
Valorization of Citrus Reticulata Peels for Flavonoids and Antioxidant Enhancement by Solid-State Fermentation Using Aspergillus niger CGMCC 3.6189


 and
and
Abstract

1. Introduction
2. Results
2.1. Effect of pH on TPC, TFC, Antioxidant Activity, and Phytochemical Compositions in CRPP
2.2. Effect of Fermentation Temperature on TPC, TFC, Antioxidant Activity, and Phytochemical Compositions in CRPP
2.3. Effect of Moisture Content on TPC, TFC, Antioxidant Activity, and Phytochemical Compositions in CRPP
2.4. Effect of Spore Concentration on TPC, TFC, Antioxidant Activity, and Phytochemical Compositions in CRPP
2.5. Effect of Fermentation Times on TPC, TFC, Antioxidant Activity, and Phytochemical Compositions in CRPP
2.6. Effect of Fermentation Conditions on the Color of CRPP
2.7. Pearson Correlation between SSF Conditions and Quality Attributes of CRPP
2.8. Principal Component Analysis of the Quality Attributes of CRPP
3. Discussion
4. Materials and Methods
4.1. Materials and Chemical
4.2. Samples Preparation
4.3. Preparation of the Growth Curve and the Inoculum of Aspergillus niger CGMCC 3.6189
4.4. Solid-State Fermentation (SSF)
4.5. Extraction of CRPP
4.6. Analysis of Total Phenolic Content (TPC)
4.7. Analysis of Total Flavonoid Content (TFC)
4.8. Analysis of ABTS Radical Scavenging Capacity
4.9. Analysis of DPPH Radical Scavenging Capacity
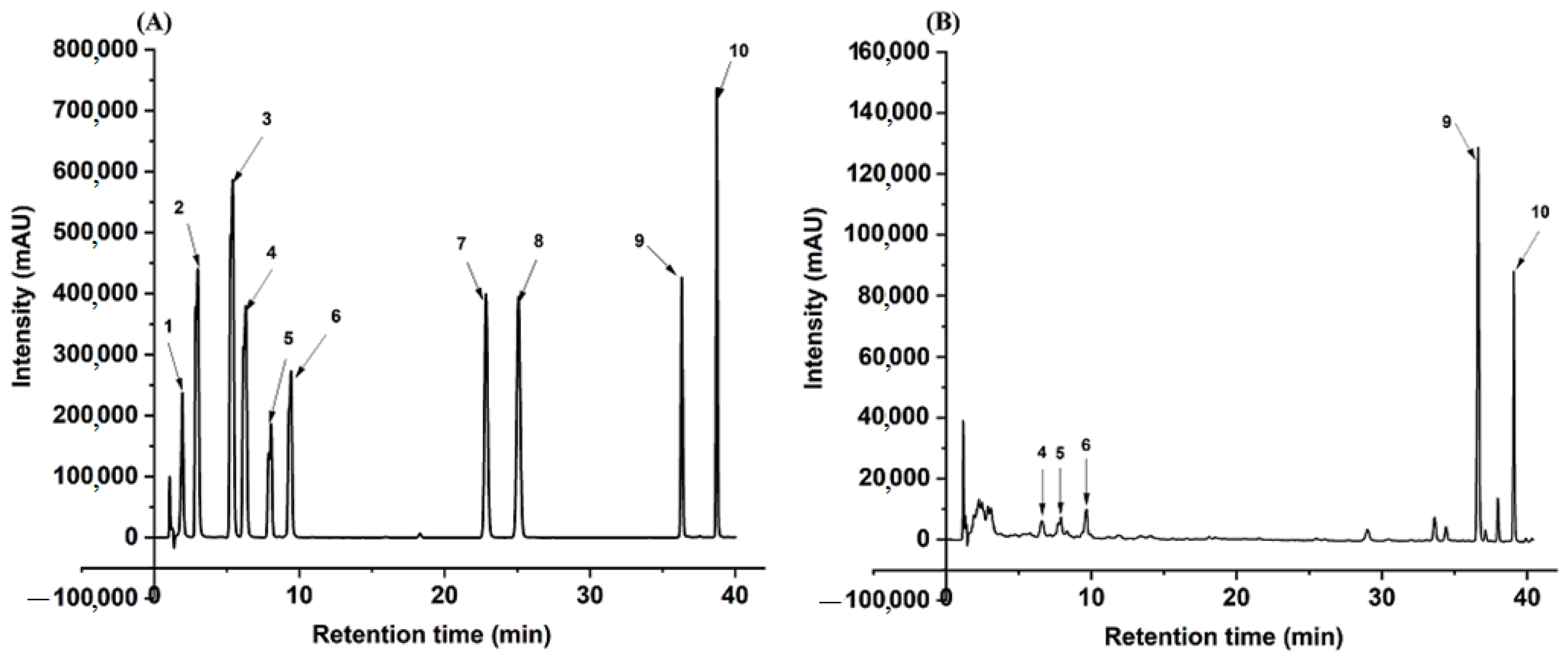
4.10. Analysis of Phytochemicals Using HPLC
4.11. Determination of Color Parameters
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- FAO. Citrus fruit fresh and processed statistical bulletin 2020. Stat. Bull. 2020, 1–40. Available online: https://www.fao.org/3/cb6492en/cb6492en.pdf. (accessed on 5 April 2022).
- Kaur, S.; Panesar, P.S.; Chopra, H.K. Citrus processing by-products: An overlooked repository of bioactive compounds. Crit. Rev. Food Sci. Nutr. 2021, 1–20. [Google Scholar] [CrossRef]
- Choi, M.Y.; Chai, C.; Park, J.H.; Lim, J.; Lee, J.; Kwon, S.W. Effects of storage period and heat treatment on phenolic compound composition in dried Citrus peels (Chenpi) and discrimination of Chenpi with different storage periods through targeted metabolomic study using HPLC-DAD analysis. J. Pharm. Biomed. Anal. 2011, 54, 638–645. [Google Scholar] [CrossRef]
- Wedamulla, N.E.; Fan, M.; Choi, Y.J.; Kim, E.K. Citrus peel as a renewable bioresource: Transforming waste to food additives. J. Funct. Foods. 2022, 95, 19. [Google Scholar] [CrossRef]
- Liu, Y.; Benohoud, M.; Galani Yamdeu, J.H.; Gong, Y.Y.; Orfila, C. Green extraction of polyphenols from citrus peel by-products and their antifungal activity against Aspergillus flavus. Food Chem. X 2021, 12, 100144. [Google Scholar] [CrossRef]
- Shakour, Z.T.A.; Fayek, N.M.; Farag, M.A. How do biocatalysis and biotransformation affect Citrus dietary flavonoids chemistry and bioactivity? A review. Crit. Rev. Biotechnol. 2020, 40, 689–714. [Google Scholar] [CrossRef]
- Wang, F.; Chen, L.; Li, F.Q.; Liu, S.J.; Chen, H.P.; Liu, Y.P. The Increase of Flavonoids in Pericarpium Citri Reticulatae (PCR) Induced by Fungi Promotes the Increase of Antioxidant Activity. Evid. Based Complement. Alternat. Med. 2018, 2018, 2506037. [Google Scholar] [CrossRef]
- Luo, Y.; Zeng, W.; Huang, K.E.; Li, D.X.; Chen, W.; Yu, X.Q.; Ke, X.H. Discrimination of Citrus reticulata Blanco and Citrus reticulata ‘Chachi’ as well as the Citrus reticulata ‘Chachi’ within different storage years using ultra high performance liquid chromatography quadrupole/time-of-flight mass spectrometry based metabol. J. Pharm. Biomed. Anal. 2019, 171, 218–231. [Google Scholar] [CrossRef]
- Yang, M.; Jiang, Z.; Wen, M.; Wu, Z.; Zha, M.; Xu, W.; Zhang, L. Chemical Variation of Chenpi (Citrus Peels) and Corresponding Correlated Bioactive Compounds by LC-MS Metabolomics and Multibioassay Analysis. Front. Nutr. 2022, 9, 825381. [Google Scholar] [CrossRef]
- Chen, J.; He, C.; He, Q.; Li, J.; Ying, F.; Chen, G. The central bacterial community in Pericarpium Citri Reticulatae ‘Chachiensis. Food Res. Int. 2019, 125, 108624. [Google Scholar] [CrossRef]
- Parshikov, I.A.; Sutherland, J.B. Biotransformation of Steroids and Flavonoids by Cultures of Aspergillus niger. Appl. Biochem. Biotechnol. 2015, 176, 903–923. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Chen, X.; Jassbi, A.R.; Xiao, J. Microbial biotransformation of bioactive flavonoids. Biotechnol. Adv. 2015, 33, 214–223. [Google Scholar] [CrossRef]
- Ahmed, T.; Rana, M.R.; Zzaman, W.; Ara, R.; Aziz, M.G. Optimization of substrate composition for pectinase production from Satkara (Citrus macroptera) peel using Aspergillus niger-ATCC 1640 in solid-state fermentation. Heliyon 2021, 7, e08133. [Google Scholar] [CrossRef]
- Shi, H.; Yang, E.; Li, Y.; Chen, X.; Zhang, J. Effect of Solid-State Fermentation on Nutritional Quality of Leaf Flour of the Drumstick Tree (Moringa oleifera Lam.). Front. Bioeng. Biotechnol. 2021, 9, 626628. [Google Scholar] [CrossRef]
- Manan, M.A.; Webb, C. Design Aspects of Solid State Fermentation as Applied to Microbial Bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 511–532. [Google Scholar] [CrossRef]
- Bose, P.; Gowrie, S.U.; Chathurdevi, G. Optimization of Culture Conditions for Growth and Production of Bioactive Metabolites by Endophytic Fungus-Aspergillus tamarii. Int. J. Pharm. Biol. Sci. 2019, 9, 469–478. [Google Scholar] [CrossRef]
- Xiong, J.; Ding, L. Optimised Aspergillus niger enzyme-assisted extraction of flavonoids from Dicranopteris and evaluation of antioxidant activity in vitro. Adv. Mater. Res. 2012, 396, 1436–1439. [Google Scholar] [CrossRef]
- Bind, A.; Singh, S.K.; Prakash, V.; Kumar, M. Evaluation of antioxidants through solid state fermentation from pomegranate peels using Aspergillus niger and it’s antibacterial properties. Indones. J. Pharm. Biol. Sci. 2014, 4, 104–112. [Google Scholar]
- Jiao, J.; Gai, Q.Y.; Wang, W.; Zang, Y.P.; Niu, L.L.; Fu, Y.J.; Wang, X. Remarkable enhancement of flavonoid production in a co-cultivation system of Isatis tinctoria L. hairy root cultures and immobilized Aspergillus niger. Ind. Crops Prod. 2018, 112, 252–261. [Google Scholar] [CrossRef]
- Ahmed, N.S.; Al- Shamary, E.I. Optimization of Phenolic Compound Production by Local Aspergillus niger B1b Isolate. IOP Conf. Ser. Earth Environ. Sci. 2021, 761, 012119. [Google Scholar] [CrossRef]
- Labrath, Y.P.; Gaikar, V.G. Solid State Fermentation of Orange Peels for Production of Cellulase, Pectinase and Recovery of Orange Oil using Aspergillus Species NCIM 1432. Res. Sq. 2020, 30. [Google Scholar] [CrossRef]
- Rinu, K.; Pandey, A. Temperature-dependent phosphate solubilization by cold- and pH-tolerant species of Aspergillus isolated from Himalayan soil. Mycoscience. 2010, 51, 263–271. [Google Scholar] [CrossRef]
- Liu, N.; Song, M.; Wang, N.; Wang, Y.; Wang, R.; An, X.; Qi, J. The effects of solid-state fermentation on the content, composition and in vitro antioxidant activity of flavonoids from dandelion. PLoS ONE 2020, 15, e0239076. [Google Scholar] [CrossRef]
- Mamma, D.; Kourtoglou, E.; Christakopoulos, P. Fungal multienzyme production on industrial by-products of the citrus-processing industry. Bioresour. Technol. 2008, 99, 2373–2383. [Google Scholar] [CrossRef] [PubMed]
- Pe´rez-Guerra, N.; Torrado-Agrasar, A.; Lo´pez-Marias, C.; Pastrana, L. Main characteristics and applications of solid substrate fermentation. Food Chem. 2003, 2, 343–350. [Google Scholar]
- Torrado, A.M.; Cortés, S.; Salgado, J.M.; Max, B.; Rodríguez, N.; Bibbins, B.P.; Converti, A.; Domínguez, J.M. Citric acid production from orange peel wastes by solid-state fermentation. Braz. J. Microbiol. 2011, 42, 394–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, L.; Chen, S.; Chen, H.; Liu, Y. Microbial biotransformation of Pericarpium Citri Reticulatae (PCR) by Aspergillus niger and effects on antioxidant activity. Food Sci. Nutr. 2021, 9, 855–865. [Google Scholar] [CrossRef]
- Cai, S.; Wang, O.; Wu, W.; Zhu, S.; Zhou, F.; Ji, B.; Gao, F.; Zhang, D. Comparative Study of the Effects of Solid-State Fermentation with Three Filamentous Fungi on the Total Phenolics Content (TPC), Flavonoids, and Antioxidant Activities of Subfractions from Oats (Avena sativa L.). Agric. Food Chem. 2012, 60, 507–5013. [Google Scholar] [CrossRef]
- Nigam, P.; Singh, D. Solid-state (substrate) fermentation systems and their applications in biotechnology. J. Basic Microbiol. 1994, 34, 405–423. [Google Scholar] [CrossRef]
- Auta, H.S.; Abidoye, K.T.; Tahir, H.; Ibrahim, A.D.; Aransiola, S.A. Citric Acid Production by Aspergillus niger Cultivated on Parkia biglobosa Fruit Pulp. Int. Sch. Res. Not. 2014, 2014, 762021. [Google Scholar] [CrossRef]
- Pérez-Nájera, V.C.; Lugo-Cervantes, E.; Amaya-Delgado, L.; Madrigal-Pulido, J.A.; Rueda-Puente, E.O.; Borboa-Flores, J.; Del-Toro-Sánchez, C.L. Biotransformation of hesperidin from lime peel (Citrus limetta risso) in solid fermentation by Aspergillus saitoi. J. Food 2018, 16, 537–543. [Google Scholar] [CrossRef]
- Santos, T.R.J.; Feitosa, P.R.B.; Gualberto, N.C.; Narain, N.; Santana, L.C.L.A. Improvement of bioactive compounds content in granadilla (Passiflora ligularis) seeds after solid-state fermentation. Food Sci. Technol. Int. 2021, 27, 234–241. [Google Scholar] [CrossRef]
- Kim, S.S.; Park, K.J.; An, H.J.; Choi, Y.H. Phytochemical, antioxidant, and antibacterial activities of fermented Citrus unshiu byproduct. Food Sci. Biotechnol. 2017, 26, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, Y.A.-G.; Assawah, S.W.; El-Sharkawy, S.H.; Abdel-Salam, A. Flavone Biotransformation by Aspergillus niger and the Characterization of Two Newly Formed Metabolites. Mycobiology 2008, 36, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, J.; Alderete, J.; Escobar, C.; Araya, R.; Cespedes, C.L. Aspergillus niger catalyzes the synthesis of flavonoids from chalcones. Biocatal. Biotransform. 2013, 31, 160–167. [Google Scholar] [CrossRef]
- Long, X.; Zeng, X.; Yan, H.; Xu, M.; Zeng, Q.; Xu, C.; Xu, Q.; Liang, Y.; Zhang, J. Flavonoids composition and antioxidant potential assessment of extracts from Gannanzao Navel Orange (Citrus sinensis Osbeck Cv. Gannanzao) peel. Nat. Prod. Res. 2021, 35, 702–706. [Google Scholar] [CrossRef]
- Guo, C.; Shan, Y.; Yang, Z.; Zhang, L.; Ling, W.; Liang, Y.; Ouyang, Z.; Zhong, B.; Zhang, J. Chemical composition, antioxidant, antibacterial, and tyrosinase inhibition activity of extracts from Newhall navel orange (Citrus sinensis Osbeck cv. Newhall) peel. J. Sci. Food Agric. 2020, 100, 2664–2674. [Google Scholar] [CrossRef]
- Muzdalifah, D.; Athaillah, Z.A.; Nugrahani, W.; Devi, A.F. Colour and pH changes of tempe during extended fermentation. AIP Conf. Proc. 2017, 1803, 020036. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, Y.; Lu, T.; Chen, X. Temporal kinetics of changes in color, phytochemicals, γ -aminobutyric acid, enzyme activity and antioxidant activity of coffee leaves during postharvest storage. Sci. Hortic. 2022, 304, 111360. [Google Scholar] [CrossRef]
- Ar, T.S.A.; Tahir, M.M.; Wijaya, H. The degreening of “selayar” orange using ethephon: The color peel changes and ethephon residue. IOP Conf. Ser. Earth Environ. Sci. 2021, 807, 022022. [Google Scholar] [CrossRef]
- Wang, J.; Cao, F.; Su, E.; Zhao, L.; Qin, W. Improvement of animal feed additives of ginkgo leaves through solid-state fermentation using Aspergillus niger. Int. J. Biol. Sci. 2018, 14, 736–747. [Google Scholar] [CrossRef]
- Bastidas, O. Cell counting with neubauer chamber, basic hemocytometer usage. Tech. Note-Neubauer Chamb. Cell Count. 2013, 6. [Google Scholar] [CrossRef]
- Luo, M.; Luo, H.; Hu, P.; Yang, Y.; Wu, B.; Zheng, G. Evaluation of chemical components in Citri Reticulatae Pericarpium of different cultivars collected from different regions by GC–MS and HPLC. Food Sci. Nutr. 2018, 6, 400–416. [Google Scholar] [CrossRef]
- Chen, X.; Ding, J.; Ji, D.; He, S.; Ma, H. Optimization of ultrasonic-assisted extraction conditions for bioactive components from coffee leaves using the Taguchi design and response surface methodology. J. Food Sci. 2020, 85, 1742–1751. [Google Scholar] [CrossRef]
- Shraim, A.M.; Ahmed, T.A.; Rahman, M.M.; Hijji, Y.M. Determination of total flavonoid content by aluminum chloride assay: A critical evaluation. LWT 2021, 150, 111932. [Google Scholar] [CrossRef]
- Chen, X.; Kitts, D.D.; Ji, D.; Ding, J. Free radical scavenging activities of phytochemical mixtures and aqueous methanolic extracts recovered from processed coffee leaves. Int. J. Food Sci. Technol. 2019, 54, 2872–2879. [Google Scholar] [CrossRef]
- Gao, L.; Mei, S.; Ma, H.; Chen, X. Ultrasound-assisted green synthesis of gold nanoparticles using citrus peel extract and their enhanced anti-inflammatory activity. Ultrason. Sonochem. 2022, 83, 105940. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Variables | TPC (mg GAE/g) | TFC (mg QE/g) | ABTS (µmol TE/g) | DPPH (µmol TE/g) |
|---|---|---|---|---|---|
| pH 1 | |||||
| Control | - | 10.77 ± 0.27 b | 4.78 ± 0.07 a | 22.19 ± 0.97 abc | 13.35 ± 0.71 a |
| FC1a | 4.0 | 11.97 ± 0.26 a | 4.83 ± 0.11 a | 25.43 ± 1.89 a | 14.05 ± 1.12 a |
| FC1b | 4.5 | 10.57 ± 0.18 b | 4.28 ± 0.15 b | 25.07 ± 1.09 ab | 14.65 ± 0.49 a |
| FC1c | 5.0 | 10.40 ± 0.08 b | 4.06 ± 0.17 b | 24.97 ± 1.99 ab | 14.61 ± 0.82 a |
| FC1d | 5.5 | 10.48 ± 0.09 b | 4.07 ± 0.09 b | 23.63 ± 0.20 ab | 13.56 ± 0.46 a |
| FC1e | 6.0 | 10.41 ± 0.30 b | 3.67 ± 0.10 c | 21.46 ± 1.91 bc | 13.65 ± 0.90 a |
| FC1f | 6.5 | 10.44 ± 0.36 b | 3.51 ± 0.06 c | 18.72 ± 0.17 c | 13.61 ± 0.92 a |
| Incubation temperature (IT, °C) 2 | |||||
| Control | - | 10.77 ± 0.27 b | 4.78 ± 0.07 a | 22.19 ± 0.97 a | 13.35 ± 0.71 a |
| FC2a | 25 | 11.41 ± 0.35 ab | 4.71 ± 0.18 a | 24.56 ± 1.69 a | 13.44 ± 0.69 a |
| FC2b | 30 | 11.97 ± 0.26 a | 4.83 ± 0.11 a | 25.43 ± 1.89 a | 14.05 ± 1.12 a |
| FC2c | 35 | 12.05 ± 0.44 a | 4.01 ± 0.14 b | 22.49 ± 1.20 a | 13.33 ± 1.44 a |
| Moisture content (MC,%, w.b.) 3 | |||||
| Control | - | 10.77 ± 0.27 c | 4.78 ± 0.07 a | 22.19 ± 0.97 b | 13.35 ± 0.71 a |
| FC3a | 70 | 11.97 ± 0.26 b | 4.83 ± 0.11 a | 25.43 ± 1.89 a | 14.05 ± 1.12 a |
| FC3b | 80 | 12.95 ± 0.59 a | 4.90 ± 0.06 a | 25.63 ± 0.47 a | 15.00 ± 1.41 a |
| FC3c | 90 | 13.43 ± 0.20 a | 4.29 ± 0.10 b | 24.73 ± 0.45 ab | 14.08 ± 0.44 a |
| Spore concentration (SC, spores/g) 4 | |||||
| Control | - | 10.77 ± 0.27 b | 4.78 ± 0.07 b | 22.19 ± 0.97 c | 13.35 ± 0.71 b |
| FC4a | 4 × 106 | 12.95 ± 0.59 a | 4.90 ± 0.06 ab | 25.63 ± 0.47 b | 15.00 ± 1.41 ab |
| FC4b | 2 × 107 | 13.17 ± 0.40 a | 5.04 ± 0.04 ab | 26.00 ± 0.21 b | 16.29 ± 1.83 ab |
| FC4c | 4 × 107 | 13.47 ± 0.36 b | 5.15 ± 0.23 a | 27.53 ± 0.24 a | 17.33 ± 1.40 a |
| Fermentation time (FT, h) 5 | |||||
| Control | - | 10.77 ± 0.27 d | 4.78 ± 0.07 d | 22.19 ± 0.97 c | 13.35 ± 0.71 c |
| FC5a | 60 | 13.47 ± 0.36 c | 5.15 ± 0.23 c | 27.53 ± 0.24 b | 17.33 ± 1.40 bc |
| FC5b | 96 | 13.77 ± 0.21 c | 5.35 ± 0.20 bc | 28.13 ± 1.36 b | 17.36 ± 1.09 bc |
| FC5c | 144 | 15.69 ± 0.33 b | 5.69 ± 0.21 ab | 29.37 ± 0.52 b | 18.99 ± 1.55 ab |
| FC5d | 192 | 18.31 ± 0.35 a | 6.06 ± 0.11 a | 36.60 ± 1.82 a | 22.91 ± 3.45 a |
| Samples | Variable | Ferulic Acid | Narirutin | Hesperidin | Nobiletin | Tangeretin |
|---|---|---|---|---|---|---|
| pH 1 | ||||||
| Control | - | 0.46 ± 0.00 a | 4.97 ± 0.07 b | 19.36 ± 0.47 a | 6.31 ± 0.11 a | 2.91 ± 0.04 a |
| FC1a | 4.0 | 0.45 ± 0.01 a | 5.53 ± 0.13 a | 19.69 ± 0.13 a | 6.36 ± 0.08 a | 2.91 ± 0.02 a |
| FC1b | 4.5 | ND | 4.25 ± 0.22 c | 13.83 ± 0.04 b | 5.63 ± 0.20 b | 2.59 ± 0.09 b |
| FC1c | 5.0 | ND | 4.23 ± 0.03 c | 9.28 ± 1.13 c | 5.76 ± 0.05 b | 2.64 ± 0.02 b |
| FC1d | 5.5 | ND | 4.25 ± 0.04 c | 7.46 ± 0.24 c | 5.73 ± 0.03 b | 2.67 ± 0.01 b |
| FC1e | 6.0 | 0.31 ± 0.00 b | 4.30 ± 0.16 c | 15.39 ± 0.72 b | 5.77 ± 0.05 b | 2.66 ± 0.02 b |
| FC1f | 6.5 | 0.24 ± 0.01 c | 4.17 ± 0.09 c | 8.41 ± 0.96 c | 5.56 ± 0.31 b | 2.54 ± 0.14 b |
| Incubation temperature (IT, °C) 2 | ||||||
| Control | - | 0.46 ± 0.00 a | 4.97 ± 0.07 bc | 19.36 ± 0.47 a | 6.31 ± 0.11 a | 2.91 ± 0.04 a |
| FC2a | 25 | 0.43 ± 0.02 a | 4.48 ± 0.33 c | 15.02 ± 0.36 b | 5.71 ± 0.05 b | 2.60 ± 0.03 b |
| FC2b | 30 | 0.45 ± 0.01 a | 5.53 ± 0.13 ab | 19.69 ± 0.13 a | 6.36 ± 0.08 a | 2.91 ± 0.02 a |
| FC2c | 35 | 0.41 ± 0.03 a | 5.76 ± 0.30 a | 16.41 ± 1.10 b | 5.45 ± 0.01 c | 2.52 ± 0.02 c |
| Moisture content (MC,%, w.b.) 3 | ||||||
| Control | - | 0.46 ± 0.00 b | 4.97 ± 0.07 b | 19.36 ± 0.47 c | 6.31 ± 0.11 a | 2.91 ± 0.04 a |
| FC3a | 70 | 0.45 ± 0.01 b | 5.53 ± 0.13 a | 19.69 ± 0.13 bc | 6.36 ± 0.08 a | 2.91 ± 0.02 a |
| FC3b | 80 | 0.53 ± 0.01 a | 5.41 ± 0.25 a | 22.73 ± 0.38 a | 6.27 ± 0.05 a | 2.63 ± 0.03 b |
| FC3c | 90 | 0.39 ± 0.01 c | 4.30 ± 0.14 c | 20.42 ± 0.47 b | 5.91 ± 0.15 b | 2.57 ± 0.07 b |
| Spore concentration (SC, spores/g) 4 | ||||||
| Control | - | 0.46 ± 0.00 b | 4.97 ± 0.07 c | 19.36 ± 0.47 c | 6.31 ± 0.11 ab | 2.91 ± 0.04 a |
| FC4a | 4x106 | 0.53 ± 0.01 a | 5.41 ± 0.25 ab | 22.73 ± 0.38 b | 6.27 ± 0.05 ab | 2.63 ± 0.03 b |
| FC4b | 2x107 | 0.39 ± 0.0 c | 5.69 ± 0.05 a | 23.85 ± 0.69 ab | 6.42 ± 0.04 ab | 3.01 ± 0.05 a |
| FC4c | 4x107 | 0.40 ± 0.01 c | 5.14 ± 0.15 bc | 24.89 ± 1.10 a | 6.46 ± 0.02 a | 3.05 ± 0.10 a |
| Fermentation time (FT, h) 5 | ||||||
| Control | - | 0.46 ± 0.00 a | 4.97 ± 0.07 c | 19.36 ± 0.47 c | 6.31 ± 0.11 b | 2.91 ± 0.04 c |
| FC5a | 60 | 0.40 ± 0.01 b | 5.14 ± 0.15 c | 24.89 ± 1.10 b | 6.46 ± 0.02 b | 3.05 ± 0.10 c |
| FC5b | 96 | 0.33 ± 0.00d | 5.56 ± 0.04 b | 27.52 ± 0.55 b | 7.91 ± 0.10 a | 3.68 ± 0.05 a |
| FC5c | 144 | 0.46 ± 0.00 a | 6.63 ± 0.03 a | 27.56 ± 0.77 b | 7.83 ± 0.04 a | 3.55 ± 0.02 ab |
| FC5d | 192 | 0.37 ± 0.01 c | 4.69 ± 0.09 d | 28.23 ± 0.76 a | 7.78 ± 0.07 a | 3.49 ± 0.06 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamy, D.; Huang, Y.; Akpabli-Tsigbe, N.D.K.; Battino, M.; Chen, X. Valorization of Citrus Reticulata Peels for Flavonoids and Antioxidant Enhancement by Solid-State Fermentation Using Aspergillus niger CGMCC 3.6189. Molecules 2022, 27, 8949. https://doi.org/10.3390/molecules27248949
Mamy D, Huang Y, Akpabli-Tsigbe NDK, Battino M, Chen X. Valorization of Citrus Reticulata Peels for Flavonoids and Antioxidant Enhancement by Solid-State Fermentation Using Aspergillus niger CGMCC 3.6189. Molecules. 2022; 27(24):8949. https://doi.org/10.3390/molecules27248949
Chicago/Turabian StyleMamy, Daniel, Yuanyuan Huang, Nelson Dzidzorgbe Kwaku Akpabli-Tsigbe, Maurizio Battino, and Xiumin Chen. 2022. "Valorization of Citrus Reticulata Peels for Flavonoids and Antioxidant Enhancement by Solid-State Fermentation Using Aspergillus niger CGMCC 3.6189" Molecules 27, no. 24: 8949. https://doi.org/10.3390/molecules27248949
APA StyleMamy, D., Huang, Y., Akpabli-Tsigbe, N. D. K., Battino, M., & Chen, X. (2022). Valorization of Citrus Reticulata Peels for Flavonoids and Antioxidant Enhancement by Solid-State Fermentation Using Aspergillus niger CGMCC 3.6189. Molecules, 27(24), 8949. https://doi.org/10.3390/molecules27248949