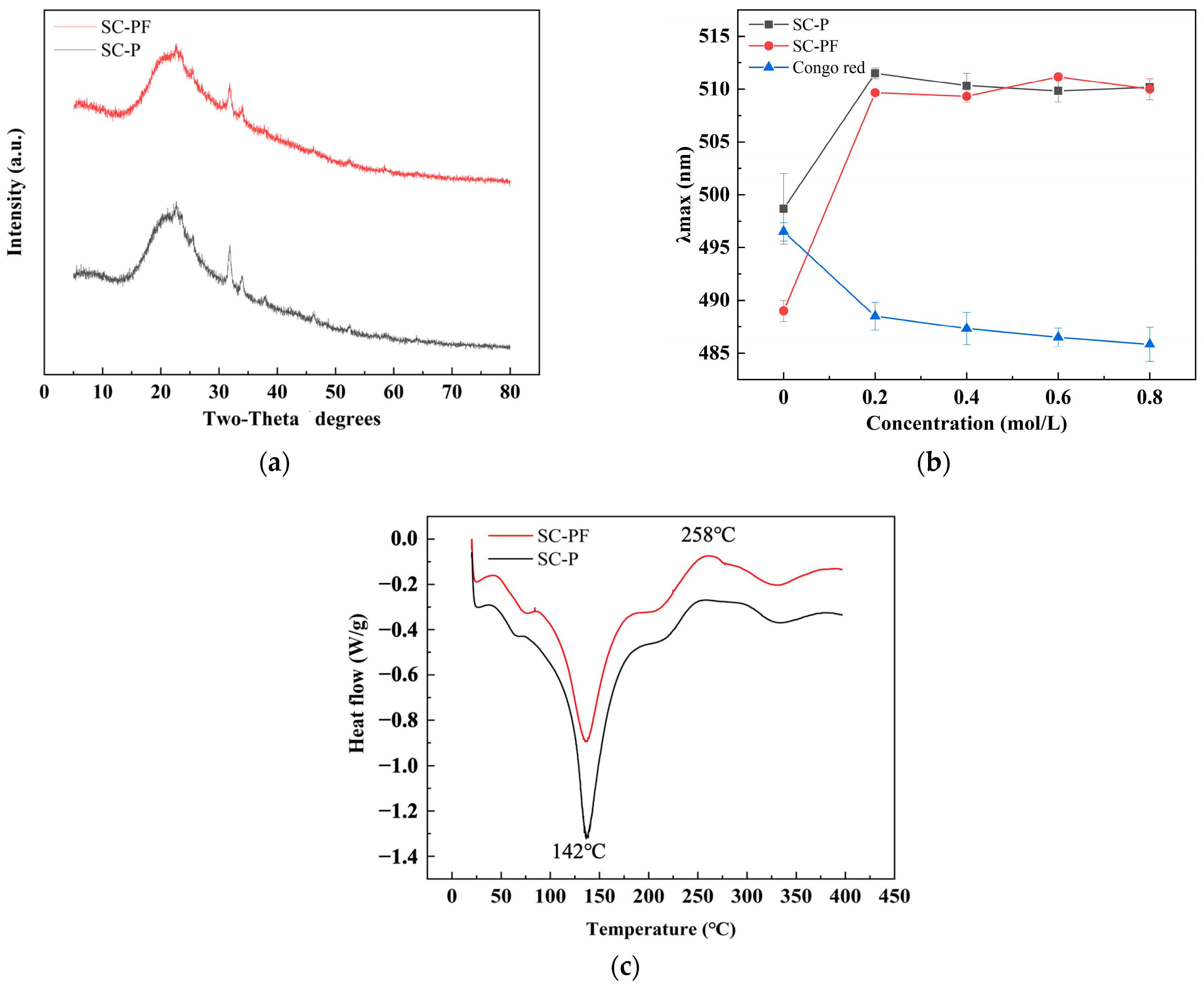
2.4. The Structure and Thermal Analysis of the Polysaccharides
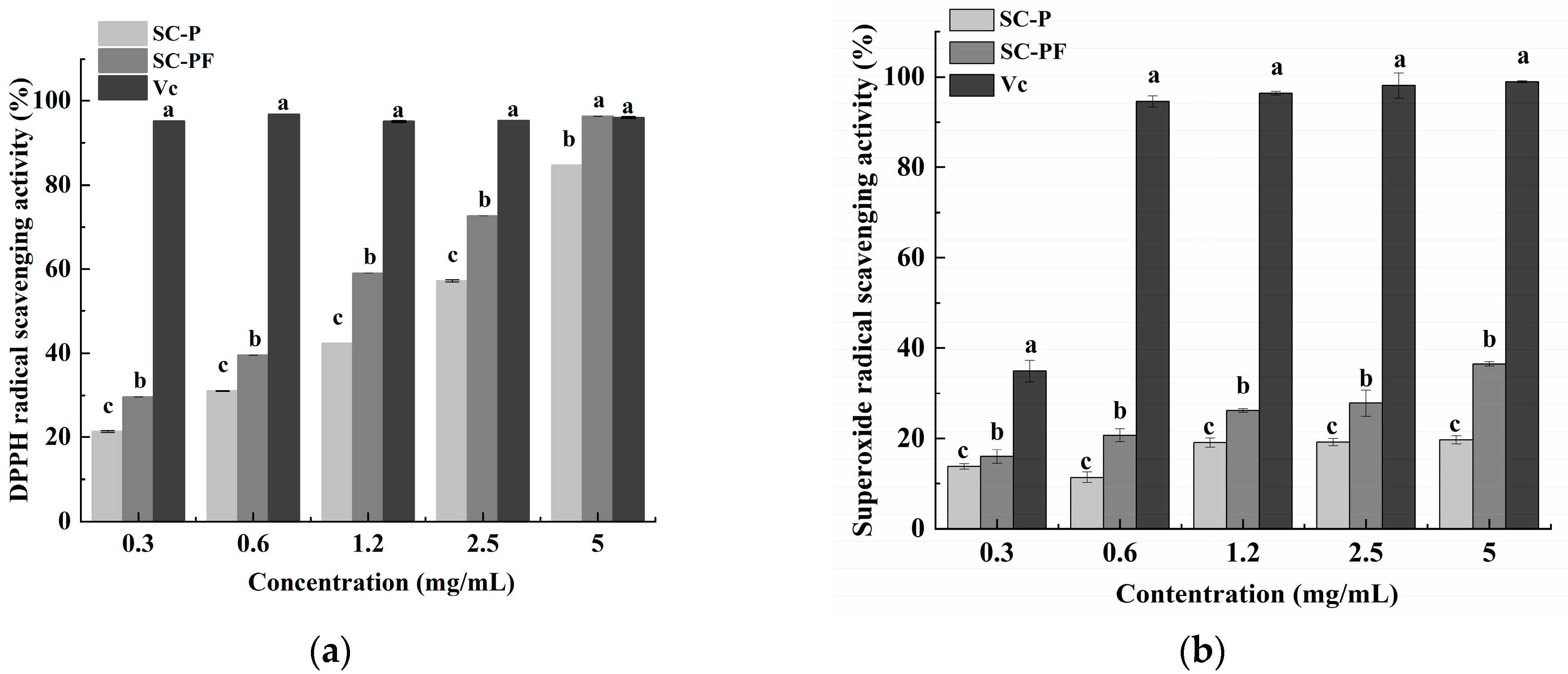
2.5. The Antioxidant Characteristics of the Polysaccharides
2.6. Hypoglycemic and Cholesterol-Absorbing Activities
3. Materials and Methods
3.1. Materials
3.2. SI Fermentation
3.3. Polysaccharide Extraction
3.4. Chemical Composition Analysis of the Polysaccharides
3.5. Characterization of Polysaccharide Molecular Mass Distribution and Monosaccharide Composition
3.6. Particle Size and Zeta Potential Profiling
3.7. Spectral and Morphology Analysis
3.7.1. Spectral Analysis
3.7.2. Scanning Electron Microscopy (SEM) Analysis
3.7.3. Atomic Force Microscopy (AFM) Analysis
3.8. Structural and Thermal Characteristic Analysis
3.8.1. X-Ray Diffraction (XRD) Analysis
3.8.2. Congo Red Analysis
3.8.3. Differential Scanning Calorimetric (DSC) Analysis
3.9. Functional Analysis
3.9.1. Determination of Antioxidant Activity
3.9.2. Inhibition Capacities of α-Amylase
3.9.3. Inhibition Capacities of α-Glucosidase
3.9.4. Glucose Absorption Capacity (GAC)
3.9.5. Cholesterol Absorption Capacity (CAC)
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Y.-C.; Xue, C.-H.; Zhang, T.-T.; Wang, Y.-M. Saponins from Sea Cucumber and Their Biological Activities. J. Agric. Food Chem. 2018, 66, 7222–7237. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Dave, D.; Shahidi, F. Antioxidant Potential of Sea Cucumbers and Their Beneficial Effects on Human Health. Mar. Drugs 2022, 20, 521. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.-X.; Wu, Y.-C.; Liu, Y.; Lv, S.-Z.; You, Y.; Zhou, Z.-L.; Chen, X.; Li, H.-J. Structure and Hypoglycemic Effect of a Neutral Polysaccharide Isolated from Sea Cucumber Stichopus japonicus. Int. J. Biol. Macromol. 2022, 216, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Pan, R.; Deng, X.; Chen, Y.; Zhao, G.; Wang, C. Separation, Purification, Anticoagulant Activity and Preliminary Structural Characterization of Two Sulfated Polysaccharides from Sea Cucumber Acaudina molpadioidea and Holothuria nobilis. Process Biochem. 2014, 49, 1352–1361. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Zhi, Z.; Wei, C.; Wang, W.; Ding, T.; Ye, X.; Hu, Y.; Linhardt, R.J.; Chen, S. Macromolecular Properties and Hypolipidemic Effects of Four Sulfated Polysaccharides from Sea Cucumbers. Carbohydr. Polym. 2017, 173, 330–337. [Google Scholar] [CrossRef]
- Qin, Y.; Yuan, Q.; Zhang, Y.; Li, J.; Zhu, X.; Zhao, L.; Wen, J.; Liu, J.; Zhao, L.; Zhao, J. Enzyme-Assisted Extraction Optimization, Characterization and Antioxidant Activity of Polysaccharides from Sea Cucumber Phyllophorus proteus. Molecules 2018, 23, 590. [Google Scholar] [CrossRef]
- Lu, X.; Wang, M.; Yue, H.; Feng, X.; Tian, Y.; Xue, C.; Zhang, T.; Wang, Y. Novel Peptides from Sea Cucumber Intestines Hydrolyzed by Neutral Protease Alleviate Exercise-induced Fatigue via Upregulating the Glutaminemediated Ca2+/Calcineurin Signaling Pathway in Mice. J. Food Sci. 2024, 89, 1727–1738. [Google Scholar] [CrossRef]
- Yue, H.; Tian, Y.; Feng, X.; Bo, Y.; Xue, C.; Dong, P.; Wang, J. Novel Peptides Derived from Sea Cucumber Intestine Promotes Osteogenesis by Upregulating Integrin-Mediated Transdifferentiation of Growth Plate Chondrocytes to Osteoblasts. J. Agric. Food Chem. 2022, 70, 13212–13222. [Google Scholar] [CrossRef]
- Yang, D.; Lin, F.; Huang, Y.; Ye, J.; Xiao, M. Separation, Purification, Structural Analysis and Immune-Enhancing Activity of Sulfated Polysaccharide Isolated from Sea Cucumber Viscera. Int. J. Biol. Macromol. 2020, 155, 1003–1018. [Google Scholar] [CrossRef]
- Ye, J.; Zheng, L.; Pan, W.; Huang, Y.; Zhang, N.; Yang, D.; Yang, Y.; Zheng, B.; Zhang, X.; Xiao, M. Sulfated Polysaccharide from Apostichopus japonicus Viscera Exhibits Anti-Inflammatory Properties in Vitro and in Vivo. Int. J. Biol. Macromol. 2024, 280, 135500. [Google Scholar] [CrossRef]
- Chen, G.; Fang, C.; Ran, C.; Tan, Y.; Yu, Q.; Kan, J. Comparison of Different Extraction Methods for Polysaccharides from Bamboo Shoots (Chimonobambusa quadrangularis) Processing by-Products. Int. J. Biol. Macromol. 2019, 130, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Wang, W.; Bian, J.; Gao, Y.; Hao, Z.; Tan, J. Recent Advances in Medicinal and Edible Homologous Polysaccharides: Extraction, Purification, Structure, Modification, and Biological Activities. Int. J. Biol. Macromol. 2022, 222, 1110–1126. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Huang, Y.; Islam, S.; Fan, B.; Tong, L.; Wang, F. Influence of the Degree of Hydrolysis on Functional Properties and Antioxidant Activity of Enzymatic Soybean Protein Hydrolysates. Molecules 2022, 27, 6110. [Google Scholar] [CrossRef] [PubMed]
- Doungapai, C.; Siriwoharn, T.; Malila, Y.; Autsavapromporn, N.; Makkhun, S.; Yarnpakdee, S.; Jantanasakulwong, K.; Regenstein, J.M.; Wangtueai, S. UV-B Protective and Antioxidant Activities of Protein Hydrolysate From Sea Cucumber (Holothuria scabra) Using Enzymatic Hydrolysis. Front. Mar. Sci. 2022, 9, 892255. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, G.; Liu, C.; Sun, Z.; Li, R.; Gao, J.; Li, M.; Sun, L. The Structural Characteristics and Bioactivity Stability of Cucumaria Frondosa Intestines and Ovum Hydrolysates Obtained by Different Proteases. Mar. Drugs 2023, 21, 395. [Google Scholar] [CrossRef]
- Wang, Y.; Song, Y.; Chang, Y.; Liu, Y.; Chen, G.; Xue, C. Dynamic Changes of Peptidome and Release of Polysaccharide in Sea Cucumber (Apostichopus japonicus) Hydrolysates Depending on Enzymatic Hydrolysis Approaches. Food Sci. Hum. Wellness 2022, 11, 1331–1341. [Google Scholar] [CrossRef]
- Ma, E.-H.; Rathnayake, A.U.; Lee, J.K.; Lee, S.-M.; Byun, H.-G. Characterization of β-Secretase Inhibitory Extracts from Sea Cucumber (Stichopus japonicus) Hydrolysis with Their Cellular Level Mechanism in SH-SY5Y Cells. Eur. Food Res. Technol. 2021, 247, 2039–2052. [Google Scholar] [CrossRef]
- Wei, C.; He, P.; He, L.; Ye, X.; Cheng, J.; Wang, Y.; Li, W.; Liu, Y. Structure Characterization and Biological Activities of a Pectic Polysaccharide from Cupule of Castanea henryi. Int. J. Biol. Macromol. 2018, 109, 65–75. [Google Scholar] [CrossRef]
- Huang, F.; Hong, R.; Zhang, R.; Yi, Y.; Dong, L.; Liu, L.; Jia, X.; Ma, Y.; Zhang, M. Physicochemical and Biological Properties of Longan Pulp Polysaccharides Modified by Lactobacillus fermentum Fermentation. Int. J. Biol. Macromol. 2019, 125, 232–237. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Ding, Y.; Li, S.; Sang, X.; Li, T.; Zhao, Q.; Yu, S. Structure, in Vitro Digestive Characteristics and Effect on Gut Microbiota of Sea Cucumber Polysaccharide Fermented by Bacillus subtilis Natto. Food Res. Int. 2023, 169, 112872. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, R.; Wang, Y.; An, X.; Liu, N.; Song, M.; Yang, Y.; Yin, N.; Qi, J. Characterization and Antioxidant Activity of Wheat Bran Polysaccharides Modified by Saccharomyces cerevisiae and Bacillus subtilis Fermentation. J. Cereal Sci. 2021, 97, 103157. [Google Scholar] [CrossRef]
- Melo, C.d.C.S.; Freire, A.d.S.; Galdeano, M.A.; Costa, C.F.d.; Gonçalves, A.P.D.d.O.; Dias, F.S.; Menezes, D.R. Probiotic Potential of Enterococcus hirae in Goat Milk and Its Survival in Canine Gastrointestinal Conditions Simulated in Vitro. Res. Vet. Sci. 2021, 138, 188–195. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Hassan, S.E.-D.; Alrefaey, H.M.A.; El-Belely, E.F.; Elsakhawy, T.; Fouda, A.; Desouky, S.G.; Khattab, S.M.R. Subsequent Improvement of Lactic Acid Production from Beet Molasses by Enterococcus hirae Ds10 Using Different Fermentation Strategies. Bioresour. Technol. Rep. 2021, 13, 100617. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, W.; Yao, J.; Niu, J. Production, Purification, Characterization, and Biological Properties of Rhodosporidium paludigenum Polysaccharide. PLoS ONE 2021, 16, e0246148. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yang, C.; Liu, R.; Sui, W.; Zhu, Q.; Jin, Y.; Wu, T.; Zhang, M. The Relationship between Preparation and Biological Activities of Animal-Derived Polysaccharides: A Comprehensive Review. Foods 2024, 13, 173. [Google Scholar] [CrossRef]
- Cheng, Y.; Huang, X.; Li, L.; Liu, L.; Zhang, C.; Fan, X.; Xie, Y.; Zou, Y.; Geng, Z.; Huang, C. Effects of Solid Fermentation on Polygonatum cyrtonema Polysaccharides: Isolation, Characterization and Bioactivities. Molecules 2023, 28, 5498. [Google Scholar] [CrossRef]
- Tao, S.; Ren, Z.; Yang, Z.; Duan, S.; Wan, Z.; Huang, J.; Liu, C.; Wei, G. Effects of Different Molecular Weight Polysaccharides From Dendrobium Officinale Kimura & Migo on Human Colorectal Cancer and Transcriptome Analysis of Differentially Expressed Genes. Front. Pharmacol. 2021, 12, 704486. [Google Scholar] [CrossRef]
- Zou, P.; Ma, S.; Yuan, Y.; Ma, J.; Yang, X.; Hu, X.; Meng, Q.; Jing, C.; Li, Y. A Glucomannan Produced by Bacillus velezensis HY23 and Its Growth Promoting Effect on Soybeans under Salt Stress. Int. J. Biol. Macromol. 2024, 275, 133474. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.-Y.; Liu, R.-Z.; Wang, G.-Y. Culturable Microorganisms Associated with Sea Cucumbers and Microbial Natural Products. Mar. Drugs 2021, 19, 461. [Google Scholar] [CrossRef]
- Mohal, M.M.; Sraboni, F.S.; Islam, S.; Zaman, S.; Uddin, M.S.; Saleh, M.A. Functional Characterization and Biotechnological Applications of Exopolysaccharides Produced by Newly Isolated Enterococcus hirae MLG3-25–1. Int. Microbiol. 2024, 1–12. [Google Scholar] [CrossRef]
- Li, M.; Zhang, H.; Hu, X.; Liu, Y.; Liu, Y.; Song, M.; Wu, R.; Wu, J. Isolation of a New Polysaccharide from Dandelion Leaves and Evaluation of Its Antioxidant, Antibacterial, and Anticancer Activities. Molecules 2022, 27, 7641. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Jia, R.-B.; Ou, Z.-R.; Li, Z.-R.; Zhao, M.; Luo, D.; Lin, L. Comparative Study on the Structural Characterization and α-Glucosidase Inhibitory Activity of Polysaccharide Fractions Extracted from Sargassum fusiforme at Different PH Conditions. Int. J. Biol. Macromol. 2022, 194, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, S. Effects of Separation and Purification on Structural Characteristics of Polysaccharide from Quinoa (Chenopodium quinoa willd). Biochem. Biophys. Res. Commun. 2020, 522, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Gao, Y.; Pan, Y.; Zou, Y.; He, M.; He, C.; Li, L.; Yin, Z.; Lv, C. Purification, Chemical Characterization and Antioxidant Activities of Polysaccharides Isolated from Mycena dendrobii. Carbohydr. Polym. 2019, 203, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hu, M.; Wang, C.; Yang, Y.; Chen, J.; Ding, J.; Guo, W. Characterization and in Vitro Antitumor Activity of Polysaccharides from the Mycelium of Sarcodon aspratus. Int. J. Biol. Macromol. 2013, 52, 52–58. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhang, X.; Ma, M.; Long, T.; Xiao, C.; Zhang, J.; Liu, J.; Zhao, L. Immunoenhancing Glucuronoxylomannan from Tremella aurantialba Bandoni et Zang and Its Low-Molecular-Weight Fractions by Radical Depolymerization: Properties, Structures and Effects on Macrophages. Carbohydr. Polym. 2020, 238, 116184. [Google Scholar] [CrossRef]
- Rozi, P.; Abuduwaili, A.; Ma, S.; Bao, X.; Xu, H.; Zhu, J.; Yadikar, N.; Wang, J.; Yang, X.; Yili, A. Isolations, Characterizations and Bioactivities of Polysaccharides from the Seeds of Three Species Glycyrrhiza. Int. J. Biol. Macromol. 2020, 145, 364–371. [Google Scholar] [CrossRef]
- Liu, X.; Ren, Z.; Yu, R.; Chen, S.; Zhang, J.; Xu, Y.; Meng, Z.; Luo, Y.; Zhang, W.; Huang, Y.; et al. Structural Characterization of Enzymatic Modification of Hericium erinaceus Polysaccharide and Its Immune-Enhancement Activity. Int. J. Biol. Macromol. 2021, 166, 1396–1408. [Google Scholar] [CrossRef]
- Shen, S.; Jia, S.; Wu, Y.; Yan, R.; Lin, Y.-H.; Zhao, D.; Han, P. Effect of Culture Conditions on the Physicochemical Properties and Antioxidant Activities of Polysaccharides from Nostoc Flagelliforme. Carbohydr. Polym. 2018, 198, 426–433. [Google Scholar] [CrossRef]
- Hui, H.; Jin, H.; Li, X.; Yang, X.; Cui, H.; Xin, A.; Zhao, R.; Qin, B. Purification, Characterization and Antioxidant Activities of a Polysaccharide from the Roots of Lilium davidii Var. Unicolor Cotton. Int. J. Biol. Macromol. 2019, 135, 1208–1216. [Google Scholar] [CrossRef]
- Nie, C.; Zhu, P.; Ma, S.; Wang, M.; Hu, Y. Purification, Characterization and Immunomodulatory Activity of Polysaccharides from Stem Lettuce. Carbohydr. Polym. 2018, 188, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Mannai, F.; Elhleli, H.; Yılmaz, M.; Khiari, R.; Belgacem, M.N.; Moussaoui, Y. Precipitation Solvents Effect on the Extraction of Mucilaginous Polysaccharides from Opuntia ficus-indica (Cactaceae): Structural, Functional and Rheological Properties. Ind. Crop. Prod. 2023, 202, 117072. [Google Scholar] [CrossRef]
- Yang, B.; Wu, Q.; Luo, Y.; Yang, Q.; Wei, X.; Kan, J. High-Pressure Ultrasonic-Assisted Extraction of Polysaccharides from Hovenia Dulcis: Extraction, Structure, Antioxidant Activity and Hypoglycemic. Int. J. Biol. Macromol. 2019, 137, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Li, C.; Zhang, C.; Zeng, R.; Fu, C. Optimization of Infrared-Assisted Extraction of Bletilla Striata Polysaccharides Based on Response Surface Methodology and Their Antioxidant Activities. Carbohydr. Polym. 2016, 148, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, X.; Wang, Z.; Li, Z.; Geng, Y. Effects of Pine Pollen Polysaccharides and Sulfated Polysaccharides on Ulcerative Colitis and Gut Flora in Mice. Polymers 2023, 15, 1414. [Google Scholar] [CrossRef]
- Deng, Y.; Huang, L.; Zhang, C.; Xie, P.; Cheng, J.; Wang, X.; Liu, L. Novel Polysaccharide from Chaenomeles speciosa Seeds: Structural Characterization, α-Amylase and α-Glucosidase Inhibitory Activity Evaluation. Int. J. Biol. Macromol. 2020, 153, 755–766. [Google Scholar] [CrossRef]
- Chen, G.; Sun, J.; Dai, Q.; Sun, M.; Hu, P. Polysaccharides from Seedless Chestnut Rose (Rosa sterilis) Fruits: Insights into Innovative Drying Technologies and Their Structural Characteristics, Antioxidant, Antiglycation, and α-Glucosidase Inhibitory Activities. Foods 2024, 13, 2483. [Google Scholar] [CrossRef]
- Fu, X.; Yang, H.; Ma, C.; Li, X.; Li, D.; Yang, Y.; Xu, Y.; Wang, L. Characterization and Inhibitory Activities on α-Amylase and α-Glucosidase of the Polysaccharide from Blue Honeysuckle Berries. Int. J. Biol. Macromol. 2020, 163, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Guan, Y.; Khoo, H.E.; Li, X. In Vitro Assessment of Chemical and Pre-Biotic Properties of Carboxymethylated Polysaccharides From Passiflora Edulis Peel, Xylan, and Citrus Pectin. Front. Nutr. 2021, 8, 778563. [Google Scholar] [CrossRef]
- He, C.; Zhang, R.; Jia, X.; Dong, L.; Ma, Q.; Zhao, D.; Sun, Z.; Zhang, M.; Huang, F. Variation in Characterization and Probiotic Activities of Polysaccharides from Litchi Pulp Fermented for Different Times. Front. Nutr. 2022, 9, 993828. [Google Scholar] [CrossRef]
- Chen, C.; You, L.-J.; Abbasi, A.M.; Fu, X.; Liu, R.H. Optimization for Ultrasound Extraction of Polysaccharides from Mulberry Fruits with Antioxidant and Hyperglycemic Activity in Vitro. Carbohydr. Polym. 2015, 130, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wang, L.; Li, L.; Zhang, Z.; Zhang, J.; Zhang, J.; Wang, J. Characterization of Polysaccharide Fractions from Allii Macrostemonis bulbus and Assessment of Their Antioxidant. LWT 2022, 165, 113687. [Google Scholar] [CrossRef]
- Li, J.; Shi, H.; Yu, J.; Lei, Y.; Huang, G.; Huang, H. Extraction and Properties of Ginkgo Biloba Leaf Polysaccharide and Its Phosphorylated Derivative. Ind. Crop. Prod. 2022, 189, 115822. [Google Scholar] [CrossRef]
- Long, X.; Hu, X.; Xiang, H.; Chen, S.; Li, L.; Qi, B.; Li, C.; Liu, S.; Yang, X. Structural Characterization and Hypolipidemic Activity of Gracilaria lemaneiformis Polysaccharide and Its Degradation Products. Food Chem. X 2022, 14, 100314. [Google Scholar] [CrossRef]
- Liu, C.; Hu, D.; Zhu, H.; Zhang, Y.; Qin, J.; Wang, F.; Zhang, Z.-D.; Lv, G. Preparation, Characterization and Immunoregulatory Activity of Derivatives of Polysaccharide from Atractylodes lancea (Thunb.) DC. Int. J. Biol. Macromol. 2022, 216, 225–234. [Google Scholar] [CrossRef]
- Ouyang, H.; Guo, B.; Hu, Y.; Li, L.; Jiang, Z.; Li, Q.; Ni, H.; Li, Z.; Zheng, M. Effect of Ultra-High Pressure Treatment on Structural and Functional Properties of Dietary Fiber from Pomelo Fruitlets. Food Biosci. 2023, 52, 102436. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Chen, G. Extraction, Structural Analysis, Derivatization and Antioxidant Activity of Polysaccharide from Chinese Yam. Food Chem. 2021, 361, 130089. [Google Scholar] [CrossRef]
- Wang, H.; Chen, J.; Ren, P.; Zhang, Y.; Onyango, S.O. Ultrasound Irradiation Alters the Spatial Structure and Improves the Antioxidant Activity of the Yellow Tea Polysaccharide. Ultrason. Sonochem. 2021, 70, 105355. [Google Scholar] [CrossRef]
- Niu, G.; You, G.; Zhou, X.; Fan, H.; Liu, X. Physicochemical Properties and in Vitro Hypoglycemic Activities of Hsian-Tsao Polysaccharide Fractions by Gradient Ethanol Precipitation Method. Int. J. Biol. Macromol. 2023, 231, 123274. [Google Scholar] [CrossRef]
- Nsor-Atindana, J.; Zhong, F.; Mothibe, K.J. In Vitro Hypoglycemic and Cholesterol Lowering Effects of Dietary Fiber Prepared from Cocoa (Theobroma cacao L.) Shells. Food Funct. 2012, 3, 1044–1050. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | SC-P | SC-PF |
|---|---|---|
| Yield (%) | 0.48 ± 0.06 b | 0.63 ± 0.06 a |
| Carbohydrate (%) | 44.40 ± 0.10 b | 50.35 ± 0.01 a |
| Protein (%) | 3.30 ± 0.00 a | 3.98 ± 0.20 a |
| Reducing sugar (%) | 3.65 ± 0.34 a | 5.10 ± 0.63 a |
| Uronic acid (%) | 0.23 ± 0.00 b | 0.29 ± 0.01 a |
| Molecular weight distribution (Da) | 35,964 | 32,022 |
| Monosaccharide composition (molar ratio, %) | ||
| Mannose (Man) | 2.96 | 5.55 |
| Ribose (Rib) | 7.80 | 10.07 |
| Glucuronic acid (GlcA) | 0.10 | 1.17 |
| Galacturonic acid (GalA) | 0.02 | 0.06 |
| Glucose (Glc) | 7.63 | 13.4 |
| Galactose (Gal) | 1.49 | 3.12 |
| Fructose (Fuc) | 1.26 | 1.72 |
| Xylose (Xyl) | - | 0.04 |
| Arabinose (Ara) | 0.19 | 0.37 |
| Particle size (nm) | 2005.09 ± 82.73 a | 1285.77 ± 168.85 b |
| Polydispersity index | 0.337 ± 0.013 a | 0.362 ± 0.023 a |
| Zeta potential (mv) | −16.49 ± 0.53 a | −21.42 ± 1.38 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, X.; Wang, X.; Cui, F.; Zeshan, A.; Wang, D.; Li, X.; Li, J. Effect of Enterococcus hirae GS22 Fermentation-Assisted Extraction on the Physicochemical and Bioactivities of Sea Cucumber Intestinal Polysaccharides. Molecules 2024, 29, 5800. https://doi.org/10.3390/molecules29235800
Tan X, Wang X, Cui F, Zeshan A, Wang D, Li X, Li J. Effect of Enterococcus hirae GS22 Fermentation-Assisted Extraction on the Physicochemical and Bioactivities of Sea Cucumber Intestinal Polysaccharides. Molecules. 2024; 29(23):5800. https://doi.org/10.3390/molecules29235800
Chicago/Turabian StyleTan, Xiqian, Xiaoqing Wang, Fangchao Cui, Ali Zeshan, Dangfeng Wang, Xuepeng Li, and Jianrong Li. 2024. "Effect of Enterococcus hirae GS22 Fermentation-Assisted Extraction on the Physicochemical and Bioactivities of Sea Cucumber Intestinal Polysaccharides" Molecules 29, no. 23: 5800. https://doi.org/10.3390/molecules29235800
APA StyleTan, X., Wang, X., Cui, F., Zeshan, A., Wang, D., Li, X., & Li, J. (2024). Effect of Enterococcus hirae GS22 Fermentation-Assisted Extraction on the Physicochemical and Bioactivities of Sea Cucumber Intestinal Polysaccharides. Molecules, 29(23), 5800. https://doi.org/10.3390/molecules29235800