
Indolylazine Derivative Induces Chaperone Expression in Aged Neural Cells and Prevents the Progression of Alzheimer’s Disease

 , ,
, , 
Abstract

1. Introduction
2. Results
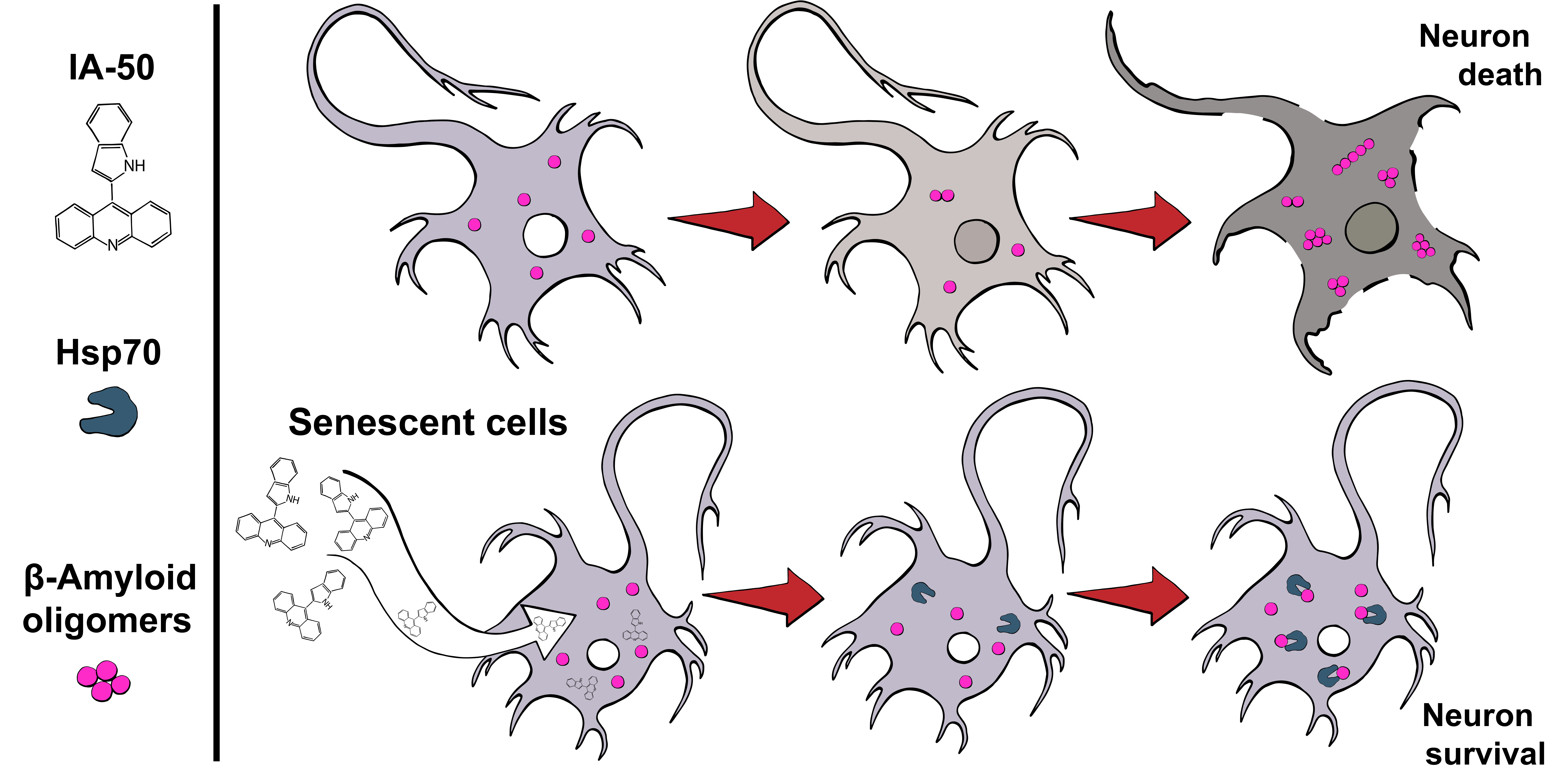
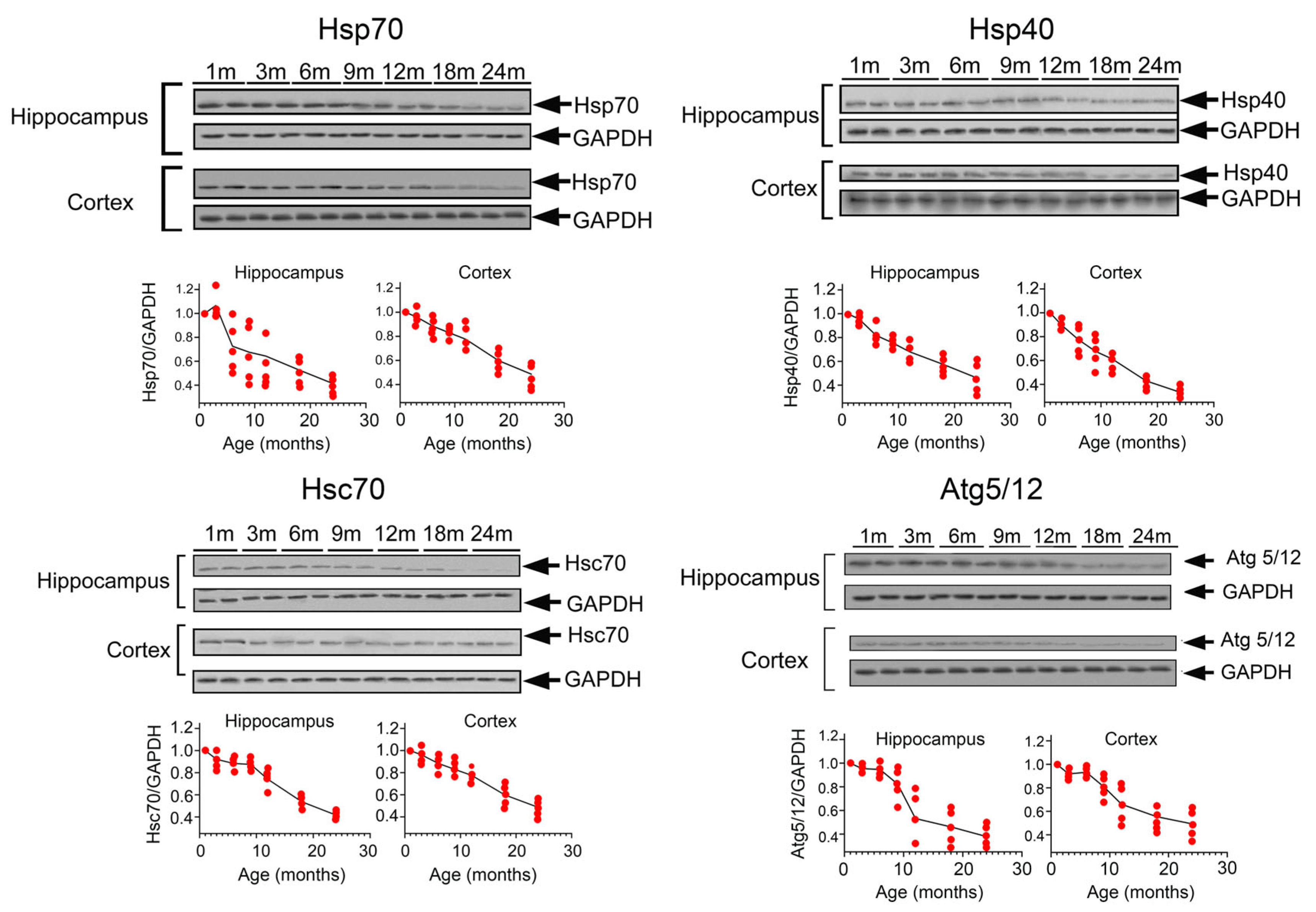
2.1. Analysis of Proteins Regulating Cell Proteostasis in Ageing
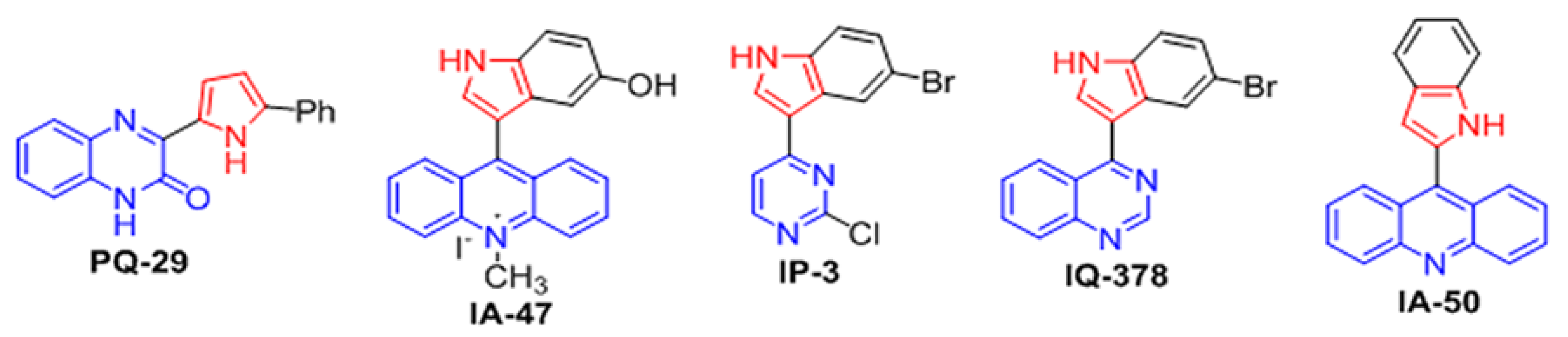
2.2. In Silico Prediction of the Indolylazine Ability to Penetrate the Cell Membrane of Senescent Neurons
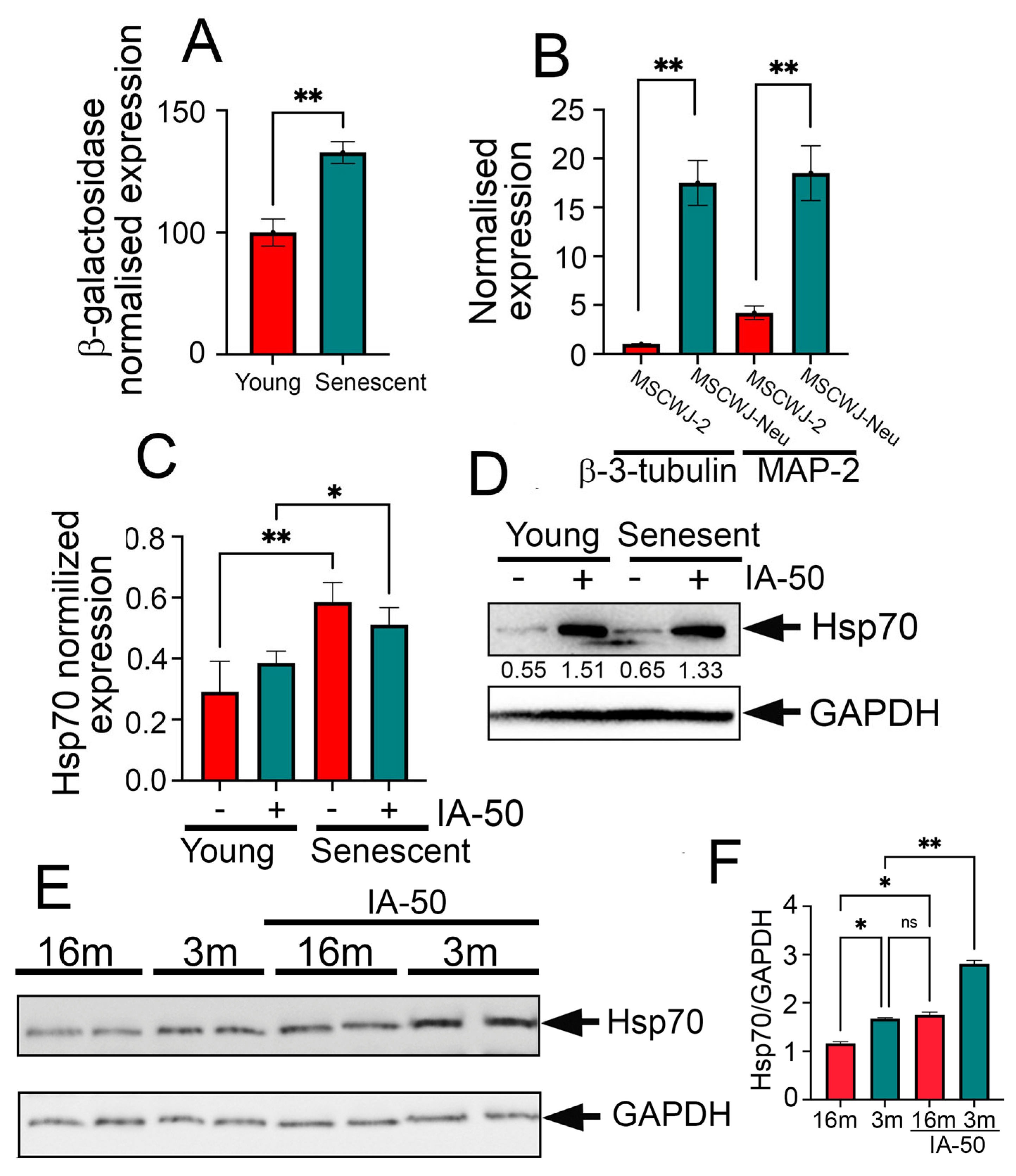
2.3. IA-50 Is Able to Induce Chaperone Accumulation in Aged Neurons
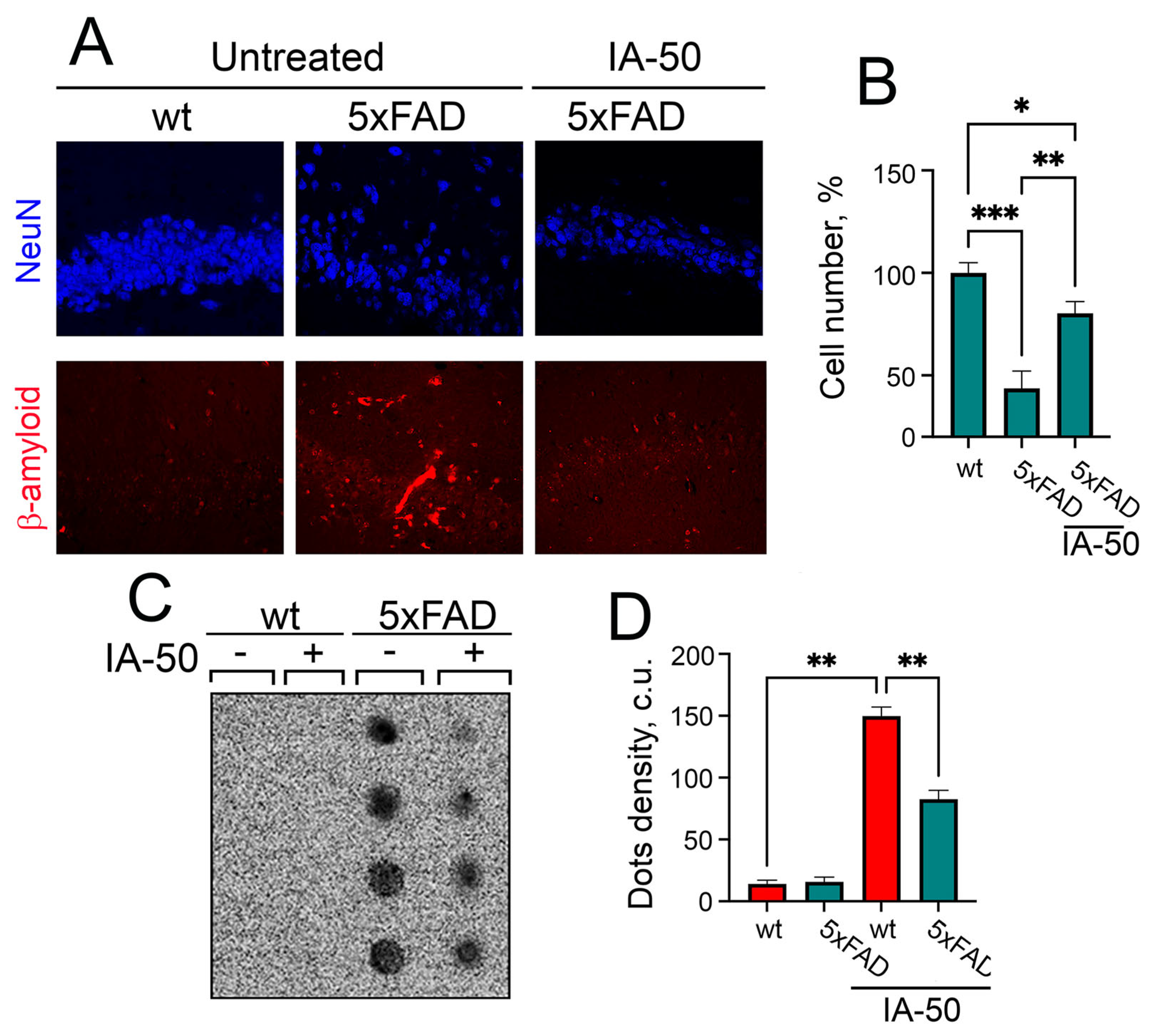
2.4. The Treatment of 5xFAD Mice with IA-50 Improves Memory Defects and Aβ42 Aggregation
3. Discussion
4. Materials and Methods
4.1. In Silico Assays
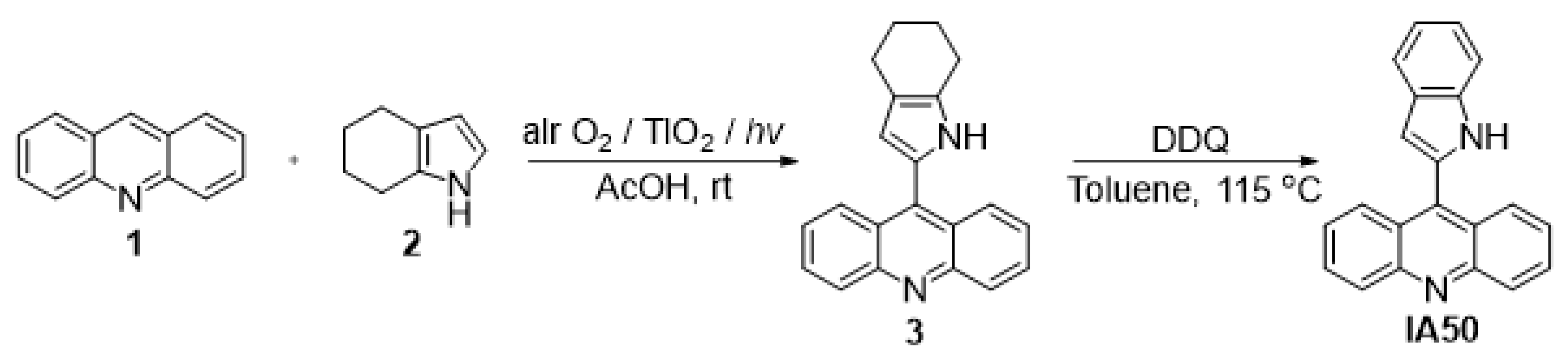
4.2. Synthesis of 9-(1H-indol-2-yl)acridine (IA-50)
4.3. Mimicking the Neuronal Cell Phenotype
4.4. RNA Isolation and Real-Time PCR
4.5. Electrophoresis and Immunoblotting
4.6. Senescence Analysis
4.7. Transgenic Mice Model of AD
4.8. Animal Behaviour Tests
4.8.1. Marble Burying Assay
4.8.2. Morris Water Test
4.9. Immunohistochemistry
4.10. Aggregation Assay
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karran, E.; Mercken, M.; De Strooper, B. The amyloid cascade hypothesis for Alzheimer’s disease: An appraisal for the development of therapeutics. Nat. Rev. Drug Discov. 2011, 10, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Shepardson, N.; Yang, T.; Chen, G.; Walsh, D.; Selkoe, D.J. Soluble amyloid beta-protein dimers isolated from Alzheimer cortex directly induce Tau hyperphosphorylation and neuritic degeneration. Proc Natl Acad Sci USA 2011, 108, 5819–5824. [Google Scholar] [CrossRef] [PubMed]
- Cascella, R.; Bigi, A.; Cremades, N.; Cecchi, C. Effects of oligomer toxicity, fibril toxicity and fibril spreading in synucleinopathies. Cell Mol. Life Sci. 2022, 79, 174. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Mahanta, S. Association of heat-shock proteins in various neurodegenerative disorders: Is it a master key to open the therapeutic door? Mol. Cell Biochem. 2014, 386, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Lazarev, V.F.; Mikhaylova, E.R.; Guzhova, I.V.; Margulis, B.A. Possible Function of Molecular Chaperones in Diseases Caused by Propagating Amyloid Aggregates. Front Neurosci. 2017, 11, 277. [Google Scholar] [CrossRef]
- Lackie, R.E.; Maciejewski, A.; Ostapchenko, V.G.; Marques-Lopes, J.; Choy, W.Y.; Duennwald, M.L.; Prado, V.F.; Prado, M.A.M. The Hsp70/Hsp90 Chaperone Machinery in Neurodegenerative Diseases. Front Neurosci. 2017, 11, 254. [Google Scholar] [CrossRef]
- Shelton, L.B.; Koren, J., 3rd; Blair, L.J. Imbalances in the Hsp90 Chaperone Machinery: Implications for Tauopathies. Front Neurosci 2017, 11, 724. [Google Scholar] [CrossRef]
- Neef, D.W.; Jaeger, A.M.; Thiele, D.J. Heat shock transcription factor 1 as a therapeutic target in neurodegenerative diseases. Nat. Rev. Drug Discov. 2011, 10, 930–944. [Google Scholar] [CrossRef]
- Margulis, B.; Tsimokha, A.; Zubova, S.; Guzhova, I. Molecular Chaperones and Proteolytic Machineries Regulate Protein Homeostasis In Aging Cells. Cells 2020, 9, 1308. [Google Scholar] [CrossRef]
- Blake, M.J.; Fargnoli, J.; Gershon, D.; Holbrook, N.J. Concomitant decline in heat-induced hyperthermia and HSP70 mRNA expression in aged rats. Am. J. Physiol. 1991, 260, R663–R667. [Google Scholar] [CrossRef]
- Faassen, A.E.; O’Leary, J.J.; Rodysill, K.J.; Bergh, N.; Hallgren, H.M. Diminished heat-shock protein synthesis following mitogen stimulation of lymphocytes from aged donors. Exp. Cell Res. 1989, 183, 326–334. [Google Scholar] [CrossRef]
- Heydari, A.R.; Wu, B.; Takahashi, R.; Strong, R.; Richardson, A. Expression of heat shock protein 70 is altered by age and diet at the level of transcription. Mol. Cell Biol. 1993, 13, 2909–2918. [Google Scholar]
- Pahlavani, M.A.; Harris, M.D.; Moore, S.A.; Weindruch, R.; Richardson, A. The expression of heat shock protein 70 decreases with age in lymphocytes from rats and rhesus monkeys. Exp. Cell Res. 1995, 218, 310–318. [Google Scholar] [CrossRef]
- Rea, I.M.; McNerlan, S.; Pockley, A.G. Serum heat shock protein and anti-heat shock protein antibody levels in aging. Exp. Gerontol. 2001, 36, 341–352. [Google Scholar] [CrossRef]
- Jin, X.; Wang, R.; Xiao, C.; Cheng, L.; Wang, F.; Yang, L.; Feng, T.; Chen, M.; Chen, S.; Fu, X.; et al. Serum and lymphocyte levels of heat shock protein 70 in aging: A study in the normal Chinese population. Cell Stress Chaperones 2004, 9, 69–75. [Google Scholar] [CrossRef]
- Calabrese, V.; Butterfield, D.A.; Scapagnini, G.; Stella, A.M.; Maines, M.D. Redox regulation of heat shock protein expression by signaling involving nitric oxide and carbon monoxide: Relevance to brain aging, neurodegenerative disorders, and longevity. Antioxid. Redox Signal. 2006, 8, 444–477. [Google Scholar] [CrossRef]
- Gleixner, A.M.; Pulugulla, S.H.; Pant, D.B.; Posimo, J.M.; Crum, T.S.; Leak, R.K. Impact of aging on heat shock protein expression in the substantia nigra and striatum of the female rat. Cell Tissue Res 2014, 357, 43–54. [Google Scholar] [CrossRef]
- Dutysheva, E.A.; Utepova, I.A.; Trestsova, M.A.; Anisimov, A.S.; Charushin, V.N.; Chupakhin, O.N.; Margulis, B.A.; Guzhova, I.V.; Lazarev, V.F. Synthesis and approbation of new neuroprotective chemicals of pyrrolyl- and indolylazine classes in a cell model of Alzheimer’s disease. Eur. J. Med. Chem. 2021, 222, 113577. [Google Scholar] [CrossRef]
- Peetla, C.; Vijayaraghavalu, S.; Labhasetwar, V. Biophysics of cell membrane lipids in cancer drug resistance: Implications for drug transport and drug delivery with nanoparticles. Adv. Drug Deliv. Rev. 2013, 65, 1686–1698. [Google Scholar] [CrossRef]
- Das, U.N. “Cell Membrane Theory of Senescence” and the Role of Bioactive Lipids in Aging, and Aging Associated Diseases and Their Therapeutic Implications. Biomolecules 2021, 11, 241. [Google Scholar] [CrossRef]
- Ertl, P.; Rohde, B.; Selzer, P. Fast calculation of molecular polar surface area as a sum of fragment-based contributions and its application to the prediction of drug transport properties. J. Med. Chem. 2000, 43, 3714–3717. [Google Scholar] [CrossRef] [PubMed]
- Oakley, H.; Cole, S.L.; Logan, S.; Maus, E.; Shao, P.; Craft, J.; Guillozet-Bongaarts, A.; Ohno, M.; Disterhoft, J.; Van Eldik, L.; et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer’s disease mutations: Potential factors in amyloid plaque formation. J. Neurosci. 2006, 26, 10129–10140. [Google Scholar] [CrossRef] [PubMed]
- Devi, L.; Ohno, M. Phospho-eIF2α level is important for determining abilities of BACE1 reduction to rescue cholinergic neurodegeneration and memory defects in 5XFAD mice. PLoS ONE 2010, 5, e12974. [Google Scholar] [CrossRef]
- Tanaka, H.; Hashimoto, M.; Fukuhara, R.; Ishikawa, T.; Yatabe, Y.; Kaneda, K.; Yuuki, S.; Honda, K.; Matsuzaki, S.; Tsuyuguchi, A.; et al. Relationship between dementia severity and behavioural and psychological symptoms in early-onset Alzheimer’s disease. Psychogeriatrics 2015, 5, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Baillon, S.; Gasper, A.; Wilson-Morkeh, F.; Pritchard, M.; Jesu, A.; Velayudhan, L. Prevalence and Severity of Neuropsychiatric Symptoms in Early- Versus Late-Onset Alzheimer’s Disease. Am. J. Alzheimers Dis. Other Demen 2019, 34, 433–438. [Google Scholar] [CrossRef]
- Njung’e, K.; Handley, S.L. Evaluation of marble-burying behavior as a model of anxiety. Pharmacol. Biochem. Behav. 1991, 38, 63–67. [Google Scholar] [CrossRef]
- Kaurav, B.P.; Wanjari, M.M.; Chandekar, A.; Chauhan, N.S.; Upmanyu, N. Influence of Withania somnifera on obsessive compulsive disorder in mice. Asian. Pac. J. Trop. Med. 2012, 5, 380–384. [Google Scholar] [CrossRef]
- Torres-Lista, V.; López-Pousa, S.; Giménez-Llort, L. Impact of Chronic Risperidone Use on Behavior and Survival of 3xTg-AD Mice Model of Alzheimer’s Disease and Mice With Normal Aging. Front. Pharmacol. 2019, 10, 1061. [Google Scholar] [CrossRef]
- Kim, T.K.; Han, H.E.; Kim, H.; Lee, J.E.; Choi, D.; Park, W.J.; Han, P.L. Expression of the plant viral protease NIa in the brain of a mouse model of Alzheimer’s disease mitigates Aβ pathology and improves cognitive function. Exp. Mol. Med. 2012, 44, 740–748. [Google Scholar] [CrossRef]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. In vivo aspects of protein folding and quality control. Science 2016, 353, aac4354. [Google Scholar] [CrossRef]
- Ciechanover, A.; Kwon, Y.T. Protein Quality Control by Molecular Chaperones in Neurodegeneration. Front. Neurosci. 2017, 11, 185. [Google Scholar] [CrossRef]
- Chung, L.; Ng, Y.C. Age-related alterations in expression of apoptosis regulatory proteins and heat shock proteins in rat skeletal muscle. Biochim. Biophys. Acta 2006, 1762, 103–109. [Google Scholar] [CrossRef]
- Hubert, V.; Weiss, S.; Rees, A.J.; Kain, R. Modulating Chaperone-Mediated Autophagy and Its Clinical Applications in Cancer. Cells 2022, 11, 2562. [Google Scholar] [CrossRef]
- Catarino, S.; Pereira, P.; Girão, H. Molecular control of chaperone-mediated autophagy. Essays Biochem. 2017, 61, 663–674. [Google Scholar]
- Pajouhesh, H.; Lenz, G.R. Medicinal chemical properties of successful central nervous system drugs. NeuroRx 2005, 2, 541–553. [Google Scholar] [CrossRef]
- Bagdi, A.K.; Rahman, M.; Bhattacherjee, D.; Zyryanov, G.V.; Ghosh, S.; Chupakhin, O.N.; Hajra, A. Visible light promoted cross-dehydrogenative coupling: A decade update. Green Chem. 2020, 22, 6632–6681. [Google Scholar] [CrossRef]
- Narayanam, J.M.R.; Stephenson, C.R.J. Visible light photoredox catalysis: Applications in organic synthesis. Chem. Soc. Rev. 2011, 40, 102–113. [Google Scholar] [CrossRef]
- Utepova, I.A.; Chupakhin, O.N.; Trestsova, M.A.; Musikhina, A.A.; Kucheryavaya, D.A.; Charushin, V.N.; Rempel, A.A.; Kozhevnikova, N.S.; Valeeva, A.A.; Mikhaleva, A.I.; et al. Direct functionalization of the C-H bond in (hetero)arenes: Aerobic photoinduced oxidative coupling of azines with aromatic nucleophiles (SNH-reactions) in the presence of a CdS/TiO2 photocatalyst. Russ. Chem. Bull 2016, 65, 445–450. [Google Scholar] [CrossRef]
- Rae, E.A.; Brown, R.E. The problem of genotype and sex differences in life expectancy in transgenic AD mice. Neurosci. Biobehav. Rev. 2015, 57, 238–251. [Google Scholar] [CrossRef]
- Kalmar, B.; Greensmith, L. Activation of the heat shock response in a primary cellular model of motoneuron neurodegeneration-evidence for neuroprotective and neurotoxic effects. Cell Mol. Biol. Lett. 2009, 14, 319–335. [Google Scholar] [CrossRef]
- Sugano, E.; Endo, Y.; Sugai, A.; Kikuchi, Y.; Tabata, K.; Ozaki, T.; Kurose, T.; Takai, Y.; Mitsuguchi, Y.; Honma, Y.; et al. Geranylgeranyl acetone prevents glutamate-induced cell death in HT-22 cells by increasing mitochondrial membrane potential. Eur. J. Pharmacol. 2020, 883, 173193. [Google Scholar] [CrossRef] [PubMed]
- Ekimova, I.V.; Plaksina, D.V.; Pastukhov, Y.F.; Lapshina, K.V.; Lazarev, V.F.; Mikhaylova, E.R.; Polonik, S.G.; Pani, B.; Margulis, B.A.; Guzhova, I.V.; et al. New HSF1 inducer as a therapeutic agent in a rodent model of Parkinson’s disease. Exp. Neurol. 2018, 306, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, T.; Roth, A.G.; Petersen, N.H.; Mahalka, A.K.; Olsen, O.D.; Moilanen, I.; Zylicz, A.; Knudsen, J.; Sandhoff, K.; Arenz, C.; et al. Hsp70 stabilizes lysosomes and reverts Niemann-Pick disease-associated lysosomal pathology. Nature 2010, 463, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Trofimov, B.A.; Mikhaleva, A.I.; Schmidt, E.Y.; Vasil’tsov, A.M.; Ivanov, A.V.; Protsuk, N.I.; Ryapolov, O.A. A new technology for the synthesis of 4,5,6,7-tetrahydroindole. Dokl. Chem. 2010, 435, 307–310. [Google Scholar] [CrossRef]
- Koltsova, A.M.; Krylova, T.A.; Musorina, A.S.; Zenin, V.V.; Turilova, V.I.; Yakovleva, T.R.; Poljanskaya, G.G. The Dynamics of Cell Properties during Long-Term Cultivation of Two Lines of Mesenchymal Stem Cells Derived from Wharton’s Jelly of Human Umbilical Cord. Cell Tissue Biol. 2018, 121, 7–19. [Google Scholar] [CrossRef]
- Guzhova, I.; Kislyakova, K.; Moskaliova, O.; Fridlanskaya, I.; Tytell, M.; Cheetham, M.; Margulis, B. In vitro studies show that Hsp70 can be released by glia and that exogenous Hsp70 can enhance neuronal stress tolerance. Brain Res. 2001, 914, 66–73. [Google Scholar] [CrossRef]
- Peters, O.M.; Shelkovnikova, T.; Tarasova, T.; Springe, S.; Kukharsky, M.S.; Smith, G.A.; Brooks, S.; Kozin, S.A.; Kotelevtsev, Y.; Bachurin, S.O.; et al. Chronic administration of Dimebon does not ameliorate amyloid-β pathology in 5xFAD transgenic mice. J. Alzheimer’s Dis. 2013, 36, 589–596. [Google Scholar] [CrossRef]
- Broekkamp, C.L.; Rijk, H.W.; Joly-Gelouin, D.; Lloyd, K.L. Major tranquillizers can be distinguished from minor tranquillizers on the basis of effects on marble burying and swim-induced grooming in mice. Eur. J. Pharmacol. 1986, 126, 223–229. [Google Scholar] [CrossRef]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Sankowski, R.; Huerta, T.S.; Kalra, R.; Klein, T.J.; Strohl, J.J.; Al-Abed, Y.; Robbiati, S.; Huerta, P.T. Large-Scale Validation of the Paddling Pool Task in the Clockmaze for Studying Hippocampus-Based Spatial Cognition in Mice. Front. Behav. Neurosci. 2019, 13, 121. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Dutysheva, E.A.; Komarova, E.Y.; Mikhaylova, E.R.; Guzhova, I.V.; Margulis, B.A. GAPDH-targeted therapy—A new approach for secondary damage after traumatic brain injury on rats. Biochem. Biophys. Res. Commun. 2018, 501, 1003–1008. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PQ-29 | IA-47 | IP-3 | IQ-378 | IA-50 | |
|---|---|---|---|---|---|
| Log Po/w | 3.24 | 3.34 | 3.33 | 3.72 | 3.78 |
| Log S | −4.10 | −6.53 | −4.2 | −4.6 | −4.38 |
| NRB | 2 | 1 | 1 | 1 | 1 |
| Number H-bond acceptors | 2 | 1 | 2 | 2 | 1 |
| Nummer H-bond donors | 2 | 2 | 1 | 1 | 1 |
| TPSA | 61.54 | 39.90 | 41.57 | 41.57 | 28,15 |
| Molecular weight | 287.32 | 452.30 | 308.57 | 324.17 | 296.35 |
| Gastrointestinal absorption | High | High | High | High | High |
| BBB permeability | Yes | Yes | Yes | Yes | Yes |
| GAPDH | Forward | 5′-ATGATTCTACCCACGGCAAG-3′ |
| Reverse | 5′-CTGGAAGATGGTGATGGGTT-3′ | |
| HSPA1A (Hsp70) | Forward | 5′-CAAGAATGCGCTCGAGTCCTA-3′ |
| Reverse | 5′-GGAGATGACCTCCTGGCACTT-3′ | |
| MAP-2 | Forward | 5′-CGCTAAATCGTAAGTGAGGGCT-3′ |
| Reverse | 5′-ATTAGAAGTCCCCGCAGTGG-3′ | |
| β-3-tubulin | Forward | 5′-AGCAAGAACAGCAGCTACTTCGT-3′ |
| Reverse | 5′-GATGAAGGTGGAGGACATCTTGA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazarev, V.F.; Dutysheva, E.A.; Mikhaylova, E.R.; Trestsova, M.A.; Utepova, I.A.; Chupakhin, O.N.; Margulis, B.A.; Guzhova, I.V. Indolylazine Derivative Induces Chaperone Expression in Aged Neural Cells and Prevents the Progression of Alzheimer’s Disease. Molecules 2022, 27, 8950. https://doi.org/10.3390/molecules27248950
Lazarev VF, Dutysheva EA, Mikhaylova ER, Trestsova MA, Utepova IA, Chupakhin ON, Margulis BA, Guzhova IV. Indolylazine Derivative Induces Chaperone Expression in Aged Neural Cells and Prevents the Progression of Alzheimer’s Disease. Molecules. 2022; 27(24):8950. https://doi.org/10.3390/molecules27248950
Chicago/Turabian StyleLazarev, Vladimir F., Elizaveta A. Dutysheva, Elena R. Mikhaylova, Maria A. Trestsova, Irina A. Utepova, Oleg N. Chupakhin, Boris A. Margulis, and Irina V. Guzhova. 2022. "Indolylazine Derivative Induces Chaperone Expression in Aged Neural Cells and Prevents the Progression of Alzheimer’s Disease" Molecules 27, no. 24: 8950. https://doi.org/10.3390/molecules27248950
APA StyleLazarev, V. F., Dutysheva, E. A., Mikhaylova, E. R., Trestsova, M. A., Utepova, I. A., Chupakhin, O. N., Margulis, B. A., & Guzhova, I. V. (2022). Indolylazine Derivative Induces Chaperone Expression in Aged Neural Cells and Prevents the Progression of Alzheimer’s Disease. Molecules, 27(24), 8950. https://doi.org/10.3390/molecules27248950