Abstract
Due to the increase in multidrug-resistant microorganisms, the investigation of novel or more efficient antimicrobial compounds is essential. The World Health Organization issued a list of priority multidrug-resistant bacteria whose eradication will require new antibiotics. Among them, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacteriaceae are in the “critical” (most urgent) category. As a result, major investigations are ongoing worldwide to discover new antimicrobial compounds. Burkholderia, specifically Burkholderia sensu stricto, is recognized as an antimicrobial-producing group of species. Highly dissimilar compounds are among the molecules produced by this genus, such as those that are unique to a particular strain (like compound CF66I produced by Burkholderia cepacia CF-66) or antimicrobials found in a number of species, e.g., phenazines or ornibactins. The compounds produced by Burkholderia include N-containing heterocycles, volatile organic compounds, polyenes, polyynes, siderophores, macrolides, bacteriocins, quinolones, and other not classified antimicrobials. Some of them might be candidates not only for antimicrobials for both bacteria and fungi, but also as anticancer or antitumor agents. Therefore, in this review, the wide range of antimicrobial compounds produced by Burkholderia is explored, focusing especially on those compounds that were tested in vitro for antimicrobial activity. In addition, information was gathered regarding novel compounds discovered by genome-guided approaches.
1. Introduction
Burkholderia sensu lato comprises more than 100 species, which were gradually discovered during 30 years of research. In recent years, using comparative genomics, this large group was divided into seven genera, namely Burkholderia sensu stricto (s.s.), Paraburkholderia, Caballeronia, Robbsia, Mycetohabitans, Trinickia, and Pararobbsia [1,2,3,4,5]. The species contained in these genera thrive in soil, water, rhizosphere, plant nodules, fungi, and in animal and human infections. Burkholderia s.s. is formed by three groups of species: (a) the Burkholderia pseudomallei group (composed of 8 species), (b) the Burkholderia species that are mostly plant pathogenic bacteria (containing 4 species), and (c) the Burkholderia cepacia complex (Bcc) (composed of 25 species). The B. pseudomallei group is of worldwide importance because the species B. pseudomallei and Burkholderia mallei cause the mortal (if not treated) melioidosis diseases in humans and animals, and glanders, specifically in equines, respectively [6,7]. Recently, “Burkholderia mayonis” and “Burkholderia savannae” were described within the B. pseudomallei group [8]. Although some species from the Bcc are plant pathogens, there is a small group where Burkholderia plantarii, Burkholderia gladioli, and Burkholderia glumae are included; however, they are not part of the Bcc. Recently, “Burkholderia perseverans” was added to this group; this species produces volatile compounds that inhibit plant pathogens but has not been described as a pathogen per se [9]. The Bcc species are best known as opportunistic pathogens, mainly in cystic fibrosis (CF) and immunocompromised patients [10]. Within the Bcc, the last species described was Burkholderia orbicola [11]. Another important feature within the Bcc is their resistance to many antibiotics [12], which especially endangers the lives of CF and immunocompromised patients.
Bcc is also known for their phenotypic and genotypic diversity [13], which includes features/functions for biotechnological uses. This functional ability has been shown through biopesticidal activity in the rhizosphere [14] and by the bioremediation of xenobiotics compounds [15,16]. The Bcc are also able to produce a large array of compounds involved in the inhibition of pathogenic bacteria, fungi, and yeasts, which is important for tackling multidrug-resistant microorganisms [17]. Interestingly, the B. pseudomallei group encodes the largest capacity for secondary metabolite biosynthesis (>11% of their genomes) [18]. Moreover, the Bcc account for significant antibiotic biosynthetic capacity, e.g., Burkholderia ambifaria involves 9% of its genome in secondary metabolism, and B. gladioli and B. glumae dedicate 10% or more of their genome to antibiotic biosynthesis. Therefore, this review aims to enumerate in detail all antimicrobial compounds produced by Burkholderia s.s., detailing activities demonstrated in vitro and reviewing the novel compounds discovered by genome-guided approaches. The compounds are grouped and discussed according to common chemical features and shown as a list in Supplementary Table S1 with the numbers given in bold.
2. N-Containing Heterocycles
The analogs of nitrogen-based heterocycles occupy an exclusive position as a valuable source of therapeutic agents in medicinal chemistry [19]. Many of these compounds are volatile organic compounds (VOCs) or volatile nitrogen compounds [20]. Pyrazine-derived compounds (VOC) produced by Burkholderia seminalis JRBHU6 have been named PPDH and identified as (pyrrolo (1,2-a) pyrazine-1,4-dione, hexahydro) (1) and PPDHMP identified as C11H18N2O2 (pyrrolo (1,2-a) pyrazine-1,4-dione, hexahydro-3(2-methyl-propyl)) (2) [21]. The pyrrolo [1,2-a] pyrazine core occurs in nature and is frequently used in drug design. Pyrrole has therapeutic significance as an anticancer, antimicrobial, and antiviral agent [22]. Compounds 1 and 2 produced by B. seminalis JRBHU6 inhibit the fungi genera Fusarium, Aspergillus, Microsporum, Trichophyton, and Trichoderma, and the bacterial genera Staphyloccous, Pseudomonas, Escherichia, Shigella, and Klebsiella [21]. Molecular docking with bioactive compounds 1 and 2 was carried out to identify protein targets. According to the analysis, 2 showed full fitness to human proteins cell division protein kinase 7 and mitogen-activated protein kinase 8, suggesting a putative role in the inhibition of protein kinase activity in sensitive microorganisms. Good binding affinity and full fitness were also found with bacterial proteins such as choloylglycine hydrolase, camphor t-monooxygenase, chitinase B, and tyrosine phenol-lyase, while 1 showed good strong full fitness only with chitinase B. Other N-containing antimicrobial compounds are iminopyrrolidines produced by B. plantarii 9424. These compounds are 2-imino-3-methylene-5-L(carboxy-L-valyl)-pyrrolidine (3) and 2-imino-3-methylene-5-L(carboxy-L-threoninyl)-pyrrolidine (4). These are amino acid conjugates and have high in vitro inhibitory activity against the bacteria Erwinia amylovora, a pathogen causing fire blight disease in apple and pear trees [23]. A pyrazole molecule that consisted of a substituted pyrazole, linked to the aspartate-b-carboxyl of the tripeptide L-alanyl-L-homoserinyl-L-aspartate, resulted in a deduced structure 3-[L-alanyl-L-homoserinyl-L-aspartyl-b-carboxy]-4-hydroxy-5-oxopyrazole (5). This compound was produced by B. glumae and was found to inhibit bacterial pathogens such as different species of Erwinia, Pectobacterium, Pseudomonas, and Xanthomonas [24]. Pyrrolnitrin (6), 3-chloro-4-(2-nitro-3-chlorophenyl)pyrrole, is a microbial halometabolite (containing a halogen moiety) with a large antimicrobial significance in agricultural, pharmaceutical, and industrial implications [25]. This compound is produced by rhizospheric fluorescent and non-fluorescent pseudomonads, Serratia and Burkholderia. Pyrrolnitrin was first discovered in Pseudomonas (now Burkholderia) pyrrocinia in 1960 [26,27]; other species such as Burkholderia cepacia and Burkholderia ambifaria are able to synthesize it as well [28,29,30,31]. A number of phytopathogens are inhibited by 6, e.g., Penicillium, Phytophthora, Fusarium, Rhizoctonia, Colletotrichum, and Sclerotinia, yeast such as Candida, Hansenula, and Saccharomyces, and bacteria such as Bacillus and Streptomyces. Interestingly, the production of pyrrolnitrin was induced when chloramphenicol was added to the culture medium of B. ambifaria AMMDT [31]. Phenazines are a large group of nitrogen-containing heterocycles with diverse chemical structures and pharmacological activity such as antimicrobial, antiparasitic, neuroprotective, insecticidal, anti-inflammatory, and anticancer [32]. There are more than 100 phenazine derivatives produced by bacteria and archaea. Burkholderia cepacia 5.5B produces the phenazine 4,9-dihydroxyphenazine-1,6-dicarboxylic acid dimethyl ester (7), which inhibits Rhizoctonia solani [33]. The production of phenazines in B. lata was strongly affected by the growth conditions, the best production being observed in culture grown in King’s B medium [34]. Moreover, the involvement of phenazine in the formation of biofilm by B. lata was analyzed using a phenazine-overproducing strain, a phenazine-deficient mutant, and the wild type of the strain. The results showed that both the wild type and the overproducing strain formed thicker biofilms and attached more quickly than the mutant, suggesting a role of phenazine in biofilm formation by B. lata and, therefore, a role in the pathogenicity of this member of Bcc. Burkholderia glumae 411gr-6 was found to synthesize phencomycin (8) (a phenazine with two substituents, a carboxyl and a carbomethoxy group) and two new derivatives, 4-hydroxyphencomycin (9) and 5,10-dihydro-4,9-dihydroxyphencomycin methyl ester (10) [35]. The three compounds inhibit several plant pathogenic fungi, yeasts, and bacteria. Burkholderia sp. HQB-1, closely related to Burkholderia stagnalis, produces PCA, phenazine-1-carboxilic acid (11), which has been proposed to protect banana against Fusarium oxysporum wilt. Compound 11 produced by strain HQB-1 also inhibits the genera Colletotrichum, Botrytis, and Curvularia [36]. Indole compounds and derivatives are N-containing heterocycles; among this kind of compounds the VOC indole (12) produced by Burkholderia cenocepacia ETR-B22 inhibited the fungi Alternaria, Aspergillus, Bipolaris, Bacillus, Fusarium, Helminthosporium, Mycosphaerella, Magnaporthe, Phyllosticta, and Rhizoctonia [37]. The pityriacitrin (13), a b-carboline alkaloid with an indole ring attached with a carbonyl group on C-1 position and the derivative pityriacitrin B (14) isolated and identified in Burkholderia sp. NBF227, was tested for cytotoxic activity against cancer cell lines, but chemically synthesized derivatives from the previous compounds were more effective [38]. Other synthesized pityriacitrin derivatives from Burkholderia sp. NBF227 were investigated for antifungal activity [39]. The fungicidal activity was tested with four taxonomically different plant pathogens (oomycetes, ascomycetes, deuteromycetes, and basidiomycetes), showing that pityriacitrin displayed broad-spectrum antifungal activity and protected pepper leaves and grapefruits against infection by P. capsici and B. cinerea, respectively. Some N-containing heterocycles are shown in Figure 1.
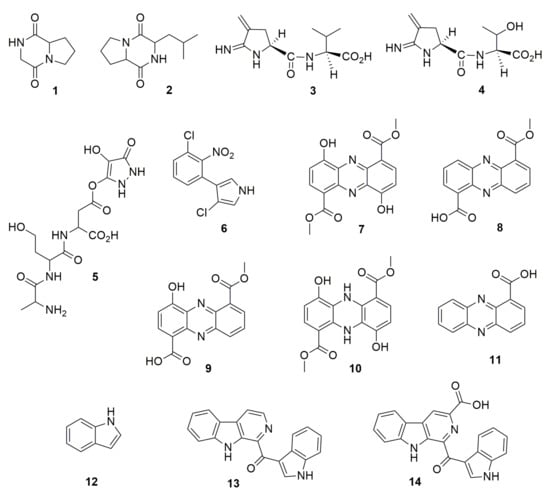
Figure 1.
N-containing heterocycles. (1) pyrrolo (1,2-a) pyrazine-1,4-dione, hexahydro. (2) pyrrolo (1,2-a) pyrazine-1,4-dione, hexahydro-3(2-methyl-propyl). (3) 2-imino-3-methylene-5-L(carboxy-L-valyl)-pyrrolidine. (4) 2-imino-3-methylene-5-L(carboxy-L-threoninyl)-pyrrolidine. (5) 3-[L-alanyl-L-homoserinyl-L-aspartyl-b-carboxy]-4-hydroxy-5-oxopyrazole. (6) pyrrolnitrin, 3-chloro-4-(2-nitro-3-chlorophenyl)pyrrole. (7) phenazine, 4,9-dihydroxyphenazine-1,6-dicarboxylic acid dimethyl ester. (8) phencomycin. (9) 4-hydroxyphencomycin. (10) 5,10-dihydro-4,9-dihydroxyphencomycin methyl ester. (11) phenazine-1-carboxilic acid. (12) índole. (13) pityriacitrin. (14) pityriacitrin B.
3. Volatile Organic Compounds
Besides the VOCs mentioned in the N-containing heterocycles section, B. cenocepacia ETR-B22 also synthetizes other VOCs that lack nitrogen in their ring structure (Figure 2). These compounds are the benzyl derivatives methyl anthranilate (15), methyl salicylate (16), methyl benzoate (17), benzyl propionate (18), benzyl acetate (19), 3,5-Di-tert-butylphenol (20), allyl benzyl ether (21), and benzyl benzoate (22) (Figure 2), which inhibit an important number of fungal plant pathogens [37]. The VOCs dimethyl trisulfide (23), nonanoic acid (24), 2-pentadecanone (25), and 3-hexen-1-ol, benzoate, (Z)- (26) produced by the strain ETR-B22 also have antifungal activity. Burkholderia gladioli strain BBB-01, isolated from rice shoots, emits the VOCs dimethyl disulfide (27) and 2,5-dimethylfuran (28) with inhibitory activity against the phytopathogenic fungi M. oryzae, Gibberella fujikuroi, Sarocladium oryzae, Phellinus noxius, and Colletotrichum fructicola and human pathogen C. albicans [40].
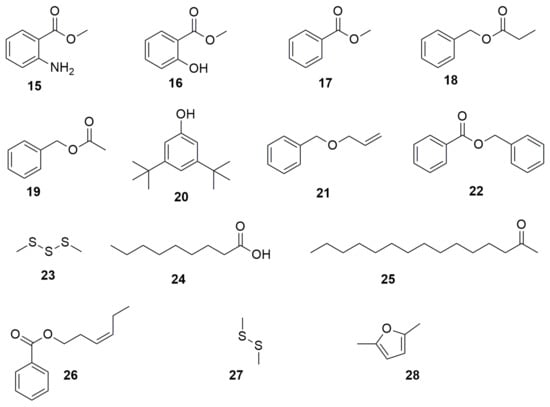
Figure 2.
Volatile organic compounds. (15) methyl anthranilate. (16) methyl salicylate. (17) methyl benzoate. (18) benzyl propionate. (19) benzyl acetate. (20) 3,5-Di-tert-butylphenol. (21) allyl benzyl ether. (22) benzyl benzoate. (23) dimethyl trisulfide. (24) nonaoic acid. (25) 2-pentadecanone. (26) 3-hexen-1-ol, benzoate, (Z)-. (27) dimethyl disulfoxide. (28) 2,5-dimethylfuran.
4. Polyenes
Polyenes are poly-unsaturated organic compounds that contain at least three alternating double and single carbon–carbon bonds. Hunter and Manter [41] reported the isolation and purification of a compound with oxidizing and antibiotic properties from B. cenocepacia P525. The structure of this compound has not been reported but the preliminary chemical study showed that the compound could be a polyene with six conjugated double bonds and bacteriostatic activity against Enterobacter soli and Enterobacter aerogenes. Burkholderia thailandensis is a close relative of B. pseudomallei and therefore used as a model to study B. pseudomallei pathogenicity and biosynthetic pathways because B. thailandensis is not a pathogen. This species produces the polyene polyketide thailandamide A (29) (Figure 3) inhibiting notably bacteria such as Bacillus subtilis, S. aureus, and Neisseria gonorrhoeae [42]. Genetic analysis showed that 29 inhibits acetyl-CoA carboxylase (ACC), an essential enzyme responsible for the first step in fatty acid biosynthesis. Moreover, B. thailandensis synthetizes thailandenes A (30), B (31) and C (32) (Figure 3), which are linear formylated or acidic polyenes containing a combination of cis and trans double bonds [43]. Compounds 30 and 31 exhibited potent antimicrobial activity against S. aureus and S. cerevisiae. A polyketide (PK) enacyloxin IIa (33) (Figure 3) and its stereoisomer, designated iso-enacyloxin IIa (34), produced by B. ambifaria AMMDT, has activity against Burkholderia multivorans, Burkholderia dolosa, and Acinetobacter baumannii [44]. Expression analysis showed that enzymes-encoding genes for enacyloxin biosynthesis were among the most highly upregulated when strain AMMDT was grown to stationary phase on glycerol. Moreover, enacyloxin targets protein biosynthesis by inhibition of the ribosomal elongation factor Tu [45]. Burkholderia gladioli pv. cocovenenans ATCC 33664T produces 33 and enacyloxin IIIa (35), and both were found to display equally potent activity against Escherichia coli and P. aeruginosa [46]. Moreover, traditionally used in food fermentations (tempe and sufu), Rhizopus microspores is accompanied by B. gladioli pv. cocovenenans. Thus, a coculture of both microorganisms showed that enacyloxins were found in high titers, with an increased production of the lethal toxin bongkrekic acid, showing the significance to food safety of this common microbial co-existence.
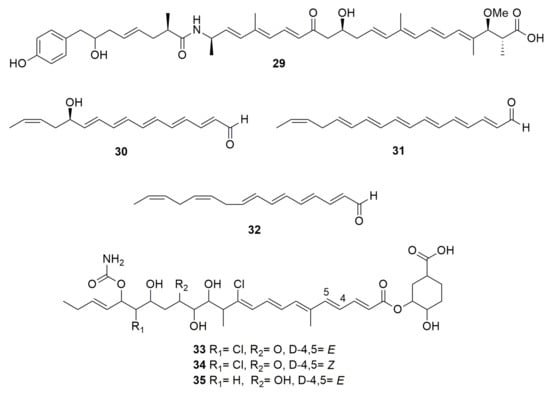
Figure 3.
Polyenes. (29) thailandamide A. (30) thailandene A. (31) thailandene B. (32) thailandene C. (33) enacyloxin IIa. (34) iso-enacyloxin IIa. (35) enacyloxin IIIa.
5. Polyynes
Polyynes are organic compounds with alternating single and triple bonds, a series of consecutive alkynes. Cepacin A (36) and cepacin B (37) (Figure 4) are two acetylenic antibiotics produced by B. cepacia SC 11,783 with a strong activity against staphylococci [47] (Parker et al. 1984). Burkholderia ambifaria BCC0191 also synthetizes the metabolite cepacin A, which mediates protection of germinating crops against Pythium damping-off disease [48]. The activity was demonstrated when no biological control was observed with the inoculation of a cepacin mutant of strain BCC0191. Burkholderia caryophylli, a plant pathogen, produces the triple-bond compounds caryoynecin A (38), B (39), and C (40) (Figure 4). Although they are unstable, they can inhibit E. coli, K. pneumoniae, and S. aureus [49]. Caryoynecin analogues synthesized chemically were found more stable and demonstrated activity against S. aureus, B. subtilis, Enterococcus faecalis, E. coli, Salmonella enteritidis, K. pneumoniae, Serratia marcescens, Proteus vulgaris, Shigella flexneri, Enterobacter cloacae, P. aeruginosa, T. mentagrophytes, Trichophyton interdigitale, and Trichophyton rubrum [50]. Burkholderia gladioli also produces caryoynencin, which has activity against Purpureocillium lilacinum and has a role in the transition of the plant pathogen to an insect-defensive mutualism [51].
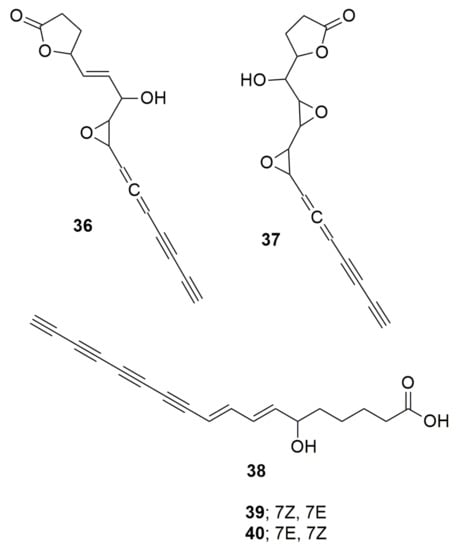
Figure 4.
Polyynes. (36) Cepacin. (37) Cepacin B. (38) Caryoynecin A. (39) Caryoynecin B. (40) Caryoynencin C.
6. Siderophores
Siderophores are low-molecular-weight organic compounds with high affinity to chelate iron (Fe). These compounds are produced by microorganisms and higher plants [52]. The typical siderophores ligands are cathecholate, a-hydrocycarboxylate, hydroxyphenyloxazolone, hydroxamate, a-aminocarboxylate, and a-hydroxyimidazole. Many bacterial siderophores are synthesized through non-ribosomal peptide synthetases (NRPS). NRPS are a large family of biosynthetic enzymes that generate relevant natural compounds from amino acid precursors [53,54]. NRPSs are frequently categorized as type I and II [55]. Type I NRPSs are large modular complexes containing all the enzymes necessary to generate a peptide product in an assembly line fashion analogous to type I fatty acid synthases (FASs) and polyketide synthases (PKSs). Type II NRPS proteins are commonly standalone enzymes or didomains that coordinate to form unique amino acid derivatives. Unlike type II FAS and PKS, the type II NRPS proteins are linear, noniterative pathways that contain specialized tailoring enzymes and combine with other pathways to generate a final product [55]. Pyochelin (41) (Figure 5), a non-ribosomal peptide (NRP) purified from P. aeruginosa PAO1, was first found to display antibiotic activity against S. aureus and moderately against several species of Xanthomonas [56]. Pyochelin produced by “Burkholderia paludis”, a non-validated species within the Bcc, inhibits three multidrug-resistant E. faecalis and four S. aureus strains but was not able to inhibit Bacillus subtilis ATCC 8188, Bacillus cereus ATCC 14579, Aeromonas hydrophila ATCC 49140, E. coli ATCC 25922, Klebsiella pneumoniae ATCC 10031, Proteus mirabilis ATCC 49140, P. vulgaris IMR, P. aeruginosa ATCC 10145 and ATCC BAA-47, Salmonella Typhimurium ATCC 14028, or Shigella flexneri ATCC 12022 [57]. This compound enhanced the production of intracellular reactive oxygen species (ROS), leading to cell death by disrupting the integrity of the bacterial membrane [58]. Pyochelin synthesized by B. seminalis TC3.4.2R3 inhibits F. oxysporum, which was demonstrated when a cepacin mutant was unable to inhibit the fungi [59]. Cepabactin (42) (Figure 5) is a 1-hydroxy-5-methoxy-6-methyl-2(1H)-pyridinone, a cyclic hydroxamate but also a heterocyclic analogue of catechol [60]. This compound produced by B. cepacia ATCC 25416T has antimicrobial activity against S. aureus, Staphylococcus epidermidis, Streptococcus faecalis, B. subtilis, Bacillus anthracis, E. coli, Salmonella Typhi, Salmonella Typhimurium, K. pneumoniae, P. vulgaris, P. mirabilis, and Proteus rettgeri [60,61,62]. The production of 42 was present in only 12% of 65 B. cepacia strains, lower than other siderophores such as ornibactin (87%) or pyochelin (60%), showing that this siderophore is not largely produced in the species [63]. Ornibactin (43) (Figure 5), a NRP produced by most Burkholderia species [64], is a tetrapeptide siderophore with an l-ornithine-d-hydroxyaspartate-l-serine-l-ornithine backbone. A study with Burkholderia contaminans MS14, isolated from soil in Mississippi, USA, using transposon mutagenesis, resulted in two strains with insertional mutations in orbI gene (mutant MT577) and a luxR family transcriptional regulatory gene (mutant MT357) [65]. Both mutants lost bactericidal activity, relating the activity to siderophore ornibactin. This compound successfully inhibited Xanthomonas citri pv. malvacearum, P. carotovorum supsp. carotovorum, Ralstonia solanacearum, P. syringae pv. syringae, E. amylovora, E. coli, Clavibacter michiganensis subsp. Michiganensis, and Bacillus megaterium. The ornibactin mutant retained antifungal activity, showing that the antibacterial and antifungal action is independent, with the antifungal activity a result of occidiofungin. Similarly, ornibactin derivatives produced by Burkholderia catarinensis 89T presented no activity against fungi [17]. Pyochelin and ornibactin are siderophores found in the genome of Burkholderia orbicola TAtl-371T, and this bacterium produces siderophores in culture medium [66]. A test removing iron from the culture medium showed that the bacteria was able to inhibit Paraburkholderia phenazinium and Candida glabrata, suggesting the involvement of these siderophores in antagonism.
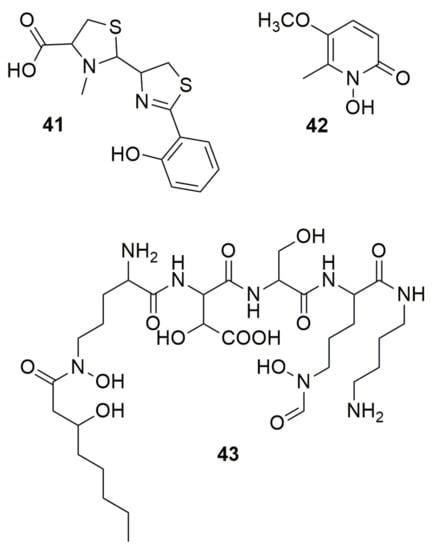
Figure 5.
Siderophores. (41) Pyochelin. (42) Cepabactin. (43) Ornibactin.
7. Macrolides
Macrolides are various types of hydrophobic compounds containing a macrocyclic lactone ring and various side chains/groups [67]. Burkholderia gladioli BCC0238 isolated from a CF patient synthesize the PK, macrolide antibiotic gladiolin (44) (Figure 6), which has a strong activity against M. tuberculosis H37Rv and several other M. tuberculosis strains, K. pneumoniae, A. baumannii, P. aeruginosa, E. clocae, Serratia plymuthica, “Ralstonia mannitolilytica”, B. multivorans, E. coli, Enterococcus faecium, S. aureus, B. subtilis, and C. albicans, and was found to exhibit low toxicity toward an ovarian cancer cell line [68]. The mode of action of gladiolin is the inhibition of the RNA polymerase. Another PK macrolide lagriene (45) (Figure 6), produced by B. gladioli Lv-StA, has activity against B. thuringiensis, M. vaccae, vancomycin-resistant E. faecalis and S. aureus [51].
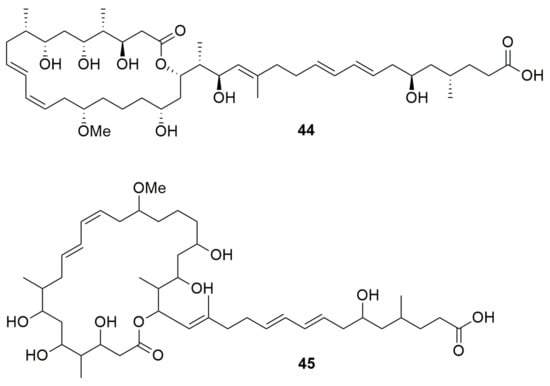
Figure 6.
Macrolides. (44) Gladiolin. (45) Lagriene.
8. Bacteriocins
Bacteriocins are a varied class of bactericidal peptides or proteins produced by bacteria and archaea with bactericidal activity and specific immunity mechanisms toward strains closely related to the producer bacteria [69]. There are two central differences between bacteriocins and antibiotics: bacteriocins are ribosomally synthesized but antibiotics are not, and bacteriocins have a somewhat narrow killing spectrum while antibiotics have an extensive killing range. Bacteriocins vary in size, microbial target, mode of action, release, and immunity mechanism, and can be divided into two groups, the ones produced by Gram-negative bacteria and those by Gram-positive bacteria. Gram-negative bacteriocins are further classified according to their size into three main groups, namely colicins, phage-tail-like bacteriocins, and microcins [70]. Microcins are low-molecular-weight compounds grouped into class I (<5 kDa) or class II (5–10 kDa). Class I is now designated as ribosomally synthesized and post-translationally modified peptides (RiPP). Burkholderia cenocepacia BC0425 synthesizes the bacteriocin tailocin, a phage tail-like compound, named BceTMilo [71]. Unlike phages, tailocin injects through the cell membrane and disrupts the proton motive force [72]. Strains belonging to Bcc are sensitive to BceTMilo, and other non-Bcc such as B. gladioli and B. glumae are also sensitive to tailocin. Lectin-like bacteriocins (LlpAs) contain two monocot mannose-binding lectin (MMBL) domains, a module predominantly and abundantly found in lectins from monocot plants. Burkholderia strains can synthesize these bacteriocins. B. cenocepacia AU1054 (now B. orbicola) [11] produces an LlpA bacteriocin that inhibits B. ambifaria, Burkholderia anthina, B. cenocepacia, B. contaminans and Burkholderia metallica [73]. The homologue LlpA88 from B. orbicola TAtl-371T inhibited the same species as strain AU1054 [66]. Burkhocins M1 and M2, colicin M-like bacteriocins called ColM in E. coli, from B. ambifaria MEX-5 and AMMDT were produced recombinantly, showing antagonistic activity against a number of Bcc strains [74]. Three strains from Burkholderia ubonensis inhibited B. pseudomallei; the antagonism from a representative strain (A21) was characterized, and a pepsin-sensitive moiety consistent with a bacteriocin-like compound was found, suggesting the antagonism is due to the production of a bacteriocin or bacteriocin-like inhibitory substance (BLIS) [75]. Lasso peptides are a structurally unique class of bioactive peptides characterized by a knotted arrangement where the C-terminus threads through an N-terminal macrolactam ring [76]. Lasso peptides are divided depending on the presence (class I) or absence (class II) of four conserved cysteine residues involved in the formation of two intramolecular disulfide bonds [77]. Burkholderia thailandensis produces the lasso peptide capistruin, a 19-amino-acid class II lasso peptide comprising an isopeptide bond between Gly1 and Asp9 resulting in a nine-residue macrolactam ring [78], which exhibits antimicrobial activity against Burkholderia (now Paraburkholderia) caledonica, E. coli, and P. aeruginosa [76]. Ubonodin, another lasso peptide produced by B. ubonensis MSMB2207, was heterologous expressed in E. coli BL21, displaying inhibition of B. cepacia, B. multivorans and B. mallei; it has a weak effect against B. thailandensis and had no effect on B. gladioli and B. pseudomallei [79]. This compound inhibits RNA polymerase in vitro and the narrow effect might allow therapeutic usage.
9. Quinolones
Quinolones were discovered as a by-product in the search for improved synthesis of the anti-malarial chloroquine; thus, they are fully synthetic molecules [80]. Today, it is known that molecules in the quinolone family are also present as natural products of plants and bacteria, although their potency has been tested only at the experimental level. The basic structure is a 3-carboxyquinolone and the first quinolone described was nalidixic acid. Burkholderia thailandensis contains a biosynthetic gene cluster (BGCs), which is a quorum-sensing-regulated hmq cluster that produces a diverse set of hydroxyalkylquinolines (HAQs). These compounds exist mainly in the 4(1H) quinolone type at neutral pH and are known as bioactive metabolites [81]. Two HAQ analogues, HMNQ (4-hydroxy-3-methyl-2-(2-nonenyl)-quinoline) (46) and HQNO (2-heptyl-4(1H)-quinolone N-oxide) (47) (Figure 7), synthesized by B. thailandensis E264T, when challenged with antibiotics inhibit B. subtilis 168 but display weak activity against E. coli K12 [82]. It was found, as well, that both quinolones act synergistically to inhibit bacterial growth. Moreover, B. thailandensis produces 46 and rhamnolipids in outer membrane vesicles (OMV), which have antimicrobial and antibiofilm properties against methyl-resistant S. aureus [83]. Bacterial OMVs contain proteins, lipids, polysaccharides, and small molecules and serve numerous and versatile roles in intra- and interspecies interactions. Burkholderia cepacia RB425, isolated from lettuce root, makes the quinolone antibiotics 2-(2-heptenyl)-3-methyl-4-quinolinol (48) and (46) (Figure 7) with high activity against fungal pathogen Verticillium dahlia, moderate inhibition of Pyricularia oryzae and Cochliobolus myyabeanus, and weak growth inhibition of R. solani, F. oxysporum, and Gaeumannomyces graminis [84]. A range of hydroxy-methyl-alkylquinolines (HMAQ) produced by B. cepacia PC-II antagonizes P. capsici, which is responsible for Phytophthora blight in red peppers and many vegetables; in particular, 48 was the most potent against the oomycetes P. capsica and Pythium ultimum and the fungi F. oxysporum and R. solanc [85]. Burkholderia sp. QN15488 produces burkholone (49) (Figure 7), a (E)-3-methyl-2-(octenyl)-4-quinolone, this compound induces cell death in 32D/GR15 cells in IGF-I-containing medium [86]. Insulin-like growth factors (IGFs) play a key role in human cancer progression and IGF signals through the IGF-1 receptor are known to be significant for tumor cell growth and survival [87].
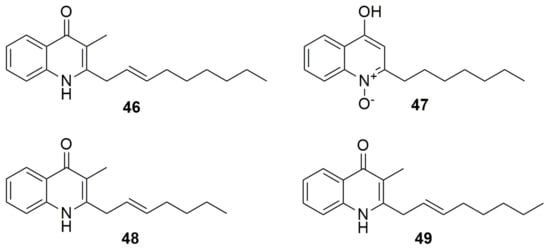
Figure 7.
Quinolones. (46) HMNQ (4-hydroxy-3-methyl-2-(2-nonenyl)-quinoline). (47) HQNO (2-heptyl-4(1H)-quinolone N-oxide). (48) 2-(2-heptenyl)-3-methyl-4-quinolinol (C7Δ2). (49) Burkholone.
10. Other NPR-PK Compounds
Gladiofungin A (50) (Figure 8) is a novel antifungal PK that is highly unusual because it harbors a butanolide moiety. This compound is produced by the insect-associated bacteria B. gladioli HKI0739, which displays activity against Penicillium notatum, Sprobolomyces salmonicolor, and P. lilacinum [88]. The strains BCC0238 and BCC1622, belonging to B. gladioli, produce a PK antibiotic named gladiostatin, which has the same structure as (50) [89]. This molecule has promising activity against several cancer cell lines such as ovarian, pancreatic, and colon cancer, and inhibits tumor cell migration. Moreover, it was found to be inactive against a lung cell line, which indicates that it may exhibit some selectivity. Gladiostatin contains an unusual 2-acyl-4-hydroxy-3-methylbutenolide in addition to the glutarimide pharmacophore that also inhibits S. cerevisiae. Glidobactins A (51), B (52), and C (53) have a common cyclized tripeptide nucleus composed of L-threonine, 4(S)-amino-2(E)-pentoic acid, and erythro-4hydroxy-L-lysine but differ from each other in the unsaturated fatty acid moiety attached to the peptide [90]. These compounds were isolated from strain K481-B101, whose 16S rRNA sequence (Accession No. AM410613) analyzed in the EzBioCloud server (https://www.ezbiocloud.net/, accessed on 1 June 2022) was identified as Schlegelella brevitalea, a member of the Burkholderiales order and Comamonadaceae family. These glidobactins compounds have antifungal and antitumor activity [91]. Later, cepafunings I, II (54), and III (55) acylpeptides produced by B. cepacia CB-3 were described by Shoji et al. [92]. The mixture of cepafungins has moderate inhibitory activity against pathogenic yeast and fungi such as C. albicans, Candida krusei, Aspergillus fumigatus, Microsporum canis, and T. mentagrophytes. It showed no curative effect in mice infected with C. albicans, and there was no activity against bacteria, but it had a moderate effect on prolonging the survival period of mice in which murine lymphatic leukemia P388 cells were implanted [92]. The elucidation of cepafungin structures showed that compound I is identical to 51 (Figure 8) [93]. Later, Schellenberg et al. [94] named the group of cepafungins as glidobactins and, while studying the genes for the synthesis of 51 in S. brevitalea K481-B101, found homologous gene clusters in B. pseudomallei and B. mallei. The production but not the antimicrobial activity of 53 synthesized by B. pseudomallei was reported by Biggins et al. [95]. Moreover, a 53 variant was described as deoxyglidobactin C, which contains a lysine instead of a 4-hydroxylisine within the structure. Occidiofungin A-D (56–59) (Figure 8), synthesized by B. contaminans MS14, are glycopeptides with antifungal activity inhibiting a large spectrum of fungal pathogens, among them Alternaria, Aspergillus, Fusarium, Geotrichum, Macrophomina, Microsporum, Penicillum, Pythium, Rhizoctonia, Trichophyton, and several Candida species [96,97,98]. Occidiofungin disrupts fungal membrane morphology and induces apoptosis [96,99]. A recent study identified actin filaments as the primary cellular target of occidiofungin in fungi [98]. It also has antiparasitic activity, damaging the parasite Cryptosporidium parvum [100]. Occidiofungin is also produced by B. pyrrocinia Lyc2, having antifungal activity, attacking Aspergillus, Cladosporium, Cochilobolus heterostrophus, Colletotrichum acutatum, Gaeumannomyces graminis, Geotrichum candidum, Glomerella cingulate and Thieloviopsis basicola [101]. Moreover, occidofungin was tested in toxicological evaluations and it was found to have minimal toxicity in human fibroblasts and has potent anticancer activity [102]. Cepacidine A1 (60) and A2 (61) (Figure 8) are glycopeptides produced by B. cepacia AF 2001. They are highly similar, with molecular weights of 1199 and 1219 Da, respectively [103]. Cepacidine A2 contains asparagine, and A1 includes b-hydroxy aspargine, which combined have potent antifungal but no antibacterial activity [104]. The cepacidine mixture inhibits C. albicans, C. glabrata, Cryptococcus neoformans, S. cerevisiae, A. niger, Microsporum gypseum, Epidermophyton floccosum, T. mentagrophyte, Trichophyton rubrum, F. oxysporum, and Rhizopus stolonifera. Moreover, cepacidine A has immunosuppressive action involving in vitro inhibition of the proliferation of murine lymphocytes [105]. Cepacidine A has moderate anthelmintic in vitro but not in vivo activity [106]. AFC-BC11 is a lipopeptide produced by B. cepacia BC11 [107]. This compound is involved in the biological control of R. solani damping-off in cotton. Icosalide A1 (62) (Figure 8) is an unusual two-tailed lipocyclopeptide antibiotic produced by B. gladioli HKI0739, which is active against entomopathogenic bacteria B. thuringiensis and Paenibacillus larvae and is involved in swarming inhibition [108]. Burkholderia gladioli BCC0238 was also found to synthesize (62), but its antimicrobial activity was not tested [109]. This compound was first reported from Aureobasidium and showed activity against Streptococcus pyogenes and E. faecalis [110]. Burkholderia thailandensis produces bactobolins A-D (63–66) (Figure 8), a group of polyketide-peptide molecules, some of which are potent antimicrobials [111]. The production of these compounds is temperature dependent with better results of production at 30 than 37 °C. The purification of the three most abundant bactobolins showed that 63 and 65 have strong activity against bacteria (Bacillus cereus, B. subtilis, B. cenocepacia, Paraburkholderia kururiensis, Burkholderia vietnamiensis, Chromobacterium violaceum, E. coli, Flavobacterium johnsoniae, K. pneumoniae, Mycobacterium marinum, P. aeruginosa, Pseudomonas fluorescens, Ralstonia pickettii, S. Typhimurium, S. aureus, and Streptococcus pyogenes) and fibroblasts. Xylocandins A1, A2, B1, B2, C1, C2, D1, and D2 were isolated from B. cepacia ATCC 3927 [112]. Xylocandins are cyclic peptides containing glycine, serine, asparagine, β-hydroxytyrosine, and an unusual amino acid with the formula C18H37NO5. The mixture of each compound showed that xylocandin A1 and A2 have a potent antifungal activity inhibiting several Candida species and dermatophytes such as T. mentagrophytes, T. rubrum, Epidermophyton floccosum, and M. canis, but does not inhibit Gram-negative and -positive bacteria, nor vaginitis in a rat model [113]. Burkholderia cenocepacia H111 was detected to produce a diazeniumdiolate metalophore compound called fragin (67) (Figure 8) [114]. When iron was added to the medium, the antifungal activity of strain H111 diminishes, suggesting that the metal chelation is the molecular basis for antifungal activity. Fragin enantiomers inhibit F. solani, B. cereus, B. subtilis, B. thuringiensis, S. aureus, and S. cerevisiae, but no Gram-negative bacteria such as C. violaceum, E. coli, Klebsiella oxytoca, and P. syringae. Besides fragin, strain H111 synthesizes a signal molecule called valdiazen, which shares a high degree of structural homology with fragin, but their function is different since valdiazen has no antimicrobial activity. Valdiazen is a diffusible signal that regulates both itself and fragin and the expression of more than 100 genes, representing a novel quorum-sensing signal. Betulinans are produced by B. pseudomallei K96243 [115]. During the screening of agonists for eukaryotic phosphodiesterase (PDE), the betulinan BTH-II0204-207:A (68) (Figure 8) compound produced by B. pseudomallei K96243 was found. PDE are divided into 11 families. PDE4 has been implicated in inflammation responses across multiple immune cell types; therefore, PDE4 inhibitors have been extensively investigated as potential therapeutic molecules for a number of inflammatory diseases. Bioactivity assays indicated that 68 is a PDE4 inhibitor. Microbial symbionts are often a source of chemicals that can contribute to host defense against antagonists. Lagria beetles live in symbiosis with multiple strains of Burkholderia that protect their offspring against pathogens. Among them, B. gladioli Lv-StB was found to produce the PK lagriamide (69) (Figure 8), which inhibits A. niger and P. lilacinum [116]. Isosulfazecin (70) (Figure 8) is a NRP b-lactam antibiotic produced by Pseudomonas mesoacidophila SB-72310 (now B. ubonensis) [117,118]. Compound 70 inhibits S. Typhimurium and moderately inhibits E. coli, P. vulgaris, P. mirabilis, S. marcescens, E. faecalis, and B. subtilis. Burkholderia ubonensis SB-72310 also produces bulgecins, glycopeptides that induce bulge formation in cooperation with b-lactams and enhance the lytic activity of b-lactam-antibiotics, but bulgecins show no antimicrobial activity [119]. Burkholderia cepacia CF-66 displays strong antifungal activity against R. solani; a compound named CF66I was purified and showed inhibition of F. oxysporum, Fusarium sambucinum, Rosselinia necatrix, Aspergillus flavus, A. niger, Cochilobus carbonum, B. cinerea, Mucor hiemolis, Penicillum chrysogenum, Rhizopus oryzae, C. albicans, C. neoformens, Pichia membranae, and S. cerevisiae but not E. coli, B. subtilis, or S. aureus [120]. Pseudomonas aeruginosa, causing nosocomial and wound infections, possess a signal molecule that integrates quorum sensing (QS) and stress response [121]. This integrated QS molecule (IQS) is identical to aeruginaldehyde from P. fluorescens [122]. IQS is effectively captured by the NRP siderophore malleobactin produced by B. thailandensis, which results in the formation of a rare nitrone bioconjugate called malleonitrone (71) (Figure 8) that is active against the IQS producer and therefore has significance from a pharmaceutical perspective [123]. Spliceostatins are spliceosome inhibitors, synthesized by a hybrid NRPS-PKS system of the trans-acyl transferase (AT) type, that show promising anticancer activity. Burkholderia sp. FERM BP-3421, identified by 16S sequence (KJ364655) as a member of the Bcc, produces the hemiketal spliceostatins, as well as analogs containing a terminal carboxylic acid [124]. Some spliceostatin analogues (72–73) and their semisynthetic analogues were evaluated in cell proliferation assays against a panel of solid tumor cell lines, showing potent cytotoxicity [125]. Diketopiperazines (DKP) are NRP-cyclized molecules comprising amino acid bounded by two peptide bonds [126]. Burkholderia cepacia CF-66 synthesizes the molecules diketopiperazines cyclo(Pro-Phe), cyclo(Pro-Tyr), cyclo(Ala-Val), cyclo(Pro-Leu), and cyclo(Pro-Val); all of these compounds are both D- and L-type [127]. These DKP showed a negative effect on the candidacidal activity of the culture supernatant extracts. Burkholderia cepacia CF-66 lacks the gen cepI that encodes for an acyl homoserine lactone (AHL), which is involved in QS; however, a study with B. cenocepacia J2315T showed that new DKP molecules can inhibit CepI in vitro, impairing the ability of B. cenocepacia to produce proteases and siderophores and to form biofilms [128].
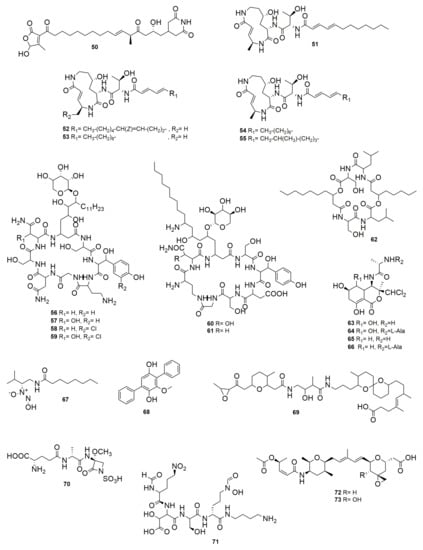
Figure 8.
Other NPR-PK compounds. (50) Gladiofungin A. (51) Glidobactin A. (52) Glidobactin B. (53) Glidobactn C. (54) Cepafungin I, II (55) Cepafungin III. (56–59) Occidiofungins A–D. (60) Cepacidine A1. (61) Cepacidine A2. (62) Icosalide A1. (63–66) Bactobolins A–D. (67) Fragin. (68) BTH-II0204-207:A. (69) Lagriamide. (70) Isosulfazecin (iSZ). (71) Malleonitrone. (72–73) Spliceostatins.
11. Other Antimicrobial Compounds
Sinapigladioside (74) (Figure 9), an aromatic glycoside, contains an isothiocyanate moiety, a rare structural feature among bacterial metabolites [51]. This compound produced by B. gladioli displays antifungal activity against P. lilacinum, which is an egg entomopathogen, A. fumigatus, and Penicillum notatum. A not fully characterized molecule referred to as “Compound 1”, an antimicrobial compound with an unknown structure but with an ion at m/z 391.2845 and produced by B. orbicola TAtl-371T, was found to inhibit only Tatumella terrea SHS 2008T [129]. Cepaciamide A (75) (Figure 9) is a (3R, 3′R, 2”R, 5”S, 6”R)-3-N-[3′-(2”-hydroxy-5”, 6”-methylenoctadecanoyl)-hexadecamido]-2-piperidinone isolated from B. cepacia D-202 [130]. This compound has toxicity activity against B. cinerea, which causes beet root rot in Japan. The bacterial type III secretion system (T3SS) acts as a complex multiunit nanomachine to translocate effector proteins across the bacterial membrane to deliver them directly into eukaryotic host cells [131]. B. gladioli NGJ1 produces a prophage tail-like protein (Bg_9562), which is a potential effector secreted by a T3SS and is essential for mycophagy in R. solani [132]. Bg_9562 protein showed antifungal activity against S. cerevisiae, C. albicans, Alternaria brassicae, M. oryzae, Venturia inaequalis, F. oxysporum 7063, Alternaria sp., Dedymella sp., Phytophthora sp., Colletotrichum sp., Ascochyta rabiei, and Neofusicoccum sp. Moreover, Bg_9562 protein showed no inhibition of E. coli, Pantoea ananatis, or B. glaidoli NGJ1. The compound 2-hydroxymethyl-chroman-4-one (76), designated as MSSP2 (Figure 9), is produced by Burkholderia sp. MSSP [133], whose 16S gene sequence (AY551271) indicates that it belongs to the Bcc. The compound MSSP2 displays an inhibitory effect against P. ultimum, P. capsica, and S. sclerotiorum. Altericidins A, B, and C produced by B. cepacia KB-1 can inhibit a wide range of fungi and yeasts, such as Alteraria kikuchiana and Ustilago maydis, but has no effect on bacteria [134]. Rhamnolipids (Rha) are glycolipidic biosurfactants consisting of rhamnose molecules linked through a β-glycosidic bond to 3-hydroxyfatty acids with various chain lengths produced by bacterial species with several functions such as antimicrobial activity [135]. The non-pathogenic B. thailandensis E264T synthesize di-rhamnolipids C14-C14 (77) and C12-C14 (78) (Figure 9), which have antibacterial and antibiofilm activity against Streptococcus sanguinis, Streptococcus oralis, Neisseria mucosa, and Actinomyces naeslundii [136]. Other non-pathogenic Burkholderia synthesize Rha, such as B. glumae and B. plantarii; however, their antagonistic activity was not tested [137,138]. Additionally, B. pseudomallei produce Rha-Rha C14-C14, which showed cytotoxic and hemolytic activities [139,140].
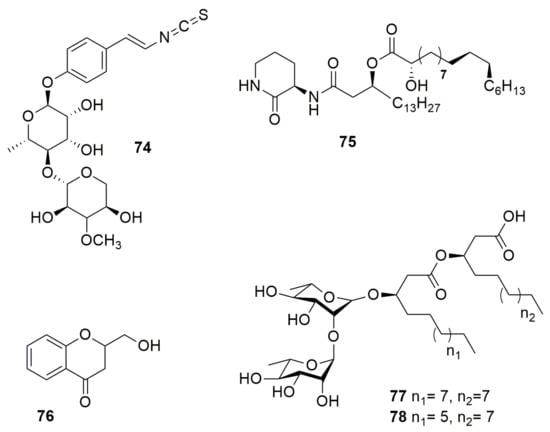
Figure 9.
Other antimicrobial compounds. (74) Sinapigladioside. (75) Cepaciamide A. (76) 2-hydroxymethyl-chroman-4-one. (77) Di-rhamnolipid C14-C14. (78) Di-rhamnolipids C12-C14.
12. Compounds with Dual Effect
Many compounds are beneficial for humans since they are antifungal, antibacterial, or anticancer molecules. However, some of these compounds have a dual effect, both beneficial and toxic. For instance, burkholdines (Figure 10) are NRP-cyclic lipopeptides produced by B. ambifaria 2.2N with potent antifungal activity [141]. Many burkholdines have been described and the analysis of five representatives (79–83) showed antifungal activity on S. cerevisiae, C. albicans, and A. niger. However, they also exhibit hemolytic activity [142]. The latter results indicate that these compounds are important for Burkholderia virulence. Tropolone (84) (Figure 10) is a troponoid containing a seven-membered aromatic ring with various substitutions, produced by B. plantarii [143]. Tropolone shows broad-spectrum antimicrobial activity against bacteria and fungi, but it is the phytotoxin responsible for rice seedling blight [144,145]. Cepalycin I and cepalycin II were isolated from B. cepacia JN106 [146], but their structure was not reported. These compounds have both hemolytic and antifungal activity, inhibiting S. cerevisiae, C. neoformans, and C. albicans.
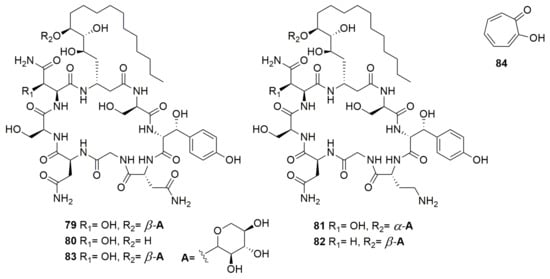
Figure 10.
Compounds with dual effect. (79–83) Burkholdines. (84) Tropolone.
13. Metabolism as Control
Fusaric acid (85) (Figure 11) is a fungal metabolite produced by several Fusarium species, which is responsible for wilts and root rot diseases in a number of plants. Burkholderia ambifaria T16 can grow with 85 as a sole carbon, nitrogen, and energy source, and showed the ability to detoxify 85 in barley seedlings, suggesting that the strain might serve as a new source of metabolites or genes for the development of novel 85-detoxification systems [147].
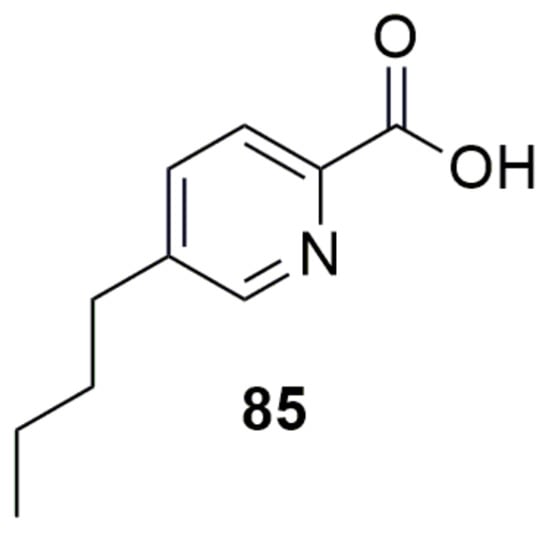
Figure 11.
Metabolism as control. (85) Fusaric acid.
14. Data Mining
Genome mining is a promising tool in the search for new bioactive compounds produced by microorganisms. This strategy has been used in several bacterial genomes to analyze their potential as sources of new compounds with pharmacological potential. Phenazines are structurally diverse, but all share a conserved seven-gene operon, phzABCDEFG, termed the “core phenazine biosynthesis genes” [148]. A genome screening of Burkholderia genomes showed that phenazine gene clusters were identified in 20 strains belonging to B. cepacia, Burkholderia lata, B. glumae, B. singularis, B. ubonensis, and some Burkholderia sp. strains [34]. A genome mining of 64 B. ambifaria strains revealed an armory of known and unknown pathways within this species, among them the biosynthetic gene cluster to produce cepacin, which was the mode of action for the biopesticidal activity of B. ambifaria [48]. In this study [48], other compounds were found in B. ambifaria genomes, such as pyrrolnitrin, burkholdines, hydroxyquinolines, bactobolins, and enacyloxina IIa. Moreover, Mullins and Mahenthiralingam [149] analyzed 4000 genomes representing the genera of Burkholderia s.l.; among them, the Burkholderia species harbored more biosynthetic gene clusters and the more diverse clusters per species compared to the remaining genera from Burkholderia s.l. These clusters include genes involved in the production of bacteriocins, phosphonates, lassopeptides, NRPS, betalactones, transAT-PKS, and terpenes. Chitinases are glycosyl hydrolases that catalyze the hydrolytic degradation of chitin, one of the major constituents of cell walls of fungi. The genome analysis of B. orbicola TAtl-371T showed the presence of the gene BCAL1722 that encodes a chitinase belonging to family 18 of the glycosyl hydrolases, as well as a gene that encodes a predicted chitinase [129]. The authors also found that several B. cenocepacia strains contain homologues; however, the activity of chitinase was not present in strain TAtl-371T. Moreover, Rojas-Rojas et al. [129] also found that the genome of strain TAtl-371T contains 30 genes reported for the biosynthesis of the bacteriocin BceTMilo. A genomic search for LlpAs in Burkholderia genomes and phylogenetic analysis showed two distinct clusters; one of them belongs to the B. pseudomallei group, including Burkholderia oklahomensis, B. pseudomallei, B. mallei, and B. thailandensis [73]. Another bacteriocin studied was ColM in Burkholderia. The colM-like bacteriocin gene was found mainly in Bcc and B. oklahomensis [74]. Ubonodin was found in the genome of 16 out of 306 B. ubonensis strains, which might be in relation to the intriguingly large size of 28 aa of the core peptides, longer than any previously characterized example [79]. HMAQ produced by the biosynthetic operon named hmqABCDEFG was searched for in the genome of Bcc strains [150]. The analysis showed that one-third of Bcc species carry a homolog of the hmqABCDEFG, and not all sequenced strains in each species possess this operon.
15. Conclusions
The ability of Burkholderia to produce antimicrobial compounds is remarkable, not just for the variety of molecules synthesized but also for the diversity of targets they attack, namely bacteria, fungi, cancer cells, tumor cells, or inflammatory processes. The compounds produced belong to a variety of chemical natures, such as N-containing heterocycles, volatile organic compounds, polyenes, polyynes, siderophores, macrolides, bacteriocins, quinolones, non-ribosomal peptides, polyketides, and other unclassified compounds such as sinapigladiosides, cepaciamide A, altericidins, and rhamnolipids, among others. Moreover, there are compounds that have both beneficial and toxic effects, such as burkholdines, tropolone, and others. The mining of genomes is another important method of finding new molecules. Certainly, Burkholderia is still a group of bacteria with as-yet unexplored compounds waiting to be discovered. Several papers about new Burkholderia strains are published daily, which may contain information about the new antimicrobial compounds they produce that have the potential to be used against multidrug-resistant microorganisms.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/molecules28041646/s1: Table S1: Antimicrobial compounds produced by Burkholderia sensu stricto. References [151,152] are cited in the Supplementary Materials.
Author Contributions
Conceptualization, methodology, validation, formal analysis, investigation, data curation, writing—original draft preparation, L.M.M.-R., M.R.-C., A.S.-G., F.U.R.-R. and P.E.-d.l.S.; writing—review and editing, visualization, supervision, project administration, funding acquisition, P.E.-d.l.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Secretaría de Investigación y Posgrado, Instituto Politécnico Nacional, grant numbers 2021-0392, 2022-0660, and 2023-0831.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
P.E.S. thanks COFAA, EDI, and SNI. We thank Martha Thayer for proofreading this manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Sawana, A.; Adeolu, M.; Gupta, R.S. Molecular signatures and phylogenomic analysis of the genus Burkholderia: Proposal for division of this genus into the emended genus Burkholderia containing pathogenic organisms and a new genus Paraburkholderia gen. nov. harboring environmental species. Front. Genet. 2014, 5, 429. [Google Scholar] [CrossRef] [PubMed]
- Dobritsa, A.P.; Samadpour, M. Transfer of eleven Burkholderia species to the genus Paraburkholderia and proposal of Caballeronia gen. nov., a new genus to accommodate twelve species of Burkholderia and Paraburkholderia. Int. J. Syst. Evol. Microbiol. 2016, 66, 2836–2846. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Santos, L.; Castro, D.B.A.; Ferreira-Tonin, M.; Correa, D.B.A.; Weir, B.S.; Park, D.; Mariscal-Ottoboni, L.M.; Rodrigues-Neto, J.; Lanza-Destefano, S.A. Reassessment of the taxonomic position of Burkholderia andropogonis and description of Robbsia andropogonis gen. nov., comb. nov. Anton. Leeuw. Int. J. Gen. 2017, 110, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Estrada-de los Santos, P.; Palmer, M.; Chavez-Ramírez, B.; Beukes, C.; Steenkamp, E.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, C.; Humm, E.; et al. Whole genome analyses suggests that Burkholderia sensu lato contains two additional novel genera (Mycetohabitans gen. nov. and Trinickia gen. nov.): Implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.H.; Lv, Y.Y.; Gao, Z.G.; Qiu, L.H. Pararobbsia silviterrae gen. nov., sp. nov., isolated from forest soil and reclassification of Burkholderia alpina as Pararobbsia alpina comb. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 1412–1420. [Google Scholar] [CrossRef]
- Khakhum, N.; Tapia, D.; Torres, A.G. Burkholderia mallei and glanders. In Defense Against Biological Attacks; Springer: Manhattan, NY, USA, 2019; pp. 161–183. [Google Scholar] [CrossRef]
- Gassiep, I.; Armostrong, M.; Norton, R. Human melioidosis. Clin. Microbiol. Rev. 2020, 33, e00006-19. [Google Scholar] [CrossRef]
- Hall, C.M.; Baker, A.L.; Sahl, J.W.; Mayo, M.; Scholz, H.C.; Kaestli, M.; Schupp, J.; Martz, M.; Settles, E.W.; Busch, J.D.; et al. Expanding the Burkholderia pseudomallei complex with the addition of two novel species: Burkholderia mayonis sp. nov. and Burkholderia savanae sp. nov. Appl. Environ. Microbiol. 2022, 11, e0158321. [Google Scholar] [CrossRef]
- Pereira-Andrade, J.; de Souza, H.G.; Carvalho-Ferreira, L.; Cnockaert, M.; De Canck, E.; Wieme, A.D.; Peeters, C.; Gross, E.; De Souza, J.T.; Santos-Marbach, P.A.; et al. Burkholderia perseverans sp. nov., a bacterium isolated from the Restinga ecosystem, a producer of volatile and diffusible compounds that inhibit plant pathogens. Braz. J. Microbiol 2021, 52, 2145–2152. [Google Scholar] [CrossRef]
- Mahenthiralingam, E.; Urban, T.A.; Goldberg, J.B. The multifarious, multireplicon Burkholderia cepacia complex. Nat. Rev. Microbiol. 2005, 3, 144–156. [Google Scholar] [CrossRef]
- Morales-Ruiz, L.M.; Rodriguez-Cisneros, M.; Kerber-Diaz, J.C.; Rojas-Rojas, F.U.; Ibarra, J.A.; Estrada-de los Santos, P. Burkholderia orbicola sp. nov., a novel species within the Burkholderia cepacia complex. Arch. Microbiol. 2022, 204, 178. [Google Scholar] [CrossRef]
- Rose, H.; Baldwin, A.; Dowson, C.G.; Mahenthiralingam, E. Biocide susceptibility of the Burkholderia cepacia complex. J. Ant. Chemother. 2009, 63, 502–510. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef]
- Parke, J.L.; Gurian-Sherman, D. Diversity of the Burkholderia cepacia complex and implications for risk assessment of biological control strains. Ann. Rev. Phytopathol. 2001, 39, 225–258. [Google Scholar] [CrossRef]
- Chain, P.S.G.; Denef, V.J.; Konstantinidis, K.T.; Vergez, L.M.; Agullo, L.; Reyes, V.L.; Hauser, L.; Cordova, M.; Gomez, L.; Gonzalez, M.; et al. Burkholderia xenovorans LB400 harbors a multi-replicon, 9.73-Mbp genome shaped for versatility. Proc. Natl. Acad. Sci. USA 2006, 103, 15280–15287. [Google Scholar] [CrossRef]
- O’Sullivan, L.A.; Weightman, A.J.; Jones, T.H.; Marchbank, A.M.; Tiedje, J.M.; Mahenthiralingam, E. Identifying the genetic basis of ecologically and biotechnologically useful functions of the bacterium Burkholderia vietnamiensis. Environ. Microbiol. 2007, 9, 1017–1034. [Google Scholar] [CrossRef]
- Bach, E.; Pereira-Passaglia, L.M.; Jiao, J.; Gross, H. Burkholderia in the genomic era: From taxonomy to the discovery of new antimicrobial secondary metabolites. Rev. Microbiol. 2022, 48, 121–160. [Google Scholar] [CrossRef]
- Depoorter, E.; Bull, M.J.; Peeters, C.; Coenye, T.; Vandmme, P.; Mahenthiralingam, E. Burkholderia: An update on taxonomy and biotechnological potential as antibiotic producers. Appl. Microbiol. Biotechnol. 2016, 100, 5215–5229. [Google Scholar] [CrossRef]
- Kerru, N.; Gummidi, L.; Maddila, S.; Gangu, K.K.; Jonnalagadda, S.B. A review on recent advances in nitrogen-containing molecules and their biological applications. Molecules 2020, 25, 1909. [Google Scholar] [CrossRef]
- Valenca, C.A.S.; Barbosa, A.A.T.; Souto, E.B.; Caramao, E.B.; Jain, S. Volatile nitrogenous compounds from bacteria: Source of novel bioactive compounds. Chem. Biodivers. 2021, 18, e2100549. [Google Scholar] [CrossRef]
- Prasad, J.; Pandey, P.; Anand, R.; Raghuwanshi, R. Drought Exposed Burkholderia seminalis JRBHU6 exhibits antimicrobial potential through pyrazine-1,4-dione derivatives targeting multiple bacterial and fungal proteins. Front. Microbiol. 2021, 12, 633036. [Google Scholar] [CrossRef]
- Petri, G.L.; Spano, V.; Spatola, R.; Holl, R.; Raimondi, M.V.; Barraja, P.; Montalbano, A. Bioactive pyrrole-based compound with target selectivity. Eur. J. Med. Chem. 2020, 208, 112783. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.E.; Teh, K.L. Antibacterial iminopyrrolidines from Burkholderia plantarii, a bacterial pathogen of rice. Org. Biomol. Chem. 2005, 3, 3540. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.E.; Greenwood, D.R.; Sarajioni, V. An antibacterial pyrazole derivative from Burkholderia glumae, a bacterial pathogen of rice. Phytochemistry 2008, 69, 2704–2707. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.; Chaudhari, A.; Prabha, R.; Shukla, R.; Singh, D.P. Microbial pyrrolnitrin: Natural metabolite with immense practical utility. Biomolecules 2019, 9, 443. [Google Scholar] [CrossRef] [PubMed]
- Arima, K.; Imanaka, H.; Kousaka, M.; Fukuta, A.; Tamura, G. Pyrrolnitrin, a new antibiotic substance produced by Pseudomonas. Agric. Biol. Chem. 1964, 28, 575–576. [Google Scholar] [CrossRef]
- Kilani, J.; Fillinger, S. Phenylpyrroles: 30 years, two molecules and (nearly) no resistance. Front. Microbiol. 2016, 7, 2014. [Google Scholar] [CrossRef]
- El-Banna, N.; Winkelmann, G. Pyrrolnitrin from Burkholderia cepacia: Antibiotic activity against fungi and novel activities against streptomycetes. J. Appl. Microbiol. 1998, 85, 69–78. [Google Scholar] [CrossRef]
- Sultan, M.Z.; Park, K.; Lee, S.Y.; Park, J.K.; Varughese, T.; Moon, S.S. Novel oxidized derivatives of antifungal pyrrolnitrin from the bacterium Burkholderia cepacia K87. J. Antibiot. 2008, 61, 420–425. [Google Scholar] [CrossRef]
- Jung, B.K.; Hong, S.J.; Park, G.S.; Kim, M.C.; Shin, J.H. Isolation of Burkholderia cepacia JBK9 with plant growth-promoting activity while producing pyrrolnitrin antagonistic to plant fungal diseases. Appl. Biol. Chem. 2018, 61, 173–180. [Google Scholar] [CrossRef]
- Webster, G.; Jones, C.; Mullins, A.J.; Mahenthiralingam, E. A rapid screening method for the detection of specialized metabolites from bacteria: Induction and suppression of metabolites from Burkholderia species. J. Microbiol. Methods 2020, 178, 106057. [Google Scholar] [CrossRef]
- Yan, J.; Liu, W.; Cai, J.; Wang, Y.; Li, D.; Hua, H.; Cao, H. Advances in phenazines over the past decade: Review of their pharmacological activities, mechanisms of action, biosynthetic pathways, and synthetic strategies. Mar. Drugs 2021, 19, 610. [Google Scholar] [CrossRef]
- Cartwright, D.K.; Chilton, W.S.; Benson, D.M. Pyrrolnitrin and phenazine production by Pseudomonas cepacia, strain 5.5B, a biocontrol agent of Rhizoctonia solani. Appl. Microbiol. Biotechnol. 1995, 43, 211–216. [Google Scholar] [CrossRef]
- Hendry, S.; Steinke, S.; Wittstein, K.; Stadler, M.; Harmrolfs, K.; Adewunmi, Y.; Sahukhal, G.; Elasri, M.; Thomashow, L.; Weller, D.; et al. Functional analysis of phenazines biosynthesis genes in Burkholderia spp. Appl. Environ. Microbiol. 2021, 87, e02348-20. [Google Scholar] [CrossRef]
- Han, J.W.; Kim, J.D.; Lee, J.M.; Ham, J.H.; Lee, D.; Kim, B.S. Structural elucidation and antimicrobial activity of new phencomycin derivatives isolated from Burkholderia glumae strain 411gr-6. J. Antibiot. 2014, 67, 721–723. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, M.; Du, J.; Huang, T.; Liu, J.; Dong, T.; Chen, Y. Isolation of Burkholderia sp. HQB-1, a promising biocontrol bacterium to protect banana against Fusarium wilt through phenazine-1-carboxylic acid secretion. Front. Microbiol. 2020, 11, 605152. [Google Scholar] [CrossRef]
- Chen, J.H.; Xiang, W.; Cao, K.X.; Lu, X.; Yao, S.C.; Hung, D.; Huang, R.S.; Li, L.B. Characterization of volatile organic compounds emitted from endophytic Burkholderia cenocepacia ETR-B22 by SPME-GC-MS and their inhibitory activity against various plant fungal pathogens. Molecules 2020, 25, 3765. [Google Scholar] [CrossRef]
- Xu, T.; Shi, L.; Zhang, Y.; Wang, K.; Yang, Z.; Ke, S. Synthesis and biological evaluation of marine alkaloid-oriented β-carboline analogues. Eur. J. Med. Chem. 2019, 25, 3765. [Google Scholar] [CrossRef]
- Huang, D.; Zhang, Z.; Li, Y.; Liu, F.; Huang, W.; Min, Y.; Wang, K.; Yang, J.; Cao, C.; Gong, Y.; et al. Carboline derivatives based on natural pityriacitrin as potential antifungal agents. Phytochemi. Lett. 2022, 48, 100–105. [Google Scholar] [CrossRef]
- Lin, Y.T.; Lee, C.C.; Leu, W.M.; Wu, J.J.; Huang, Y.C.; Meng, M. Fungicidal activity of volatile organic compounds emitted by Burkholderia gladioli strain BBB-01. Molecules 2021, 26, 745. [Google Scholar] [CrossRef]
- Hunter, W.J.; Manter, D.K. Antimicrobial properties of an oxidizer produced by Burkholderia cenocepacia P525. Curr. Microbiol. 2014, 68, 610–614. [Google Scholar] [CrossRef]
- Wu, Y.; Seyedsayamdost, M.R. The polyene natural product thailandamide A inhibits fatty acid biosynthesis in Gram-positive and Gram-negative bacteria. Biochemistry 2018, 57, 4247–4251. [Google Scholar] [CrossRef] [PubMed]
- Park, J.D.; Moon, K.; Miller, C.; Rose, J.; Xu, F.; Ebmeier, C.C.; Jacobsen, J.R.; Mao, D.; Old, W.M.; DeShazer, D.; et al. Thailandenes, cryptic polyene natural products isolated from Burkholderia thailandensis using phenotype-guided transposon mutagenesis. ACS Chem. Biol. 2020, 15, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Mahenthiralingam, E.; Song, L.; Sass, A.; White, J.; Wilmot, C.; Marchbank, A.; Boaisha, O.; Paine, J.; Knight, D.; Challis, G.L. Enacyloxins are products of an unusual hybrid modular polyketide synthase encoded by a cryptic Burkholderia ambifaria genomic island. Chem. Biol. 2011, 18, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Parmeggiani, A.; Krab, I.M.; Watanabe, T.; Nielsen, R.C.; Dahlberg, C.; Nyborg, J.; Nissen, P. Enacyloxin IIa pinpoints a binding pocket of elongation factor Tu for development of novel antibiotics. J. Biol. Chem. 2006, 281, 2893–2900. [Google Scholar] [CrossRef]
- Ross, C.; Opel, V.; Scherlach, K.; Hertweck, C. Biosynthesis of antifungal and antibacterial polyketides by Burkholderia gladioli in coculture with Rhizopus microsporus. Mycoses 2014, 57, 48–55. [Google Scholar] [CrossRef]
- Parker, W.L.; Rathnum, M.L.; Seiner, V.; Trejo, W.H.; Principe, P.A.; Sykes, R.B. Cepacin A and cepacin B, two new antibiotics produced by Pseudomonas cepacia. J. Antibiot. 1984, 37, 431–440. [Google Scholar] [CrossRef]
- Mullins, A.J.; Murray, J.A.H.; Bull, M.J.; Jenner, M.; Jones, C.; Webster, G.; Green, A.E.; Neill, D.R.; Connor, T.R.; Parkhill, J.; et al. Genome mining identifies cepacin as a plant-protective metabolite of the biopesticidal bacterium Burkholderia ambifaria. Nat. Microbiol. 2019, 4, 996–1005. [Google Scholar] [CrossRef]
- Kusumi, T.; Ohtani, I.; Nishiyama, K.; Kakisawa, H. Caryoynencins, potent antibiotics from a plant pathogen. Tetrahedron 1987, 28, 3981–3984. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Park, H.J.; Ishizuka, S.; Omata, K.; Hirama, M. Chemistry and antimicrobial activity of caryoynencin analogs. J. Med. Chem. 1995, 38, 5015–5022. [Google Scholar] [CrossRef]
- Florez, L.V.; Scherlach, K.; Gaube, P.; Ross, C.; Sitte, E.; Hermes, C.; Rodrigues, A.; Hertweck, C.; Kaltenpoth, M. Antibiotic-producing symbionts dynamically transition between plant pathogenicity and insect-defensive mutualism. Nat. Commun. 2017, 8, 15172. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637. [Google Scholar] [CrossRef]
- Finking, R.; Marahiel, M.A. Biosynthesis of nonribosomal peptides. Annu. Rev. Microbiol. 2004, 58, 453–488. [Google Scholar] [CrossRef]
- Hur, G.H.; Vickery, C.R.; Burkart, M.D. Explorations of catalytic domains in non-ribosomal peptide synthetase enzymology. Nat. Prod. Rep. 2012, 29, 1074–1098. [Google Scholar] [CrossRef]
- Jaremko, M.J.; Davis, T.D.; Corpuz, J.C.; Burkart, M.D. Type II non-ribosomal peptide synthetase proteins: Structure, mechanism, and protein–protein interactions. Nat. Prod. Rep. 2020, 37, 355–379. [Google Scholar] [CrossRef]
- Adler, C.; Corbalan, N.S.; Seyedsayamdost, M.R.; Pomares, M.F.; de Cristobal, R.E.; Clardy, J.; Kolter, R.; Vincent, P.A. Catecholate siderophores protect bacteria from pyochelin toxicity. PLoS ONE 2012, 7, e46754. [Google Scholar] [CrossRef]
- Ong, K.S.; Aw, Y.K.; Lee, L.H.; Yule, C.M.; Cheow, Y.L.; Lee, S.M. Burkholderia paludis sp. nov., an antibiotic-siderophore producing novel Burkholderia cepacia complex species, isolated from Malaysian tropical peat swamp soil. Front. Microbiol. 2016, 7, 2046. [Google Scholar] [CrossRef]
- Ong, K.S.; Cheow, Y.L.; Lee, S.M. The role of reactive oxygen species in the antimicrobial activity of pyochelin. J. Adv. Res. 2017, 8, 393–398. [Google Scholar] [CrossRef]
- da Araujo, F.D.S.; Araujo, W.L.; Eberlin, M.N. Potential of Burkholderia seminalis TC3.4.2R3 as biocontrol agent against Fusarium oxysporum evaluated by mass spectrometry imaging. J. Am. Soc. Mass Spectrom. 2017, 28, 901–907. [Google Scholar] [CrossRef]
- Meyer, J.M.; Hohnadel, D.; Halle, F. Cepabactin from Pseudomonas cepacia, a new type of siderophore. J. Gen. Microbiol. 1989, 135, 1479–1487. [Google Scholar] [CrossRef]
- Itoh, J.; Miyadoh, S.; Takahasi, S.; Amano, S.; Ezaki, N.; Yamada, Y. Studies on antibiotics BN-227 and BN-227-F, new antibiotics. I. Taxonomy, isolation and characterization. J. Antibiot. 1979, 32, 1089–1095. [Google Scholar] [CrossRef]
- Itoh, J.; Amano, S.; Ogawa, Y.; Kodama, Y.; Ezaki, N.; Yamada, Y. Studies on antibiotics BN-227 and BN-227-F, new antibiotics. II. Chemical structure of antibiotics BN-227 and BN-227-F. J. Antibiot. 1980, 33, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Darling, P.; Chan, M.; Cox, A.D.; Sokol, P. Siderophore production by cystic fibrosis isolates of Burkholderia cepacia. Infec. Immun. 1998, 66, 874–877. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.S. Iron acquisition mechanisms of the Burkholderia cepacia complex. BioMetals 2007, 20, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.; Foxfire, A.; Xu, J.; Baird, S.M.; Jia, J.; Delgado, K.H.; Shin, R.; Smith, L.; Lu, S.E. The siderophore product ornibactin is required for the bactericidal activity of Burkholderia contaminans MS14. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Rojas-Rojas, F.U.; Salazar-Gomez, A.; Vargas-Diaz, M.E.; Vasquez-Murrieta, M.S.; Hirsch, A.M.; De Mot, R.; Ghequire, M.G.K.; Ibarra, J.A.; Estrada-de los Santos, P. Broad-spectrum antimicrobial activity by Burkholderia cenocepacia TAtl-371, a strain isolated from the tomato rhizosphere. Microbiology 2018, 164, 1072–1086. [Google Scholar] [CrossRef]
- Lenz, K.D.; Klosterman, K.E.; Mukundan, H.; Kubicek-Sutherland, J.Z. Macrolides: From toxins to therapeutics. Toxins 2021, 13, 347. [Google Scholar] [CrossRef]
- Song, L.; Jenner, M.; Masschelein, J.; Jones, C.; Bull, M.J.; Harris, S.R.; Harkoorn, R.C.; Vocat, A.; Romero-Canelon, I.; Coupland, P.; et al. Discovery and biosynthesis of gladiolin: A Burkholderia gladioli antibiotic with promising activity against Mycobacterium tuberculosis. J. Am. Chem. Soc. 2017, 139, 7974–7981. [Google Scholar] [CrossRef]
- Riley, M.A. Bacteriocins, biology, ecology, and evolution. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 32–44. [Google Scholar]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, potent antimicrobial peptides and the fight against multi drug resistant species: Resistance is futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef]
- Yao, G.W.; Duarte, I.; Le, T.T.; Carmody, L.; LiPuma, J.J.; Young, R.; Gonzalez, C.F. A broad-host-range tailocin from Burkholderia cenocepacia. Appl. Environ. Microbiol. 2017, 83, e03414-16. [Google Scholar] [CrossRef]
- Principe, A.; Fernandez, M.; Torasso, M.; Godino, A.; Fischer, S. Effectiveness of tailocins produced by Pseudomonas fluorescens SF4c in controlling the bacterial-spot disease in tomatoes caused by Xanthomonas vesicatoria. Microbiol. Res. 2018, 212–213, 94–102. [Google Scholar] [CrossRef]
- Ghequire, M.G.K.; De Canck, E.; Wattiau, P.; Van Winge, I.; Loris, R.; Coenye, T.; De Mot, R. Antibacterial activity of a lectin-like Burkholderia cenocepacia protein. MicrobiologyOpen 2013, 2, 566–575. [Google Scholar] [CrossRef]
- Ghequire, M.G.K.; De Mot, R. Distinct colicin M-like bacteriocin-immunity pairs in Burkholderia. Sci. Rep. 2015, 5, 17368. [Google Scholar] [CrossRef]
- Marshall, K.; Shakya, S.; Greenhill, A.R.; Padilla, G.; Baker, A.; Warner, J.M. Antibiosis of Burkholderia ubonensis against Burkholderia pseudomallei, the causative agent for melioidosis. J. Trop. Med. Public Health 2010, 41, 904–912. [Google Scholar]
- Knappe, T.A.; Linne, U.; Zirah, S.; Rebuffat, S.; Xie, X.; Marahiel, M.A. Isolation and structural characterization of capistruin, a lasso peptide predicted from the genome sequence of Burkholderia thailandensis E264. Chem. Biol. 2008, 16, 1290–1298. [Google Scholar] [CrossRef]
- Rebufat, S.; Blond, A.; Destoumieux-Garzon, D.; Goulard, C.; Peduzzi, J. Microcin J25, from the macrocyclic to the lasso structure: Implications for biosynthetic, evolutionary, and biotechnological perspectives. Curr. Protein Pept. Sci. 2004, 5, 383–391. [Google Scholar] [CrossRef]
- Knappe, T.A.; Linne, U.; Robbel, L.; Marahiel, M.A. Insights into the biosynthesis and stability of the lasso peptide capistruin. Chem. Biol. 2009, 16, 1290–1298. [Google Scholar] [CrossRef]
- Cheung-Lee, W.L.; Parry, M.E.; Zong, C.; Jaramillo-Cartagena, A.; Darst, S.A.; Connell, N.D.; Rusoo, R.; Link, A.J. Discovery of ubonodin, an antimicrobial lasso peptide active against members of the Burkholderia cepacia complex. ChemBioChem. 2020, 21, 1335–1340. [Google Scholar] [CrossRef]
- Millanao, A.; Mora, A.; Villagra, N.; Bucarey, S.; Hidalgo, A. Biological effects of quinolones: A family of broad-spectrum antimicrobial agents. Molecules 2021, 26, 7153. [Google Scholar] [CrossRef]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Camara, M. Quinolones: From antibiotics to autoinducers. FEMS Microbiol. Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef]
- Wu, Y.; Seyedsayamdost, M.R. Synergy and target promiscuity drive structural divergence in bacterial alkylquinolone biosynthesis. Cell Chem. Biol. 2017, 24, 1437–1444. [Google Scholar] [CrossRef]
- Wang, Y.; Hoffmann, J.P.; Chou, C.W.; Honer zu Bentrup, K.; Fuselier, J.A.; Bitoun, J.P.; Wimley, W.C.; Morici, L.A. Burkholderia thailandensis outer membrane vesicles exert antimicrobial activity against drug-resistant and competitor microbial species. J. Microbiol. 2020, 58, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Yoshihisa, H.; Sato, Z.; Hirayama, F.; Konno, K.; Shirahama, H.; Suzui, T. Production of antibiotics by Pseudomonas cepacia as an agent for biological control of soilborne plant pathogens. Soil Biol. Biochem. 1989, 21, 723–728. [Google Scholar] [CrossRef]
- Saalim, M.; Villegas-Moreno, J.; Clark, B.R. Bacterial alkyl-4-quinolones: Discovery, structural diversity and biological properties. Molecules 2020, 25, 5689. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; Yamashita, T.; Furihata, K.; Nagai, K.; Suzuki, K.I.; Hayakawa, Y.; Shin-ya, K. Burkholone, a new cytotoxic antibiotic against IGF-I dependent cells from Burkholderia sp. J. Antibiot. 2007, 60, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Baserga, R.; Hongo, A.; Rubini, M.; Prisco, M.; Valentinis, B. The IGF-I receptor in cell growth, transformation and apoptosis. Biochim. Biophys. Acta 1997, 1332, F105–F126. [Google Scholar] [CrossRef]
- Niehs, S.P.; Kumpfmuller, J.; Dose, B.; Little, R.F.; Ishida, K.; Florez, L.V.; Kaltenpoth, M.; Hertweck, C. Insect-associated bacteria assemble the antifungal butanolide gladiofungin by non-canonical polyketide chain termination. Angew. Chem. Int. Ed. Engl. 2020, 59, 23122–23126. [Google Scholar] [CrossRef]
- Nakou, I.T.; Jenner, M.; Dashti, Y.; Romero-Canelón, I.; Masschelein, J.; Mahenthiralingam, E.; Challis, G.L. Genomics-driven discovery of a novel glutarimide antibiotic from Burkholderia gladioli reveals an unusual polyketide synthase chain release mechanism. Angew. Chem. Int. Ed. Engl. 2020, 59, 23145–23153. [Google Scholar] [CrossRef]
- Oka, M.; Yaginuma, K.; Numata, K.; Konishi, M.; Oki, T.; Kawaguchi, H. Glidobactins A, B and C, new antitumor antibiotics. II. Structure elucidation. J. Antibiot. 1988, 41, 1338–1350. [Google Scholar] [CrossRef]
- Oka, M.; Nishiyama, Y.; Ohta, S.; Kamei, H.; Konishi, M.; Miyaki, T.; Oki, T.; Kawaguchi, H. Glidobactins A, B and C, new antitumor antibiotics. I. Production, isolation, chemical properties and biological activity. J. Antibiot. 1988, 41, 1331–1337. [Google Scholar] [CrossRef]
- Shoji, J.; Hinoo, H.; Kato, T.; Hattori, T.; Hirooka, K.; Tawara, K.; Shiratori, O.; Terui, Y. Isolation of cepafungins I, II and III from Pseudomonas species. J. Antibiot. 1990, 43, 783–787. [Google Scholar] [CrossRef]
- Terui, Y.; Nishikawa, J.; Hinoo, H.; Kato, T.; Shoji, J. Structures of cepafungins I, II and III. J. Antibiot. 1990, 43, 788–795. [Google Scholar] [CrossRef]
- Schellenberg, B.; Bigles, L.; Dudler, R. Identification of genes involved in the biosynthesis of the cytotoxic compound glidobactin from a soil bacterium. Environ. Microbiol. 2007, 9, 1640–1650. [Google Scholar] [CrossRef]
- Biggins, J.B.; Kang, H.S.; Ternei, M.A.; DeShazer, D.; Brady, S.F. The chemical arsenal of Burkholderia pseudomallei is essential for pathogenicity. J. Am. Chem. Soc. 2014, 136, 9484–9490. [Google Scholar] [CrossRef]
- Lu, S.E.; Novak, J.; Austin, F.W.; Gu, G.; Ellis, D.; Kirk, M.; Wilson-Stanford, S.; Tonelli, M.; Smith, L. Occidiofungin, a unique antifungal glycopeptide produced by a strain of Burkholderia contaminans. Biochemistry 2009, 48, 8312–8321. [Google Scholar] [CrossRef]
- Ellis, D.; Gosai, J.; Emrick, C.; Heintz, R.; Romans, L.; Gordon, D.; Lu, S.E.; Austin, F.; Smith, L. Occidiofungin’s chemical stability and in vitro potency against Candida species. Antimicrob. Agents Chemother. 2012, 56, 765–769. [Google Scholar] [CrossRef]
- Ravichandran, A.; Geng, M.; Hull, K.G.; Romo, D.; Lu, S.E.; Albee, A.; Nutter, C.; Gordon, D.M.; Ghannoum, M.A.; Lockless, S.W.; et al. Occidiofungin, and actin binding antifungal with in vivo efficacy in a vulvovaginal candidiasis infection. bioRxiv 2018, 368720. [Google Scholar] [CrossRef]
- Emrick, D.; Ravichandran, A.; Gosai, J.; Lu, S.; Gordon, D.M.; Smith, L. The antifungal occidiofungin triggers an apoptotic mechanism of cell death in yeast. J. Nat. Prod. 2013, 76, 829–838. [Google Scholar] [CrossRef]
- Ma, J.; Guo, F.; Jin, Z.; Geng, M.; Ju, M.; Ravichandran, A.; Orugunty, R.; Smith, L.; Zhu, G.; Zhang, H. Novel antiparasitic activity of the antifungal lead occidiofungin. Antimicrob. Agents Chemother. 2020, 64, e00244-20. [Google Scholar] [CrossRef]
- Wang, X.Q.; Liu, A.X.; Guerrero, A.; Liu, J.; Yu, X.Q.; Deng, P.; Ma, L.; Baird, S.M.; Smith, L.; Lu, S.E. Occidiofungin is an important component responsible for the antifungal activity of Burkholderia pyrrocinia strain Lyc2. J. Appl. Microbiol. 2016, 120, 607–618. [Google Scholar] [CrossRef]
- Hing, S.L.; Ravichandran, A.; Escano, J.; Cooley, J.; Autin, F.; Lu, S.E.; Pruett, S.; Smith, L. Toxicological evaluation of occidiofungin against mice and human cancer cell lines. Sci. Res. 2014, 5, 1085–1093. [Google Scholar] [CrossRef]
- Lim, Y.; Suh, J.W.; Kim, S.; Hyun, B.; Kim, C.; Lee, C. Cepacidine A, a novel antifungal antibiotic produced by Pseudomonas cepacia. II. Physico-chemical properties and structure elucidation. J. Antibiot. 1994, 47, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Kim, S.; Hyun, B.; Suh, J.W.; Yon, C.; Kim, C.; Lim, Y.; Kim, C. Cepacidine A, a novel antifungal antibiotic produced by Pseudomonas cepacia. I. Taxonomy, production, isolation and biological activity. J. Antibiot. 1994, 47, 1402–1405. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Suh, J.W.; Cho, Y.H. Immunosuppressive activity of cepacidine A, a novel antifungal antibiotic produced by Pseudomonas cepacia. J. Microbiol. Biotechnol. 1999, 9, 672–674. [Google Scholar]
- Lee, C.H.; Kempf, H.J.; Lim, Y.; Cho, Y.H. Biocontrol activity of Pseudomonas cepacia AF2001 and anthelmintic activity of its novel metabolite, cepacidine A. J. Microbiol. Biotechnol. 2000, 10, 568–571. [Google Scholar]
- Kang, Y.; Carlson, R.; Tharpe, W.; Schell, M.A. Characterization of genes involved in biosynthesis of a novel antibiotic from Burkholderia cepacia BC11 and their role in biological control of Rhizoctonia solani. Appl. Environ. Microbiol. 1998, 64, 3939–3947. [Google Scholar] [CrossRef]
- Dose, B.; Niehs, S.P.; Scherlach, K.; Florez, L.V.; Kaltenpoth, M.; Hertweck, C. Unexpected bacterial origin of the antibiotic icosalide: Two-tailed depsipeptide assembly in multifarious Burkholderia symbionts. ACS Chem. Biol. 2018, 13, 2414–2420. [Google Scholar] [CrossRef]
- Jenner, M.; Jian, X.; Dashti, Y.; Masschelein, J.; Hobson, C.; Roberts, D.; Jones, C.; Harris, S.; Parkhill, J.; Raja, H.A.; et al. An unusual Burkholderia gladioli double chain-initiating nonribosomal peptide synthetase assembles “fungal” icosalide antibiotics. Chem. Sci. 2019, 10, 5489–5494. [Google Scholar] [CrossRef]
- Boros, C.; Smith, C.J.; Vasina, Y.; Che, Y.; Dix, A.B.; Darveaux, B.; Pearce, C. Isolation and identification of the icosalides—Cyclic peptolides with selective antibiotic and cytotoxic activities. J. Antibiot. 2006, 59, 486–494. [Google Scholar] [CrossRef]
- Chandler, J.R.; Truong, T.T.; Silva, P.M.; Seyedsayamdost, M.R.; Carr, G.; Radey, M.; Jacobs, M.A.; Sims, E.H.; Clardy, J.; Greenberg, E.P. Bactobolin resistance is conferred by mutations in the L2 ribosomal protein. mBio 2012, 3, e00499-12. [Google Scholar] [CrossRef]
- Bisacchi, G.S.; Hockstein, D.R.; Koster, W.H.; Parker, W.L.; Rathnum, M.L.; Unger, S.E. Xylocandin: A new complex of antifungal peptides. II. Structural studies and chemical modifications. J. Antibiot. 1987, 40, 1520–1529. [Google Scholar] [CrossRef]
- Meyers, E.; Bissachi, G.S.; Dean, L.; Liu, W.C.; Minassian, B.; Slusarchyk, D.S.; Sykes, R.B.; Tanaka, S.K.; Trejo, W. Xylocandin: A new complex of antifungal peptides. I. Taxonomy, isolation and biological activity. J. Antibiot. 1987, 40, 1515–1519. [Google Scholar] [CrossRef]
- Jenul, C.; Sieber, S.; Daeppen, C.; Mathew, A.; Lardi, M.; Pessi, G.; Hoepfner, D.; Neuburger, M.; Linden, A.; Gademann, K.; et al. Biosynthesis of fragin is controlled by a novel quorum sensing signal. Nat. Commun. 2018, 9, 1297. [Google Scholar] [CrossRef]
- Biggins, J.B.; Liu, X.; Feng, Z.; Brady, S.F. Metabolites from the induced expression of cryptic single operons found in the genome of Burkholderia pseudomallei. J. Am. Chem. Soc. 2011, 133, 1638–1641. [Google Scholar] [CrossRef]
- Florez, L.V.; Scherlach, K.; Miller, I.J.; Rodrigues, A.; Kwan, J.C.; Hertweck, C.; Kaltenpoth, M. An antifungal polyketide associated with horizontally acquired genes supports symbiont-mediated defense in Lagria villosa beetles. Nat. Commun. 2018, 9, 2478. [Google Scholar] [CrossRef]
- Imada, A.; Kitano, K.; Kintaka, K.; Muroi, M.; Asai, M. Sulfazecin and isosulfazecin, novel β-lactam antibiotics of bacterial origin. Nature 1981, 289, 590–591. [Google Scholar] [CrossRef]
- Loveridge, E.J.; Jones, C.; Bull, M.J.; Moody, S.C.; Kahl, M.W.; Khan, Z.; Neilson, L.; Tomeva, M.; Adams, S.E.; Wood, A.C.; et al. Reclassification of the specialized metabolite producer Pseudomonas mesoacidophila ATCC 31433 as a member of the Burkholderia cepacia complex. J. Bacteriol. 2017, 199, e00125-17. [Google Scholar] [CrossRef]
- Imada, A.; Kintaka, K.; Nakao, M.; Shinagawa, S. Bulgecin, a bacterial metabolite which in concert with β-lactam antibiotics causes bulge formation. J. Antibiot. 1982, 35, 1400–1403. [Google Scholar] [CrossRef]
- Quan, C.S.; Zheng, W.; Liu, Q.; Ohta, Y.; Fan, S.D. Isolation and characterization of a novel Burkholderia cepacia with strong antifungal activity against Rhizoctornia solani. Appl. Environ. Biotechnol. 2006, 72, 1276–1284. [Google Scholar] [CrossRef]
- Lee, J.; Wu, J.; Deng, Y.; Wang, J.; Wang, C.; Wang, J.; Chang, C.; Dong, Y.; Williams, P.; Zhang, L.H. A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol. 2013, 9, 339–343. [Google Scholar] [CrossRef]
- Ye, L.; Cornelis, P.; Guillemyn, K.; Ballet, S.; Christophersen, C.; Hammerich, O. Structure revision of N-mercapto-4-formylcarbostyril produced by Pseudomonas fluorescens G308 to 2-(2-hydroxyphenyl)thiazole-4-carbaldehyde (aeruginaldehyde). Nat. Prod. Commun. 2014, 9, 789–794. [Google Scholar] [CrossRef]
- Trottmann, F.; Franke, J.; Ishida, K.; Garcia-Altares, M.; Hertweck, C. A pair of bacterial siderophores releases and traps an intercellular signal molecule: An unusual case of natural nitrone bioconjugation. Angew. Chem. Int. Ed. 2019, 58, 200–2004. [Google Scholar] [CrossRef] [PubMed]
- Eustaquio, A.S.; Janso, J.E.; Ratnayake, A.S.; O’Donnell, C.J.; Koehn, F.E. Spliceostatin hemiketal biosynthesis in Burkholderia spp. is catalyzed by an iron/-ketoglutarate-dependent dioxygenase. Proc. Natl. Acad. Sci. USA 2014, 111, E3376–E3385. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ratnayake, A.S.; Janso, J.E.; Min, H.; Yang, H.Y.; Loganzo, F.; Shor, B.; O’Donnell, J.C.; Koehn, F.E. Cytotoxic spliceostatins from Burkholderia sp. and their semisynthetic analogues. J. Nat. Prod. 2014, 77, 1864–1870. [Google Scholar] [CrossRef] [PubMed]
- Prasad, C. Bioactive cyclic dipeptides. Peptides 1995, 16, 151–164. [Google Scholar] [CrossRef]
- Wang, J.H.; Quan, C.S.; Qi, X.H.; Li, X.; Fan, S.D. Determination of diketopiperazines of Burkholderia cepacia CF-66 by gas chromatography–mass spectrometry. Anal. Bioanal. Chem. 2010, 396, 1773–1779. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Chiarelli, L.R.; Makarov, V.; Brackman, G.; Israyilova, A.; Azzalin, A.; Forneris, F.; Riabova, O.; Savina, S.; Coenye, T.; et al. Discovery of new diketopiperazines inhibiting Burkholderia cenocepacia quorum sensing in vitro and in vivo. Sci. Rep. 2016, 6, 32487. [Google Scholar] [CrossRef]
- Rojas-Rojas, F.U.; Sanchez-Lopez, D.; Tapia-Garcia, E.Y.; Arroyo-Herrera, I.; Maymon, M.; Humm, E.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; et al. Draft genome of Burkholderia cenocepacia TAtl-371, a strain from the Burkholderia cepacia complex retains antagonism in different carbon and nitrogen sources. Curr. Microbiol. 2019, 76, 566–574. [Google Scholar] [CrossRef]
- Jiao, Y.; Yoshihara, T.; Ishikuri, S.; Uchino, H.; Ichihara, A. Structural identification of cepaciamide A, a novel fungitoxic compound from Pseudomonas cepacia D-202. Tetrahedron 1996, 37, 1039–1042. [Google Scholar] [CrossRef]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef]
- Swain, D.M.; Yadav, S.K.; Tyagi, I.; Kumar, R.; Kumar, R.; Ghosh, S.; Das, J.; Jha, G. A prophage tail-like protein is deployed by Burkholderia bacteria to feed on fungi. Nat. Commun. 2017, 8, 404. [Google Scholar] [CrossRef]
- Kang, J.G.; Shin, S.Y.; Kim, M.J.; Bajpai, V.; Maheshwari, D.K.; Kang, S.C. Isolation and anti-fungal activities of 2-hydroxymethyl-chroman-4-one produced by Burkholderia sp. MSSP. J. Antibiot. 2004, 57, 726–731. [Google Scholar] [CrossRef]
- Kirinuki, T.; Ichiba, T.; Katamaya, K. General survey of action site of altericidins on metabolism of Alternaria kikuchiana and Ustilago maydis. J. Pestic. Sci. 1984, 9, 601–610. [Google Scholar] [CrossRef]
- Abdel-Mawgoud, A.M.; Lepine, F.; Deziel, E. Rhamnolipids: Diversity of structures, microbial origins, and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef]
- Elshikh, M.; Funston, S.; Chebbi, A.; Ahmed, S.; Marchant, R.; Banat, I.M. Rhamnolipids from non-pathogenic Burkholderia thailandensis E264: Physicochemical characterization, antimicrobial and antibiofilm efficacy against oral hygiene related pathogens. N. Biotechnol. 2017, 36, 26–36. [Google Scholar] [CrossRef]
- Hormann, B.; Muller, M.M.; Syldatk, C.; Hausmann, R. Rhamnolipid production by Burkholderia plantarii DSM 9509T. Eur. J. Lipid Sci. Technol. 2010, 112, 674–680. [Google Scholar] [CrossRef]
- Costa, S.; Deziel, E.; Lepine, F. Characterization of rhamnolipid production by Burkholderia glumae. Lett. Appl. Microbiol. 2011, 53, 620–627. [Google Scholar] [CrossRef]
- Funston, S.J.; Tsaousi, K.; Rudden, M.; Smyth, T.J.; Stevenson, P.S.; Marchant, R.; Banat, I.M. Characterizing rhamnolipid production in Burkholderia thailandensis E264, a non-pathogenic producer. Appl. Microbiol. Biotechnol. 2016, 100, 7945–7956. [Google Scholar] [CrossRef]
- Haubler, S.; Nimtz, M.; Domke, T.; Wray, V.; Steinmetz, I. Purification and characterization of a cytotoxic exolipid of Burkholderia pseudomallei. Infect. Immun. 1998, 66, 1588–1593. [Google Scholar]
- Tawfik, K.A.; Jeffs, P.; Bray, B.; Dubay, G.; Falkinham, J.O.; Mesbah, M.; Youssef, D.; Khalifa, S.; Schmidt, E.W. Burkholdines 1097 and 1229, potent antifungal peptides from Burkholderia ambifaria 2.2N. Org. Lett. 2010, 12, 664–666. [Google Scholar] [CrossRef]
- Lin, Z.; Falkinham, J.O.; Tawfik, K.A.; Jeffs, P.; Bray, B.; Dubay, G.; Cox, J.E.; Schmidt, E.W. Burkholdines from Burkholderia ambifaria: Antifungal agents and possible virulence factors. J. Nat. Prod. 2012, 75, 1518–1523. [Google Scholar] [CrossRef]
- Wang, M.; Tachibana, S.; Murai, Y.; Li, L.; Lau, S.Y.L.; Cao, M.; Zhu, G.; Hashimoto, M.; Hashidoko, Y. Indole-3-acetic acid produced by Burkholderia heleia acts as a phenylacetic acid antagonist to disrupt tropolone biosynthesis in Burkholderia plantarii. Sci. Rep. 2016, 6, 22596. [Google Scholar] [CrossRef] [PubMed]
- Azegami, K.; Nishiyama, K.; Watanabe, Y.; Suzuki, T.; Yoshida, M.; Nose, K.; Toda, S. Tropolone as a root growth-inhibitor produced by a plant pathogenic Pseudomonas sp. causing seedling blight of rice. Jpn. J. Phytopathol. 1985, 51, 315–317. [Google Scholar] [CrossRef]
- Wakimoto, S.; Hirayae, K.; Tsuchiya, K.; Kushima, Y.; Furuya, N.; Matsuyama, N. Production of antibiotics by plant pathogenic pseudomonads. Ann. Phytopathol. Soc. Jpn. 1986, 52, 835–842. [Google Scholar] [CrossRef]
- Abe, M.; Nakazawa, T. Characterization of hemolytic and antifungal substance, cepalycin, from Pseudomonas cepacia. Microbiol. Immunol. 1994, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, E.; Roberts, I.N.; Montecchia, M.S.; Gutierrez-Boem, F.H.; Gomez, F.M.; Ruiz, J.A. A novel Burkholderia ambifaria strain able to degrade the mycotoxin fusaric acid and to inhibit Fusarium spp. growth. Microbiol. Res. 2018, 206, 50–59. [Google Scholar] [CrossRef]
- Guttenberger, N.; Blankenfeldt, W.; Breinbauer, R. Recent developments in the isolation, biological function, biosynthesis, and synthesis of phenazine natural products. Bioorg. Med. Chem. 2017, 25, 6149–6166. [Google Scholar] [CrossRef]
- Mullins, A.J.; Mahenthiralingam, E. The hidden genomic diversity, specialized metabolic capacity, and revised taxonomy of Burkholderia sensu lato. Front. Microbiol. 2021, 12, 726847. [Google Scholar] [CrossRef]
- Coulon, P.M.L.; Groleau, M.C.; Deziel, E. Potential of the Burkholderia cepacia complex to produce 4-hydroxy-3-methyl-2-alkyquinolines. Front. Cell. Infect. Microbiol. 2019, 9, 33. [Google Scholar] [CrossRef]
- Matthew, A.; Jenul, C.; Carlier, A.L.; Eberl, L. The role of siderophores in metal homeostasis of member of the genus Burkholderia. Environ. Microbiol. Rep. 2016, 8, 103–109. [Google Scholar] [CrossRef]
- Li, X.; Quan, C.S.; Fan, S.D. Antifungal activity of a novel compound from Burkholderia cepacia against plant pathogenic fungi. Lett. Appl. Microbiol. 2007, 45, 508–514. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).