
Exploring the Anti-Cancer Effects of Fish Bone Fermented Using Monascus purpureus: Induction of Apoptosis and Autophagy in Human Colorectal Cancer Cells

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
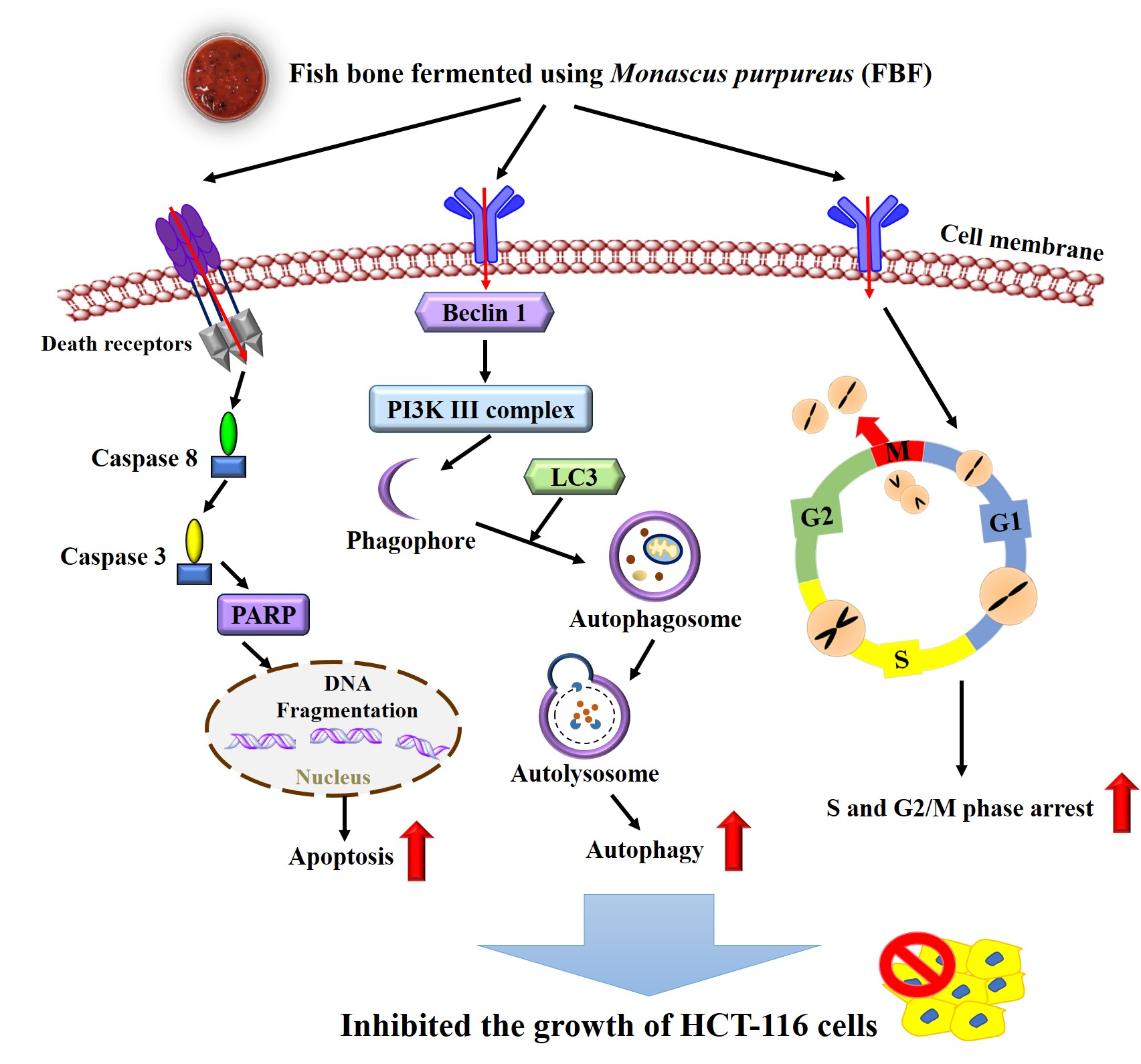
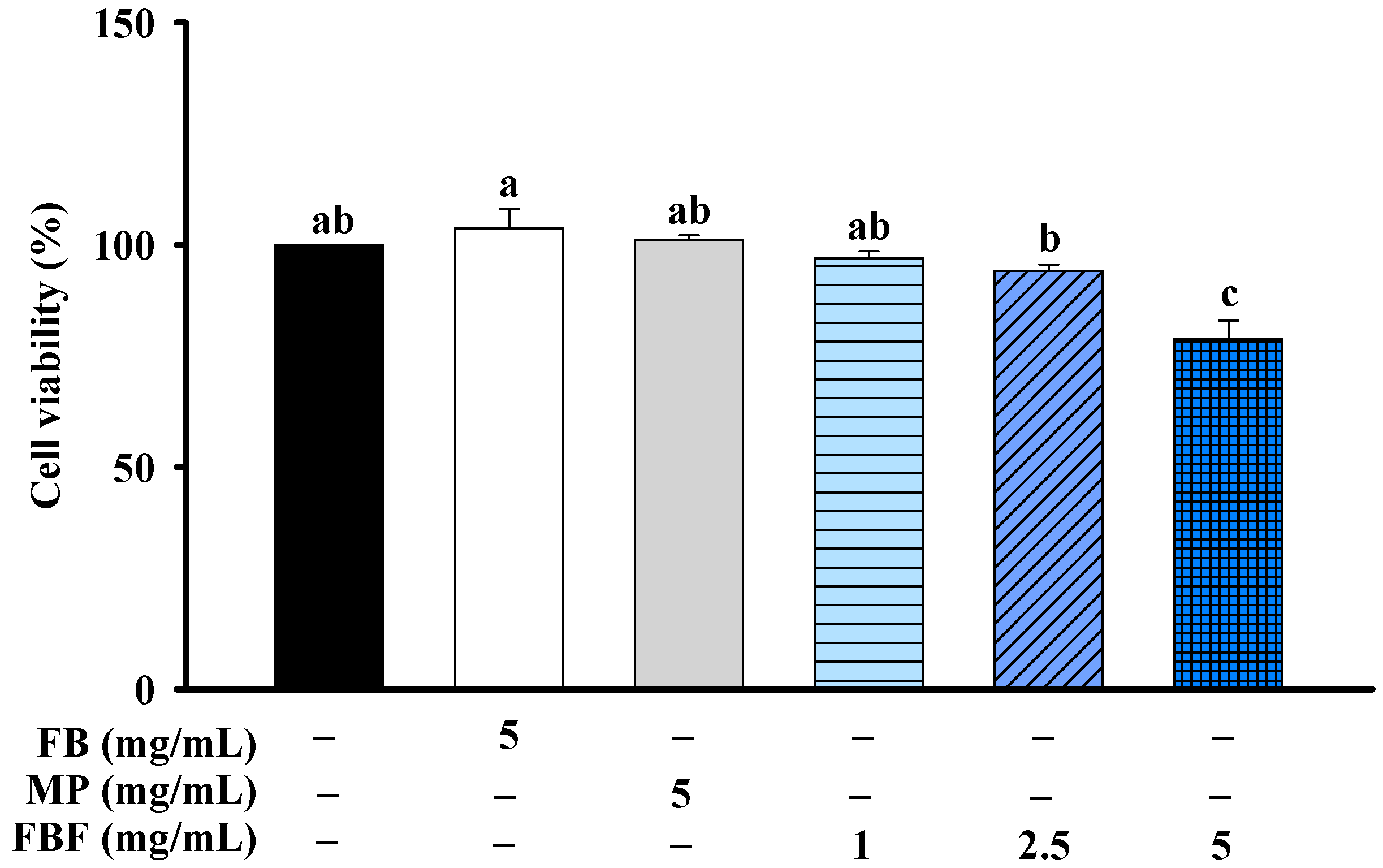
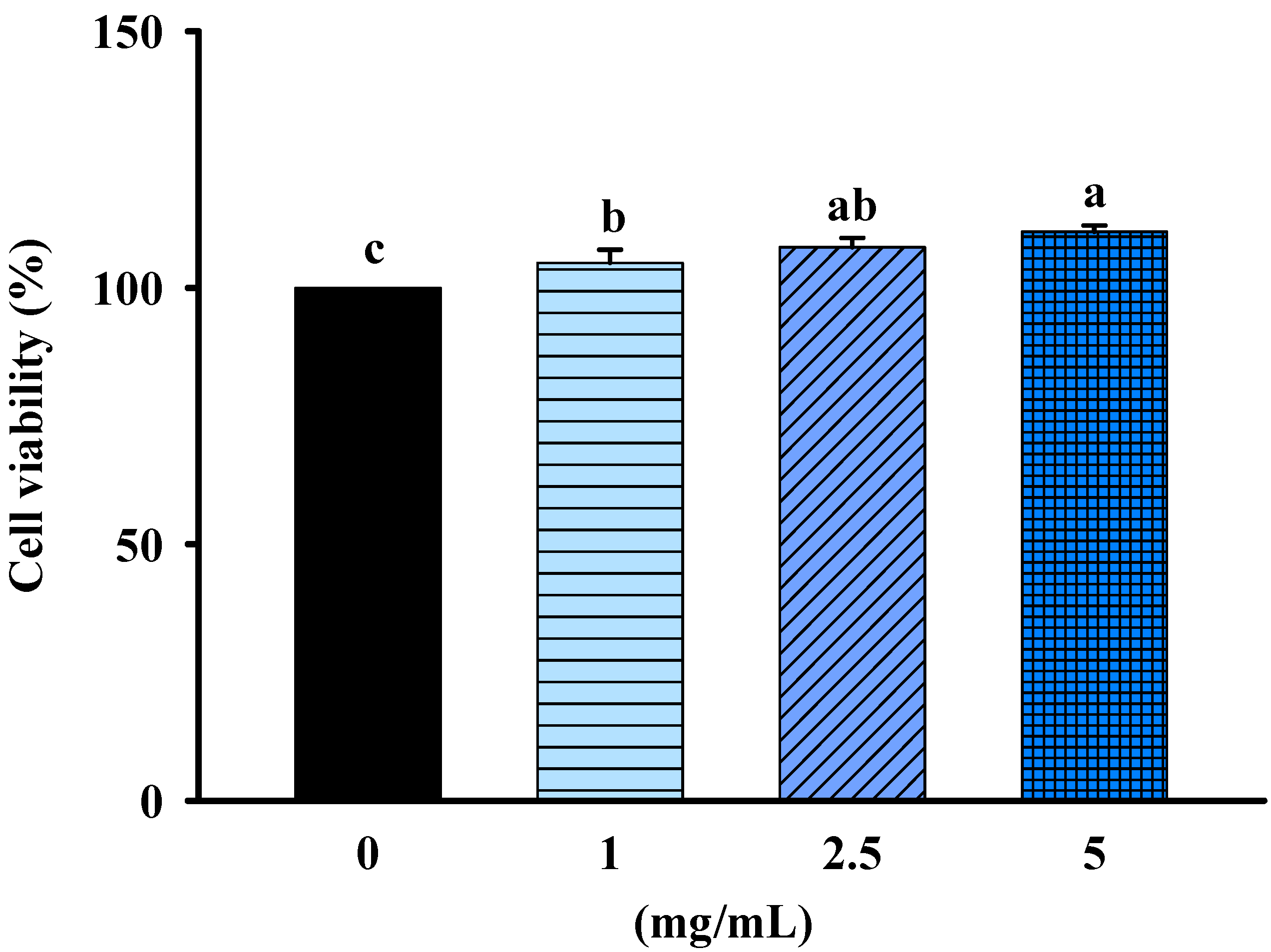
2.1. Effects of FBF on Cell Viability
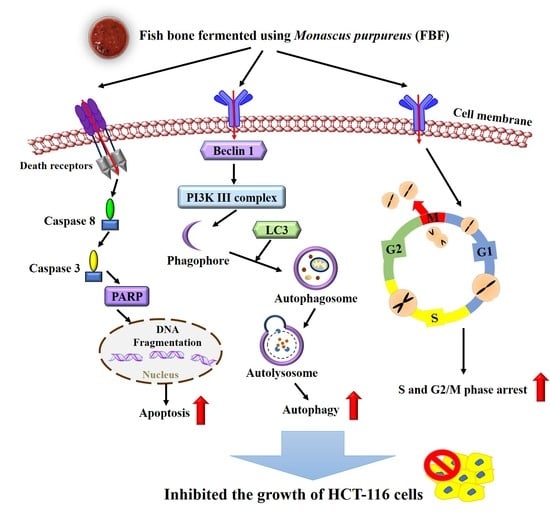
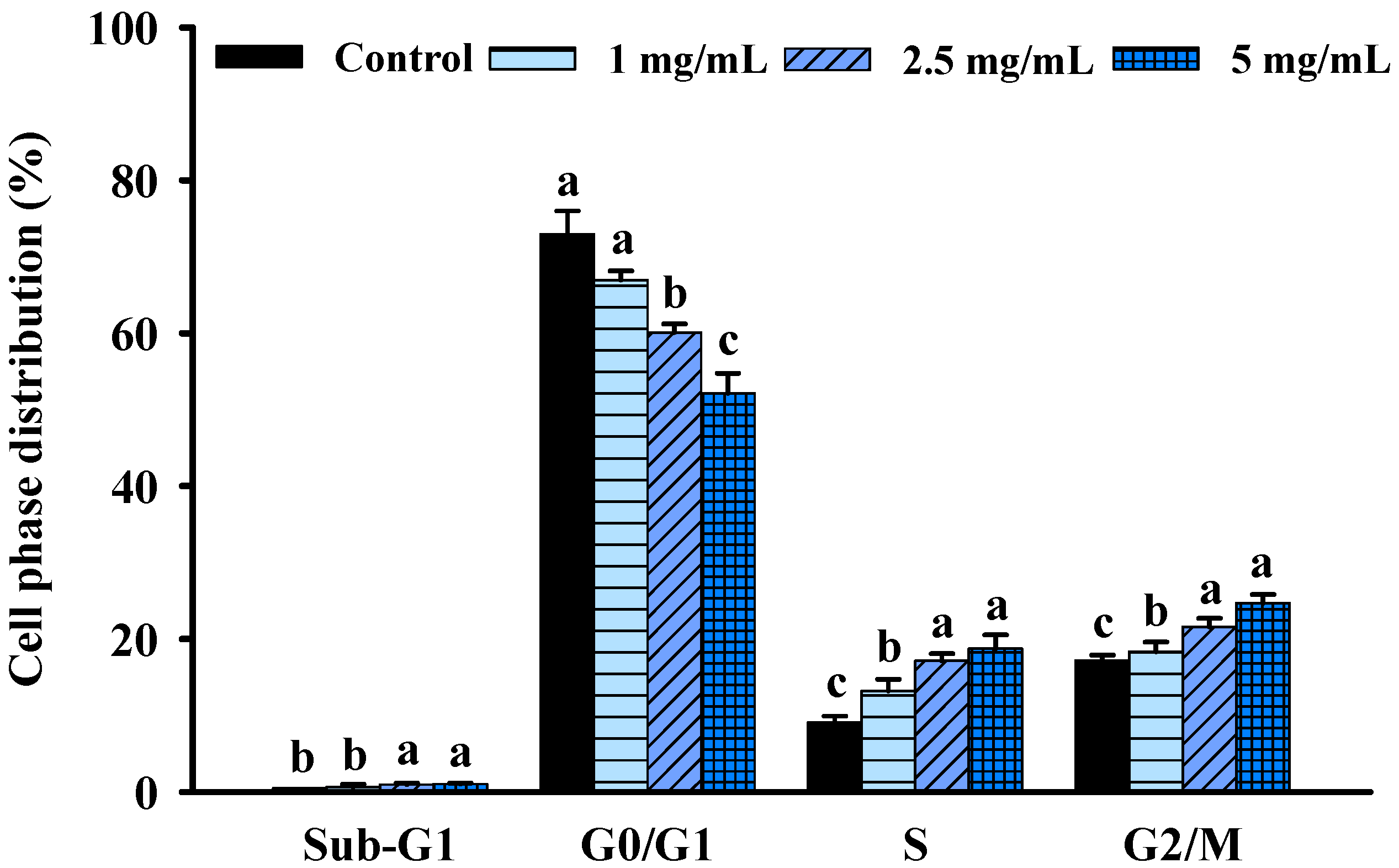
2.2. FBF Inhibits the Growth of HCT-116 Cells by Inducing S and G2/M-Phase Arrest
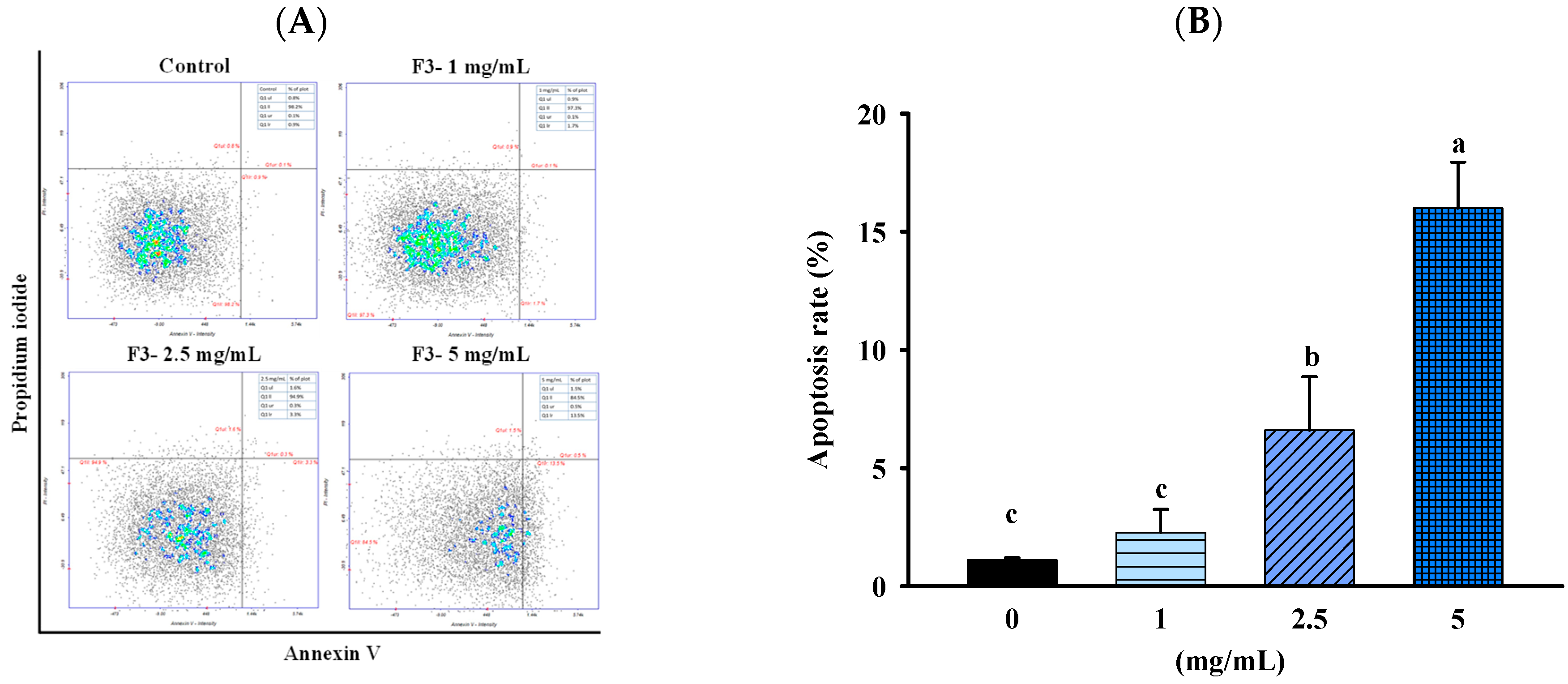
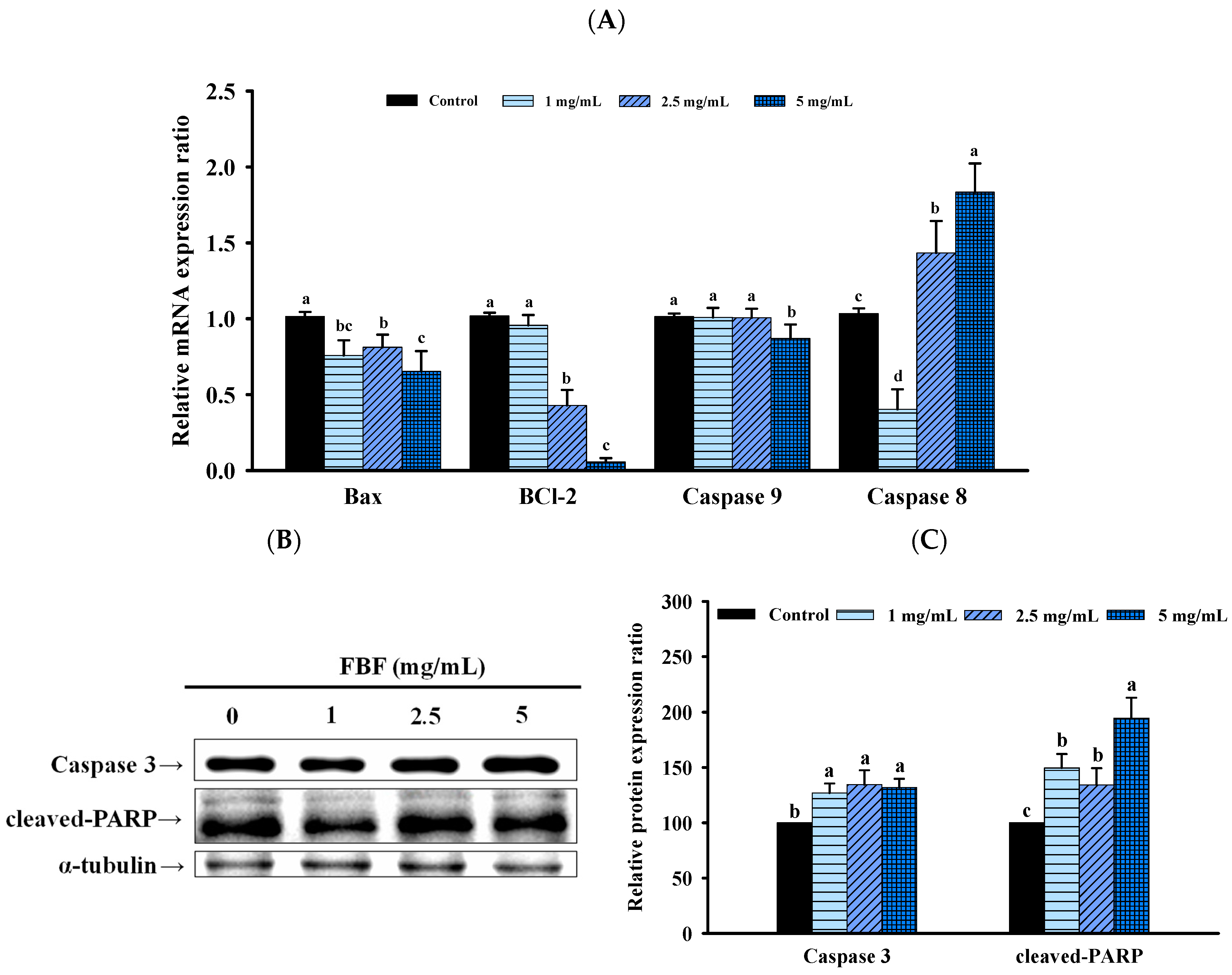
2.3. FBF Inhibits the Growth of HCT-116 Cells by Inducing Apoptosis
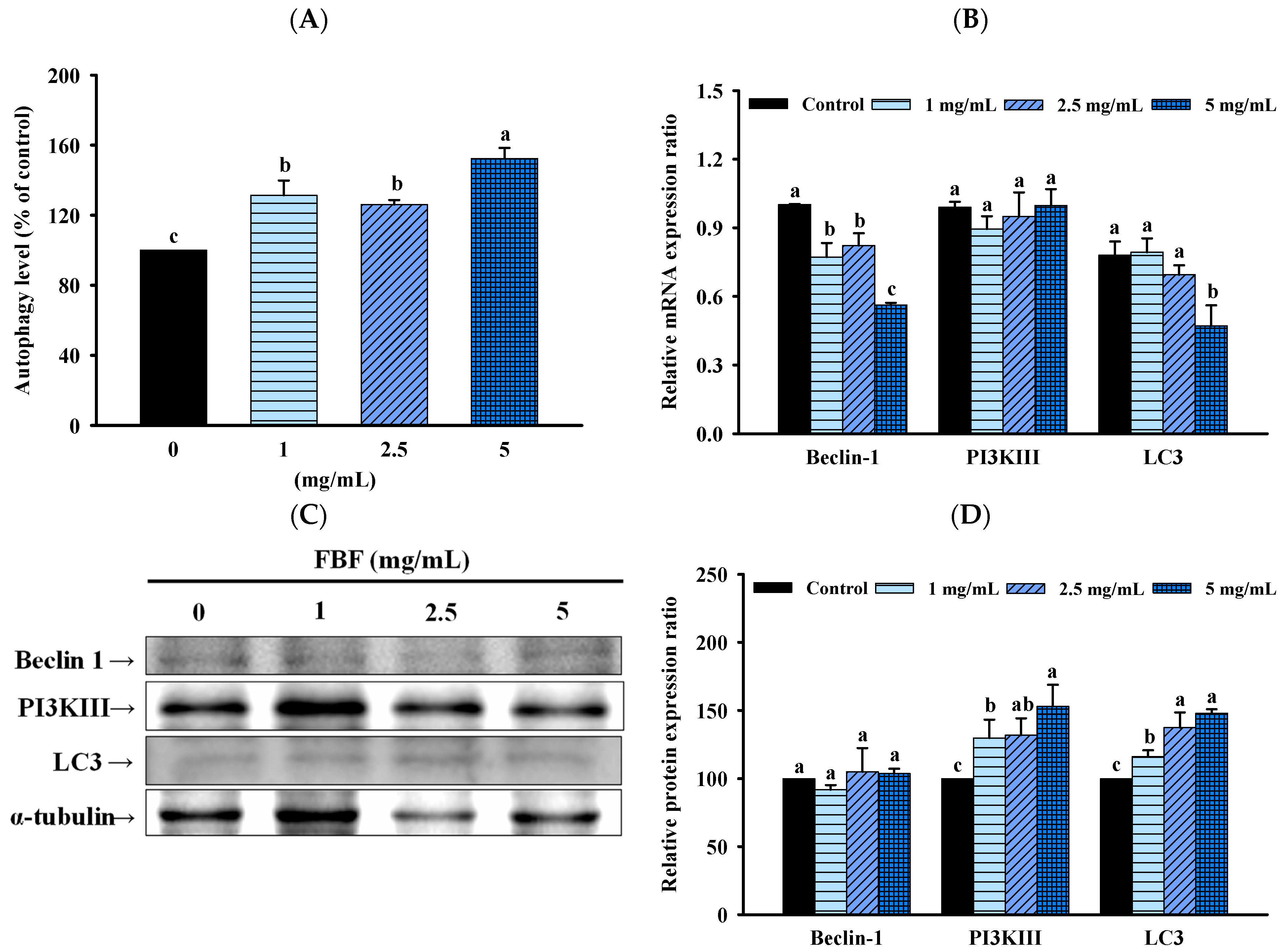
2.4. FBF Inhibits the Growth of HCT-116 Cells by Inducing Autophagy
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Preparation of FBF
4.3. Cell Culture and Treatment
4.4. Cell Viability Analysis
4.5. Cell Cycle Distribution Analysis
4.6. Apoptosis Percentage Analysis
4.7. Autophagy Percentage Analysis
4.8. Relative mRNA Expression Analysis
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Obenauf, A.C.; Zou, Y.; Ji, A.L.; Vanharanta, S.; Shu, W.; Shi, H.; Kong, X.; Bosenberg, M.C.; Wiesner, T.; Rosen, N.; et al. Therapy-induced tumour secretomes promote resistance and tumour progression. Nature 2015, 520, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Mun, J.G.; Jeon, H.D.; Kee, J.Y.; Hong, S.H. Betulin inhibits lung metastasis by inducing cell cycle arrest, autophagy, and apoptosis of metastatic colorectal cancer cells. Nutrients 2020, 12, 66. [Google Scholar] [CrossRef]
- Coates, J.M.; Galante, J.M.; Bold, R.J. Cancer therapy beyond apoptosis: Autophagy and anoikis as mechanisms of cell death. J. Surg Res. 2010, 164, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Dhanalakshmi, S.; Agarwal, R. Phytochemicals as cell cycle modulators a less toxic approach in halting human cancers. Cell Cycle 2002, 1, 155–160. [Google Scholar] [CrossRef]
- Hartwell, L.H.; Kastan, M.B. Cell cycle control and cancer. Science 1994, 266, 1821–1828. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Takada, Y.; Oommen, O.V. From chemoprevention to chemotherapy: Common targets and common goals. Expert Opin. Investig. Drugs 2004, 13, 1327–1338. [Google Scholar] [CrossRef]
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316–323. [Google Scholar] [CrossRef]
- Jain, M.V.; Paczulla, A.M.; Klonisch, T.; Dimgba, F.N.; Rao, S.B.; Roberg, K.; Schweizer, F.; Lengerke, C.; Davoodpour, P.; Palicharla, V.R.; et al. Interconnections between apoptotic, autophagic and necrotic pathways: Implications for cancer therapy development. J. Cell Mol. Med. 2013, 17, 12–29. [Google Scholar] [CrossRef]
- Menon, M.B.; Dhamija, S. Beclin 1 phosphorylation-at the center of autophagy regulation. Front. Cell Dev. Biol. 2018, 6, 137. [Google Scholar] [CrossRef]
- Yun, C.W.; Lee, S.H. The roles of autophagy in cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef]
- Tasdemir, S.S.; Sanlier, N. An insight into the anticancer effects of fermented foods: A review. J. Funct. Foods 2020, 75, 104281. [Google Scholar] [CrossRef]
- Behera, S.S.; Bal, P.; Das, S.; Panda, S.H.; Mohanty, N. Advances in Microbial Fermentation and Fermented Food for Health. In Innovations in Technologies for Fermented Food and Beverage Industries. Food Microbiology and Food Safety; Panda, S., Shetty, P., Eds.; Springer: Cham, Switzerland, 2018; pp. 53–70. [Google Scholar] [CrossRef]
- Lee, C.I.; Hsieh, S.L.; Wu, C.C.; Lee, C.L.; Chang, Y.P.; Wang, J.J. Proteomic analysis of human breast cancer cells treated with Monascus-fermented red mold rice extracts. J. Food Nutr. Res. 2015, 3, 325–329. [Google Scholar] [CrossRef]
- Zeng, H.; Qin, L.; Liu, X.; Miao, S. Increases of lipophilic antioxidants and anticancer activity of coix seed fermented by Monascus purpureus. Foods 2021, 10, 566. [Google Scholar] [CrossRef]
- Chang, W.T.; Chuang, C.H.; Lee, W.J.; Huang, C.S. Extract of Monascus purpureus CWT715 fermented from sorghum liquor biowaste inhibits migration and invasion of SK-Hep-1 human hepatocarcinoma cells. Molecules 2016, 21, 1691. [Google Scholar] [CrossRef]
- Chen, Y.T.; Hsieh, S.L.; Gao, W.S.; Yin, L.J.; Dong, C.D.; Chen, C.W.; Singhania, R.R.; Hsieh, S.; Chen, S.J. Evaluation of chemical compositions, antioxidant capacity and intracellular antioxidant action in fish bone fermented with Monascus purpureus. Molecules 2021, 26, 5288. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Chen, S.J.; Yin, L.J.; Hu, C.Y.; Dong, C.D.; Singhania, R.R.; Hsieh, S.L. Anti-inflammatory effects of fish bone fermented using Monascus purpureus in LPS-induced RAW264. 7 cells by regulating NF-κB pathway. J. Food Sci. Technol. 2022, 60, 958–965. [Google Scholar] [CrossRef]
- Liu, J.; Tan, F.; Liu, X.; Yi, R.; Zhao, X. Exploring the antioxidant effects and periodic regulation of cancer cells by polyphenols produced by the fermentation of grape skin by Lactobacillus plantarum KFY02. Biomolecules 2019, 9, 575. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tan, F.; Liu, X.; Yi, R.; Zhao, X. Grape skin fermentation by Lactobacillus fermentum CQPC04 has anti-oxidative effects on human embryonic kidney cells and apoptosis-promoting effects on human hepatoma cells. RSC Adv. 2020, 10, 4607–4620. [Google Scholar] [CrossRef]
- Rashad, M.M.; Mahmoud, A.E.; Ali, M.M.; Nooman, M.U.; Al-Kashef, A.S. Antioxidant and anticancer agents produced from pineapple waste by solid state fermentation. Int. J. Toxicol. Pharmacol. Res. 2015, 7, 287–296. [Google Scholar]
- Nakayama, K.I.; Nakayama, K. Ubiquitin Ligases: Cell-Cycle Control and Cancer. Nat. Rev. Cancer 2006, 6, 369. [Google Scholar] [CrossRef]
- Wang, Y.; Ji, P.; Liu, J.; Broaddus, R.R.; Xue, F.; Zhang, W. Centrosome-associated regulators of the G 2/M checkpoint as targets for cancer therapy. Mol. Cancer 2009, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Luo, Y.; Xia, Y.; Zhang, X.; Qin, Y.; Liu, W.; Li, M.; Liu, X.; Zheng, Q.; Li, D. Cinobufagin induces cell cycle arrest at the S phase and promotes apoptosis in nasopharyngeal carcinoma cells. Biomed. Pharmacother. 2020, 122, 109763. [Google Scholar] [CrossRef]
- Sun, N.X.; Liu, H.P.; Liu, S.J.; Zhang, X.Y.; Chen, P.; Li, W.H.; Xu, X.X.; Tian, W.T. Purification, preliminary structure and antitumor activity of exopolysaccharide produced by Streptococcus thermophilus CH9. Molecules 2018, 23, 2898. [Google Scholar] [CrossRef]
- Zhang, L.; Zheng, Y.; Deng, H.; Liang, L.; Peng, J. Aloperine induces G2/M phase cell cycle arrest and apoptosis in HCT116 human colon cancer cells. Int. J. Mol. Med. 2014, 33, 1613–1620. [Google Scholar] [CrossRef]
- Ma, L.; Xu, G.B.; Tang, X.; Zhang, C.; Zhao, W.; Wang, J.; Chen, H. Anti-cancer potential of polysaccharide extracted from hawthorn (Crataegus.) on human colon cancer cell line HCT116 via cell cycle arrest and apoptosis. J. Funct. Foods 2020, 64, 103677. [Google Scholar] [CrossRef]
- Priyadarsini, R.V.; Murugan, R.S.; Maitreyi, S.; Ramalingam, K.; Karunagaran, D.; Nagini, S. The flavonoid quercetin induces cell cycle arrest and mitochondria-mediated apoptosis in human cervical cancer (HeLa) cells through p53 induction and NF-κB inhibition. Eur. J. Pharmacol. 2010, 649, 84–91. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Y.S.; Thakur, K.; Hussain, S.S.; Zhang, J.G.; Xiao, G.R.; Wei, Z.J. Licochalcone A from licorice root, an inhibitor of human hepatoma cell growth via induction of cell apoptosis and cell cycle arrest. Food Chem. Toxicol. 2018, 120, 407–417. [Google Scholar] [CrossRef]
- Wu, L.; Wang, L.; Tian, X.; Zhang, J.; Feng, H. Germacrone exerts anti-cancer effects on gastric cancer through induction of cell cycle arrest and promotion of apoptosis. BMC Complement Med. Ther. 2020, 20, 21. [Google Scholar] [CrossRef] [PubMed]
- Kesika, P.; Sivamaruthi, B.S.; Chaiyasut, C. Health promoting effects of fermented foods against cancer: An updated concise review. Food Sci. Technol. 2020, 42, e18220. [Google Scholar] [CrossRef]
- Hawk, E.T.; Levin, B. Colorectal cancer prevention. J. Clin. Oncol. 2005, 23, 378–391. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, C.M.; Singh, A.T. Apoptosis: A target for anticancer therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed]
- van Loo, G.; Saelens, X.; Van Gurp, M.; MacFarlane, M.; Martin, S.J.; Vandenabeele, P. The role of mitochondrial factors in apoptosis: A Russian roulette with more than one bullet. Cell Death Differ. 2002, 9, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef]
- Koh, E.M.; Lee, E.K.; Song, J.; Kim, S.J.; Song, C.H.; Seo, Y.; Chae, C.H.; Jung, K.J. Anticancer activity and mechanism of action of fermented wheat germ extract against ovarian cancer. J. Food Biochem. 2018, 42, e12688. [Google Scholar] [CrossRef]
- Khan, I.; Kang, S.C. Apoptotic activity of Lactobacillus plantarum DGK-17-fermented soybean seed extract in human colon cancer cells via ROS-JNK signaling pathway. J. Food Sci. 2017, 82, 1475–1483. [Google Scholar] [CrossRef]
- Otto, C.; Hahlbrock, T.; Eich, K.; Karaaslan, F.; Jürgens, C.; Germer, C.T.; Wiegering, A.; Kämmerer, U. Antiproliferative and antimetabolic effects behind the anticancer property of fermented wheat germ extract. BMC Complement Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef]
- Ryu, J.Y.; Kang, H.R.; Cho, S.K. Changes over the fermentation period in phenolic compounds and antioxidant and anticancer activities of blueberries fermented by Lactobacillus plantarum. J. Food Sci. 2019, 84, 2347–2356. [Google Scholar] [CrossRef]
- Michalak, M.; Szwajgier, D.; Paduch, R.; Kukula-Koch, W.; Waśko, A.; Polak-Berecka, M. Fermented curly kale as a new source of gentisic and salicylic acids with antitumor potential. J. Funct. Foods 2020, 67, 103866. [Google Scholar] [CrossRef]
- Khamphio, M.; Barusrux, S.; Weerapreeyakul, N. Sesamol induces mitochondrial apoptosis pathway in HCT116 human colon cancer cells via pro-oxidant effect. Life Sci. 2016, 158, 46–56. [Google Scholar] [CrossRef]
- Li, G.; Kong, B.; Tong, Q.; Li, Y.; Chen, L.; Zeng, J.; Yu, H.; Xie, X.; Zhang, J. Vanillin downregulates NNMT and attenuates NNMT-related resistance to 5-fluorouracil via ROS-induced cell apoptosis in colorectal cancer cells. Oncol. Rep. 2021, 45, 1–15. [Google Scholar] [CrossRef]
- El-Ella, D.M.A. Autophagy/apoptosis induced by geraniol through HIF-1α/BNIP3/Beclin-1 signaling pathway in A549 CoCl2 treated cells. Adv. Pharm. Bull. 2022, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Chiangjong, W.; Chutipongtanate, S.; Hongeng, S. Anticancer peptide: Physicochemical property, functional aspect and trend in clinical application. Int. J. Oncol. 2020, 57, 678–696. [Google Scholar] [CrossRef] [PubMed]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival: Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Ramdall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer sequence (5′-3′) | Reference (NCBI GenBank) |
|---|---|---|
| β-actin | F-ATGTGCAAGGCCGGCTTC R-GAATCCTTCTGACCCATGCC | NM_001101.3 |
| Bax | F-TGTTTTCTGACGGCAACTTCA R-AGCCCATGATGGTTCTGATCA | NM_001291428.1 |
| Bcl-2 | F-CCTGTGGATGACTGAGTACCTGAAC R-CAGCCAGGAGAAATCAAACAGA | NM_000633.2 |
| Caspase 8 | F-TCCAAATGCAAACTGGATGATG R-TTTTCAGGATGTCCAACTTTCCTT | NM_001080124.1 |
| Caspase 9 | F-AGCTGGACGCCATATCTAGTTTG R-AACGTACCAGGAGCCACTCTTG | NM_001229.4 |
| PI3K III | F-TTGGAGACAGGCACCTGGAT R-CCATTTCTTTATTCAGCTTCATTGG | NM_001308020.1 |
| Beclin 1 | F-CTGGACACGAGTTTCAAGATCCT R-GTTAGTCTCTTCCTCCTGGGTCTCT | NM_001313998.1 |
| LC3 | F-TCCTGGACAAGACCAAGTTTTTG R-ACCATGCTGTGCTGGTTCAC | NM_032514.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-T.; Chen, S.-J.; Hu, C.-Y.; Dong, C.-D.; Chen, C.-W.; Singhania, R.R.; Hsieh, S.-L. Exploring the Anti-Cancer Effects of Fish Bone Fermented Using Monascus purpureus: Induction of Apoptosis and Autophagy in Human Colorectal Cancer Cells. Molecules 2023, 28, 5679. https://doi.org/10.3390/molecules28155679
Chen Y-T, Chen S-J, Hu C-Y, Dong C-D, Chen C-W, Singhania RR, Hsieh S-L. Exploring the Anti-Cancer Effects of Fish Bone Fermented Using Monascus purpureus: Induction of Apoptosis and Autophagy in Human Colorectal Cancer Cells. Molecules. 2023; 28(15):5679. https://doi.org/10.3390/molecules28155679
Chicago/Turabian StyleChen, Ya-Ting, Shu-Jen Chen, Chun-Yi Hu, Cheng-Di Dong, Chiu-Wen Chen, Reeta Rani Singhania, and Shu-Ling Hsieh. 2023. "Exploring the Anti-Cancer Effects of Fish Bone Fermented Using Monascus purpureus: Induction of Apoptosis and Autophagy in Human Colorectal Cancer Cells" Molecules 28, no. 15: 5679. https://doi.org/10.3390/molecules28155679
APA StyleChen, Y.-T., Chen, S.-J., Hu, C.-Y., Dong, C.-D., Chen, C.-W., Singhania, R. R., & Hsieh, S.-L. (2023). Exploring the Anti-Cancer Effects of Fish Bone Fermented Using Monascus purpureus: Induction of Apoptosis and Autophagy in Human Colorectal Cancer Cells. Molecules, 28(15), 5679. https://doi.org/10.3390/molecules28155679