


Two-Dimensional-PAGE Coupled with nLC-MS/MS-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in MCF7 Breast Cancer Cells Transfected for JTB Protein Silencing


 ,
,  and
and
Abstract
1. Introduction
2. Results and Discussion
2.1. JTB Silencing Is Associated with Neoplastic Abilities of MCF7 Transfected Cells
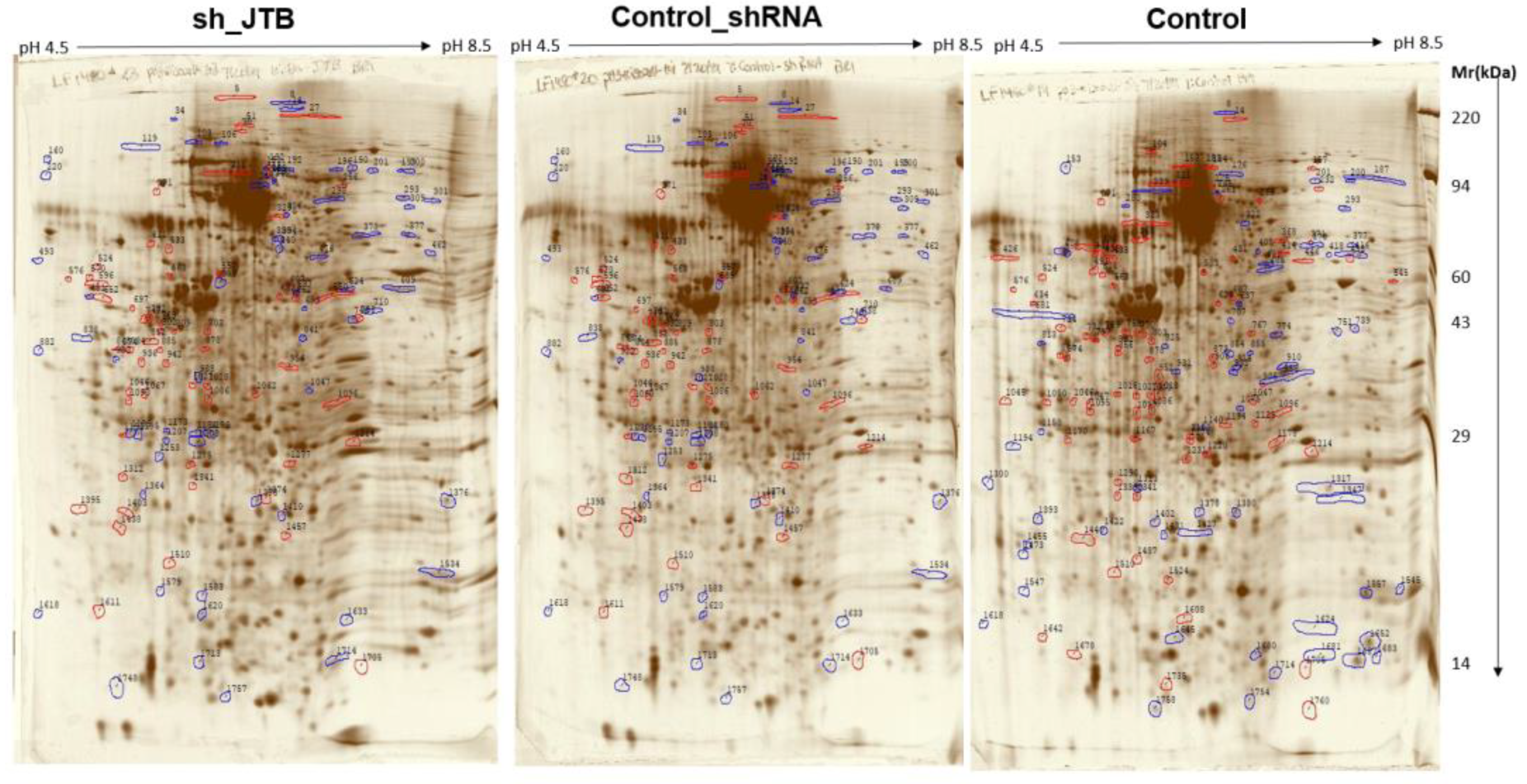
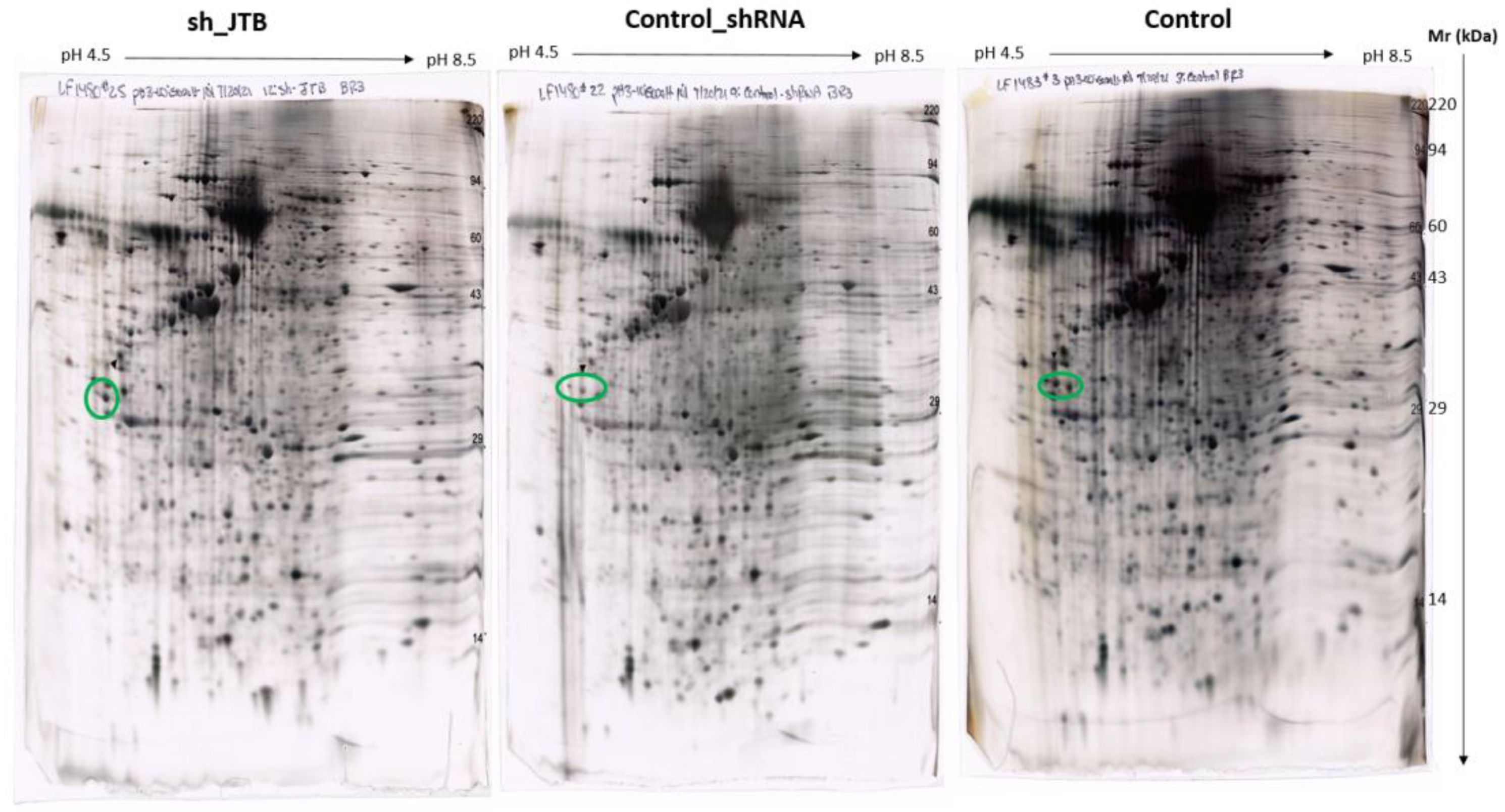
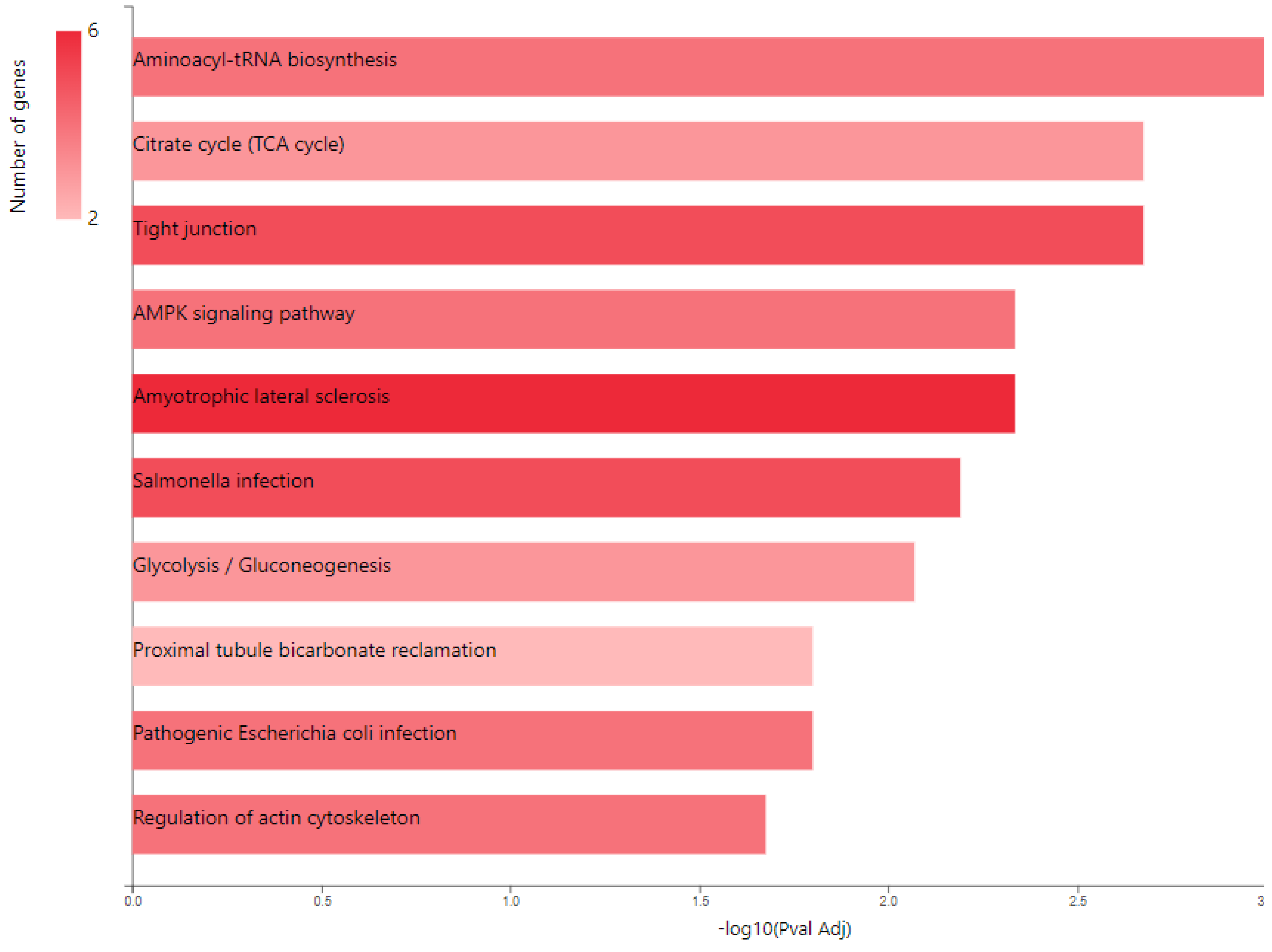
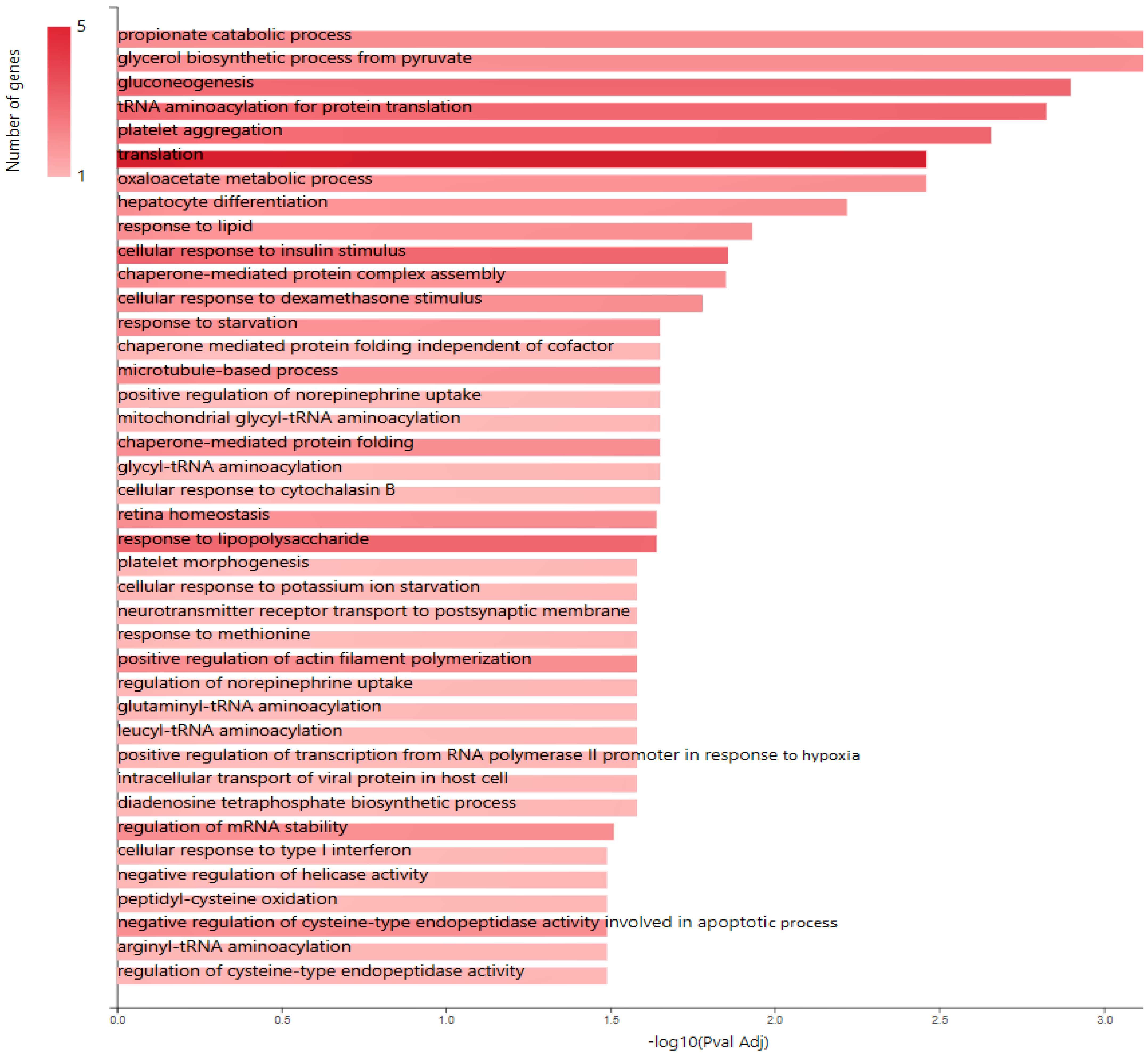
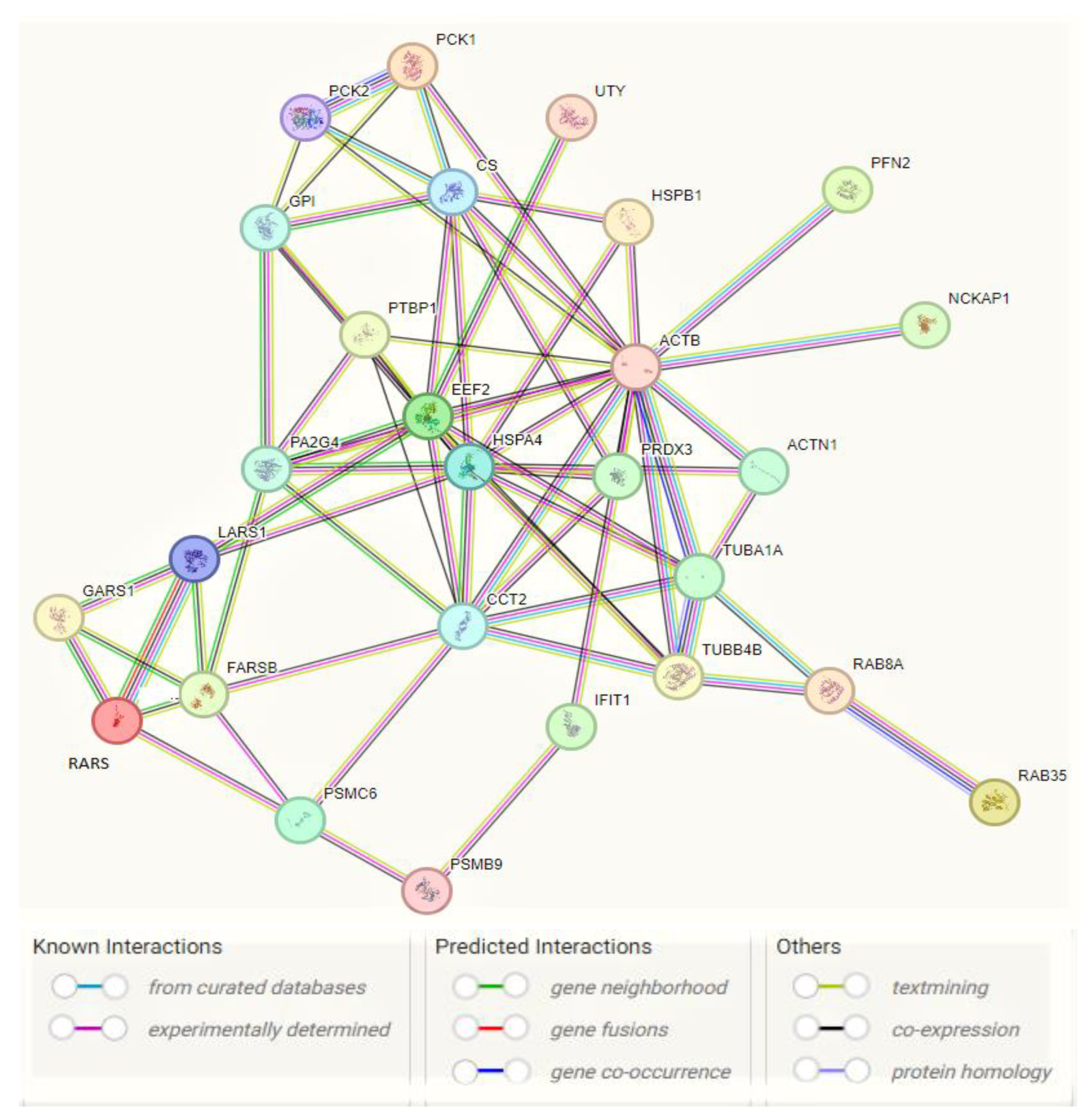
2.2. Glucose Metabolism Reprogramming in JTB Downregulated Condition
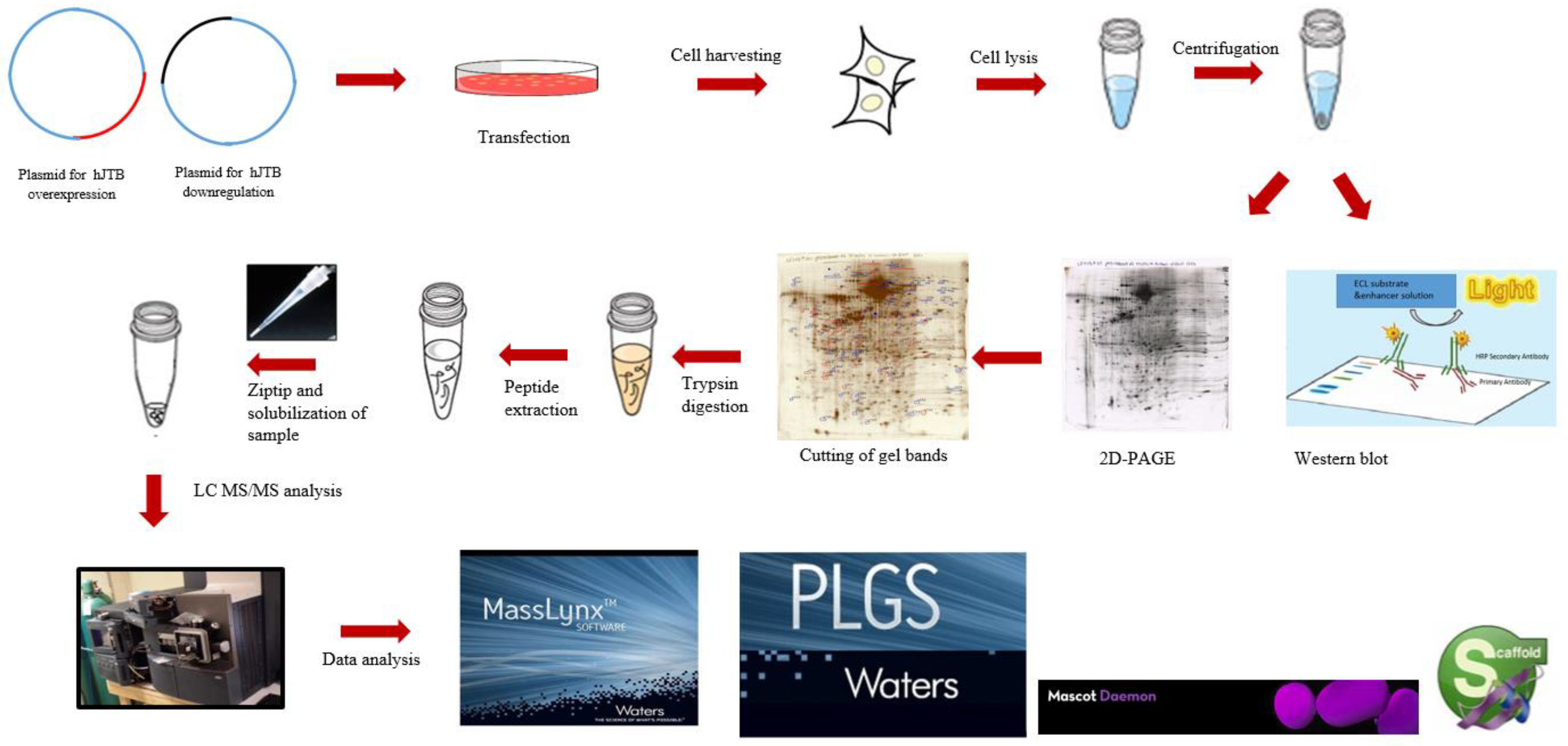
3. Materials and Methods
3.1. Cell Culture
3.2. Plasmids for Downregulation
3.3. Transfection into MCF7 Cells
3.4. Western Blot Analysis
3.5. 2D-PAGE & Proteomic Analysis
3.6. Data Sharing
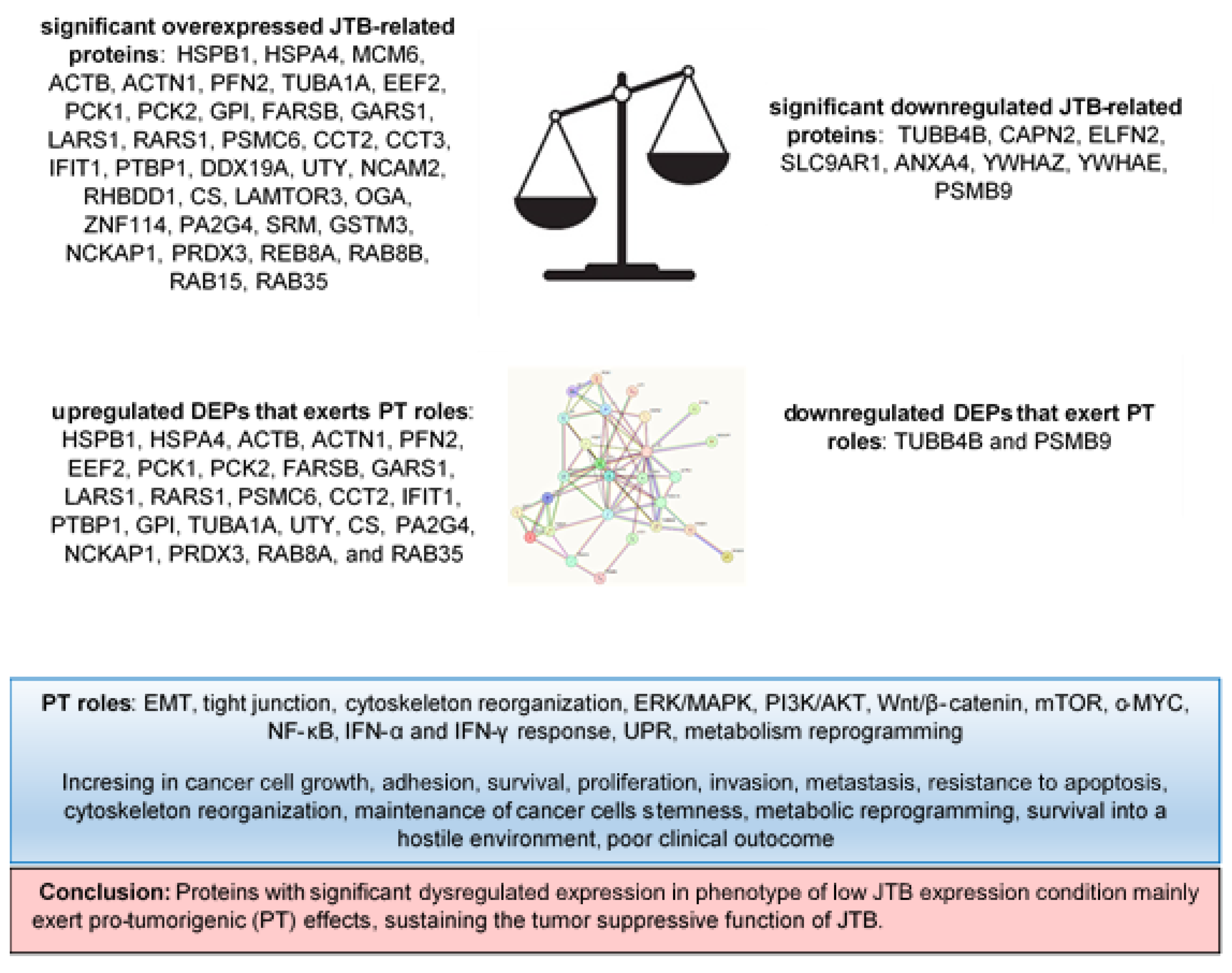
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, R.; Sultan, A.; Ishrat, R.; Haque, S.; Khan, N.J.; Prieto, M.A. Identification of New Key Genes and Their Association with Breast Cancer Occurrence and Poor Survival Using In Silico and In Vitro Methods. Biomedicines 2023, 11, 1271. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, S.; Osawa, M.; Omine, M.; Ishikawa, F. JTB: A novel membrane protein gene at 1q21 rearranged in a jumping translocation. Oncogene 1999, 18, 2085–2090. [Google Scholar] [CrossRef] [PubMed]
- Tyszkiewicz, T.; Jarzab, M.; Szymczyk, C.; Kowal, M.; Krajewska, J.; Jaworska, M.; Fraczek, M.; Krajewska, A.; Hadas, E.; Swierniak, M.; et al. Epidermal differentiation complex (locus 1q21) gene expression in head and neck cancer and normal mucosa. Folia Histochem. et Cytobiol. 2014, 52, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Hogenbirk, M.A.; Heideman, M.R.; de Rink, I.; Velds, A.; Kerkhoven, R.M.; Wessels, L.F.A.; Jacobs, H. Defining chromosomal translocation risks in cancer. Proc. Natl. Acad. Sci. USA 2016, 113, E3649–E3656. [Google Scholar] [CrossRef] [PubMed]
- Stankiewicz, P.; Cheung, S.; Shaw, C.; Saleki, R.; Szigeti, K.; Lupski, J. The donor chromosome breakpoint for a jumping translocation is associated with large low-copy repeats in 21q21.3. Cytogenet. Genome Res. 2003, 101, 118–123. [Google Scholar] [CrossRef]
- Kanome, T.; Itoh, N.; Ishikawa, F.; Mori, K.; Kim-Kaneyama, J.R.; Nose, K.; Shibanuma, M. Characterization of Jumping translocation breakpoint (JTB) gene product isolated as a TGF-β1-inducible clone involved in regulation of mitochondrial function, cell growth and cell death. Oncogene 2007, 26, 5991–6001. [Google Scholar] [CrossRef]
- Platica, O.; Chen, S.; Ivan, E.; Lopingco, M.C.; Holland, J.F.; Platica, M. PAR, a novel androgen regulated gene, ubiquitously expressed in normal and malignant cells. Int. J. Oncol. 2000, 16, 1055–1116. [Google Scholar] [CrossRef]
- Platica, M.; Ionescu, A.; Ivan, E.; Holland, J.F.; Mandeli, J.; Platica, O. PAR, a protein involved in the cell cycle, is functionally related to chromosomal passenger proteins. Int. J. Oncol. 2011, 38, 777–785. [Google Scholar] [CrossRef]
- Pan, J.-S.; Cai, J.-Y.; Xie, C.-X.; Zhou, F.; Zhang, Z.-P.; Dong, J.; Xu, H.-Z.; Shi, H.-X.; Ren, J.-L. Interacting with HBsAg compromises resistance of Jumping translocation breakpoint protein to ultraviolet radiation-induced apoptosis in 293FT cells. Cancer Lett. 2009, 285, 151–156. [Google Scholar] [CrossRef]
- de Abreu Pereira, D.; Sandim, V.; Fernandes, T.F.; Almeida, V.H.; Rocha, M.R.; do Amaral, R.J.; Rossi, M.I.D.; Kalume, D.E.; Zingali, R.B. Proteomic Analysis of HCC-1954 and MCF-7 Cell Lines Highlights Crosstalk between αv and β1 Integrins, E-Cadherin and HER-2. Int. J. Mol. Sci. 2022, 23, 10194. [Google Scholar] [CrossRef]
- Somasekhara, D.; Dammalli, M.; Nadumane, V.K. Proteomic Analysis of Human Breast Cancer MCF-7 Cells to Identify Cellular Targets of the Anticancer Pigment OR3 from Streptomyces coelicolor JUACT03. Appl. Biochem. Biotechnol. 2023, 195, 236–252. [Google Scholar]
- Jayathirtha, M.; Whitham, D.; Alwine, S.; Donnelly, M.; Neagu, A.-N.; Darie, C.C. Investigating the Function of Human Jumping Translocation Breakpoint Protein (hJTB) and Its Interacting Partners through In-Solution Proteomics of MCF7 Cells. Molecules 2022, 27, 8301. [Google Scholar] [CrossRef]
- Jayathirtha, M.; Neagu, A.-N.; Whitham, D.; Alwine, S.; Darie, C.C. Investigation of the effects of overexpression of jumping translocation breakpoint (JTB) protein in MCF7 cells for potential use as a biomarker in breast cancer. Am. J. Cancer Res. 2022, 12, 1784–1823. [Google Scholar]
- Jayathirtha, M.; Channaveerappa, D.; Darie, C. Investigation and Characterization of the Jumping Translocation Breakpoint (JTB) Protein using Mass Spectrometry based Proteomics. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Jayathirtha, M.; Neagu, A.-N.; Whitham, D.; Alwine, S.; Darie, C.C. Investigation of the effects of downregulation of jumping translocation breakpoint (JTB) protein expression in MCF7 cells for potential use as a biomarker in breast cancer. Am. J. Cancer Res. 2022, 12, 4373–4398. [Google Scholar] [PubMed]
- Aslebagh, R.; Channaveerappa, D.; Arcaro, K.F.; Darie, C.C. Comparative two-dimensional polyacrylamide gel electrophoresis (2D-PAGE) of human milk to identify dysregulated proteins in breast cancer. Electrophoresis 2018, 39, 1723–1734. [Google Scholar] [CrossRef]
- Arrigo, A.-P.; Gibert, B. HspB1, HspB5 and HspB4 in Human Cancers: Potent Oncogenic Role of Some of Their Client Proteins. Cancers 2014, 6, 333–365. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Liu, T.T.; Wang, H.H.; Hong, H.M.; Yu, A.L.; Feng, H.P.; Chang, W.W. Hsp27 participates in the maintenance of breast cancer stem cells through regulation of epithelial-mesenchymal transition and nuclear factor-κB. Breast Cancer Res. 2011, 13, R101. [Google Scholar] [CrossRef] [PubMed]
- Shang, B.-B.; Chen, J.; Wang, Z.-G.; Liu, H. Significant correlation between HSPA4 and prognosis and immune regulation in hepatocellular carcinoma. PeerJ 2021, 9, e12315. [Google Scholar] [CrossRef]
- Zhang, M.; Dai, W.; Li, Z.; Tang, L.; Chen, J.; Chen, C. HSPA4 Knockdown Retarded Progression and Development of Colorectal Cancer. Cancer Manag. Res. 2021, 13, 4679–4690. [Google Scholar] [CrossRef]
- Fan, G.; Tu, Y.; Wu, N.; Xiao, H. The expression profiles and prognostic values of HSPs family members in Head and neck cancer. Cancer Cell Int. 2020, 20, 220. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wu, Q.; Tang, Z.; Tan, Z.; Pu, D.; Tan, W.; Zhang, W.; Liu, S. Comprehensive Analysis of Necroptosis-Related Genes as Prognostic Factors and Immunological Biomarkers in Breast Cancer. J. Pers. Med. 2022, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Morisaki, T.; Yashiro, M.; Kakehashi, A.; Inagaki, A.; Kinoshita, H.; Fukuoka, T.; Kasashima, H.; Masuda, G.; Sakurai, K.; Kubo, N.; et al. Comparative proteomics analysis of gastric cancer stem cells. PLoS ONE 2014, 9, e110736. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Guan, Y.; Li, Y.-K.; Wu, Q.; Tang, X.-J.; Zeng, X.; Ling, H.; Zou, J. The DNA replication regulator MCM6: An emerging cancer biomarker and target. Clin. Chim. Acta 2021, 517, 92–98. [Google Scholar] [CrossRef]
- Liu, M.; Hu, Q.; Tu, M.; Wang, X.; Yang, Z.; Yang, G.; Luo, R. MCM6 promotes metastasis of hepatocellular carcinoma via MEK/ERK pathway and serves as a novel serum biomarker for early recurrence. J. Exp. Clin. Cancer Res. 2018, 37, 10. [Google Scholar] [CrossRef]
- Gu, Y.; Tang, S.; Wang, Z.; Cai, L.; Lian, H.; Shen, Y.; Zhou, Y. A pan-cancer analysis of the prognostic and immunological role of β-actin (ACTB) in human cancers. Bioengineered 2021, 12, 6166–6185. [Google Scholar] [CrossRef]
- Misawa, A.; Takayama, K.I.; Fujimura, T.; Homma, Y.; Suzuki, Y.; Inoue, S. Androgen-induced lncRNA POTEF-AS1 regulates apoptosis-related pathway to facilitate cell survival in prostate cancer cells. Cancer Sci. 2017, 108, 373–379. [Google Scholar] [CrossRef]
- Javaid, N.; Choi, S. Toll-like Receptors from the Perspective of Cancer Treatment. Cancers 2020, 12, 297. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, J.; Chen, T.; Wang, J.; Wang, Y.; Yu, Z.; Zhao, K.; Zheng, K.; Chen, Y.; Wang, Z.; et al. α-Actinin1 promotes tumorigenesis and epithelial-mesenchymal transition of gastric cancer via the AKT/GSK3β/β-Catenin pathway. Bioengineered 2021, 12, 5688–5704. [Google Scholar] [CrossRef]
- Kovac, B.; Mäkelä, T.P.; Vallenius, T. Increased α-actinin-1 destabilizes E-cadherin-based adhesions and associates with poor prognosis in basal-like breast cancer. PLoS ONE 2018, 13, e0196986. [Google Scholar] [CrossRef]
- Ling, Y.; Cao, Q.; Liu, Y.; Zhao, J.; Zhao, Y.; Li, K.; Chen, Z.; Du, X.; Huo, X.; Kang, H.; et al. Profilin 2 (PFN2) promotes the proliferation, migration, invasion and epithelial-to-mesenchymal transition of triple negative breast cancer cells. Breast Cancer 2021, 28, 368–378. [Google Scholar] [CrossRef]
- Cui, X.-B.; Zhang, S.-M.; Xu, Y.-X.; Dang, H.-W.; Liu, C.-X.; Wang, L.-H.; Yang, L.; Hu, J.-M.; Liang, W.-H.; Jiang, J.-F.; et al. PFN2, a novel marker of unfavorable prognosis, is a potential therapeutic target involved in esophageal squamous cell carcinoma. J. Transl. Med. 2016, 14, 137. [Google Scholar] [CrossRef] [PubMed]
- Meric-Bernstam, F.; Chen, H.; Akcakanat, A.; Do, K.-A.; Lluch, A.; Hennessy, B.T.; Hortobagyi, G.N.; Mills, G.B.; Gonzalez-Angulo, A.M. Aberrations in translational regulation are associated with poor prognosis in hormone receptor-positive breast cancer. Breast Cancer Res. 2012, 14, R138. [Google Scholar] [CrossRef]
- Oji, Y.; Tatsumi, N.; Fukuda, M.; Nakatsuka, S.-I.; Aoyagi, S.; Hirata, E.; Nanchi, I.; Fujiki, F.; Nakajima, H.; Yamamoto, Y.; et al. The translation elongation factor eEF2 is a novel tumor-associated antigen overexpressed in various types of cancers. Int. J. Oncol. 2014, 44, 1461–1469. [Google Scholar] [CrossRef]
- Song, Y.; Sun, B.; Hao, L.; Hu, J.; Du, S.; Zhou, X.; Zhang, L.; Liu, L.; Gong, L.; Chi, X.; et al. Elevated eukaryotic elongation factor 2 expression is involved in proliferation and invasion of lung squamous cell carcinoma. Oncotarget 2016, 7, 58470–58482. [Google Scholar] [CrossRef] [PubMed]
- Tuo, L.; Xiang, J.; Pan, X.; Hu, J.; Tang, H.; Liang, L.; Xia, J.; Hu, Y.; Zhang, W.; Huang, A.; et al. PCK1 negatively regulates cell cycle progression and hepatoma cell proliferation via the AMPK/p27Kip1 axis. J. Exp. Clin. Cancer Res. 2019, 38, 50. [Google Scholar] [CrossRef]
- Shi, L.; An, S.; Liu, Y.; Liu, J.; Wang, F. PCK1 Regulates Glycolysis and Tumor Progression in Clear Cell Renal Cell Carcinoma Through LDHA. OncoTargets Ther. 2020, 13, 2613–2627. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Gou, D.; Xiang, J.; Pan, X.; Gao, Q.; Zhou, P.; Liu, Y.; Hu, J.; Wang, K.; Tang, N. O-GlcNAc modified-TIP60/KAT5 is required for PCK1 deficiency-induced HCC metastasis. Oncogene 2021, 40, 6707–6719. [Google Scholar] [CrossRef]
- Montal, E.D.; Dewi, R.; Bhalla, K.; Ou, L.; Hwang, B.J.; Ropell, A.E.; Gordon, C.; Liu, W.-J.; DeBerardinis, R.J.; Sudderth, J.; et al. PEPCK Coordinates the Regulation of Central Carbon Metabolism to Promote Cancer Cell Growth. Mol. Cell 2015, 60, 571–583. [Google Scholar] [CrossRef]
- Hsu, H.; Chu, P.; Chang, T.; Huang, K.; Hung, W.; Jiang, S.S.; Lin, H.; Tsai, H. Mitochondrial phosphoenolpyruvate carboxykinase promotes tumor growth in estrogen receptor-positive breast cancer via regulation of the mTOR pathway. Cancer Med. 2022, 12, 1588–1601. [Google Scholar] [CrossRef]
- Wang, Z.; Dong, C. Gluconeogenesis in Cancer: Function and Regulation of PEPCK, FBPase, and G6Pase. Trends Cancer 2019, 5, 30–45. [Google Scholar] [CrossRef]
- Ma, X.; Gao, Y.; Liu, J.; Xu, L.; Liu, W.; Huna, A.; Wang, X.; Gong, W. Low expression of PCK2 in breast tumors contributes to better prognosis by inducing senescence of cancer cells. IUBMB Life 2022, 74, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Sangha, A.K.; Kantidakis, T. The Aminoacyl-tRNA Synthetase and tRNA Expression Levels Are Deregulated in Cancer and Correlate Independently with Patient Survival. Curr. Issues Mol. Biol. 2022, 44, 3001–3017. [Google Scholar] [CrossRef]
- Gao, X.; Guo, R.; Li, Y.; Kang, G.; Wu, Y.; Cheng, J.; Jia, J.; Wang, W.; Li, Z.; Wang, A.; et al. Contribution of upregulated aminoacyl-tRNA biosynthesis to metabolic dysregulation in gastric cancer. J. Gastroenterol. Hepatol. 2021, 36, 3113–3126. [Google Scholar] [CrossRef]
- Sung, Y.; Yoon, I.; Han, J.M.; Kim, S. Functional and pathologic association of aminoacyl-tRNA synthetases with cancer. Exp. Mol. Med. 2022, 54, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.H.; Kim, H.S.; Jung, S.H.; Xu, H.D.; Jeong, Y.B.; Chung, Y.J. Implication of leucyl-tRNA synthetase 1 (LARS1) over-expression in growth and migration of lung cancer cells detected by siRNA targeted knock-down analysis. Exp. Mol. Med. 2008, 40, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.; Gogonea, V.; Fox, P.L. Aminoacyl-tRNA synthetases of the multi-tRNA synthetase complex and their role in tumorigenesis. Transl. Oncol. 2022, 19, 101392. [Google Scholar] [CrossRef]
- Bottoni, A.; Vignali, C.; Piccin, D.; Tagliati, F.; Luchin, A.; Zatelli, M.C.; Uberti, E.C.D. Proteasomes and RARS modulate AIMP1/EMAP II secretion in human cancer cell lines. J. Cell. Physiol. 2007, 212, 293–297. [Google Scholar] [CrossRef]
- Zhang, J.-Y.; Shi, K.-Z.; Liao, X.-Y.; Li, S.-J.; Bao, D.; Qian, Y.; Li, D.-J. The Silence of PSMC6 Inhibits Cell Growth and Metastasis in Lung Adenocarcinoma. BioMed Res. Int. 2021, 2021, 9922185. [Google Scholar] [CrossRef]
- Kao, T.-J.; Wu, C.-C.; Phan, N.N.; Liu, Y.-H.; Ta, H.D.K.; Anuraga, G.; Wu, Y.-F.; Lee, K.-H.; Chuang, J.-Y.; Wang, C.-Y. Prognoses and genomic analyses of proteasome 26S subunit, ATPase (PSMC) family genes in clinical breast cancer. Aging 2021, 13, 17970. [Google Scholar] [CrossRef]
- Liu, Q.; Qi, Y.; Kong, X.; Wang, X.; Zhang, W.; Zhai, J.; Yang, Y.; Fang, Y.; Wang, J. Molecular and Clinical Characterization of CCT2 Expression and Prognosis via Large-Scale Transcriptome Profile of Breast Cancer. Front. Oncol. 2021, 11, 614497. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lu, Y.; Yan, X.; Lu, Q.; Sun, Y.; Wan, X.; Li, Y.; Zhao, J.; Li, Y.; Jiang, G. Current understanding on the role of CCT3 in cancer research. Front. Oncol. 2022, 12, 961733. [Google Scholar] [CrossRef] [PubMed]
- Boudiaf-Benmammar, C.; Cresteil, T.; Melki, R. The cytosolic chaperonin CCT/TRiC and cancer cell proliferation. PLoS ONE 2013, 8, e60895. [Google Scholar]
- Li, T.H.; Zhao, B.B.; Qin, C.; Wang, Y.Y.; Li, Z.R.; Cao, H.T.; Yang, X.Y.; Zhou, X.T.; Wang, W.B. IFIT1 modulates the proliferation, migration and invasion of pancreatic cancer cells via Wnt/β-catenin signaling. Cell. Oncol. 2021, 44, 1425–1437. [Google Scholar] [CrossRef]
- Danish, H.H.; Goyal, S.; Taunk, N.K.; Wu, H.; Moran, M.S.; Haffty, B.G. Interferon-induced protein with tetratricopeptide repeats 1 (IFIT1) as a prognostic marker for local control in T1-2 N0 breast cancer treated with breast-conserving surgery and radiation therapy (BCS + RT). Breast J. 2013, 19, 231–239. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Wu, M.-M.; Yen, A.-H.; Pidugu, H.B.; Chang, K.-W.; Liu, C.-J.; Lee, T.-C. IFIT1 and IFIT3 promote oral squamous cell carcinoma metastasis and contribute to the anti-tumor effect of gefitinib via enhancing p-EGFR recycling. Oncogene 2019, 38, 3232–3247. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Fan, Y.; Yu, X.; Mao, X.; Jin, F. PTBP1 promotes the growth of breast cancer cells through the PTEN/Akt pathway and autophagy. J. Cell. Physiol. 2018, 233, 8930–8939. [Google Scholar] [CrossRef]
- Li, S.; Shen, L.; Huang, L.; Lei, S.; Cai, X.; Breitzig, M.; Zhang, B.; Yang, A.; Ji, W.; Huang, M.; et al. PTBP1 enhances exon11a skipping in Mena pre-mRNA to promote migration and invasion in lung carcinoma cells. Biochim. Biophys. Acta BBA-Gene Regul. Mech. 2019, 1862, 858–869. [Google Scholar]
- Zhu, W.; Zhou, B.-L.; Rong, L.-J.; Ye, L.; Xu, H.-J.; Zhou, Y.; Yan, X.-J.; Liu, W.-D.; Zhu, B.; Wang, L.; et al. Roles of PTBP1 in alternative splicing, glycolysis, and oncogensis. J. Zhejiang Univ. Sci. B 2020, 21, 122–136. [Google Scholar] [PubMed]
- Jiang, Y.; Wang, B.; Li, Y.; Shen, J.; Wei, Y.; Li, H.; Chen, S.; Yang, H.; Zeng, F.; Liu, C.; et al. DDX19A Promotes Metastasis of Cervical Squamous Cell Carcinoma by Inducing NOX1-Mediated ROS Production. Front. Oncol. 2021, 11, 6299744. [Google Scholar] [CrossRef]
- Cai, W.; Xiong Chen, Z.; Rane, G.; Satendra Singh, S.; Choo, Z.E.; Wang, C.; Yuan, Y.; Zea Tan, T.; Arfuso, F.; Yap, C.T.; et al. Wanted DEAD/H or Alive: Helicases Winding Up in Cancers. JNCI J. Natl. Cancer Inst. 2017, 109, djw278. [Google Scholar]
- Ma, Y.-T.; Xing, X.-F.; Dong, B.; Cheng, X.-J.; Guo, T.; Du, H.; Wen, X.-Z.; Ji, J.-F. Higher autocrine motility factor/glucose-6-phosphate isomerase expression is associated with tumorigenesis and poorer prognosis in gastric cancer. Cancer Manag. Res. 2018, 10, 4969–4980. [Google Scholar] [CrossRef]
- Zeng, J.; Yi, J.; Tan, S.; Zeng, Y.; Zou, L.; Zhang, C.; Liu, L.; Yi, P.; Fan, P.; Yu, J. GPI: An indicator for immune infiltrates and prognosis of human breast cancer from a comprehensive analysis. Front. Endocrinol. 2022, 13, 995972. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Deng, X.; Sun, R.; Luo, M.; Liang, M.; Gu, B.; Zhang, T.; Peng, Z.; Lu, Y.; Tian, C.; et al. GPI Is a Prognostic Biomarker and Correlates With Immune Infiltrates in Lung Adenocarcinoma. Front. Oncol. 2021, 11, 752642. [Google Scholar]
- Gallardo-Pérez, J.C.; Rivero-Segura, N.A.; Marín-Hernández, A.; Moreno-Sánchez, R.; Rodríguez-Enríquez, S. GPI/AMF inhibition blocks the development of the metastatic phenotype of mature multi-cellular tumor spheroids. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2014, 1843, 1043–1053. [Google Scholar]
- Nami, B.; Wang, Z. Genetics and Expression Profile of the Tubulin Gene Superfamily in Breast Cancer Subtypes and Its Relation to Taxane Resistance. Cancers 2018, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jiao, Z.; Ji, Y.; Zhang, S. Elevated TUBA1A Might Indicate the Clinical Outcomes of Patients with Gastric Cancer, Being Associated with the Infiltration of Macrophages in the Tumor Immune Microenvironment. J. Gastrointest. Liver Dis. 2020, 29, 509–522. [Google Scholar] [CrossRef]
- Liang, L.; Sun, H.; Zhang, W.; Zhang, M.; Yang, X.; Kuang, R.; Zheng, H. Meta-Analysis of EMT Datasets Reveals Different Types of EMT. PLoS ONE 2016, 11, e0156839. [Google Scholar]
- Hua, C.; Chen, J.; Li, S.; Zhou, J.; Fu, J.; Sun, W.; Wang, W. KDM6 Demethylases and Their Roles in Human Cancers. Front. Oncol. 2021, 11, 779918. [Google Scholar]
- Sterling, J.; Menezes, S.V.; Abbassi, R.H.; Munoz, L. Histone lysine demethylases and their functions in cancer. Int. J. Cancer 2020, 148, 2375–2388. [Google Scholar]
- Takahashi, S.; Kato, K.; Nakamura, K.; Nakano, R.; Kubota, K.; Hamada, H. Neural cell adhesion molecule 2 as a target molecule for prostate and breast cancer gene therapy. Cancer Sci. 2011, 102, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, Y.; Wang, C.; Ju, H.; Liu, W.; Zhang, X.; Miao, S.; Wang, L.; Sun, Q.; Song, W. Rhomboid domain-containing protein 1 promotes breast cancer progression by regulating the p-Akt and CDK2 levels. Cell Commun. Signal. 2018, 16, 65. [Google Scholar] [PubMed]
- Ren, X.; Song, W.; Liu, W.; Guan, X.; Miao, F.; Miao, S.; Wang, L. Rhomboid domain containing 1 inhibits cell apoptosis by upregulating AP-1 activity and its downstream target Bcl-3. FEBS Lett. 2013, 587, 1793–1798. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Deng, Y.; Ye, J.; Zhuo, Y.; Liu, Z.; Liang, Y.; Zhang, H.; Zhu, X.; Luo, Y.; Feng, Y.; et al. Aberrant Expression of Citrate Synthase is Linked to Disease Progression and Clinical Outcome in Prostate Cancer. Cancer Manag. Res. 2020, 12, 6149–6163. [Google Scholar] [PubMed]
- Schlichtholz, B.; Turyn, J.; Goyke, E.; Biernacki, M.; Jaskiewicz, K.; Sledzinski, Z.; Swierczynski, J. Enhanced Citrate Synthase Activity in Human Pancreatic Cancer. Pancreas 2005, 30, 99–104. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.; Zhou, J.; Wang, Y.; Wang, X.; Di, W.; Zhang, S. Citrate synthase expression affects tumor phenotype and drug resistance in human ovarian carcinoma. PLoS ONE 2014, 9, e115708. [Google Scholar]
- Lin, C.-C.; Cheng, T.-L.; Tsai, W.-H.; Tsai, H.-J.; Hu, K.-H.; Chang, H.-C.; Yeh, C.-W.; Chen, Y.-C.; Liao, C.-C.; Chang, W.-T. Loss of the respiratory enzyme citrate synthase directly links the Warburg effect to tumor malignancy. Sci. Rep. 2012, 2, 785. [Google Scholar]
- MacPherson, S.; Horkoff, M.; Gravel, C.; Hoffmann, T.; Zuber, J.; Lum, J.J. STAT3 Regulation of Citrate Synthase Is Essential during the Initiation of Lymphocyte Cell Growth. Cell Rep. 2017, 19, 910–918. [Google Scholar] [CrossRef]
- Gong, Y.; Lv, Y.; Xu, F.; Xiu, Y.; Lu, Y.; Liu, Z.; Deng, L. LAMTOR3 is a prognostic biomarker in kidney renal clear cell carcinoma. J. Clin. Lab. Anal. 2022, 36, e24648. [Google Scholar]
- Marina, M.; Wang, L.; Conrad, S.E. The scaffold protein MEK Partner 1 is required for the survival of estrogen receptor positive breast cancer cells. Cell Commun. Signal. 2012, 10, 18. [Google Scholar] [CrossRef]
- De Araujo, M.E.; Erhart, G.; Buck, K.; Müller-Holzner, E.; Hubalek, M.; Fiegl, H.; Campa, D.; Canzian, F.; Eilber, U.; Chang-Claude, J.; et al. Polymorphisms in the gene regions of the adaptor complex LAMTOR2/LAMTOR3 and their association with breast cancer risk. PLoS ONE 2013, 8, e53768. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.S.; Hong, S.K.; Kwon, S.J.; Go, Y.H.; Oh, E.; Cha, H.J. BCL2 induced by LAMTOR3/MAPK is a druggable target of chemoradioresistance in mesenchymal lung cancer. Cancer Lett. 2017, 403, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.P.; Qian, K.; Lee, J.S.; Zhou, J.; Han, X.; Zhang, B.; Ong, Q.; Ni, W.; Jiang, M.; Ruan, H.B.; et al. O-GlcNAcase targets pyruvate kinase M2 to regulate tumor growth. Oncogene 2020, 39, 560–573. [Google Scholar] [CrossRef] [PubMed]
- Olechnowicz, A.; Oleksiewicz, U.; Machnik, M. KRAB-ZFPs and cancer stem cells identity. Genes Dis. 2022, 10, 1820–1832. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Wang, L.; Zhang, H.; Liang, C.; Li, B. Identification of a 5-Gene Signature Predicting Progression and Prognosis of Clear Cell Renal Cell Carcinoma. Med. Sci. Monit. 2019, 25, 4401–4413. [Google Scholar] [CrossRef]
- Oleksiewicz, U.; Gładych, M.; Raman, A.T.; Heyn, H.; Mereu, E.; Chlebanowska, P.; Andrzejewska, A.; Sozańska, B.; Samant, N.; Fąk, K.; et al. TRIM28 and Interacting KRAB-ZNFs Control Self-Renewal of Human Pluripotent Stem Cells through Epigenetic Repression of Pro-differentiation Genes. Stem Cell Rep. 2017, 9, 2065–2080. [Google Scholar] [CrossRef]
- Sun, S.; Liu, Y.; Zhou, M.; Wen, J.; Xue, L.; Han, S.; Liang, J.; Wang, Y.; Wei, Y.; Yu, J.; et al. PA2G4 promotes the metastasis of hepatocellular carcinoma by stabilizing FYN mRNA in a YTHDF2-dependent manner. Cell Biosci. 2022, 12, 55. [Google Scholar] [CrossRef]
- Novita Sari, I.; Setiawan, T.; Seock Kim, K.; Toni Wijaya, Y.; Won Cho, K.; Young Kwon, H. Metabolism and function of polyamines in cancer progression. Cancer Lett. 2021, 519, 91–104. [Google Scholar] [CrossRef]
- Akinyele, O.; Wallace, H.M. Characterising the Response of Human Breast Cancer Cells to Polyamine Modulation. Biomolecules 2021, 11, 743. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Krasnov, G.S.; Lipatova, A.V.; Sadritdinova, A.F.; Kardymon, O.L.; Fedorova, M.S.; Melnikova, N.V.; Stepanov, O.A.; Zaretsky, A.R.; Kaprin, A.D. The Dysregulation of Polyamine Metabolism in Colorectal Cancer Is Associated with Overexpression of c-Myc and C/EBPβ rather than Enterotoxigenic Bacteroides fragilis Infection. Oxidative Med. Cell. Longev. 2016, 2016, 2353560. [Google Scholar] [CrossRef]
- Gamble, L.D.; Hogarty, M.D.; Liu, X.; Ziegler, D.S.; Marshall, G.; Norris, M.D.; Haber, M. Polyamine pathway inhibition as a novel therapeutic approach to treating neuroblastoma. Front. Oncol. 2012, 2, 162. [Google Scholar] [CrossRef] [PubMed]
- Checa-Rojas, A.; Delgadillo-Silva, L.F.; Velasco-Herrera, M.d.C.; Andrade-Domínguez, A.; Gil, J.; Santillán, O.; Lozano, L.; Toledo-Leyva, A.; Ramírez-Torres, A.; Talamas-Rohana, P.; et al. GSTM3 and GSTP1: Novel players driving tumor progression in cervical cancer. Oncotarget 2018, 9, 21696–21714. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, J.; You, L.; Dai, M.; Zhao, Y. GSTM3 Function and Polymorphism in Cancer: Emerging but Promising. Cancer Manag. Res. 2020, 12, 10377–10388. [Google Scholar] [CrossRef] [PubMed]
- Di Lollo, V.; Canciello, A.; Orsini, M.; Bernabò, N.; Ancora, M.; Federico, M.; Curini, V.; Mattioli, M.; Russo, V.; Mauro, A.; et al. Transcriptomic and computational analysis identified LPA metabolism, KLHL14 and KCNE3 as novel regulators of Epithelial-Mesenchymal Transition. Sci. Rep. 2020, 10, 4180. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; He, L.; Shay, C.; Lang, L.; Loveless, J.; Yu, J.; Chemmalakuzhy, R.; Jiang, H.; Liu, M.; Teng, Y. Nck-associated protein 1 associates with HSP90 to drive metastasis in human non-small-cell lung cancer. J. Exp. Clin. Cancer Res. 2019, 38, 122. [Google Scholar] [CrossRef]
- Teng, Y.; Qin, H.; Bahassan, A.; Bendzunas, N.G.; Kennedy, E.J.; Cowell, J.K. The WASF3-NCKAP1-CYFIP1 Complex Is Essential for Breast Cancer Metastasis. Cancer Res. 2016, 76, 5133–5142. [Google Scholar] [CrossRef]
- Chen, J.; Ge, J.; Zhang, W.; Xie, X.; Zhong, X.; Tang, S. NCKAP1 is a Prognostic Biomarker for Inhibition of Cell Growth in Clear Cell Renal Cell Carcinoma. Front. Genet. 2022, 13, 764957. [Google Scholar] [CrossRef]
- Zhong, X.P.; Kan, A.; Ling, Y.H.; Lu, L.H.; Mei, J.; Wei, W.; Li, S.H.; Guo, R.P. NCKAP1 improves patient outcome and inhibits cell growth by enhancing Rb1/p53 activation in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 369. [Google Scholar] [CrossRef]
- Nicolussi, A.; D’Inzeo, S.; Capalbo, C.; Giannini, G.; Coppa, A. The role of peroxiredoxins in cancer. Mol. Clin. Oncol. 2017, 6, 139–153. [Google Scholar] [CrossRef]
- Park, M.H.; Jo, M.; Kim, Y.R.; Lee, C.-K.; Hong, J.T. Roles of peroxiredoxins in cancer, neurodegenerative diseases and inflammatory diseases. Pharmacol. Ther. 2016, 163, 1–23. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Gao, X.; Ma, Q.; Yu, Z.; Huang, S. Rab8A promotes breast cancer progression by increasing surface expression of Tropomyosin-related kinase B. Cancer Lett. 2022, 535, 215629. [Google Scholar] [CrossRef] [PubMed]
- Aboubakr, H.; Lavanya, S.; Thirupathi, M.; Rohini, R.; Sarita, R. Human Rab8b Protein as a Cancer Target—An In Silico Study. J. Comput. Sci. Syst. Biol. 2016, 9, 132–149. [Google Scholar]
- Demir, K.; Kirsch, N.; Beretta, C.A.; Erdmann, G.; Ingelfinger, D.; Moro, E.; Argenton, F.; Carl, M.; Niehrs, C.; Boutros, M. RAB8B Is Required for Activity and Caveolar Endocytosis of LRP6. Cell Rep. 2013, 4, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, H.-T.; Wang, Y.-C. Rab-mediated vesicle trafficking in cancer. J. Biomed. Sci. 2016, 23, 70. [Google Scholar] [CrossRef]
- Jia, N.; Song, Z.; Chen, B.; Cheng, J.; Zhou, W. A Novel Circular RNA circCSPP1 Promotes Liver Cancer Progression by Sponging miR-1182. OncoTargets Ther. 2021, 14, 2829–2838. [Google Scholar] [CrossRef]
- Nishimura, N.; Pham, T.V.H.; Hartomo, T.B.; Lee, M.J.; Hasegawa, D.; Takeda, H.; Kawasaki, K.; Kosaka, Y.; Yamamoto, T.; Morikawa, S.; et al. Rab15 expression correlates with retinoic acid-induced differentiation of neuroblastoma cells. Oncol. Rep. 2011, 26, 145–151. [Google Scholar] [CrossRef][Green Version]
- Villagomez, F.R.; Medina-Contreras, O.; Cerna-Cortes, J.F.; Patino-Lopez, G. The role of the oncogenic Rab35 in cancer invasion, metastasis, and immune evasion, especially in leukemia. Small GTPases 2020, 11, 334–345. [Google Scholar] [CrossRef]
- Dharmapal, D.; Jyothy, A.; Mohan, A.; Balagopal, P.G.; George, N.A.; Sebastian, P.; Maliekal, T.T.; Sengupta, S. β-Tubulin Isotype, TUBB4B, Regulates The Maintenance of Cancer Stem Cells. Front. Oncol. 2021, 11, 788024. [Google Scholar] [CrossRef]
- Sobierajska, K.; Ciszewski, W.M.; Wawro, M.E.; Wieczorek-Szukała, K.; Boncela, J.; Papiewska-Pajak, I.; Niewiarowska, J.; Kowalska, M.A. TUBB4B Downregulation Is Critical for Increasing Migration of Metastatic Colon Cancer Cells. Cells 2019, 8, 810. [Google Scholar] [CrossRef]
- Li, P.; Miao, C.; Liang, C.; Shao, P.; Wang, Z.; Li, J. Silencing CAPN2 Expression Inhibited Castration-Resistant Prostate Cancer Cells Proliferation and Invasion via AKT/mTOR Signal Pathway. BioMed Res. Int. 2017, 2017, 2593674. [Google Scholar] [CrossRef]
- Miao, C.; Liang, C.; Tian, Y.; Xu, A.; Zhu, J.; Zhao, K.; Zhang, J.; Hua, Y.; Liu, S.; Dong, H.; et al. Overexpression of CAPN2 promotes cell metastasis and proliferation via AKT/mTOR signaling in renal cell carcinoma. Oncotarget 2017, 8, 97811–97821. [Google Scholar] [CrossRef]
- Peng, X.; Yang, R.; Song, J.; Wang, X.; Dong, W. Calpain2 Upregulation Regulates EMT-Mediated Pancreatic Cancer Metastasis via the Wnt/β-Catenin Signaling Pathway. Front. Med. 2022, 9, 783592. [Google Scholar] [CrossRef]
- Liu, C.; Haijuan, F.; Liu, X.; Lei, Q.; Zhang, Y.; She, X.; Liu, Q.; Liu, Q.; Sun, Y.; Li, G.; et al. LINC00470 Coordinates the Epigenetic Regulation of ELFN2 to Distract GBM Cells Autophagy. Mol. Ther. 2018, 26, 2267–2281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Zhang, J.J.; Song, H.J.; Li, D.W. Overexpression of CST4 promotes gastric cancer aggressiveness by activating the ELFN2 signaling pathway. Am. J. Cancer Res. 2017, 7, 2290–2304. [Google Scholar] [PubMed]
- Wang, B.; Yang, Y.; Friedman, P.A. Na/H Exchange Regulatory Factor 1, a Novel AKT-associating Protein, Regulates Extracellular Signal-regulated Kinase Signaling through a B-Raf–Mediated Pathway. Mol. Biol. Cell 2008, 19, 1637–1645. [Google Scholar] [CrossRef]
- Crocamo, S.; Binato, R.; Santos, E.; de Paula, B.; Abdelhay, E. Translational Results of Zo-NAnTax: A Phase II Trial of Neoadjuvant Zoledronic Acid in HER2-Positive Breast Cancer. Int. J. Mol. Sci. 2022, 23, 15515. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Jiao, Y.; Hao, Y.; Yan, S.; Lyu, N.; Gao, H.; Li, D.; Liu, Q.; Zheng, J.; Song, N. Targeting of NHERF1 through RNA interference inhibits the proliferation and migration of metastatic prostate cancer cells. Oncol. Lett. 2016, 11, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Zhu, H.; Sun, H.; Hua, Y.; Zhang, G.; Jiang, J.; Wang, X. Adipose Mesenchymal Stem Cell-Derived Exosomal microRNA-1236 Reduces Resistance of Breast Cancer Cells to Cisplatin by Suppressing SLC9A1 and the Wnt/β-Catenin Signaling. Cancer Manag. Res. 2020, 12, 8733–8744. [Google Scholar] [CrossRef]
- Takahashi, Y.; Morales, F.C.; Kreimann, E.L.; Georgescu, M.-M. PTEN tumor suppressor associates with NHERF proteins to attenuate PDGF receptor signaling. EMBO J. 2006, 25, 910–920. [Google Scholar] [CrossRef]
- Choi, C.H.; Chung, J.-Y.; Chung, E.J.; Sears, J.D.; Lee, J.-W.; Bae, D.-S.; Hewitt, S.M. Prognostic significance of annexin A2 and annexin A4 expression in patients with cervical cancer. BMC Cancer 2016, 16, 448. [Google Scholar] [CrossRef]
- Yao, H.; Sun, C.; Hu, Z.; Wang, W. The role of annexin A4 in cancer. Front. Biosci. Landmark Ed. 2016, 21, 949–957. [Google Scholar] [PubMed]
- Lin, L.-L.; Huang, H.-C.; Juan, H.-F. Revealing the Molecular Mechanism of Gastric Cancer Marker Annexin A4 in Cancer Cell Proliferation Using Exon Arrays. PLoS ONE 2012, 7, e44615. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Guo, C.; Liu, S.; Sun, M.Z. Annexin A4 and cancer. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 447, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Lee, Y.C.; Wang, Y.Y.; Wang, C.H.; Hou, M.F.; Yuan, S.S.F. YWHAE promotes proliferation, metastasis, and chemoresistance in breast cancer cells. Kaohsiung J. Med. Sci. 2019, 35, 408–416. [Google Scholar] [CrossRef]
- Gan, Y.; Ye, F.; He, X.-X. The role of YWHAZ in cancer: A maze of opportunities and challenges. J. Cancer 2020, 11, 2252–2264. [Google Scholar] [CrossRef]
- Yu, C.C.; Li, C.F.; Chen, I.H.; Lai, M.T.; Lin, Z.J.; Korla, P.K.; Chai, C.Y.; Ko, G.; Chen, C.M.; Hwang, T. YWHAZ amplification/overexpression defines aggressive bladder cancer and contributes to chemo-/radio-resistance by suppressing caspase-mediated apoptosis. J. Pathol. 2019, 248, 476–487. [Google Scholar] [CrossRef]
- Liu, J.; Yang, X.; Ji, Q.; Yang, L.; Li, J.; Long, X.; Ye, M.; Huang, K.; Zhu, X. Immune Characteristics and Prognosis Analysis of the Proteasome 20S Subunit Beta 9 in Lower-Grade Gliomas. Front. Oncol. 2022, 12, 875131. [Google Scholar] [CrossRef]
- Hayashi, T.; Horiuchi, A.; Sano, K.; Hiraoka, N.; Kasai, M.; Ichimura, T.; Sudo, T.; Tagawa, Y.-I.; Nishimura, R.; Ishiko, O.; et al. Potential role of LMP2 as tumor-suppressor defines new targets for uterine leiomyosarcoma therapy. Sci. Rep. 2011, 1, 180. [Google Scholar] [CrossRef]
- Tian, T.; Li, X.; Zhang, J. mTOR Signaling in Cancer and mTOR Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Rascio, F.; Spadaccino, F.; Rocchetti, M.T.; Castellano, G.; Stallone, G.; Netti, G.S.; Ranieri, E. The Pathogenic Role of PI3K/AKT Pathway in Cancer Onset and Drug Resistance: An Updated Review. Cancers 2021, 13, 3949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X. Targeting the Wnt/β-catenin signaling pathway in cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-signaling pathway in cancer. OncoTargets Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [PubMed]
- Madden, E.; Logue, S.E.; Healy, S.J.; Manie, S.; Samali, A. The role of the unfolded protein response in cancer progression: From oncogenesis to chemoresistance. Biol. Cell 2019, 111, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Alrefaei, A.F.; Abu-Elmagd, M. LRP6 Receptor Plays Essential Functions in Development and Human Diseases. Genes 2022, 13, 120. [Google Scholar] [CrossRef]
- Gomes, A.P.; Ilter, D.; Low, V.; Drapela, S.; Schild, T.; Mullarky, E.; Han, J.; Elia, I.; Broekaert, D.; Rosenzweig, A.; et al. Altered propionate metabolism contributes to tumour progression and aggressiveness. Nat. Metab. 2022, 4, 435–443. [Google Scholar] [CrossRef]
- Grasmann, G.; Smolle, E.; Olschewski, H.; Leithner, K. Gluconeogenesis in cancer cells—Repurposing of a starvation-induced metabolic pathway? Biochim. Biophys. Acta BBA-Rev. Cancer 2019, 1872, 24–36. [Google Scholar] [CrossRef]
- Li, X.; Han, M.; Zhang, H.; Liu, F.; Pan, Y.; Zhu, J.; Liao, Z.; Chen, X.; Zhang, B. Structures and biological functions of zinc finger proteins and their roles in hepatocellular carcinoma. Biomark. Res. 2022, 10, 2. [Google Scholar] [CrossRef]
- Ma, R.; Ji, T.; Zhang, H.; Dong, W.; Chen, X.; Xu, P.; Chen, D.; Liang, X.; Yin, X.; Liu, Y.; et al. A Pck1-directed glycogen metabolic program regulates formation and maintenance of memory CD8+ T cells. Nat. Cell Biol. 2018, 20, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, L.; Sandhu, J.K.; Harper, M.-E.; Cuperlovic-Culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef]
- Halama, A.; Suhre, K. Advancing Cancer Treatment by Targeting Glutamine Metabolism—A Roadmap. Cancers 2022, 14, 553. [Google Scholar] [CrossRef]
- Altman, B.J.; Stine, Z.E.; Dang, C.V. From Krebs to clinic: Glutamine metabolism to cancer therapy. Nat. Rev. Cancer 2016, 16, 619–634. [Google Scholar] [PubMed]
- Liu, M.-X.; Jin, L.; Sun, S.-J.; Liu, P.; Feng, X.; Cheng, Z.-L.; Liu, W.-R.; Guan, K.-L.; Shi, Y.-H.; Yuan, H.-X.; et al. Metabolic reprogramming by PCK1 promotes TCA cataplerosis, oxidative stress and apoptosis in liver cancer cells and suppresses hepatocellular carcinoma. Oncogene 2018, 37, 1637–1653. [Google Scholar] [CrossRef]
- Millward, C.A.; DeSantis, D.; Hsieh, C.-W.; Heaney, J.D.; Pisano, S.; Olswang, Y.; Reshef, L.; Beidelschies, M.; Puchowicz, M.; Croniger, C.M. Phosphoenolpyruvate carboxykinase (Pck1) helps regulate the triglyceride/fatty acid cycle and development of insulin resistance in mice. J. Lipid Res. 2010, 51, 1452–1463. [Google Scholar] [CrossRef]
- Leithner, K.; Triebl, A.; Trötzmüller, M.; Hinteregger, B.; Leko, P.; Wieser, B.I.; Grasmann, G.; Bertsch, A.L.; Züllig, T.; Stacher, E.; et al. The glycerol backbone of phospholipids derives from noncarbohydrate precursors in starved lung cancer cells. Proc. Natl. Acad. Sci. USA 2018, 115, 6225–6230. [Google Scholar] [CrossRef] [PubMed]
- Sadria, M.; Layton, A.T. Interactions among mTORC, AMPK and SIRT: A computational model for cell energy balance and metabolism. Cell Commun. Signal. 2021, 19, 57. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Peng, D.; Cai, Z.; Lin, H.-K. AMPK signaling and its targeting in cancer progression and treatment. Semin. Cancer Biol. 2022, 85, 52–68. [Google Scholar]
- Penugurti, V.; Mishra, Y.G.; Manavathi, B. AMPK: An odyssey of a metabolic regulator, a tumor suppressor, and now a contextual oncogene. Biochim. Biophys. Acta BBA-Rev. Cancer 2022, 1877, 188785. [Google Scholar] [CrossRef]
- Furukawa, F.; Tseng, Y.-C.; Liu, S.-T.; Chou, Y.-L.; Lin, C.-C.; Sung, P.-H.; Uchida, K.; Lin, L.-Y.; Hwang, P.-P. Induction of Phosphoenolpyruvate Carboxykinase (PEPCK) during Acute Acidosis and Its Role in Acid Secretion by V-ATPase-Expressing Ionocytes. Int. J. Biol. Sci. 2015, 11, 712–725. [Google Scholar] [CrossRef]
- Yu, S.; Meng, S.; Xiang, M.; Ma, H. Phosphoenolpyruvate carboxykinase in cell metabolism: Roles and mechanisms beyond gluconeogenesis. Mol. Metab. 2021, 53, 101257. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathways | NES | FDR q-Val | |
|---|---|---|---|
| Upregulated | INTERFERON_ALPHA_RESPONSE (IFN-α) | 1.52 | 0.223 |
| INTERFERON_GAMMA_RESPONSE (IFN-γ) | 1.47 | 0.165 | |
| MYC_TARGETS_V1 | 1.04 | 0.971 | |
| UNFOLDED_PROTEIN_RESPONSE | 1 | 0.831 | |
| Downregulated | ESTROGEN_RESPONSE_LATE | −1.17 | 1 |
| ESTROGEN_RESPONSE_EARLY | −1.07 | 0.948 |
| Gene Name | Gene Description | Role | Expression in Malignancies and Putative Neoplastic Effects | Pathways | Neoplastic Condition | |
|---|---|---|---|---|---|---|
| Proteins enriched in phenotype of low JTB expression condition | ||||||
| HSPB1/HSP27 | Heat shock 27 kDa protein 1 | protein folding/CMPF, cell adhesion, cell migration, cytoskeleton dependent intracellular transport, cell death in response to OS, programmed cell death | overexpressed in many cancers [17]: including BC [18] | PT | APOPTOSIS, regulates EMT process and NF-κB activity [18] | poor clinical outcome, cell invasion, metastasis, resistance to apoptosis [17]; participates in maintenance of BCSCs [18] |
| HSPA4/HSP70/Apg-2 | Heat shock 70 kDa protein family A (HSP70) member 4 isoform a variant | molecular chaperone, induced by oncogenic stress, autophagy, regulation of protein ubiquitination, cadherin binding, cell adhesion molecule binding, MHC class II protein complex binding [19] | overexpressed in many cancers, including HCC [19], CRC [20], HNSC [21], involved in progression of BC [22], GC [23] | PT | MTORC1_SIGNALING; associated with CSCs proprieties via chaperones for EMT-associated proteins, inducing migration [23]; silencing reduced activation of PI3K/Akt signaling and increase in apoptosis [20]; activates mTOR pathway [21] | facilitates cancer cell survival, inhibits apoptosis, promotes proliferation, immune regulation [19], accumulation of misfolded proteins, ROS, and DNA damage [21], considered as a necroptosis-related gene in BC that activates cell cycle [22] |
| MCM6 | Minichromosome maintenance complex component 6 | significant DNA replication regulator, plays a key role in cell cycle progression [24] | overexpressed in many cancers [25]: BC, CRC, HCC [25], glioma, endometrial adenocarcinoma, cervical cancer, Merkel cell carcinoma, lung cancer [24] | PT | promotes EMT and activates MEK/ERK signaling, sustaining carcinogenesis [24,25] | sensitive, specific biomarker for GSTM cancer, involved in cell proliferation, metastasis, migration, invasion, immune response [24,25] |
| POTEF/POTEACTIN/ACTB | POTE ankyrin domain family member F/Beta-actin | cytoskeleton protein that belongs to the actin family, involved in motility, polarity, chemotaxis andimmune cell infiltration [26] | expressed in many cancers: BC cell lines, overexpressed in CRPC [27] | PT | AJ; represses the AT effect of Toll-like receptor (TLR) signaling pathway [27,28]; involved in cell migration by NF-κB and Wnt/β-catenin pathway [26] | promotes cell growth [27], migration, invasion [26] |
| ACTN1 | Alpha-actinin-1 (isoform X5) | actin cross-linking protein involved in cytokinesis, cell adhesion and migration [29] | overexpressed in BC tissue, BL-BC cell lines [30], GC [29] | PT | AJ; EMT via AKT/GSK3β/β-catenin pathway and FAK/Src/JAK2/STAT3 signaling [29] | promotes cell proliferation, invasion, migration, and inhibits apoptosis [29]; loss of polarity by cytoskeleton reorganization and E-cadherin-based adhesion, lack of ER expression and poor survival in BL-BC [30] |
| PFN2 | Profilin-2 (isoform b) | actin binding protein involved in cytoskeleton organization, vesicle mediated transport, signaling, cell junction organization, cell motility | overexpressed in TNBC [31], ESCC [32] | PT | EMT [31] | promotes cell proliferation, migration and invasion of TNBC cells [31] |
| EEF2 | Eukaryotic elongation factor 2 | translation elongation factor involved in polypeptide chain elongation step, cell cycle progression | highly expressed in various malignant tumors: BC [33], GC, CRC, esophageal, pancreatic, PCa, HNSCC, GBM [34], LSCC tissues and cell lines [35] | PT | UPR; promotes G2/M progression in cell cycle activating Akt signaling [34], CDC2/Cyclin B1 and EMT-related proteins [35] | associated with node positivity [33]; plays an oncogenic role, promotes cancer cell growth [34], migration, invasion [35] |
| 1KHB/PCK1/PEPCK-C (cytoplasmic isoenzyme) | Phosphoenolpyruvate carboxykinase 1 (PEPCK) Complex With Nonhydrolyzable GTP Analog, Mad Data (chain A) | rate-limiting gluconeogenesis enzyme | oncogene overexpressed in colon cancer and melanoma [36]; downregulated and tumor suppressor in gluconeogenic tissues (liver and kidney): HCC [36] and ccRCC [37] | PT | GLYCOLYSIS; acts via AMPK/p27Kip1 axis [36]; depletion promotes EMT in HCC [38] | promotes cell proliferation via mTORC1 (oncogenic function) [39]; AT in kidney and liver: suppressed ccRCC cell growth and metastasis, inhibited tumorigenesis by blocking aerobic glycolysis pathway [37]; suppresses liver tumor growth, cell cycle progression and proliferation [36] |
| PCK2/PEPCK-M (mitochondrial isoenzyme) | Phosphoenolpyruvate carboxykinase | rate-limiting gluconeogenesis enzyme | overexpressed many cancers, including ER+ BC [40], lung, prostate, thyroid, bladder, BC, cervical cancer [41] | PT | REACTOME_GLUCONEOGENESIS; activation of mTORC1 and E2F1 pathways [40]; silencing contributes to cellular senescence, inhibiting EMT in BC cells [42] | promotes tumor growth, proliferation and cell cycle progression [40] |
| FARSB/HSPC173 | Phenylalanyl-tRNA ligase beta subunit | cytoplasmic aminoacyl-tRNA synthetase (ARS/AARS) involved in tRNA metabolic process, amino acid metabolic process, protein-containing complex assembly | overexpressed in in tumor samples compared to adjacent normal tissues [43]: GC [44] | PT | aminoacyl-tRNA synthesis pathway [44] | promotes cancer progression, poor prognosis, metastasis [44]; worse patient survival in BC [43] |
| GARS1 | Glycyl-tRNA synthetase 1 | cytoplasmic and mitochondrial ARS | overexpressed in tumor samples compared to adjacent normal tissues [43], displays androgen-dependent transcriptional initiation in several hormone-responsive cells, overexpressed in PCa [45] | PT | may deactivate ERK signaling pathway [45] | worse patient survival in BC [43]; could induce tumor regression [45] |
| LARS1 | Leucyl-tRNA synthetase 1 (Editing Domain) | cytoplasmic ARS | overexpressed in some cancers: myeloid leukemia, pancreatic cancer, renal, cervical, skin cancer [45], lung cancer cell lines and tissues [46] | PT | senses intracellular leucine levels to activate mTORC1 pathway [45] | lower patient survival in BC [43], promotes cell proliferation, growth [45], and migration [46] |
| RARS1 | Arginyl--tRNA synthetase 1 | cytoplasmic ARS involved in protein synthesis | important tumorigenic activity, overexpressed in hepatoma cells [47], associated with an increased risk of BC [45] | PT | impairment of AIMP1/EMAPII secretion in MCF7 cells [48] | lower patient survival in BC [43], increases growth rate in hepatoma cells, induces stem cell-like features in head and neck tumors [47] |
| PSMC6 | Proteasome subunit p42/proteasome 26S subunit ATPase 6 | protein catabolic process, regulation of DNA-template transcription, protein-containing complex assembly | highly upregulated in LUAD [49], BC tissues, especially in luminal cancer [50] | PT | MTORC1_SIGNALING; REACTOME_REGULATION_OF_MITOTIC_CELL_CYCLE; activation of Wnt signaling via degrading AXIN proteins [49] | oncogenic effect [50], poor prognosis, silencing inhibits cell growth, migration and invasion [49] |
| CCT2/CCTβ | Chaperonin containing TCP1 Subunit 2 | involved in cell cycle regulation, protein folding and binding biological processes [51] | overexpressed in various tumors and cell lines, such as HER2+ BC, liver, prostate, cholecyst, lung, CRC, BC [51], glioblastoma [52] | PT | P53 signaling [51] | worse prognosis, especially in luminal A subtype, promotes cell growth/survival, invasion and proliferation [51] |
| CCT3/TRiC | Cytosolic chaperonin containing t-complex polypeptide 1 (TCP1) subunit 3/TCP1 ring complex (hTRiC5), partial | molecular chaperone involved in proteostasis, folding of tubulin and actins and many proteins involved in cancer [53], cell division, proliferation andapoptosis [52] | overexpressed in some tumors: BC, HCC, LUAD and LUSC, NSCLC, cervical and CRC, AML, multiple myeloma, papillary thyroid carcinoma, melanoma, GC [52] | PT | might regulate IGF-1 signaling; actin cytoskeletal signaling, and PTEN signaling, Wnt/β-catenin, JAK2/STAT3, PI3K/Akt [52] | oncogene, promotes cell growth, survival, proliferation, cell cycle progression and anti-apoptosis [52] |
| IFIT1 | Interferon-induced protein with tetratricopeptide repeats 1 (isoform 2) | inflammation-related protein, RNA-binding protein modulated by JAK/STAT pathway [54], involved in regulation of translation [55] | overexpressed in PDAC [54], OSCC [56] | PT | INTERFERON_GAMM_RESPONSE; UPR; EMT [56], Wnt/β-catenin activation [54], increasing levels of p-EGFR and p-Akt [56] | increases cell proliferation, migration, invasion [54], tumor growth, regional and distant metastasis [56] |
| PTBP1/hnRNP1 | Polypyrimidine tract-binding protein 1 (isoform a) | RNA-binding protein, key factor in the control of RNA metabolism, regulates mRNA alternative splicing (AS) events, mRNA stability, mRNA localization [57] | overexpressed in human epithelial ovarian tumors, BC tissues and cell lines, glioblastomas [57], LUAD tissues and cell lines [58] | PT | regulates PTEN-PI3K/Akt and autophagy [57]; enhances EGFR signaling, MAPK, hypoxia inducible factor-1α pathways [59]; associated with HSP progress [58] | associated with breast tumorigenesis, promotes tumor cell growth, invasion and maintenance of metastasis [57], regulates apoptosis and cell proliferation [59] |
| DDX19A | DEAD-box helicase 19A/ATP-dependent RNA helicase DDX19A | RNA helicases involved in RNA metabolic process including transcription, RNA transport, RNA degradation [60] | overexpressed in CSCC [60], BC cell lines [61] | PT | induces EMT [60] | promotes cell migration, invasion, metastasis, NOX1 expression and ROS production [60] |
| GPI | Glucose-6-phosphate isomerase/neuroleukin | cytoplasmic glycolytic-related enzyme, secreted in ECM of cancer cells it is called autocrine motility factor (AMF) [62] and functions as a cytokine or growth factor [63] | overexpressed in BC [63], LUAD/NSCLC, glioblastoma, ccRCC [64], GC [62] | PT | REACTOME_GLUCONEOGENESIS; OXPHOS; glycolysis and gluconeogenesis [64], correlated with cell cycle regulatory genes, immune cell infiltration, gene alteration, ferroptosis genes [63]; AMF induces EMT [65] | involved in cell cycle, cell proliferation, correlates with immune infiltration, cell migration, invasion [64]; silencing suppressed proliferation, migration, invasion, glycolysis, and induced apoptosis [62] |
| TUBA1A | Tubulin alpha-I a | cell division, cell movement, microtubule based process, cell junction organization, cytoskeleton organization and cytoskeleton dependent intracellular transport | upregulated in BC tissues [66], GC [67] | PT | involved in infiltration of macrophages to the tumor microenvironment [67], involved in EMT related to re-organization of cell-cell contact [68] | overexpression was correlated with poor overall survival and a more aggressive phenotype in GC [67] |
| UTY/KDM6C | Ubiquitously transcribed tetratricopeptide repeat protein Y-linked transcript variant 83 | member of lysine (K)-specific dimethylase (KDM6) family that act as dynamic regulators of gene expression by histone demethylation [69], chromatin organization, anatomical structure development, gene transcription regulation [70] | not | controversial | REACTOME_CHROMATIN_MODIFYING_ENZYMES, transcriptional dysregulation in cancer (KEGG) | oncogenic or tumor suppressive roles [70] |
| NCAM2 | Neural cell adhesion molecule 2 | involved in cell adhesion, differentiation and anatomical structure development | overexpressed in some prostate and BC cell lines, such as ER-dependent BC cell lines MCF7 and T47D [71] | not known | not known | not known |
| RHBDD1 | Rhomboid-related protein 4 isoform X1/rhomboid domain-containing protein 1 | intramembrane/cytoplasmic-cleaving serine protease involved in intracellular protein transport, programmed cell death, protein catabolic process, cell differentiation, cell growth, protein maturation, participates in ER quality control system [72] and regulation of mitochondrial membrane remodeling [73] | highly upregulated in BC, CRC tissue and cell lines [72] | PT | positive correlation with p-Akt and CDK2 [72], c-Jun [73] | important in tumorigenesis, poor prognosis in ER+, ER+PR+, HER2+, and TNBC, inhibits apoptosis by activation of c-Jun and Bcl-3 [73]; deletion suppresses BC cell survival, migration, invasion, cycle progression and G1/S phase transition and increases apoptosis and ERAD [72] |
| CS | Citrate synthase | rate-limiting respiratory enzyme in the TCA cycle, involved in cell lipid metabolism (conversion of glucose to lipids) and mitochondrial function [74] | overexpressed in various cancers: PCa [74], pancreatic ductal carcinoma [75], ovarian carcinoma [76]; downregulated in human cervical carcinoma cells [77] | PT | OXPHOS; lipid metabolism signaling [74]; downregulation could induce EMT [77] | aggressive tumor progression, poor prognosis, increases cell proliferation, growth, colony formation, migration, invasion, and cell cycle [74]; silencing induces reduction of cell proliferation, invasion, migration, and enhances apoptosis [76], cells being unable to grow or proliferate in response to extracellular growth factors [78] |
| LAMTOR3/MAPKSP1/MP1 | Late endosomal/lysosomal adaptor, MAPK and MTOR activator 3/mitogen activated protein kinase scaffold protein 1/MEK partner 1 | member of the Ragulator complex involved in multiple signaling pathways that acts as a scaffold protein complex [79] | overexpressed in ER+ and ER- BC cell lines and in non-tumorigenic mammary epithelial cell lines [80]; downregulated in KIRC [79] | PT | considered to be a convergence point for MAPK and mTOR pathways [81]; targeting MEK1/MP1/ERK1/BCL2 axis may improve clinical outcome of MLCC patients [82] | required for pro-survival signaling from PI3K/AKT pathway in ER+ BC cells [80]; upregulation induces BCL2 expression (anti-apoptotic protein) [82] |
| OGA | Protein O-GlcNAcase (isoform a) | involved in protein glycosylation and protein catabolic process | overexpressed in numerous cancers [83] | PT | drives aerobic glycolysis and tumor growth by inhibiting PKM2 [83] | enhances tumor progression [83]; |
| ZNF114 | ZNF114 protein, partial | DNA-binding protein involved in transcription; member of KRAB-ZEPs family of transcription regulators [84] | overexpressed in ccRCC [85] | PT | pluripotency maintenance; repression of differentiation gene DPYSL4 [86] | involved in maintenance of cell pluripotency and stemness [84,86]; shorter overall survival [85] |
| PA2G4/EBP1 | Proliferation associated protein 2G4/ErbB3-binding protein 1 | found in cytoplasm and nucleus, regulates cell growth and differentiation, being a ribosomal constituent, transcriptional regulator, RNA/DNA-binding protein, mediates rRNA processing, DNA transcription, mRNA translation, protein stability and signal transduction [87] | overexpressed in HCC, cervical cancer, CC, NPC, salivary ACC, downregulated in HER2+ BC and bladder cancer [87] | PT | tumor formation via Ebp1/p38/HIF1α signaling and proto-oncogene MDM2-mediated downregulation of p53, promotes EMT [87] | intensively involved in tumorigenesis and cancer progression/metastasis, promotes cell proliferation and soft agar colony generation [87] |
| SRM | spermidine synthase | essential polyamine for cell proliferation, differentiation, development [88], regulation of gene expression, apoptosis, cell cycle progression and signaling pathways [89] | biosynthesis is upregulated in BC and contribute to disease progression [89], overexpression in CRC [90] | PT | MYC_TARGETS_V2; interferes with mTOR and RAS oncogenic pathways [88], c-MYC target [91] and C/ERPβ may serve as regulators of SRM [90] | cell growth [89] |
| GSTM3 | Glutathione S-transferase mu3 | enzyme involved in xenobiotic metabolism/detoxification, apoptosis inhibition [92], regulates ROS and participates in OS-mediated pathology [93] | mRNA expression level high in HER2+ or ER+ BC [93], overexpressed in cervical cancer, colon cancer [92] | PT | EMT inducer [94], cellular stress response via NF-κB and MAPK pathway during tumor progression [92] | cancer cell maintenance, survival and tumor progression [92] |
| NCKAP1/NAP1 | Noncatalytic region of tyrosine kinase (Nck)-associated protein 1 | associates with Src homology 3 (SH3) domain of NCK protein that localizes along the lamellipodia and mediates contact-cell dependent migration [95], member of WASF3 regulatory complex [96]; involved in cytoskeleton organization through actin polymerization, programmed cell death, signaling, cell differentiation and protein-containing complex assembly | overexpressed in NSCLC [95]; downregulated in ccRCC [97] | PT | ribosomal signaling, OXPHOS, TGF-β, EMT-related signaling pathways [97]; involved in HSP90-mediated invasion and metastasis by provoking MMP9 activation and EMT [95] | overexpression is associated with poorer survival in BC patients; essential for cell motility, adhesion, invasion and metastasis [95]; knockdown in BC cell lines, MDA-MB-231 and Hs578T, leads to a significant reduction in invasion and suppresses metastasis [96]; tumor suppressive in ccRCC [97] and HCC [98] |
| PRDX3 | Thioredoxin-dependent peroxide reductase, mitochondrial isoform a precursor | mitochondrial member of the antioxidant family of thioredoxin peroxidase [99] required for mitochondrial homeostasis [100] | overexpressed in HCC, malignant mesothelioma, BC, PCa, lung cancer, cervical carcinoma [99]; overexpression associated with ER and PR [100] | PT | OXPHOS; defense against H2O2 produced by mitochondrial respiratory chain [99], c-Myc target gene [100] | tumor promoting effects [99], involved in regulation of cell proliferation, differentiation and antioxidant function, overexpression protects cells from oxidative stress and apoptosis [100] |
| RAB8A | Ras-related protein Rab-8A | intracellular protein transport, intracellular membrane trafficking, autophagy, vesicle-mediated transport, signaling, cell differentiation, membrane organization | overexpressed in BC tissues [101] | PT | activates AKT and ERK1/2 signaling pathways [101] | increases cell growth, proliferation, migration, invasion [101] |
| RAB8B | Ras-related protein Rab-8B | member of the Rab small G protein family, immune system process, intracellular trafficking, peroxisome organization, vesicle-mediated transport, signaling, cell junction organization, membrane organization, protein localization to plasma membrane | overexpressed in TC [102] | PT | required for Wnt/β-catenin signaling [103] | overexpression and loss of functioning adherence junction accelerate tumorigenesis in testis [102] |
| RAB15 | Ras-related protein Rab-15 (isoform AN2) | involved in trafficking of cargos through the apical recycling endosome (ARE) to mediate transcytosis [104] | overexpressed in liver cancer cells [105] | PT | regulates the endocytic recycling pathway [106] | associated with the susceptibility of cells to DNA damage-induced cell death [105] |
| RAB35 | Ras-related protein Rab-35 (isoform 1) | Rab GTPase located in plasma membrane/endosomes, involved in vesicular trafficking, actin dynamics, cytokinesis, apical-basal polarity, endocytosis, phagocytosis, autophagy, exosome release [107] | overexpressed in OC [107] | PT | activator of PI3K/AKT pathway [107] | oncogenic protein, enhances BC cells invasion and metastasis [107] |
| Proteins with downregulated expression in phenotype of low JTB expression condition | ||||||
| TUBB4B | tubulin beta-4B chain | constituent of microtubules involved in mitotic cycle, immune system process and cytoskeleton organization | overexpressed in membranes of stem cells enriched cultures, PCa, OC, glioblastoma, metastatic CRC [108] | PT | downregulation is essential for the initiation of EMT [108], for microtubule-VIM interaction and contributes to the maintenance of polarity in migrating cells [108] | decreased level correlates with increased cell migration [109] |
| CAPN2 | Calpain 2/m-calpain | calcium-dependent, non-lysosomal cysteine protease involved in lipid metabolic process, cytoskeleton organization, programmed cell death, cell differentiation, protein catabolic process, protein maturation and anatomical structure development | overexpressed in various malignancies: CRPC cell lines (DU145, PC3), BL and TNBC [110], RCC [111], PC [112] | AT | acts via AKT/mTOR signaling pathway [111], and regulates Wnt/β-catenin signaling pathway-mediated EMT [112]; silencing may inhibit EMT [111,112] | oncogene involved in carcinogenesis and tumor progression, and metastasis silencing inhibits cell proliferation, migration and invasion by reducing MMP-2 and MMP-9 activation and regulation of invadopodia dynamics [111] |
| ELFN2/LRRC62 | Extracellular leucine rich repeat and fibronectin type III domain-containing 2/protein phosphatase 1 regulatory subunit 29 | putative oncogene, hypomethylation gene [113] | overexpressed in GC tissues and cell lines [114]; oncogene in GBM [113] | AT | interacts with AurkA and eukaryotic translation initiation factor 2 subunit alpha (EIF2α) and regulates the kinase activity of AurkA to promote cell autophagy [113] | knockdown inhibits cell proliferation, migration, invasion, increases E-cadherin and decreases N-cadherin [114] |
| SLC9AR1/NHERF1/ERB50 | Solute carrier (SLC) family 9 (Na(+)/H(+) exchanger), member 3 regulator 1/sodium-hydrogen exchanger regulatory factor 1/ERM-binding phosphoprotein | multifunctional cytoplasmic adaptor involved in growth factor signaling [115]; interacts with several proteins related to the estrogen pathway and tumorigenesis: EGFR, PTEN, PDGFR, beta catenin, EZR [116] | in BC acts as a tumor suppressor protein; oncogene in glioma and other cancers; overexpressed in PCa tissue and cell lines [117] | AT/PT | MTORC1_SIGNALING; ESTROGEN_RESPONSE_LATE; downregulation is associated with Wnt/β-catenin inactivation [118]; knockdown enhances PDGF-induced cytoskeletal rearrangements and chemotactic migration of cells [119]; AKT-associated protein [115] | knockdown suppresses proliferation and migration of metastatic PCa cells and promotes apoptosis [117] |
| ANXA4 | Annexin 4/lipocortin IV/endonexin I [120] | intracellular Ca2+ sensor that modulates membrane permeability and membrane trafficking, participates in cell growth, apoptosis [121], cell cycle and anticoagulation [122] | overexpressed in various tumors: chemoresistant LC, ESCC, GC, CRC, PCC, gallbladder, HCC, cholangiocarcinoma, BC, RCC, OCCC, laryngeal and PCa, MM [121,123], cervical cancer [120] | AT | overexpressed, is related to AKT, CDK1, and tumor suppressor p21 [122] | knockdown attenuates migration in OC and BC cells [121] |
| YWHAE | 14-3-3 protein epsilon | mitochondrial import stimulation factor L subunit [124] involved in mitotic cell cycle, intracellular protein transport, nucleocytoplasmic transport, signaling, cell differentiation, cell motility and transmembrane transport | overexpressed in BC tissue [124] | AT | MITOTIC_SPINDLE; MYC_TARGETS_V1 | knockdown reduces expression of Snail and Twist [124] |
| YWHAZ | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta | central hub protein involved in many signal transduction pathways [125] | oncogene overexpressed in multiple cancers: HCC, CRC, LUAD, BC [125], urothelial carcinomas [126] | AT | UPR | knockdown decreases cell growth, proliferation, invasion, enhances apoptosis and tamoxifen-induced inhibition of cell viability [125] |
| PSMB9/LMP2 | Proteasome 20S subunit 9 beta/low molecular weight protein 2 | immunoproteasome functions; major enzyme in ubiquitin-dependent protein degradation and inactivation [127] | overexpressed in tumor tissues: LGG [127]; deficient in uLMS [128] | PT | INTERFERON_ALPHA_RESPONSE; INTERFERON_GAMMA_RESPONSE; downregulation is associated with inhibition of pathways related to formation and development of ECM through proto-oncogene tyrosine-protein kinase (SRC) gene | defective expression contributes to abnormal cell proliferation and tumor progression [128] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jayathirtha, M.; Jayaweera, T.; Whitham, D.; Sullivan, I.; Petre, B.A.; Darie, C.C.; Neagu, A.-N. Two-Dimensional-PAGE Coupled with nLC-MS/MS-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in MCF7 Breast Cancer Cells Transfected for JTB Protein Silencing. Molecules 2023, 28, 7501. https://doi.org/10.3390/molecules28227501
Jayathirtha M, Jayaweera T, Whitham D, Sullivan I, Petre BA, Darie CC, Neagu A-N. Two-Dimensional-PAGE Coupled with nLC-MS/MS-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in MCF7 Breast Cancer Cells Transfected for JTB Protein Silencing. Molecules. 2023; 28(22):7501. https://doi.org/10.3390/molecules28227501
Chicago/Turabian StyleJayathirtha, Madhuri, Taniya Jayaweera, Danielle Whitham, Isabelle Sullivan, Brîndușa Alina Petre, Costel C. Darie, and Anca-Narcisa Neagu. 2023. "Two-Dimensional-PAGE Coupled with nLC-MS/MS-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in MCF7 Breast Cancer Cells Transfected for JTB Protein Silencing" Molecules 28, no. 22: 7501. https://doi.org/10.3390/molecules28227501
APA StyleJayathirtha, M., Jayaweera, T., Whitham, D., Sullivan, I., Petre, B. A., Darie, C. C., & Neagu, A.-N. (2023). Two-Dimensional-PAGE Coupled with nLC-MS/MS-Based Identification of Differentially Expressed Proteins and Tumorigenic Pathways in MCF7 Breast Cancer Cells Transfected for JTB Protein Silencing. Molecules, 28(22), 7501. https://doi.org/10.3390/molecules28227501