


Assessing the Protective Role of Epigallocatechin Gallate (EGCG) against Water-Pipe Smoke-Induced Toxicity: A Comparative Study on Gene Expression and Histopathology

 , ,
, ,  ,
, 
Abstract
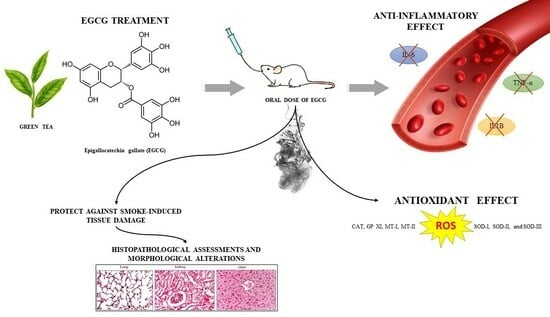
:
1. Introduction
2. Results
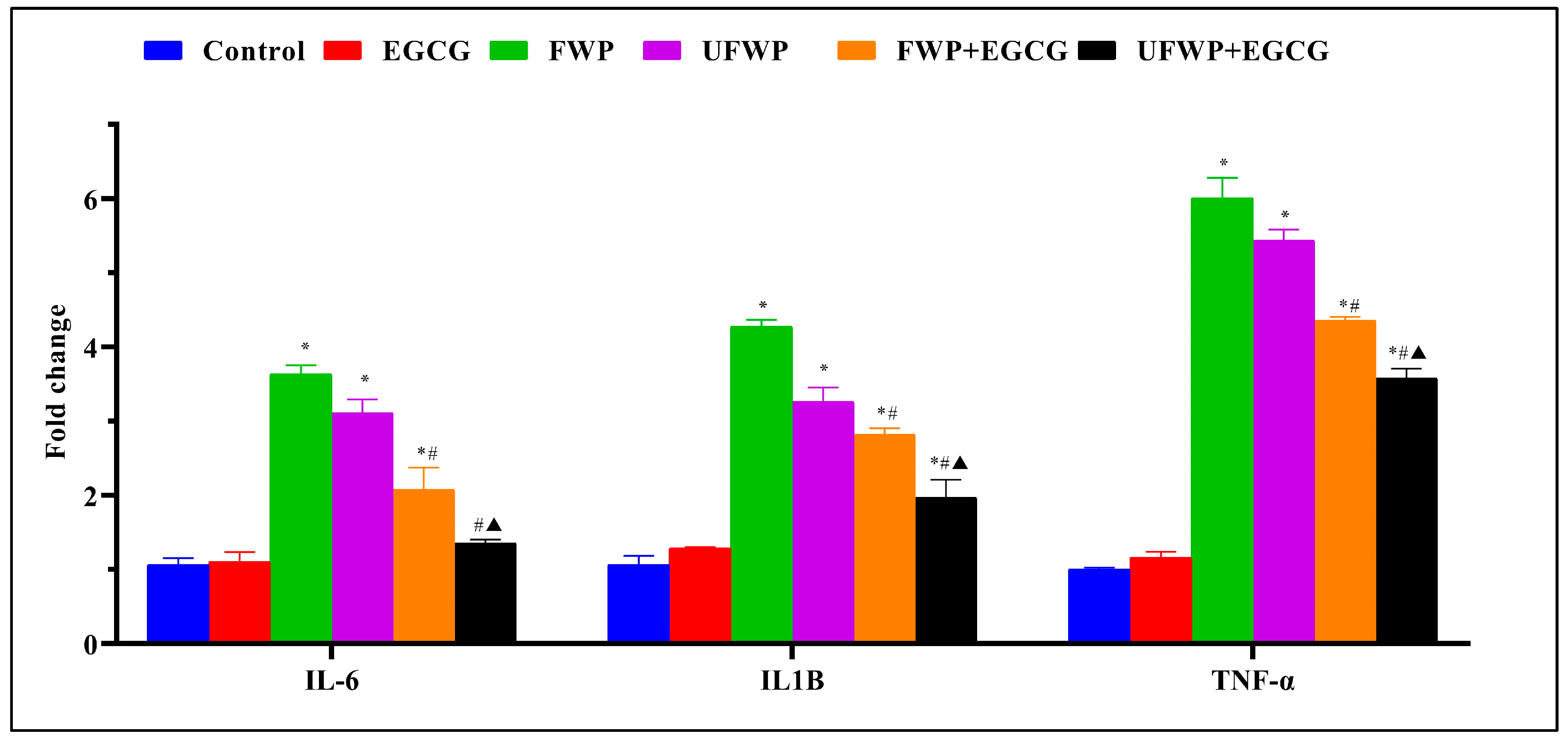
2.1. The Anti-Inflammatory Effects of Epigallocatechin gallate (EGCG) on Inflammatory Marker (IL-6, IL1B, and TNF-α) Expression in BALB/c mice Exposed to Flavored and Unflavored Water-Pipe Smoke
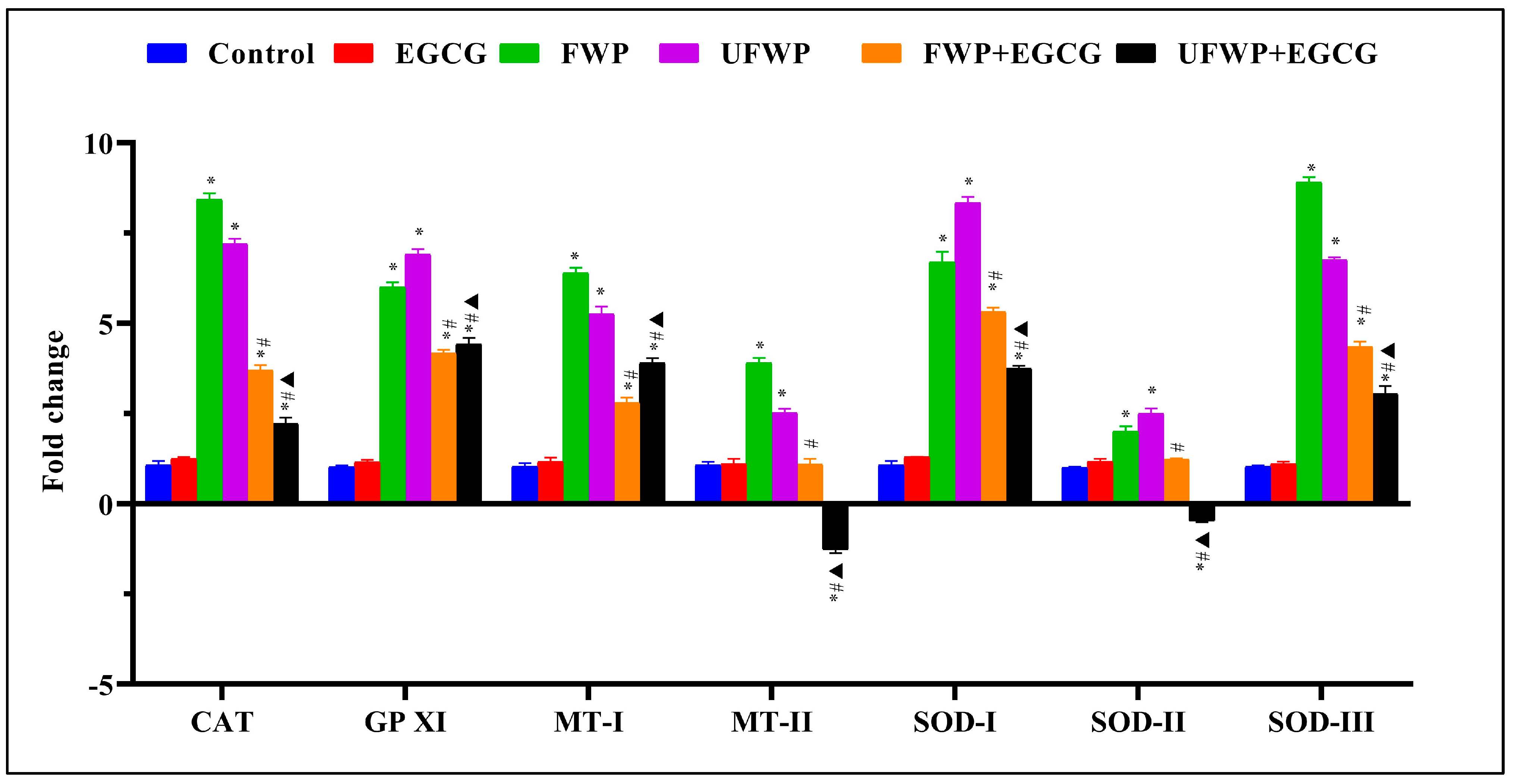
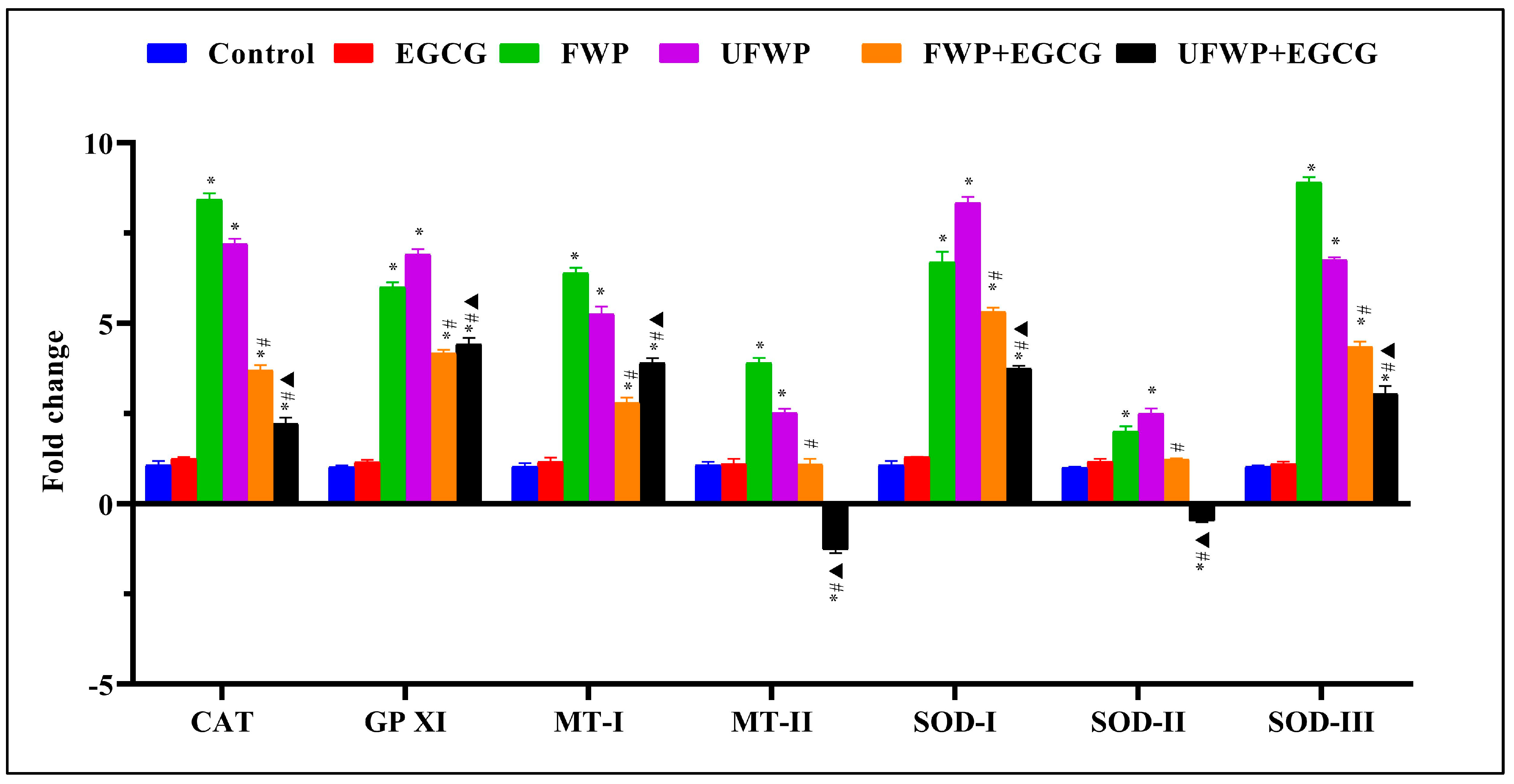
2.2. Modulating Effects of Epigallocatechin gallate (EGCG) on Antioxidant Gene Expression in Kidney Tissues Exposed to Flavored and Unflavored Water-Pipe Smoke
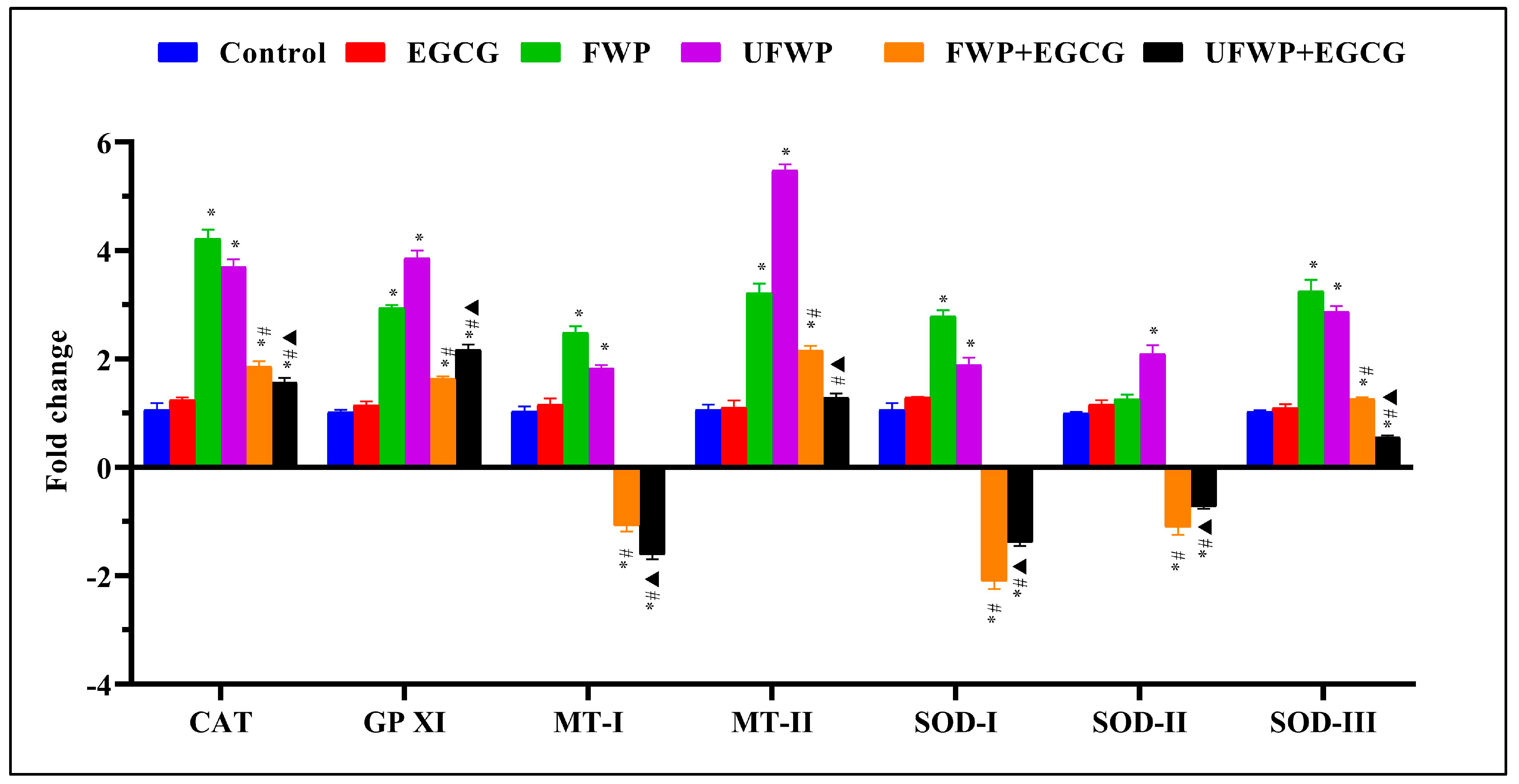
2.3. Modulating Effects of Epigallocatechin Gallate (EGCG) on Antioxidant Gene Expression in Liver Tissues Exposed to Flavored and Unflavored Water-Pipe Smoke
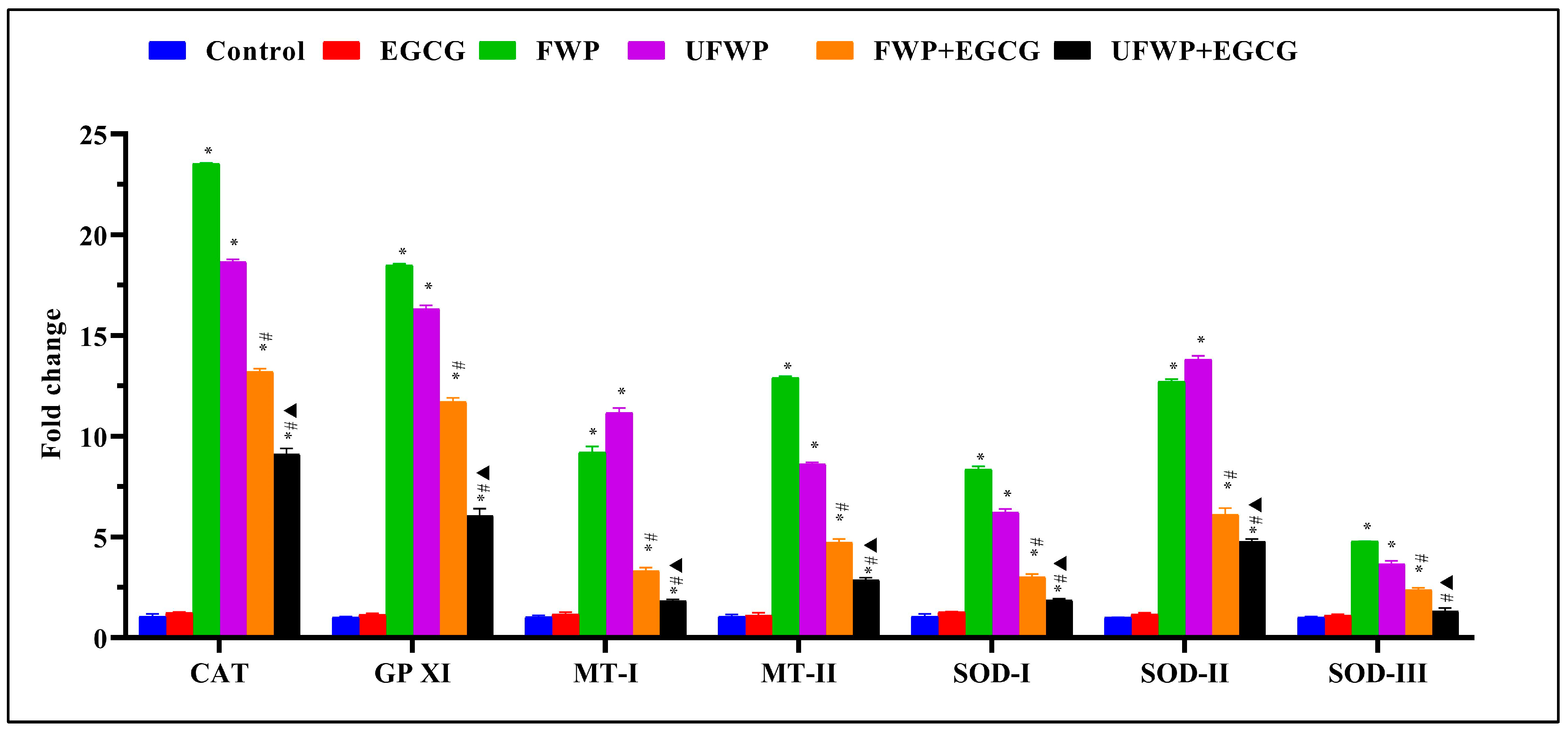
2.4. Modulating Effects of Epigallocatechin Gallate (EGCG) on Antioxidant Gene Expression in Lung Tissues Exposed to Flavored and Unflavored Water-Pipe Smoke
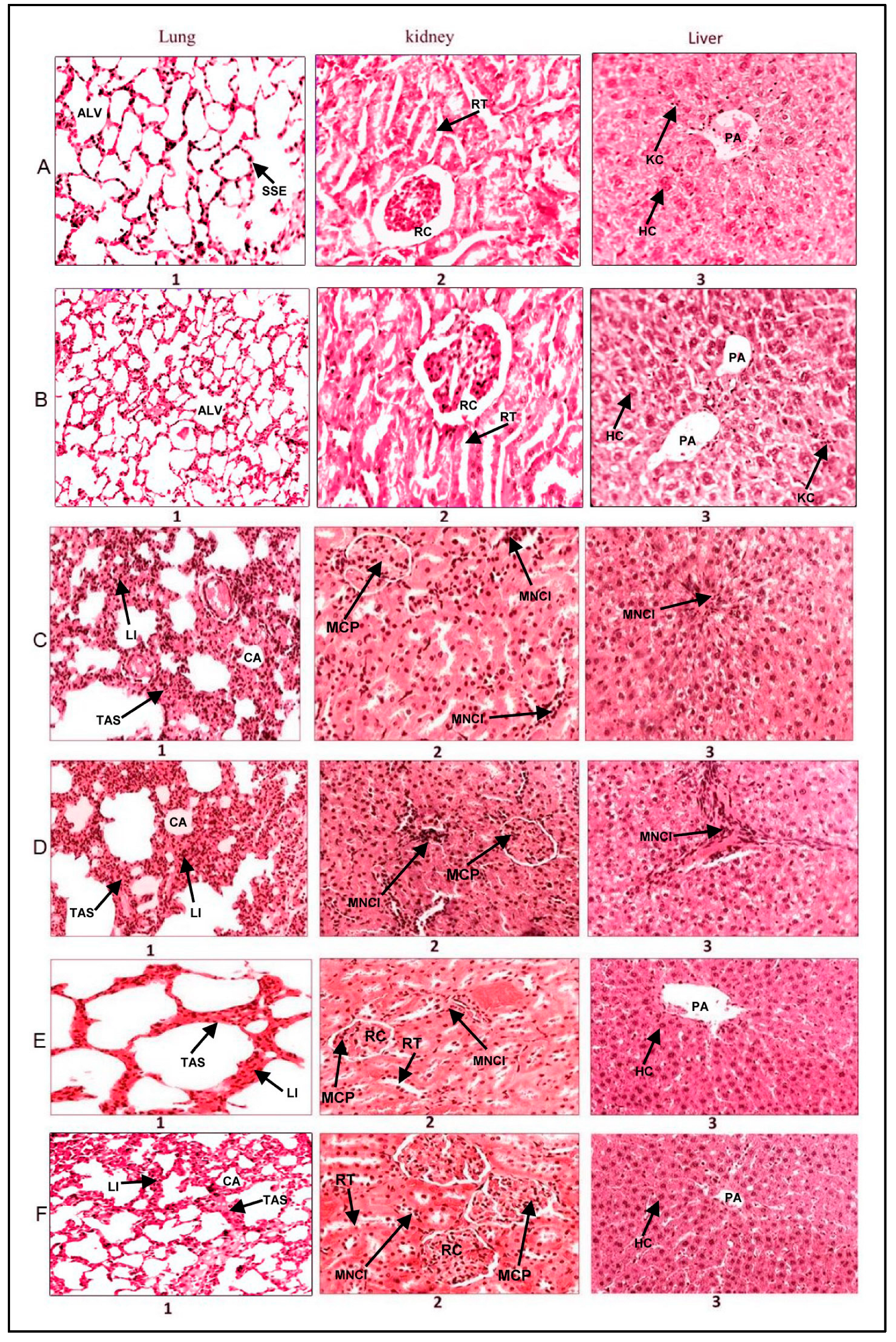
2.5. Histopathological Analysis
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Conditions
4.1.1. Animal Model
4.1.2. Grouping and Treatment Regimen
- Control Group: Subjected to ambient air without any external treatment.
- Flavored Smoke Group: Exposed to flavored water-pipe smoke.
- Unflavored Smoke Group: Exposed to unflavored water-pipe smoke.
- Unflavored Smoke + EGCG Group: Exposed to unflavored smoke and administered EGCG at a dosage of 50 mg/kg.
- Ambient Air + EGCG Group: Subjected to ambient air and given EGCG at a dosage of 50 mg/kg.
4.1.3. Housing Conditions
4.2. Smoke Exposure Mechanism
4.2.1. Device
4.2.2. Exposure Protocol
- In the first 30 s, mice were exposed to either flavored or unflavored tobacco smoke.
- In the subsequent 30 s, fresh air was introduced into the chamber to purge any remaining smoke.
- The final 30 s allowed the mice a period for respiration, during which they inhaled the introduced fresh air.
4.3. RNA Isolation and cDNA Synthesis
4.3.1. Sample Preparation
4.3.2. RNA Purification and cDNA Synthesis
4.4. Quantitative PCR (qPCR) and Gene Expression Analysis
4.4.1. System and Reagents
4.4.2. Normalization and Gene Targeting
4.5. Histopathological Analysis
4.5.1. Sample Preparation
4.5.2. Staining and Imaging
4.6. Statistical Analysis
4.6.1. Software Utilization
4.6.2. Data Normality
4.6.3. Experimental Replicates
4.6.4. Statistical Tests
4.6.5. Significance Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maziak, W.; Taleb, Z.B.; Bahelah, R.; Islam, F.; Jaber, R.; Auf, R.; Salloum, R.G. The global epidemiology of waterpipe smoking. Tob. Control 2015, 24, i3–i12. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Xie, Z.; Li, D. Discussion of waterpipe tobacco smoking on reddit. Heliyon 2022, 8, e10635. [Google Scholar] [CrossRef] [PubMed]
- Al-Awaida, W.; Najjar, H.; Shraideh, Z. Structural characterization of rat ventricular tissue exposed to the smoke of two types of waterpipe. Iran. J. Basic Med. Sci. 2015, 18, 942. [Google Scholar] [CrossRef]
- Khwaldeh, A.; Siyam, A.A.; Alzbeede, A.; Farajallah, M.; Shraideh, Z.; Badran, D. Ameliorative effects of curcumin and caffeic acid against short term exposure of waterpipe tobacco smoking on lung, heart and kidney in mice. Anat. Cell Biol. 2021, 54, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Unsal, V.; Dalkıran, T.; Çiçek, M.; Kölükçü, E. The role of natural antioxidants against reactive oxygen species produced by cadmium toxicity: A review. Adv. Pharm. Bull. 2020, 10, 184. [Google Scholar] [CrossRef] [PubMed]
- Al-Awaida, W.J.; Zihlif, M.A.; Al-Ameer, H.J.; Sharab, A.; Akash, M.; Aburubaiha, Z.A.; Fattash, I.A.; Imraish, A.; Ali, K.H. The effect of green tea consumption on the expression of antioxidant-and inflammation-related genes induced by nicotine. J. Food Biochem. 2019, 43, e12874. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Sulik, K.K.; Chen, S.-y. Nrf2-mediated transcriptional induction of antioxidant response in mouse embryos exposed to ethanol in vivo: Implications for the prevention of fetal alcohol spectrum disorders. Antioxid. Redox Signal. 2008, 10, 2023–2033. [Google Scholar] [CrossRef]
- Jiang, Q.; Qiao, B.; Lin, X.; Cao, J.; Zhang, N.; Guo, H.; Liu, W.; Zhu, L.; Xie, X.; Wan, L. A hydrogen peroxide economizer for on-demand oxygen production-assisted robust sonodynamic immunotherapy. Theranostics 2022, 12, 59. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants 2021, 7, 403–412. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Zhou, J.; Shao, Q. Glutathione peroxidase GPX1 and its dichotomous roles in cancer. Cancers 2022, 14, 2560. [Google Scholar] [CrossRef]
- Nordberg, M.; Nordberg, G.F. Metallothionein and cadmium toxicology—Historical review and commentary. Biomolecules 2022, 12, 360. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Gao, J.-Y.; Ma, J.; Xu, X.; Wang, Q.; Xiong, L.; Yang, J.; Ren, J. Cardiac-specific overexpression of metallothionein attenuates myocardial remodeling and contractile dysfunction in l-NAME-induced experimental hypertension: Role of autophagy regulation. Toxicol. Lett. 2015, 237, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef]
- Wang, H.; Chen, H.; Fu, Y.; Liu, M.; Zhang, J.; Han, S.; Tian, Y.; Hou, H.; Hu, Q. Effects of smoking on inflammatory-related cytokine levels in human serum. Molecules 2022, 27, 3715. [Google Scholar] [CrossRef] [PubMed]
- Aldaham, S.; Foote, J.A.; Chow, H.-H.S.; Hakim, I.A. Smoking status effect on inflammatory markers in a randomized trial of current and former heavy smokers. Int. J. Inflamm. 2015, 2015, 439396. [Google Scholar] [CrossRef] [PubMed]
- Kochman, J.; Jakubczyk, K.; Antoniewicz, J.; Mruk, H.; Janda, K. Health benefits and chemical composition of matcha green tea: A review. Molecules 2020, 26, 85. [Google Scholar] [CrossRef]
- Al-Awaida, W.; Akash, M.; Aburubaiha, Z.; Talib, W.H.; Shehadeh, H. Chinese green tea consumption reduces oxidative stress, inflammation and tissues damage in smoke exposed rats. Iran. J. Basic Med. Sci. 2014, 17, 740. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial properties of green tea catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef]
- Barreira, S.; Moutinho, C.; Silva, A.M.; Neves, J.; Seo, E.-J.; Hegazy, M.-E.F.; Efferth, T.; Gomes, L.R. Phytochemical characterization and biological activities of green tea (Camellia sinensis) produced in the Azores, Portugal. Phytomedicine Plus 2021, 1, 100001. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydhi, F.A.; Alsahli, M.A.; Rahmani, A.H. Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 2020, 25, 3146. [Google Scholar] [CrossRef]
- Hong, Y.; Zhou, X.; Li, Q.; Chen, J.; Wei, Y.; Shen, L.; Long, C.; Wu, S.; Wei, G. Epigallocatechin gallate alleviates mono-2-ethylhexyl phthalate-induced male germ cell pyroptosis by inhibiting the ROS/mTOR/NLRP3 pathway. Toxicol. Vitr. 2023, 2023, 105626. [Google Scholar] [CrossRef]
- Ferrari, E.; Bettuzzi, S.; Naponelli, V. The potential of epigallocatechin gallate (EGCG) in targeting autophagy for cancer treatment: A narrative review. Int. J. Mol. Sci. 2022, 23, 6075. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, S.; Maqsood, M.; Mahtab, N.; Khan, M.I.; Sahar, A.; Zaib, S.; Gul, S. Epigallocatechin gallate: Phytochemistry, bioavailability, utilization challenges, and strategies. J. Food Biochem. 2022, 46, e14189. [Google Scholar] [CrossRef]
- Lange, K.W.; Lange, K.M.; Nakamura, Y. Green tea, epigallocatechin gallate and the prevention of Alzheimer’s disease: Clinical evidence. Food Sci. Hum. Wellness 2022, 11, 765–770. [Google Scholar] [CrossRef]
- Payne, A.; Nahashon, S.; Taka, E.; Adinew, G.M.; Soliman, K.F. Epigallocatechin-3-Gallate (EGCG): New therapeutic perspectives for neuroprotection, aging, and neuroinflammation for the modern age. Biomolecules 2022, 12, 371. [Google Scholar] [CrossRef] [PubMed]
- Ntamo, Y.; Jack, B.; Ziqubu, K.; Mazibuko-Mbeje, S.E.; Nkambule, B.B.; Nyambuya, T.M.; Mabhida, S.E.; Hanser, S.; Orlando, P.; Tiano, L.; et al. Epigallocatechin gallate as a nutraceutical to potentially target the metabolic syndrome: Novel insights into therapeutic effects beyond its antioxidant and anti-inflammatory properties. Crit. Rev. Food Sci. Nutr. 2022, 2022, 2104805. [Google Scholar] [CrossRef]
- Beyaz, S.; Özlem, G.; Aslan, A. The therapeutic effects and antioxidant properties of epigallocatechin-3 gallate: A new review. Int. J. Second. Metab. 2022, 9, 125–136. [Google Scholar] [CrossRef]
- Chan, P.C.; Ramot, Y.; Malarkey, D.E.; Blackshear, P.; Kissling, G.E.; Travlos, G.; Nyska, A. Fourteen-week toxicity study of green tea extract in rats and mice. Toxicol. Pathol. 2010, 38, 1070–1084. [Google Scholar] [CrossRef]
- Isbrucker, R.; Edwards, J.; Wolz, E.; Davidovich, A.; Bausch, J. Safety studies on epigallocatechin gallate (EGCG) preparations. Part 2: Dermal, acute and short-term toxicity studies. Food Chem. Toxicol. 2006, 44, 636–650. [Google Scholar] [CrossRef]
- Takami, S.; Imai, T.; Hasumura, M.; Cho, Y.-M.; Onose, J.; Hirose, M. Evaluation of toxicity of green tea catechins with 90-day dietary administration to F344 rats. Food Chem. Toxicol. 2008, 46, 2224–2229. [Google Scholar] [CrossRef]
- Chengelis, C.P.; Kirkpatrick, J.B.; Regan, K.S.; Radovsky, A.E.; Beck, M.J.; Morita, O.; Tamaki, Y.; Suzuki, H. 28-Day oral (gavage) toxicity studies of green tea catechins prepared for beverages in rats. Food Chem. Toxicol. 2008, 46, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-W.; Tsai, C.-F.; Chen, W.-K.; Huang, C.-F.; Yen, C.-C. A subacute toxicity evaluation of green tea (Camellia sinensis) extract in mice. Food Chem. Toxicol. 2011, 49, 2624–2630. [Google Scholar] [CrossRef] [PubMed]
- Saleh, I.G.; Ali, Z.; Abe, N.; Wilson, F.D.; Hamada, F.M.; Abd-Ellah, M.F.; Walker, L.A.; Khan, I.A.; Ashfaq, M.K. Effect of green tea and its polyphenols on mouse liver. Fitoterapia 2013, 90, 151–159. [Google Scholar] [CrossRef]
- Santos, C.N.; Gomes, A.; Oudot, C.; Dias-Pedroso, D.; Rodriguez-Mateos, A.; Vieira, H.L.; Brenner, C. Pure polyphenols applications for cardiac health and disease. Curr. Pharm. Des. 2018, 24, 2137–2156. [Google Scholar] [CrossRef]
- Hazimeh, D.; Massoud, G.; Parish, M.; Singh, B.; Segars, J.; Islam, M.S. Green Tea and Benign Gynecologic Disorders: A New Trick for An Old Beverage? Nutrients 2023, 15, 1439. [Google Scholar] [CrossRef]
- Li, Y.; Karim, M.R.; Wang, B.; Peng, J. Effects of Green Tea (−)-Epigallocatechin-3-Gallate (EGCG) on Cardiac Function-A Review of the Therapeutic Mechanism and Potentials. Mini Rev. Med. Chem. 2022, 22, 2371–2382. [Google Scholar] [CrossRef]
- Stringer, M.; Abeysekera, I.; Thomas, J.; LaCombe, J.; Stancombe, K.; Stewart, R.J.; Dria, K.J.; Wallace, J.M.; Goodlett, C.R.; Roper, R.J. Epigallocatechin-3-gallate (EGCG) consumption in the Ts65Dn model of Down syndrome fails to improve behavioral deficits and is detrimental to skeletal phenotypes. Physiol. Behav. 2017, 177, 230–241. [Google Scholar] [CrossRef]
- Kang, S.U.; Lee, B.-S.; Lee, S.-H.; Baek, S.J.; Shin, Y.S.; Kim, C.-H. Expression of NSAID-activated gene-1 by EGCG in head and neck cancer: Involvement of ATM-dependent p53 expression. J. Nutr. Biochem. 2013, 24, 986–999. [Google Scholar] [CrossRef]
- Liu, T. miR-937 serves as an inflammatory inhibitor in cigarette smoke extract-induced human bronchial epithelial cells by targeting IL1B and regulating TNF-α/IL-17 signaling pathway. Tob. Induc. Dis. 2021, 19, 138227. [Google Scholar] [CrossRef]
- Chaouachi, K. Public health intervention for narghile (hookah, shisha) use requires a radical critique of the related “standardised” smoking machine. J. Public Health 2010, 18, 69–73. [Google Scholar] [CrossRef]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic effects of green tea polyphenol (−)-Epigallocatechin-3-Gallate (EGCG) in relation to molecular pathways controlling inflammation, oxidative stress, and apoptosis. Int. J. Mol. Sci. 2022, 24, 340. [Google Scholar] [CrossRef] [PubMed]
- Hassani, S.; Maqbool, F.; Salek-Maghsoudi, A.; Rahmani, S.; Shadboorestan, A.; Nili-Ahmadabadi, A.; Amini, M.; Norouzi, P.; Abdollahi, M. Alteration of hepatocellular antioxidant gene expression pattern and biomarkers of oxidative damage in diazinon-induced acute toxicity in Wistar rat: A time-course mechanistic study. EXCLI J. 2018, 17, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, J.; Byun, H.; Kim, S.-J.; Joo, J.; Park, H.H.; Shin, H. Evaluation of the anti-oxidative and ROS scavenging properties of biomaterials coated with epigallocatechin gallate for tissue engineering. Acta Biomater. 2021, 124, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, M.M.; Solanki, P.; Singh, P. Oxidative stress, antioxidants, hormesis and calorie restriction: The current perspective in the biology of aging. Arch. Gerontol. Geriatr. 2021, 95, 104413. [Google Scholar] [CrossRef] [PubMed]
- Raad, D.; Gaddam, S.; Schunemann, H.J.; Irani, J.; Abou Jaoude, P.; Honeine, R.; Akl, E.A. Effects of water-pipe smoking on lung function: A systematic review and meta-analysis. Chest 2011, 139, 764–774. [Google Scholar] [CrossRef]
- Delwing-Dal Magro, D.; Roecker, R.; Junges, G.M.; Rodrigues, A.F.; Delwing-de Lima, D.; da Cruz, J.G.; Wyse, A.T.; Pitz, H.S.; Zeni, A.L. Protective effect of green tea extract against proline-induced oxidative damage in the rat kidney. Biomed. Pharmacother. 2016, 83, 1422–1427. [Google Scholar] [CrossRef]
- Naware, N.S.; Ambatkar, S.S.; Kamble, T.S.; Bangar, S.; Uppar, K.B.; Shirke, K.; Patil, M.; Jain, A. A review focusing on the benefits of green tea catechins as nutraceuticals. Sci. Phytochem. 2023, 2, 1–12. [Google Scholar] [CrossRef]
- Chacko, S.M.; Thambi, P.T.; Kuttan, R.; Nishigaki, I. Beneficial effects of green tea: A literature review. Chin. Med. 2010, 5, 1–9. [Google Scholar] [CrossRef]
- Dawood, M.A.; Zommara, M.; Eweedah, N.M.; Helal, A.I. The evaluation of growth performance, blood health, oxidative status and immune-related gene expression in Nile tilapia (Oreochromis niloticus) fed dietary nanoselenium spheres produced by lactic acid bacteria. Aquaculture 2020, 515, 734571. [Google Scholar] [CrossRef]
- Takano, S.; Uchida, K.; Miyagi, M.; Inoue, G.; Fujimaki, H.; Aikawa, J.; Iwase, D.; Minatani, A.; Iwabuchi, K.; Takaso, M. Nerve growth factor regulation by TNF-α and IL-1β in synovial macrophages and fibroblasts in osteoarthritic mice. J. Immunol. Res. 2016, 2016, 5706359. [Google Scholar] [CrossRef]
- Pankhurst, M.W.; Gell, D.A.; Butler, C.W.; Kirkcaldie, M.T.; West, A.K.; Chung, R.S. Metallothionein (MT)-I and MT−II expression are induced and cause zinc sequestration in the liver after brain injury. PLoS ONE 2012, 7, e31185. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|
| IL1β | AAGGATGACGACAAGCCAAC | CGCTGTGCTGATGTACCAGT | [49] |
| IL6 | CTGCAAGAGACTTCCATCCAG | AGTGGTATAGACAGGTCTGTTGG | [50] |
| TNFα | GAGGCCAATAAAATCATCATCCC | CTTCCCATAGACTCTGAGTAGCG | [49] |
| MT1 | GCTGTCCTCTAAGCGTCACC | AGGAGCAGCAGCTCTTCTTG | [51] |
| MT2 | CAAACCGATCTCTCGTCGAT | AGGAGCAGCAGCTTTTCTTG | [51] |
| SOD1 | GTGATTGGGATTGCGCAGTA | TGGTTTGAGGGTAGCAGATGAGT | [7] |
| SOD2 | TTAACGCGCAGATCATGCA | GGTGGCGTTGAGATTGTTCA | [7] |
| SOD3 | CATGCAATCTGCAGGGTACAA | AGAACCAAGCCGGTGATCTG | [7] |
| GPX1 | GAAGAACTTGGGCCATTTGG | TCTCGCCTGGCTCCTGTTT | [7] |
| CAT | TGAGAAGCCTAAGAACGCAATTC | CCCTTCGCAGCCATGTG | [7] |
| GAPDH | AACGACCCCTTCATTGAC | TCCACGACATACTCAGCAC | [7] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Awaida, W.; Goh, K.W.; Al-Ameer, H.J.; Gushchina, Y.S.; Torshin, V.I.; Severin, A.E.; Al Bawareed, O.; Srour, B.; Al Farraj, J.; Hamad, I. Assessing the Protective Role of Epigallocatechin Gallate (EGCG) against Water-Pipe Smoke-Induced Toxicity: A Comparative Study on Gene Expression and Histopathology. Molecules 2023, 28, 7502. https://doi.org/10.3390/molecules28227502
Al-Awaida W, Goh KW, Al-Ameer HJ, Gushchina YS, Torshin VI, Severin AE, Al Bawareed O, Srour B, Al Farraj J, Hamad I. Assessing the Protective Role of Epigallocatechin Gallate (EGCG) against Water-Pipe Smoke-Induced Toxicity: A Comparative Study on Gene Expression and Histopathology. Molecules. 2023; 28(22):7502. https://doi.org/10.3390/molecules28227502
Chicago/Turabian StyleAl-Awaida, Wajdy, Khang Wen Goh, Hamzeh J. Al-Ameer, Yulia Sh. Gushchina, Vladimir I. Torshin, Alexandr E. Severin, Omar Al Bawareed, Besan Srour, Jude Al Farraj, and Islam Hamad. 2023. "Assessing the Protective Role of Epigallocatechin Gallate (EGCG) against Water-Pipe Smoke-Induced Toxicity: A Comparative Study on Gene Expression and Histopathology" Molecules 28, no. 22: 7502. https://doi.org/10.3390/molecules28227502
APA StyleAl-Awaida, W., Goh, K. W., Al-Ameer, H. J., Gushchina, Y. S., Torshin, V. I., Severin, A. E., Al Bawareed, O., Srour, B., Al Farraj, J., & Hamad, I. (2023). Assessing the Protective Role of Epigallocatechin Gallate (EGCG) against Water-Pipe Smoke-Induced Toxicity: A Comparative Study on Gene Expression and Histopathology. Molecules, 28(22), 7502. https://doi.org/10.3390/molecules28227502