

Hydrolytic Mechanism of a Metalloenzyme Is Modified by the Nature of the Coordinated Metal Ion




Abstract
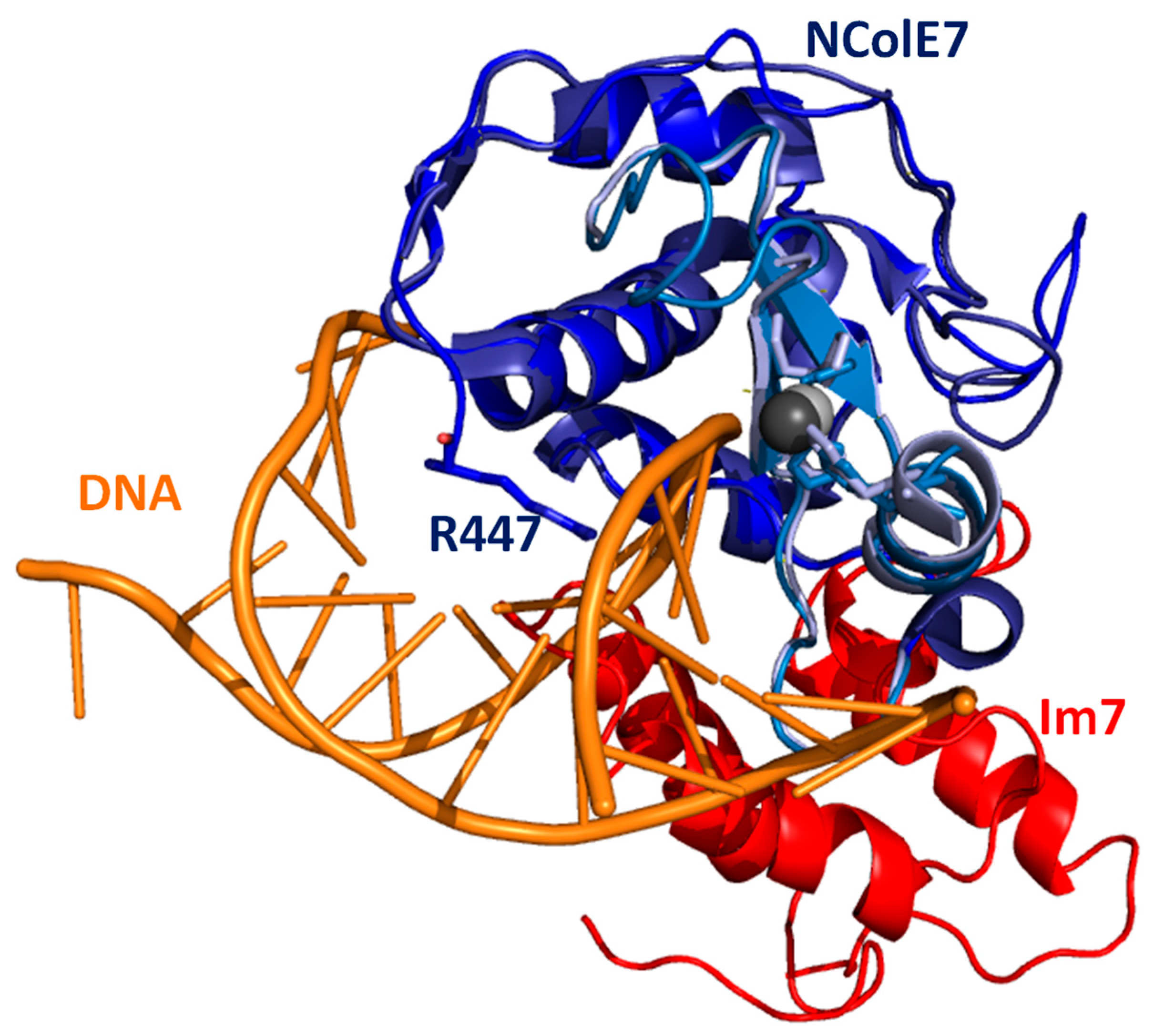
1. Introduction
2. Results and Discussion
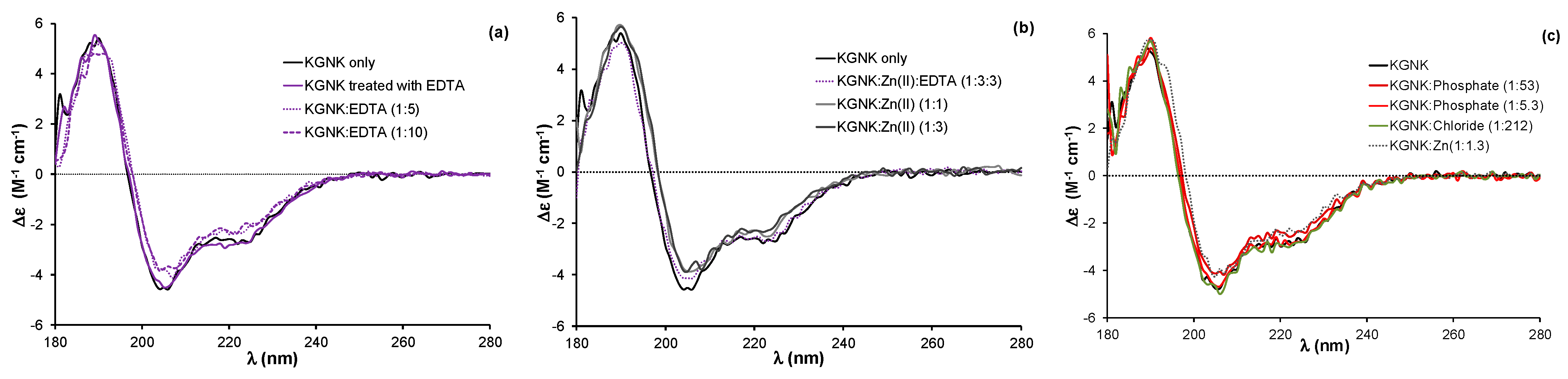
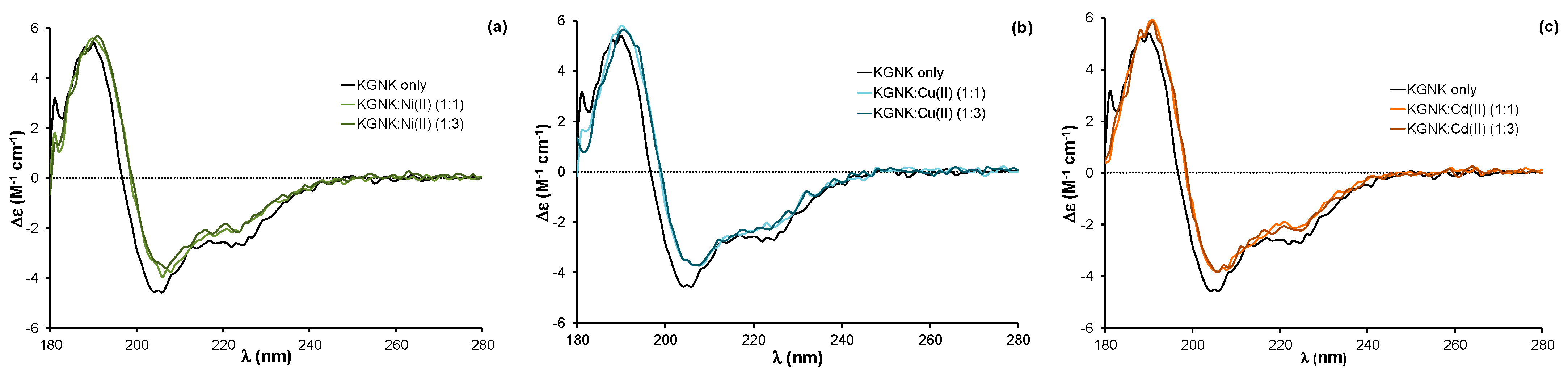
2.1. Structural Aspects of the Interaction of the KGNK Protein with Metal Ions as Observed via Circular Dichroism Spectroscopy
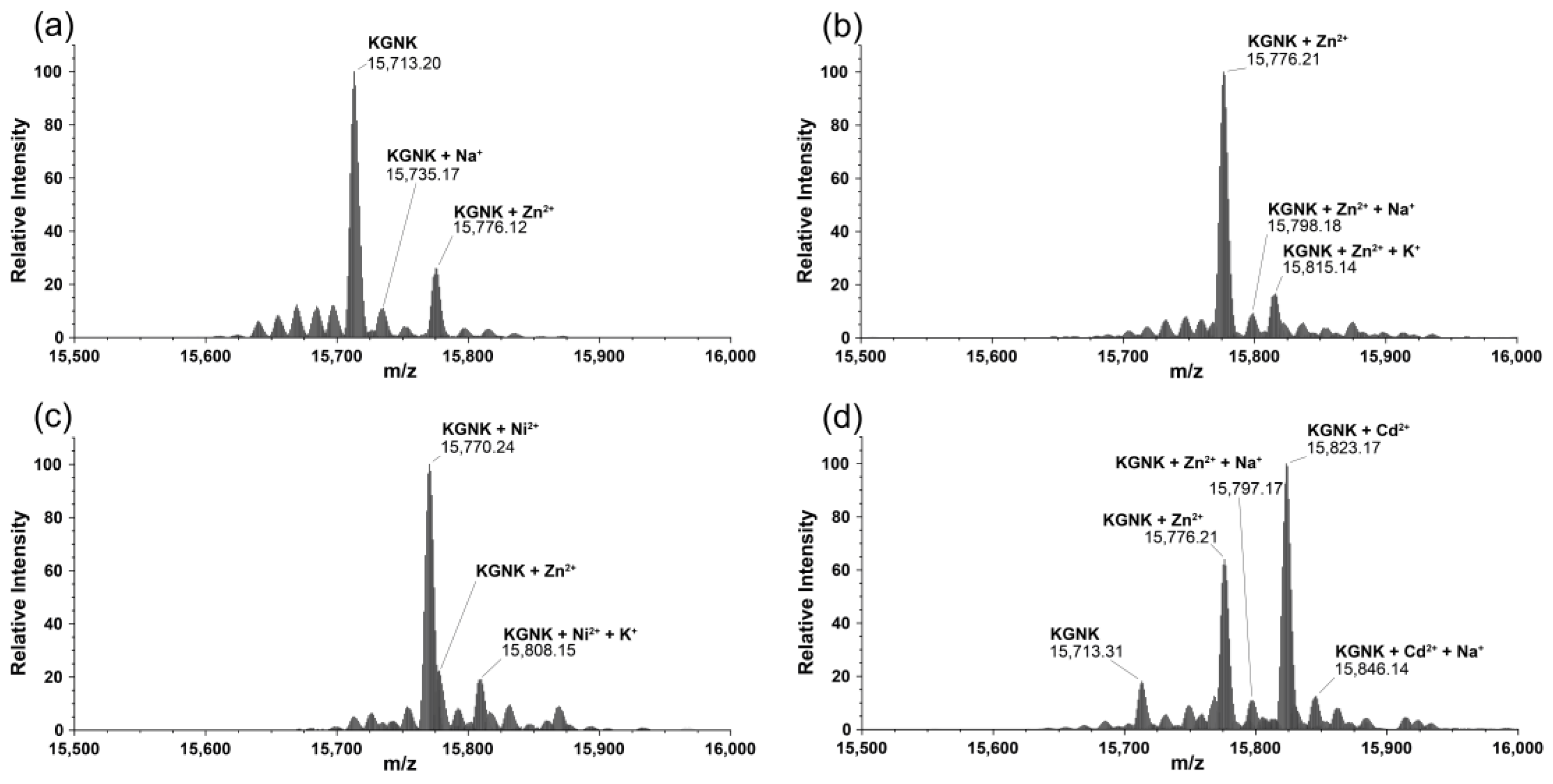
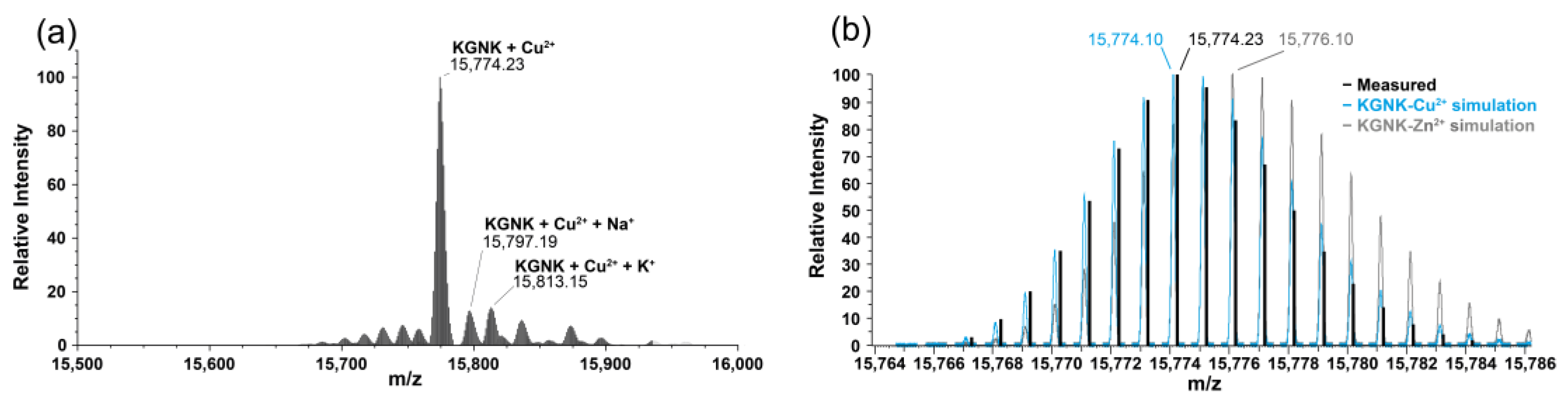
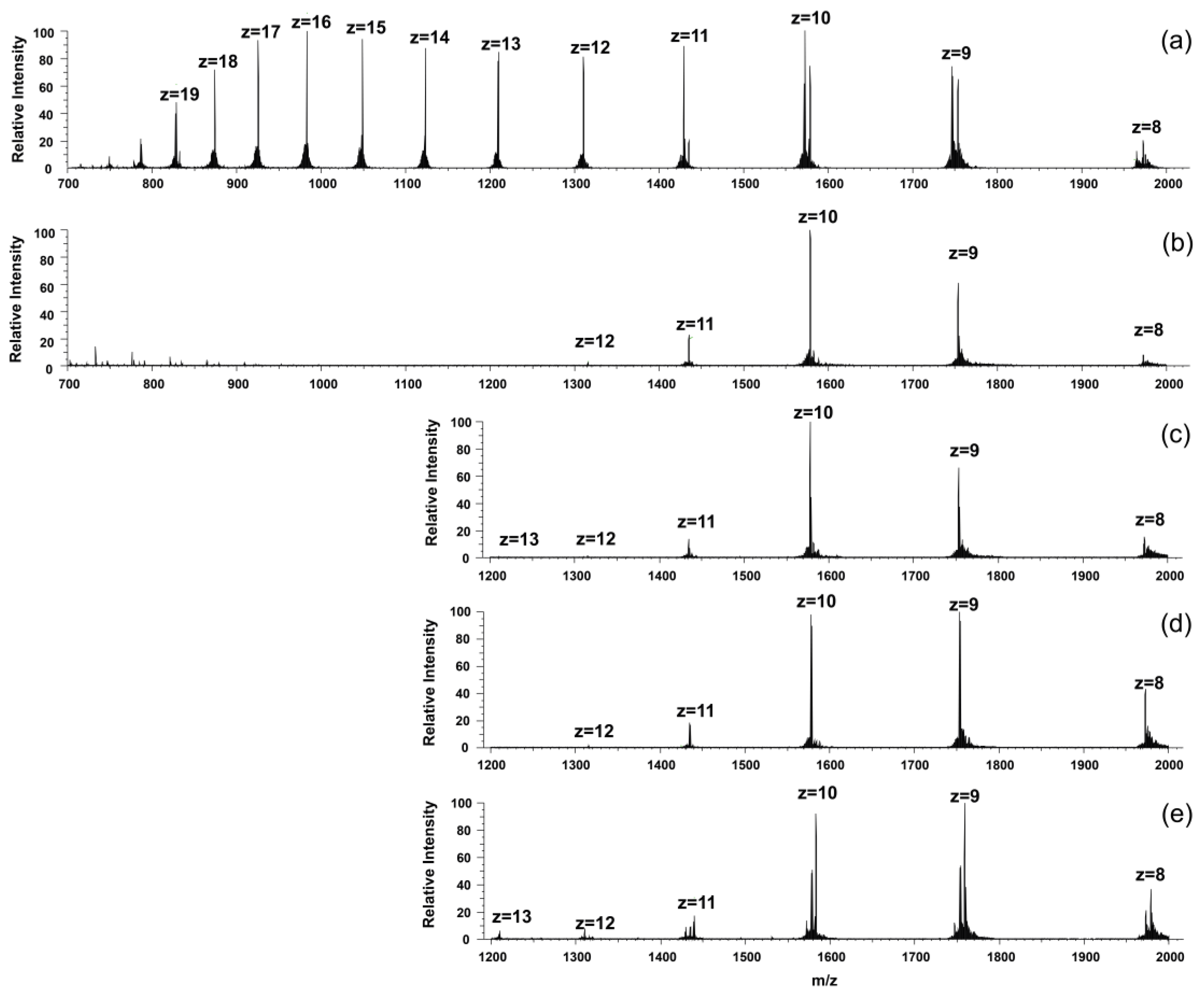
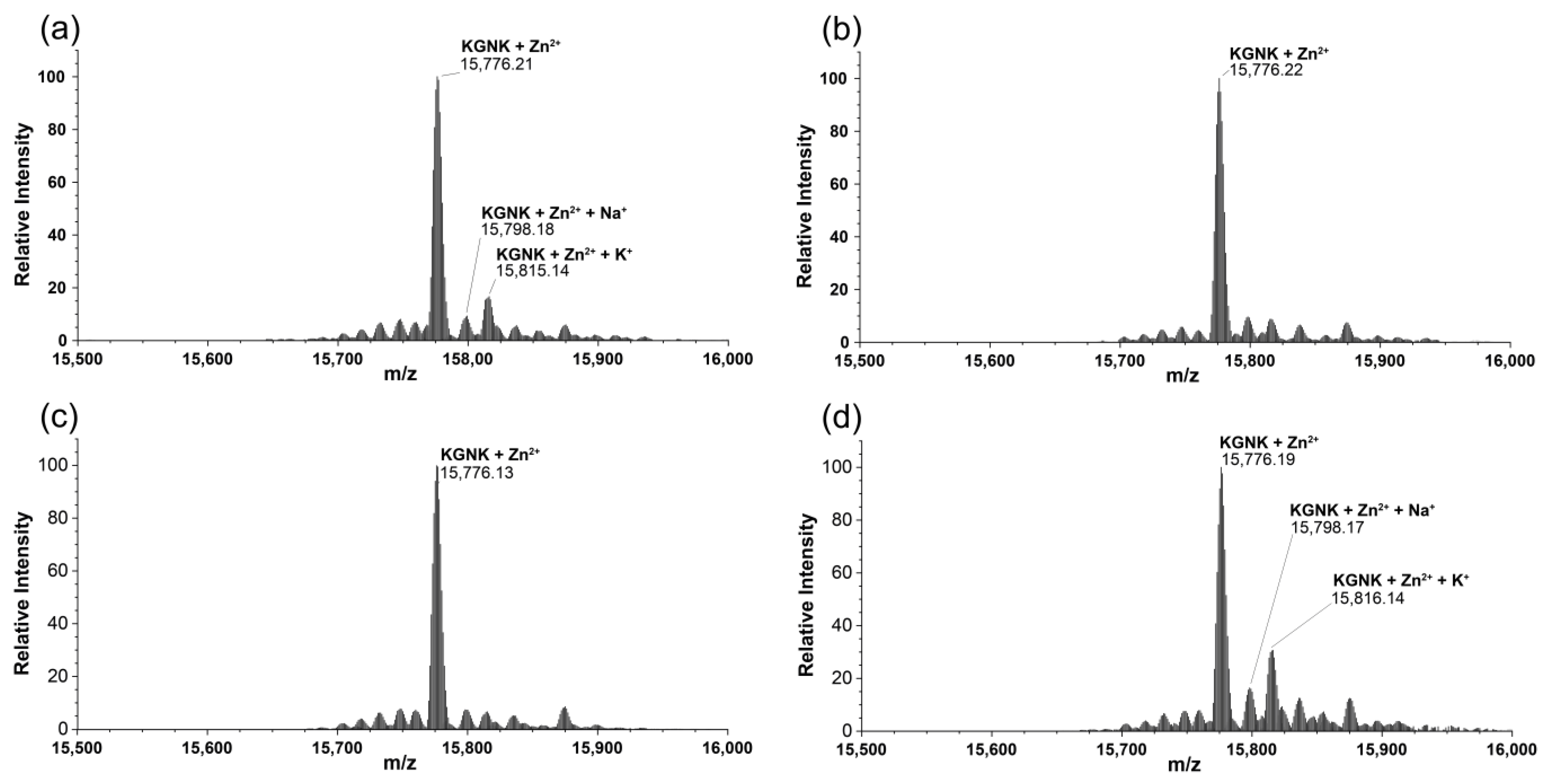
2.2. Mass Spectrometric Monitoring of Metal Ion Binding to KGNK
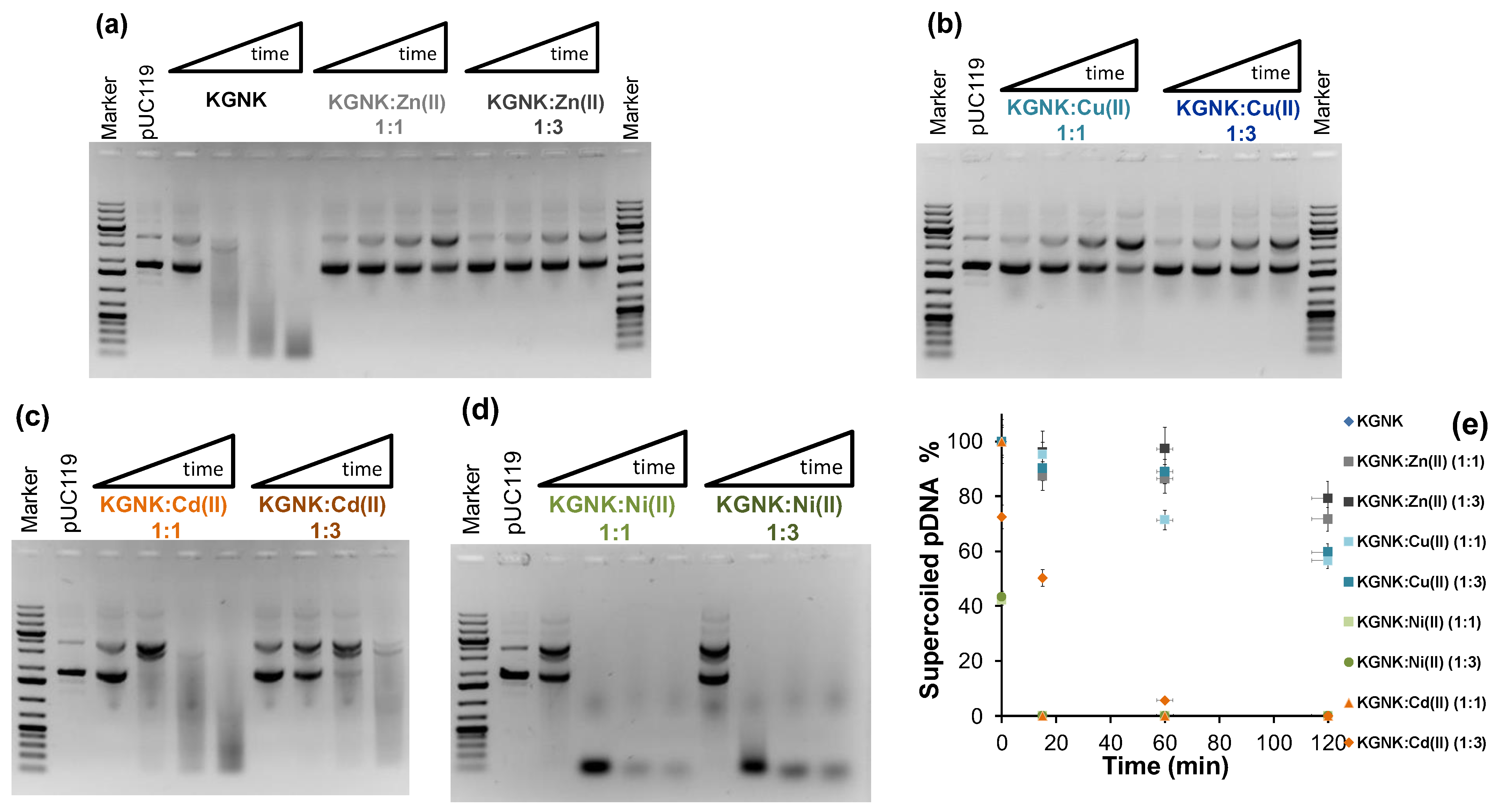
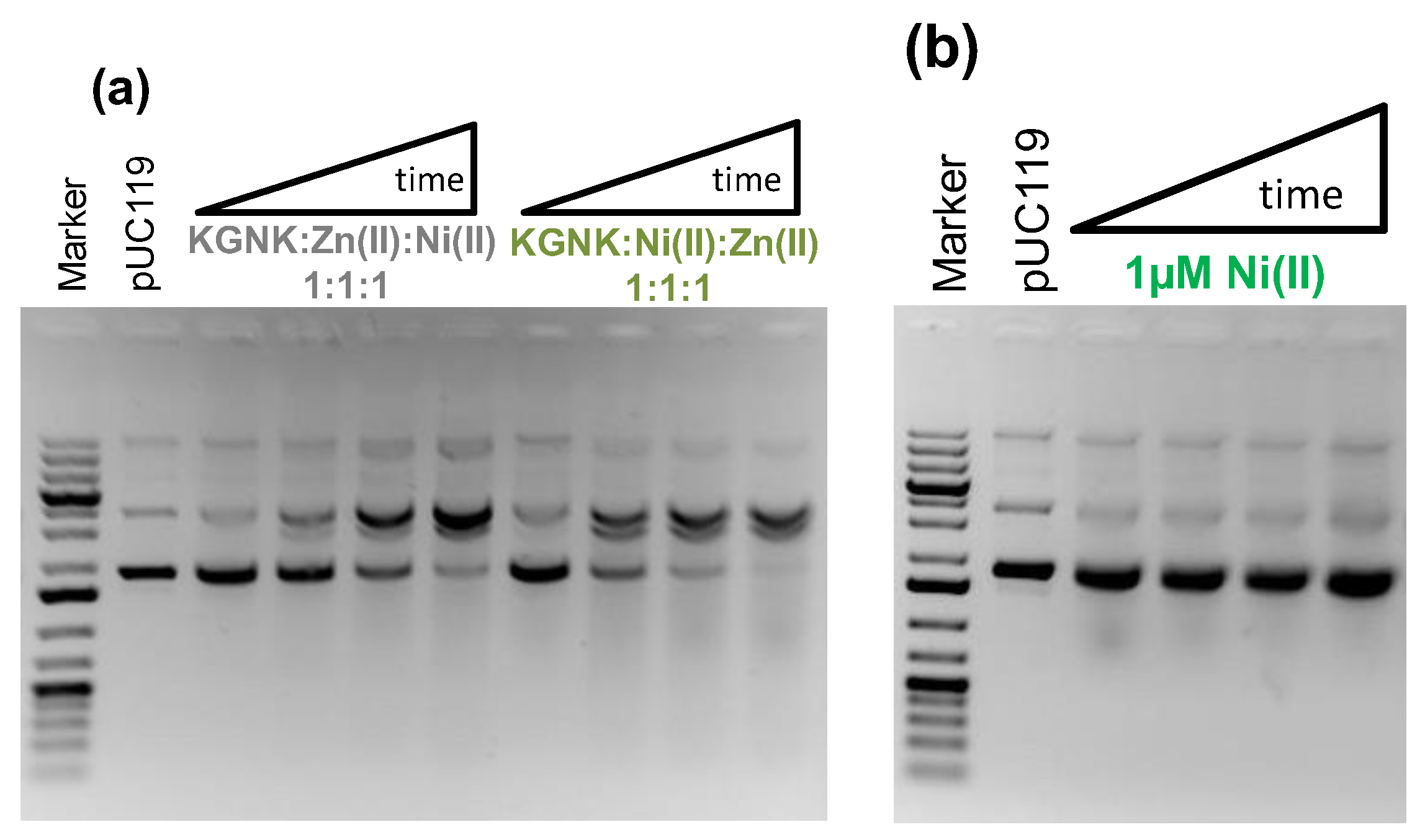
2.3. The Effect of Metal Ions on the Catalytic Activity of KGNK
3. Materials and Methods
3.1. Reagents
3.2. Recombinant NColE7-KGNK Expression and Purification
3.3. Mass Spectrometric Experiments
3.4. Circular Dichroism Spectroscopic Measurements
3.5. Catalytic Activity Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Klein, A.; Wojdyla, J.A.; Joshi, A.; Josts, I.; McCaughey, L.C.; Housden, N.G.; Kaminska, R.; Byron, O.; Walker, D.; Kleanthous, C. Structural and biophysical analysis of nuclease protein antibiotics. Biochem. J. 2016, 473, 2799–2812. [Google Scholar] [CrossRef]
- Marković, K.G.; Grujović, M.Ž.; Koraćević, M.G.; Nikodijević, D.D.; Milutinović, M.G.; Semedo-Lemsaddek, T.; Djilas, M.D. Colicins and Microcins Produced by Enterobacteriaceae: Characterization, Mode of Action, and Putative Applications. Int. J. Environ. Res. Public Health 2022, 19, 11825. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duche, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef]
- Papadakos, G.; Wojdyla, J.A.; Kleanthous, C. Nuclease colicins and their immunity proteins. Q. Rev. Biophys. 2012, 45, 57–103. [Google Scholar] [CrossRef] [PubMed]
- Chak, K.F.; Kuo, W.S.; Lu, F.M.; James, R. Cloning and characterization of the ColE7 plasmid. J. Gen. Microbiol. 1991, 137, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Hsia, K.C.; Li, C.-L.; Yuan, H.S. Structural and functional insight into sugar-nonspecific nucleases in host defense. Curr. Opin. Struct. Biol. 2005, 15, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.C.; Hsiao, K.C.; Liu, Y.W.; Leng, P.H.; Yuen, H.S.; Chak, K.-F. Processing of DNase domain during translocation of colicin E7 across the membrane of Escherichia coli. Biochem. Biophys. Res. Commun. 2001, 284, 556–562. [Google Scholar] [CrossRef]
- Hsia, K.C.; Chak, K.F.; Liang, P.H.; Cheng, Y.S.; Ku, W.Y.; Yuan, H.S. Crystal structures of nuclease-ColE7 complexed with octamer DNA. Structure 2004, 12, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; de Zamaroczy, M. In Vivo Processing of DNase Colicins E2 and E7 Is Required for Their Import into the Cytoplasm of Target Cells. PLoS ONE 2014, 9, e96549. [Google Scholar] [CrossRef]
- Cheng, Y.S.; Hsia, K.C.; Doudeva, L.G.; Chak, K.F.; Yuan, H.S. The crystal structure of the nuclease domain of colicin E7 suggests a mechanism for binding to double-stranded DNA by the H-N-H endonucleases. J. Mol. Biol. 2002, 324, 227–236. [Google Scholar]
- Eastberg, J.H.; Eklund, J. Mutability of an HNH Nuclease Imidazole General Base and Exchange of a Deprotonation Mechanism. Biochemistry 2007, 46, 7215–7225. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yuan, H.S. The Conserved Asparagine in the HNH Motif Serves an Important Structural Role in Metal Finger Endonucleases. J. Mol. Biol. 2007, 368, 812–821. [Google Scholar] [CrossRef] [PubMed]
- Levin, K.B.; Dym, O.; Albeck, S.; Magdassi, S.; Keeble, A.H.; Kleanthous, C.; Tawfik, D.S. Following evolutionary paths to protein-protein interactions with high affinity and selectivity. Nat. Struct. Mol. Biol. 2009, 16, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.; Liao, C.; Ku, W.; Chak, K.; Yuan, H.S. The crystal structure of the DNase domain of colicin E7 in complex with its inhibitor Im7 protein. Structure 1999, 7, 91–102. [Google Scholar] [CrossRef]
- Sui, M.; Tsai, L.; Hsia, K.; Doudeva, L.; Ku, W.; Han, G.; Yuan, H. Metal ions and phosphate binding in the H-N-H motif: Crystal structures of the nuclease domain of ColE7/Im7 in complex with a phosphate ion and different divalent metal ions. Protein Sci. 2002, 11, 2947–2957. [Google Scholar] [CrossRef]
- Czene, A.; Tóth, E.; Németh, E.; Otten, H.; Poulsen, J.N.; Christensen, H.E.M.; Rulíšek, L.; Nagata, K.; Larsen, S.; Gyurcsik, B. A new insight into the zinc-dependent DNA-cleavage by the colicin E7 nuclease: A crystallographic and computational study. Metallomics 2014, 6, 2090–2099. [Google Scholar] [CrossRef]
- Doudeva, L.; Huang, D.; Hsia, K.; Shi, Z.; Li, C.; Shen, Y.; Cheng, Y.; Yuan, H. Crystal structural analysis and metal-dependent stability and activity studies of the ColE7 endonuclease domain in complex with DNA/Zn2+ or inhibitor/Ni2+. Protein Sci. 2006, 15, 269–280. [Google Scholar]
- Wang, Y.; Yang, W.; Li, C.; Doudeva, L.G.; Yuan, H.S. Structural basis for sequence-dependent DNA cleavage by nonspecific endonucleases. Nucleic Acids Res. 2007, 35, 584–594. [Google Scholar] [CrossRef]
- Czene, A.; Németh, E.; Zóka, I.G.; Jakab-Simon, N.I.; Körtvélyesi, T.; Nagata, K.; Christensen, H.E.M.; Gyurcsik, B. The role of the N-terminal loop in the function of the colicin E7 nuclease domain. J. Biol. Inorg. Chem. 2013, 18, 309–321. [Google Scholar]
- Németh, E.; Körtvélyesi, T.; Thulstrup, P.W.; Christensen, H.E.M.; Kožíšek, M.; Nagata, K.; Czene, A.; Gyurcsik, B. Fine tuning of the catalytic activity of colicin E7 nuclease domain by systematic N-terminal mutations. Protein Sci. 2014, 23, 1113–1122. [Google Scholar] [CrossRef]
- Ku, W.; Liu, Y.; Hsu, Y.; Liao, C.; Liang, P.; Yuan, H.; Chak, K. The zinc ion in the HNH motif of the endonuclease domain of colicin E7 is not required for DNA binding but is essential for DNA hydrolysis. Nucleic Acids Res. 2002, 30, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Németh, E.; Körtvélyesi, T.; Kožíšek, M.; Thulstrup, P.W.; Christensen, H.E.M.; Asaka, M.N.; Nagata, K.; Gyurcsik, B. Substrate binding activates the designed triple mutant of the colicin E7 metallonuclease. J. Biol. Inorg. Chem. 2014, 19, 12951303. [Google Scholar] [CrossRef] [PubMed]
- Németh, E.; Kožíšek, M.; Schilli, G.K.; Gyurcsik, B. Preorganization of the catalytic Zn2+ binding site in the HNH nuclease motif-A solution study. J. Inorg. Biochem. 2015, 151, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Németh, E.; Balogh, R.K.; Borsos, K.; Czene, A.; Thulstrup, P.W.; Gyurcsik, B. Intrinsic protein disorder could be overlooked in cocrystallization conditions: An SRCD case study. Protein Sci. 2016, 25, 1977–1988. [Google Scholar] [CrossRef]
- Shi, Z.; Chak, K.F.; Yuan, H.S. Identification of an essential cleavage site in ColE7 required for import and killing of cells. J. Biol. Chem. 2005, 280, 24663–24668. [Google Scholar] [CrossRef]
- Shlyapnikov, S.V.; Lunin, V.V.; Perbandt, M.; Polyakov, K.M.; Lunin, V.Y.; Levdikov, V.M.; Betzel, C.; Mikhailov, A.M. Atomic structure of the Serratia marcescens endonuclease at 1.1 A resolution and the enzyme reaction mechanism. Acta Crystallogr. D Biol. Crystallogr. 2000, 56, 567–572. [Google Scholar] [CrossRef]
- Pommer, A.J.; Kuhlmann, U.C.; Cooper, A.; Hemmings, A.M.; Moore, G.R.; James, R.; Kleanthous, C. Homing in on the role of transition metals in the HNH motif of colicin endonuclease. J. Biol. Chem. 1999, 274, 27153–27160. [Google Scholar] [CrossRef]
- van den Bremer, E.T.; Jiskoot, W.; James, R.; Moore, G.R.; Kleanthous, C.; Heck, A.J.; Maier, C.S. Probing metal ion binding and conformational properties of the colicin E9 endonuclease by electrospray ionization time-of-flight mass spectrometry. Protein Sci. 2002, 11, 1738–1752. [Google Scholar] [CrossRef]
- van den Bremer, E.T.; Keeble, A.H.; Jiskoot, W.; Spelbrink, R.E.; Maier, C.S.; van Hoek, A.; Visser, A.J.; James, R.; Moore, G.R.; Kleanthous, C.; et al. Distinct conformational stability and functional activity of four highly homologous endonuclease colicins. Protein Sci. 2004, 13, 1391–1401. [Google Scholar] [CrossRef]
- Grant, S.G.N.; Jessee, J.; Bloom, F.R.; Hanahan, D. Differential plasmid rescue from transgenic mouse DNAs into Escherichia coli methylation-restriction mutants. Proc. Natl. Acad. Sci. USA 1990, 87, 4645–4649. [Google Scholar] [CrossRef]
- Studier, F.W.; Rosenberg, A.H.; Dunn, J.J.; Dubendorff, J.W. Use of T7 RNA polymerase to direct expression of cloned genes. Methods Enzymol. 1990, 185, 60–89. [Google Scholar] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the Expasy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Lagunavicius, A.; Sasnauskas, G.; Halford, S.E.; Siksnys, V. The metal-independent type IIs restriction enzyme BfiI is a dimer that binds two DNA sites but has only one catalytic centre. J. Mol. Biol. 2003, 326, 1051–1064. [Google Scholar] [CrossRef] [PubMed]
- Pohlman, R.F.; Liu, F.; Wang, L.; More, M.I.; Winans, S.C. Genetic and biochemical analysis of an endonuclease encoded by the IncN plasmid pKM101. Nucleic Acids Res. 1993, 21, 4867–4872. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
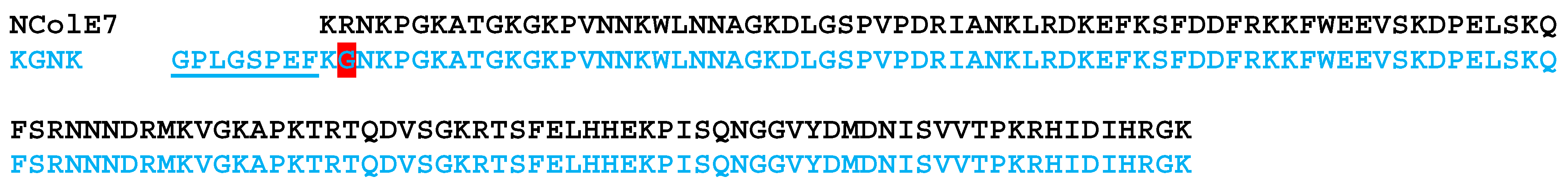{kind=link}
| Species | Formula | Calculated Mass of MH+ (Da) | Measured Mass of MH+ (Da) |
|---|---|---|---|
| KGNK apo | C690H1101N211O206S2 | 15,713.19 | 15,713.20 |
| KGNK + 1Zn2+–2H+ | C690H1099N211O206S2Zn | 15,776.10 | 15,776.12 |
| KGNK + 1Ni2+–2H+ | C690H1099N211O206S2Ni | 15,770.11 | 15,770.24 |
| KGNK + 1Cu2+–2H+ | C690H1099N211O206S2Cu | 15,774.10 | 15,774.23 |
| KGNK + 1Cd2+–2H+ | C690H1099N211O206S2Cd | 15,823.08 | 15,823.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nafaee, Z.H.; Hajdu, B.; Hunyadi-Gulyás, É.; Gyurcsik, B. Hydrolytic Mechanism of a Metalloenzyme Is Modified by the Nature of the Coordinated Metal Ion. Molecules 2023, 28, 5511. https://doi.org/10.3390/molecules28145511
Nafaee ZH, Hajdu B, Hunyadi-Gulyás É, Gyurcsik B. Hydrolytic Mechanism of a Metalloenzyme Is Modified by the Nature of the Coordinated Metal Ion. Molecules. 2023; 28(14):5511. https://doi.org/10.3390/molecules28145511
Chicago/Turabian StyleNafaee, Zeyad H., Bálint Hajdu, Éva Hunyadi-Gulyás, and Béla Gyurcsik. 2023. "Hydrolytic Mechanism of a Metalloenzyme Is Modified by the Nature of the Coordinated Metal Ion" Molecules 28, no. 14: 5511. https://doi.org/10.3390/molecules28145511
APA StyleNafaee, Z. H., Hajdu, B., Hunyadi-Gulyás, É., & Gyurcsik, B. (2023). Hydrolytic Mechanism of a Metalloenzyme Is Modified by the Nature of the Coordinated Metal Ion. Molecules, 28(14), 5511. https://doi.org/10.3390/molecules28145511