

Biochemical, Biophysical, and Structural Analysis of an Unusual DyP from the Extremophile Deinococcus radiodurans

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
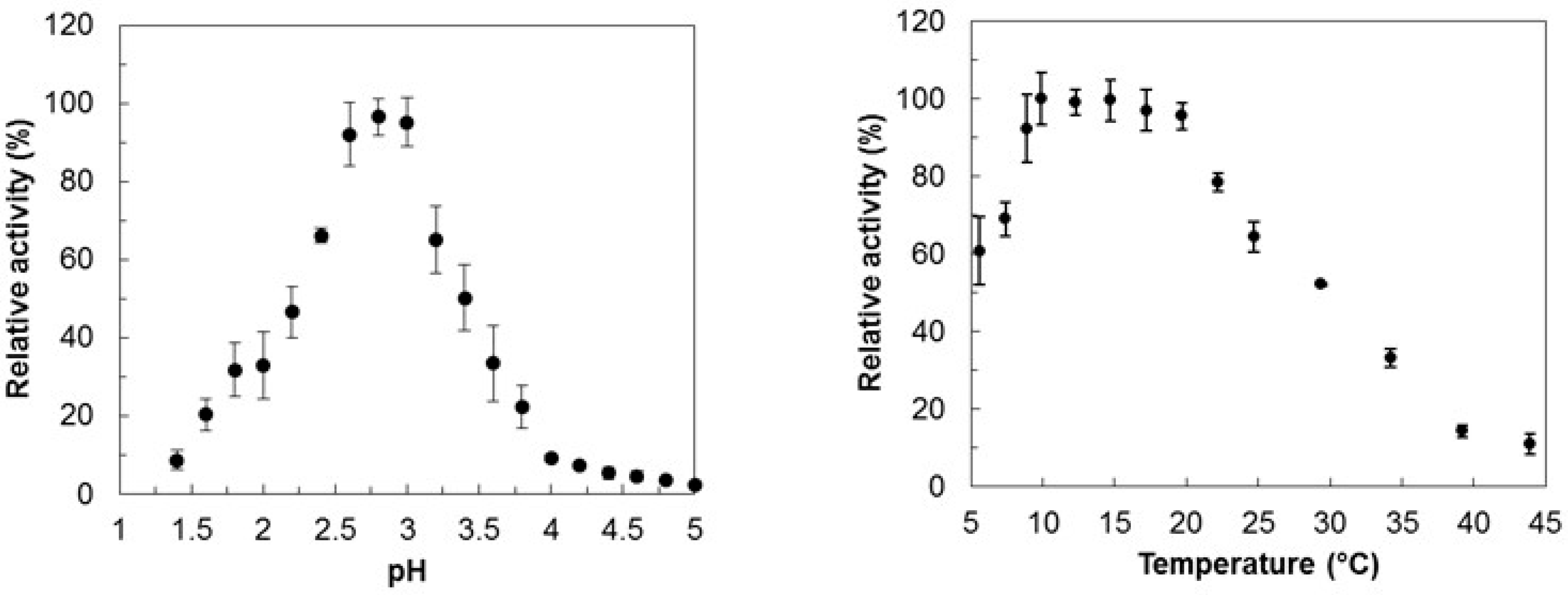
2.1. Sequence and Activity Analysis
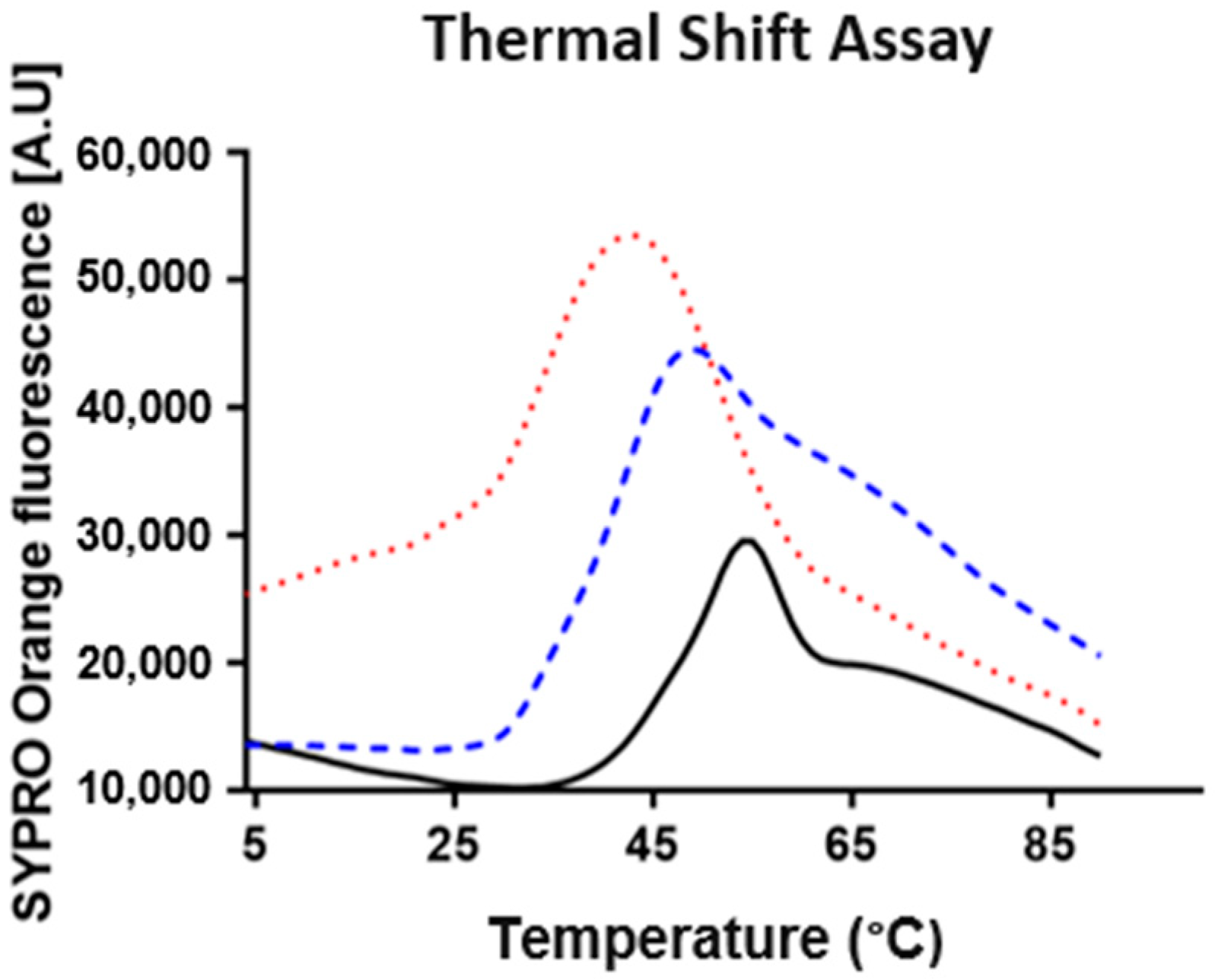
2.2. Buffer-Stability Analysis
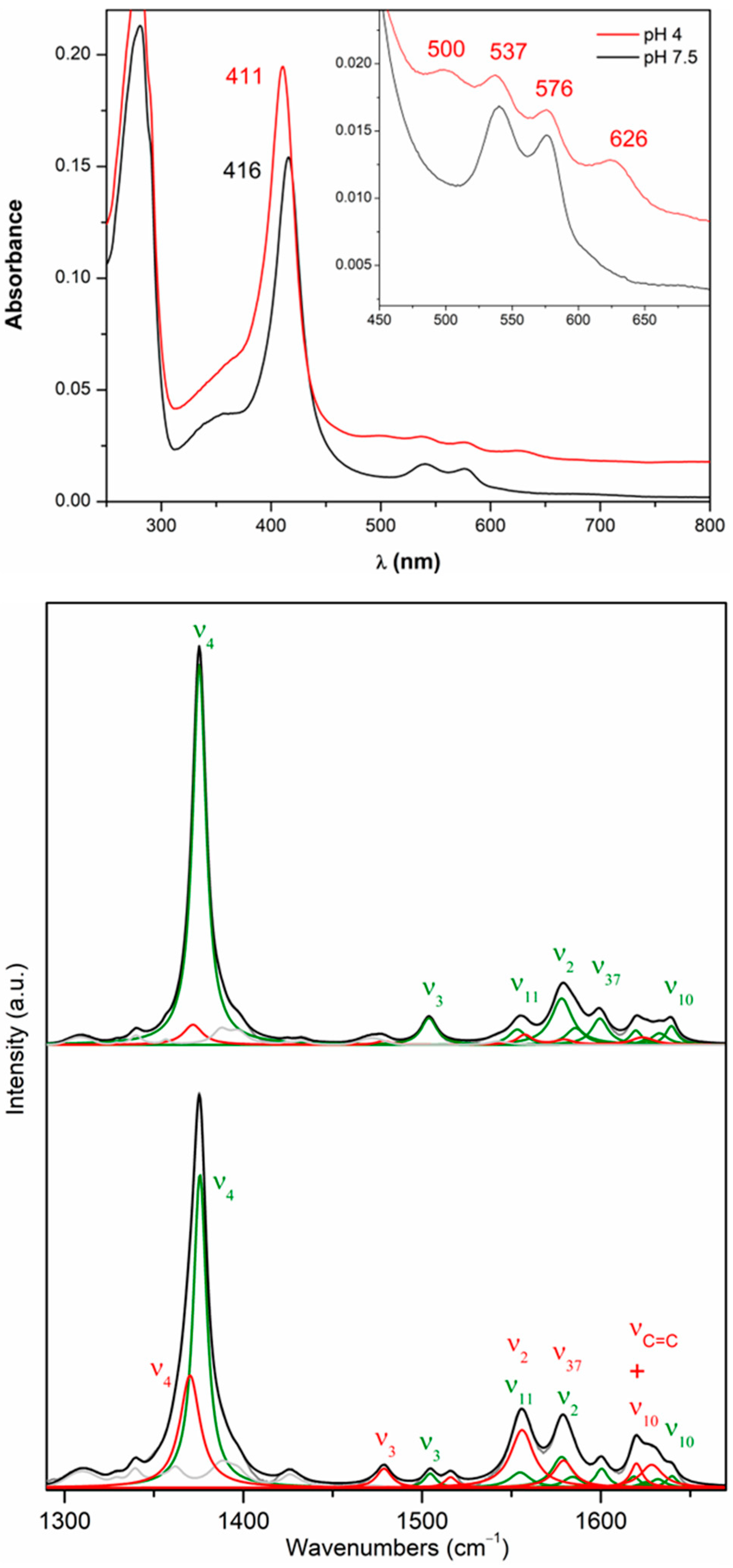
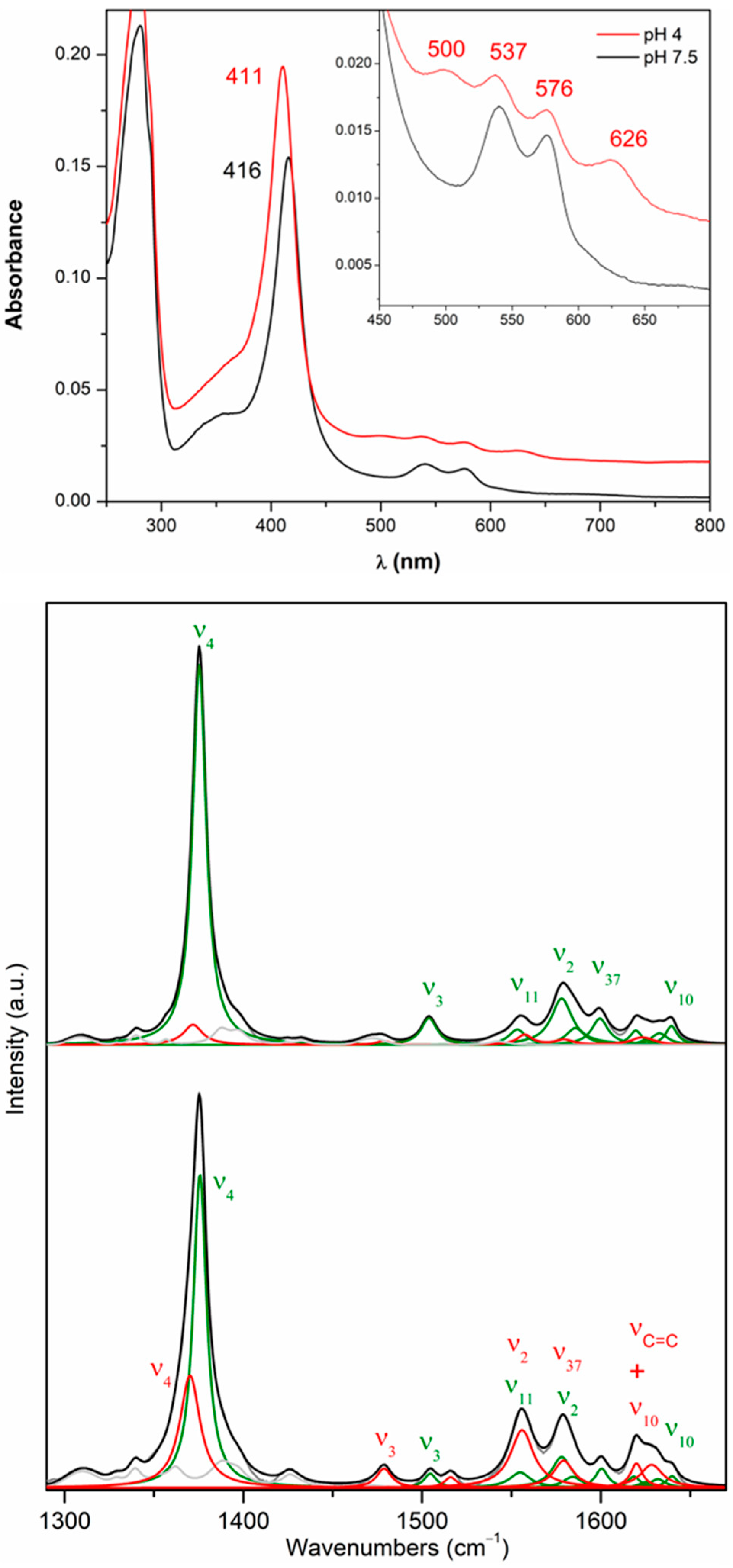
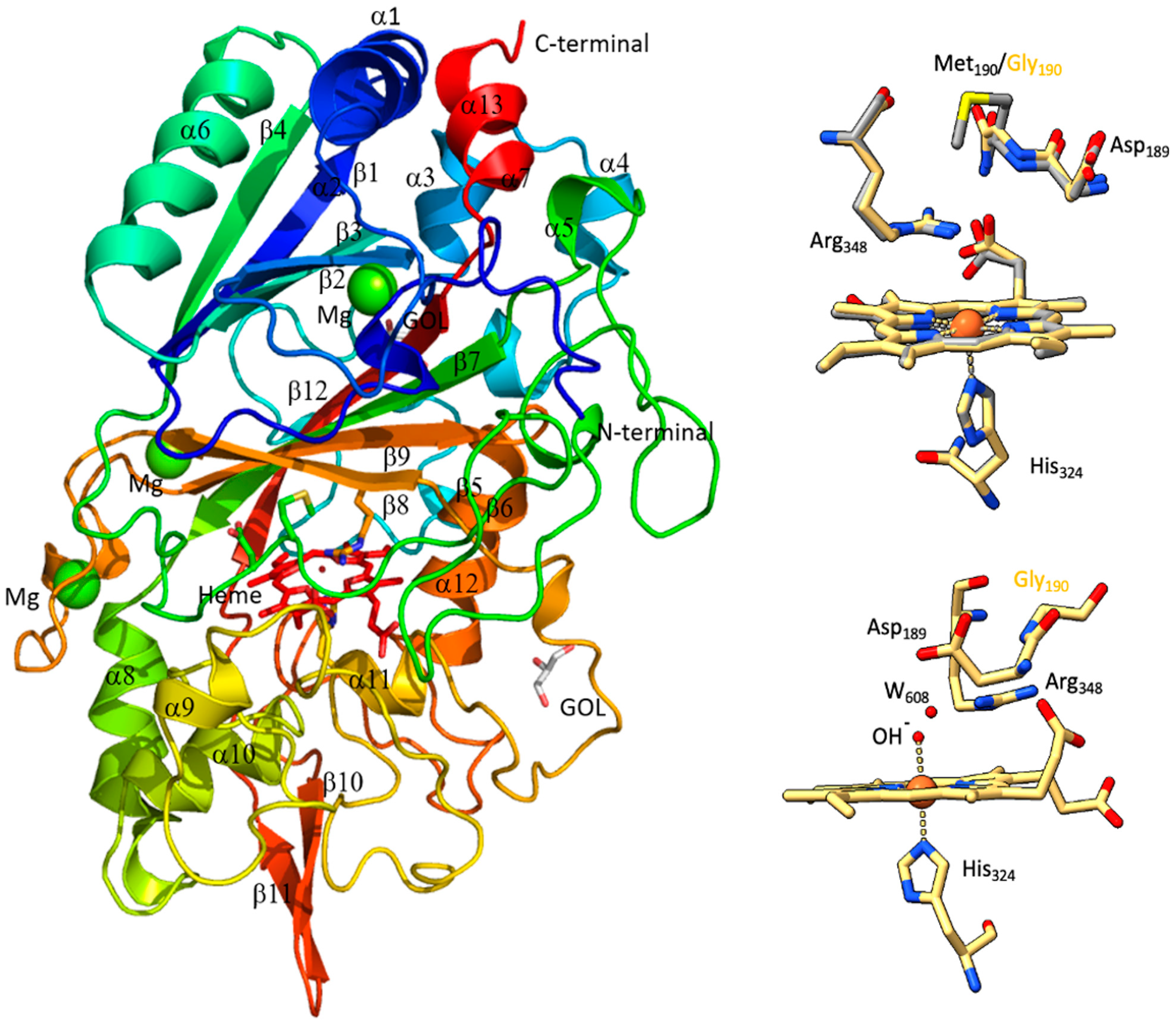
2.3. Structural Analysis of Heme in DrDyp and DrDypM190G
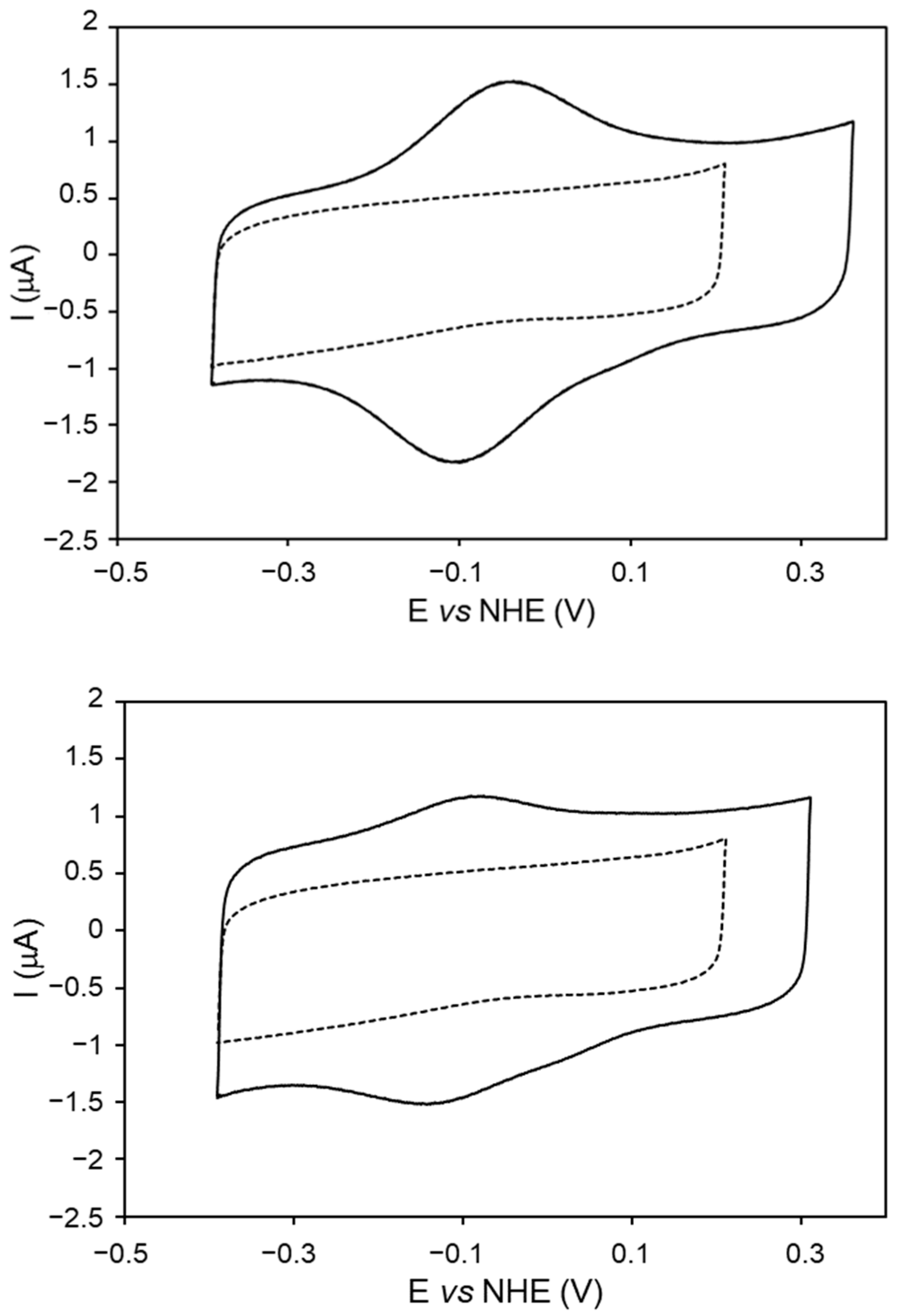
2.4. Redox Properties of DrDyP
2.5. Crystal-Structure Determination and Analysis
3. Materials and Methods
3.1. Recombinant Protein Production
3.2. UV-Visible and Resonance Raman Spectroscopy
3.3. Electrochemistry
3.4. Biochemical Assays
3.5. Buffer Stability Analysis (Thermofluor)
3.6. Crystallization, Data Collection, Processing, Structure Determination, and Refinement
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Gonzalo, G.; Colpa, D.I.; Habib, M.H.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef]
- Chen, C.; Li, T. Bacterial dye-decolorizing peroxidases: Biochemical properties and biotechnological opportunities. Phys. Sci. Rev. 2016, 1, 20160051. [Google Scholar] [CrossRef]
- Kim, S.J.; Shoda, M. Purification and characterization of a novel peroxidase from Geotrichum candidum dec 1 involved in decolorization of dyes. Appl. Environ. Microbiol. 1999, 65, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Eltis, L.D. The multihued palette of dye-decolorizing peroxidases. Arch. Biochem. Biophys. 2015, 574, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Colpa, D.I.; Fraaije, M.W.; van Bloois, E. DyP-type peroxidases: A promising and versatile class of enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1–7. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Bugg, T.D. Characterisation of Dyp-type peroxidases from Pseudomonas fluorescens Pf-5: Oxidation of Mn(II) and polymeric lignin by Dyp1B. Arch. Biochem. Biophys. 2015, 574, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Salvachua, D.; Prieto, A.; Martinez, A.T.; Martinez, M.J. Characterization of a novel dye-decolorizing peroxidase (DyP)-type enzyme from Irpex lacteus and its application in enzymatic hydrolysis of wheat straw. Appl. Environ. Microbiol. 2013, 79, 4316–4324. [Google Scholar] [CrossRef] [PubMed]
- Mendes, S.; Brissos, V.; Gabriel, A.; Catarino, T.; Turner, D.L.; Todorovic, S.; Martins, L.O. An integrated view of redox and catalytic properties of B-type PpDyP from Pseudomonas putida MET94 and its distal variants. Arch. Biochem. Biophys. 2015, 574, 99–107. [Google Scholar] [CrossRef]
- Upadhyay, P.; Shrivastava, R.; Agrawal, P.K. Bioprospecting and biotechnological applications of fungal laccase. 3 Biotech 2016, 6, 15. [Google Scholar] [CrossRef]
- Barbosa, C.; Silveira, C.M.; Silva, D.; Brissos, V.; Hildebrandt, P.; Martins, L.O.; Todorovic, S. Immobilized dye-decolorizing peroxidase (DyP) and directed evolution variants for hydrogen peroxide biosensing. Biosens. Bioelectron. 2020, 153, 112055. [Google Scholar] [CrossRef]
- Hofbauer, S.; Pfanzagl, V.; Michlits, H.; Schmidt, D.; Obinger, C.; Furtmüller, P.G. Understanding molecular enzymology of porphyrin-binding α + β barrel proteins—One fold, multiple functions. Biochim. Biophys. Acta BBA Proteins Proteom. 2021, 1869, 140536. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Sasaki, M.; Tanaka, Y.; Ishimori, K. A Dye-Decolorizing Peroxidase from Vibrio cholerae. Biochemistry 2015, 54, 6610–6621. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Sugano, Y. A structural and functional perspective of DyP-type peroxidase family. Arch. Biochem. Biophys. 2015, 574, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Poulos, T.L. Thirty years of heme peroxidase structural biology. Arch. Biochem. Biophys. 2010, 500, 3–12. [Google Scholar] [CrossRef]
- Furukawa, T.; Bello, F.O.; Horsfall, L. Microbial enzyme systems for lignin degradation and their transcriptional regulation. Front. Biol. 2014, 9, 448–471. [Google Scholar] [CrossRef]
- Pfanzagl, V.; Nys, K.; Bellei, M.; Michlits, H.; Mlynek, G.; Battistuzzi, G.; Djinovic-Carugo, K.; Van Doorslaer, S.; Furtmüller, P.G.; Hofbauer, S.; et al. Roles of distal aspartate and arginine of B-class dye-decolorizing peroxidase in heterolytic hydrogen peroxide cleavage. J. Biol. Chem. 2018, 293, 14823–14838. [Google Scholar] [CrossRef]
- Lučić, M.; Svistunenko, D.A.; Wilson, M.T.; Chaplin, A.K.; Davy, B.; Ebrahim, A.; Axford, D.; Tosha, T.; Sugimoto, H.; Owada, S.; et al. Serial Femtosecond Zero Dose Crystallography Captures a Water-Free Distal Heme Site in a Dye-Decolorising Peroxidase to Reveal a Catalytic Role for an Arginine in FeIV=O Formation. Angew. Chem. Int. Ed. 2020, 59, 21656–21662. [Google Scholar] [CrossRef]
- Chen, C.; Shrestha, R.; Jia, K.; Gao, P.F.; Geisbrecht, B.V.; Bossmann, S.H.; Shi, J.; Li, P. Characterization of Dye-decolorizing Peroxidase (DyP) from Thermomonospora curvata Reveals Unique Catalytic Properties of A-type DyPs. J. Biol. Chem. 2015, 290, 23447–23463. [Google Scholar] [CrossRef]
- Silveira, C.M.; Moe, E.; Fraaije, M.; Martins, L.O.; Todorovic, S. Resonance Raman view of the active site architecture in bacterial DyP-type peroxidases. RSC Adv. 2020, 10, 11095–11104. [Google Scholar] [CrossRef]
- Frade, K.S.T.; Fernandes, A.C.P.; Silveira, C.M.; Frazao, C.; Moe, E. A novel bacterial class V dye-decolourizing peroxidase from the extremophile Deinococcus radiodurans: Cloning, expression optimization, purification, crystallization, initial characterization and X-ray diffraction analysis. Acta Crystallogr. F Struct. Biol. Commun. 2018, 74, 419–424. [Google Scholar] [CrossRef]
- Fawal, N.; Li, Q.; Savelli, B.; Brette, M.; Passaia, G.; Fabre, M.; Mathe, C.; Dunand, C. PeroxiBase: A database for large-scale evolutionary analysis of peroxidases. Nucleic Acids Res. 2013, 41, D441–D444. [Google Scholar] [CrossRef] [PubMed]
- Sezer, M.; Santos, A.; Kielb, P.; Pinto, T.; Martins, L.O.; Todorovic, S. Distinct structural and redox properties of the heme active site in bacterial dye decolorizing peroxidase-type peroxidases from two subfamilies: Resonance Raman and electrochemical study. Biochemistry 2013, 52, 3074–3084. [Google Scholar] [CrossRef] [PubMed]
- Feis, A.; Howes, B.D.; Indiani, C.; Smulevich, G. Resonance Raman and electronic absorption spectra of horseradish peroxidase isozyme A2: Evidence for a quantum-mixed spin species. J. Raman Spectrosc. 1998, 29, 933–938. [Google Scholar] [CrossRef]
- Brown, M.E.; Barros, T.; Chang, M.C.Y. Identification and Characterization of a Multifunctional Dye Peroxidase from a Lignin-Reactive Bacterium. ACS Chem. Biol. 2012, 7, 2074–2081. [Google Scholar] [CrossRef]
- Pfanzagl, V.; Bellei, M.; Hofbauer, S.; Laurent, C.V.F.P.; Furtmüller, P.G.; Oostenbrink, C.; Battistuzzi, G.; Obinger, C. Redox thermodynamics of B-class dye-decolorizing peroxidases. J. Inorg. Biochem. 2019, 199, 110761. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Tsuge, H.; Konno, H.; Hisabori, T.; Sugano, Y. The catalytic mechanism of dye-decolorizing peroxidase DyP may require the swinging movement of an aspartic acid residue. FEBS J. 2011, 278, 2387–2394. [Google Scholar] [CrossRef] [PubMed]
- Strittmatter, E.; Liers, C.; Ullrich, R.; Wachter, S.; Hofrichter, M.; Plattner, D.A.; Piontek, K. First Crystal Structure of a Fungal High-redox Potential Dye-decolorizing Peroxidase. J. Biol. Chem. 2013, 288, 4095–4102. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fueyo, E.; Davó-Siguero, I.; Almendral, D.; Linde, D.; Baratto, M.C.; Pogni, R.; Romero, A.; Guallar, V.; Martínez, A.T. Description of a Non-Canonical Mn(II)-Oxidation Site in Peroxidases. ACS Catal. 2018, 8, 8386–8395. [Google Scholar] [CrossRef]
- Li, L.; Wang, T.; Chen, T.; Huang, W.; Zhang, Y.; Jia, R.; He, C. Revealing two important tryptophan residues with completely different roles in a dye-decolorizing peroxidase from Irpex lacteus F17. Biotechnol. Biofuels 2021, 14, 128. [Google Scholar] [CrossRef]
- Lučić, M.; Wilson, M.T.; Svistunenko, D.A.; Owen, R.L.; Hough, M.A.; Worrall, J.A.R. Aspartate or arginine? Validated redox state X-ray structures elucidate mechanistic subtleties of FeIV = O formation in bacterial dye-decolorizing peroxidases. JBIC J. Biol. Inorg. Chem. 2021, 26, 743–761. [Google Scholar] [CrossRef]
- Sarre, A.; Okvist, M.; Klar, T.; Moe, E.; Timmins, J. Expression, purification and crystallization of two endonuclease III enzymes from Deinococcus radiodurans. Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 1688–1692. [Google Scholar] [CrossRef] [PubMed]
- Kapust, R.B.; Tozser, J.; Fox, J.D.; Anderson, D.E.; Cherry, S.; Copeland, T.D.; Waugh, D.S. Tobacco etch virus protease: Mechanism of autolysis and rational design of stable mutants with wild-type catalytic proficiency. Protein Eng. 2001, 14, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Döpner, S.; Hildebrandt, P.; Grant Mauk, A.; Lenk, H.; Stempfle, W. Analysis of vibrational spectra of multicomponent systems. Application to pH-dependent resonance Raman spectra of ferricytochrome c. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 1996, 52, 573–584. [Google Scholar] [CrossRef]
- Santos, S.P.; Bandeiras, T.M.; Pinto, A.F.; Teixeira, M.; Carrondo, M.A.; Romão, C.V. Thermofluor-based optimization strategy for the stabilization and crystallization of Campylobacter jejuni desulforubrerythrin. Protein Expr. Purif. 2012, 81, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Kozak, S.; Lercher, L.; Karanth, M.N.; Meijers, R.; Carlomagno, T.; Boivin, S. Optimization of protein samples for NMR using thermal shift assays. J. Biomol. NMR 2016, 64, 281–289. [Google Scholar] [CrossRef]
- Kabsch, W. Xds. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Tickle, I.J.; Flensburg, C.; Keller, P.; Paciorek, W.; Sharff, A.; Vonrhein, C.; Bricogne, G. STARANISO; Global Phasing Ltd.: Cambridge, UK, 2018. [Google Scholar]
- Matthews, B.W. Solvent content of protein crystals. J. Mol. Biol. 1968, 33, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Kantardjieff, K.A.; Rupp, B. Matthews coefficient probabilities: Improved estimates for unit cell contents of proteins, DNA, and protein–nucleic acid complex crystals. Protein Sci. 2009, 12, 1865–1871. [Google Scholar] [CrossRef]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phasercrystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2009, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DyP | DrDyPM190G | DrDyP |
|---|---|---|
| Beamline | Petra-P13 | DLS-I03 |
| Space group a | P212121 | P32 |
| Wavelength (Å) | 0.99999 | 0.97625 |
| Resolution (Å) b | 74.74–1.51 (1.65–1.51) | 55.54–2.20 (2.33–2.20) |
| Unit-cell parameters (Å) | a = 82.090 b = 97.70 c = 116.04 | a = b = 64.130 c = 111.330 |
| Data processing | AutoPROC/STARANISO | XDS |
| Resolution limits of ellipsoid fitted to resolution cut-off surface (Å) | 2.03, 1.49, 1.60 | n/a |
| Resolution, spherical limits (Å) | 74.74–1.51 (1.65–1.51) | 55.54–2.20 (2.33–2.20) |
| No. of observations | 567,144 (24,552) | 104,559 (16,905) |
| No. of unique reflections | 98,273 (4914) | 25,853 (4196) |
| Multiplicity | 5.8 (5.0) | 4.04 (4.06) |
| <I>/<σ(I)> | 11.3 (1.7) | 10.96 (0.97) |
| %CC 1/2 | 99.7 (78.4) | 99.8 (50.8) |
| % Rmerg | 7.1 (84.9) | 6.6 (108.1) |
| % Rmeas | 7.8 (94.6) | 7.6 (124.4) |
| % Rpim | 3.2 (40.8) | 3.52 (34.7) |
| Data completeness (%), sperical (%) | 67.2 (14.2) | 99.5 (99.1) |
| Data completeness, ellipsoidal (%) | 93.4 (57.4) | n/a |
| Solvent contents (%) | 45.8 | 36.33 |
| Wilson B factor (Å2) | 17.99 | 55.82 |
| Refinement | ||
| Resolution range | 74.74–1.51 (1.53–1.51) | 39.31–2.20 (2.26–2.20) |
| No. protein molecules in a.u. | 2 | 1 |
| No. of residues per chain (total) | 444; 444 (888) | 441 (441) |
| No. of solvent waters | 653 | 50 |
| No. of Haem groups per chain | 1, 1 (2) | 1 |
| No. of glycerol molecules per chain | - | 2 |
| No. of Ca2+ ions | 1, 1 (2) | - |
| No. of Mg2+ ions | - | 3 |
| No. of Cl− ions | 1, 1 (2) | - |
| Rwork | 0.167 (0.317) | 0.183 (0.360) |
| Rfree | 0.195 (0.187) | 0.237 (0.398) |
| Estimated coordinate error | 0.13 | 0.34 |
| <adp>s of protein chains (Å2) | 26.2, 24.2 | 59.09 |
| <adp>s of haem groups per chain (Å2) | - | - |
| <adp>s of solvent waters (Å2) | 29.3 | 53.10 |
| R.m.s.d deviations from ideal values | ||
| Bonds (Å) | 0.010 | 0.008 |
| Angles (°) | 1.050 | 0.963 |
| Ramachandran plot analysis: %residues in | ||
| Favoured regions (%) | 97.97 | 98.18 |
| Allowed regions | 1.80 | 1.82 |
| Disallowed regions | 0.23 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frade, K.; Silveira, C.M.; Salgueiro, B.A.; Mendes, S.; Martins, L.O.; Frazão, C.; Todorovic, S.; Moe, E. Biochemical, Biophysical, and Structural Analysis of an Unusual DyP from the Extremophile Deinococcus radiodurans. Molecules 2024, 29, 358. https://doi.org/10.3390/molecules29020358
Frade K, Silveira CM, Salgueiro BA, Mendes S, Martins LO, Frazão C, Todorovic S, Moe E. Biochemical, Biophysical, and Structural Analysis of an Unusual DyP from the Extremophile Deinococcus radiodurans. Molecules. 2024; 29(2):358. https://doi.org/10.3390/molecules29020358
Chicago/Turabian StyleFrade, Kelly, Célia M. Silveira, Bruno A. Salgueiro, Sónia Mendes, Lígia O. Martins, Carlos Frazão, Smilja Todorovic, and Elin Moe. 2024. "Biochemical, Biophysical, and Structural Analysis of an Unusual DyP from the Extremophile Deinococcus radiodurans" Molecules 29, no. 2: 358. https://doi.org/10.3390/molecules29020358
APA StyleFrade, K., Silveira, C. M., Salgueiro, B. A., Mendes, S., Martins, L. O., Frazão, C., Todorovic, S., & Moe, E. (2024). Biochemical, Biophysical, and Structural Analysis of an Unusual DyP from the Extremophile Deinococcus radiodurans. Molecules, 29(2), 358. https://doi.org/10.3390/molecules29020358