1. Introduction
During the last decade, functional foods have gained attention from the food industry because of their possible role in the maintenance of health and wellbeing [
1]. Grain pulses, protein rich and affordable foods, are grown both for seed production and consumption and contribute to a balanced diet. Among grain pulses, lupins have recently gained considerable interest for a number of attractive nutritional attributes relating to their high protein and dietary fiber and negligible starch contents [
2]. Lupin seeds are a food ingredient characterized by good nutritional value and technological flexibility. Their high protein content makes lupin seeds an alternative protein source that can substitute for soy, egg, and other animal proteins, and recent clinical investigations have indicated that the consumption of lupins may be beneficial due to their hypocholesterolemic [
3,
4,
5], anti-atherogenic [
6], hypotensive [
7], and hypoglycemic activities [
8,
9].
The genus
Lupinus comprises more than 170 species that have been described in the Old World and only 12 species from Europe [
10]. Domesticated lupins include only four species:
L. albus L. (white lupin), originating from the Mediterranean costal region and grown in the Mediterranean area and Southern Europe;
L. angustifolius L. (narrow-leafed lupin, blue lupin), widely grown in South-Western Australia;
L. luteus L. (yellow lupin) grown in Central Europe; and
L. mutabilis (Tarwe, Andean, or Pearl lupin), originating from South America and grown in the Andes (Chile and Peru). White lupin is currently the only species grown for human nutrition, but narrow-leafed lupin is increasingly used for animal feed, as well as human food.
Certain elements are essential for plant growth and animal and human health. However, if present in excessive concentrations, they become toxic. Zelalem and Chandravanshi [
11] determined the concentration of macro- (K, Na, Mg, Ca); micro- (Cr, Mn, Fe, Co, Ni, Cu, Zn); and toxic (Pb, Cd) elements in dry raw and processed white lupin samples grown in Ethiopia. The authors found that the concentrations of all analyzed elements were higher in raw compared to processed white lupin samples, except for Ca, Co, Cu, and Zn, which appeared at higher concentrations in the processed samples than the raw white lupin samples from all of the sampling sites. The authors concluded that raw (dried) and processed
Lupinus albus could be good sources of major, minor, and trace elements essential to humans. Other authors [
12] studied yellow and blue lupin genotypes and found the highest contents of Mn, Fe, Zn, and Se in
Lupinus luteus seeds. The authors concluded that the concentration of nonessential elements in the tested lupin seeds was far below the levels that would cause a health risk.
Lupin seeds are commonly used for dietary consumption due to their valuable traits, such as their high protein content, beneficial relative protein profile, high dietary fiber content, and favorable fat content. Although most of the studies available in the literature have focused on the seeds’ protein quality, attention has also been paid to their oil content and quality. Several studies have shown that the amount of oil in lupin seeds varies depending on the genotype. Andean lupin showed a considerable amount of oil in its seeds, ranging from 16 to 20% [
13], while lower amounts (9.76–12.1%) were recorded for white lupin. The oil contents in narrow-leafed and yellow lupin [
12] were found to be less representative, at 6.5 and 5.1%, respectively [
14]. The quality rather than the quantity of oil contained in lupin seeds is much more relevant in terms of dietary purposes and applications for both humans and animals. In general, the fatty acid profile and the ratios between individual acids are used as markers of the overall seed oil quality. It has previously been demonstrated that lupin seed oil has a balanced fatty acid composition, with an average total unsaturated fatty acid percentage of 90% [
15] and interesting phytosterol, triterpene alcohol, and phospholipid contents [
16]. It must be pointed out that the quali-quantitative composition of the seed oil can vary greatly depending on factors such as the genotype, ripening phase, growing environment, pedoclimatic conditions, cultural practices, and extraction technology and/or solvents used. In the past decade, several studies have been performed to determine the effects of genotype and environment on lupin seed oil quality [
16,
17,
18,
19,
20,
21,
22]; however, to the best of our knowledge, no data are available on the effects of the extraction technology on the oil yield. Supercritical carbon dioxide (SC-CO
2) has been employed to extract seed oils with encouraging results for several decades [
23,
24,
25], and it is widely recognized as a green and environmentally friendly alternative to organic solvent extraction for the production of vegetable fats and oils. Taking into consideration the fact that lupin seeds are recognized as a protein-rich crop and their use as a dietary protein source is of increasing interest, the employment of SC-CO
2 extraction as a defatting process could address consumers’ demands for natural proteins isolated from natural resources through chemical-free processes.
Lupin seed proteins are classified as albumins (water-soluble), globulins (salt-soluble), prolamins (alcohol-soluble), and glutelins (acid/alkali-soluble). Globulins (frequently referred to as conglutins, including in the remainder of this paper) constitute the major fraction of lupin seed proteins, followed by albumins (the globulin-to-albumin ratio is 9:1), while prolamins and glutelins contribute a negligible amount. Based on the sedimentation coefficient, lupin seed storage proteins are categorized as 2S (generally termed albumins), 7S, and 11S (sub-classes of conglutins). Conglutins are classified into four families: α-, β-, γ-, and δ-conglutins. α-conglutins, members of the “legumin-like” or 11S globulin family, comprise hexamers of high molecular weight (acidic subunits), belonging to the N-terminal, and low molecular weight (basic subunits), related to the C-terminal region. These are linked by disulphide bonds [
26], with the high molecular weight subunits being glycosylated [
27]. Β-conglutins, also referred to as “vicilin-like” or 7S globulins, are trimeric proteins of 16–70 kDa monomers, characterized by the absence of any disulphide linkage and a great extent of glycosylation. Both α- and β-conglutins originate from proteolytic cleavages of precursor molecules. Γ-conglutins, accounting for about 5% of the total protein content, may not fall into the classical seed storage category, as these proteins are not cleaved during seed germination [
28]. Lupin γ-conglutin, a 7S protein, is a homo-tetrameric basic glycoprotein with a relative molecular mass of around 47 kDa, whose monomeric units consist of two disulphides linked to heterogenous subunits of 17 and 29 kDa [
29]. γ-conglutin displays unique properties: since its amino acid sequence does not match any other legume protein canonical sequence [
30], it binds divalent metal ions, especially Zn
2+ and Ni
2+ [
31], and carries one N-linked oligosaccharide chain [
32]. The last group of the lupin seed conglutin family, δ-conglutins, has been the least studied: these are small proteins, similar to other 2S sulphur-rich albumins, comprising two chains of 4 and 9 kDa linked by disulphide bridges. Recently, the complexity of these four conglutin families has been investigated in different lupin species in terms of their gene structure, phylogenetic relationship, and relative RNA and protein abundance during seed development: the results of this investigation demonstrated differential conglutin RNA expression across lupin species with a high level of γ-conglutins in
Lupinus albus and
L. cosentinii [
33]. γ-conglutins play an important role due to their health-promoting benefits. It has been demonstrated that purified or enriched γ-conglutin lowered blood glucose in hyperglycemic rats [
34] and had a substantial hypoglycemic effect in a glucose-overload trial in healthy humans and rats [
35]. γ-conglutin is therefore a potential antidiabetic agent [
34], and a patent has been deposited for its use in the treatment of type II diabetes [
36]. The molecular characterization of bioactive food components is necessary for understanding the mechanisms of their beneficial or detrimental effects on human health, and mass spectrometry is the leading technique for tracing and quantifying the differential expression of specific proteins such as bioactive or allergenic proteins. The knowledge of the protein composition is helpful for understanding the relationship between the protein content and the nutritional and technological properties of a foodstuff [
37], and mass spectrometry coupled to electrophoresis and chromatographic separation is the method of choice for protein food analysis [
38,
39,
40].
The present study comprised a comparative multielemental, nutritional, and proteomic characterization of different genotypes of Lupinus ssp. (L. albus, L. angustifolius, and L. luteus) grown in the same environmental conditions, with respect to a commercial lupin flour (L. albus, cv. Multitalia) available on the food and feed market. The considerable diversity of the Lupinus genus is still widely unexplored and unexploited regarding its potential for improving health and wellbeing. Useful data for the optimization of the SC-CO2 extraction process to obtain lupin seed oil and protein isolates of good quality are reported. The fatty acid and tocopherol profiles of the obtained oils are also presented herein in order to promote their use as an ingredient for human and animal diets. Moreover, physiological and health-promoting effects are highly influenced by protein structure. Therefore, an analysis of the diversity of lupin seed proteins was conducted to provide a deeper understanding of the relationships between structure and bioactive properties in order to improve lupin seeds’ nutraceutical potential. For this reason, a proteomic approach involving 2D-PAGE separations and mass spectrometric analysis was performed to obtain the protein profiles of the different genotypes and to evaluate the presence of protein groups characteristic of specific genotypes that might be employed as markers in future investigations.
3. Materials and Methods
3.1. Plant Material
Seeds of Lupinus albus (cv. Luxor, cv. Multitalia, and Modica ecotype); L. luteus (cv. Dukat, Mister, and Taper); and L. angustifolius (cv. Sonet) were deposited within the germplasm collection of the Research Centre for Cereal and Industrial Crops of Acireale (Catania, Italy). The field trial was conducted from 2013 to 2014 in East Sicily (Acireale, Catania, Italy) in the ‘S. Salvatore’ Experimental Station of CREA.
All seed samples were sowed in duplicate plots of 5 m2 (2.5 m × 2.0 m). Manual seeding was completed on 4 December 2013. Fertilization was applied during sowing with the fertilizer ‘Ortofrutta’, containing 12 kg ha−1 of N, 12 kg ha−1 of P2O5, and 12 kg ha−1 of K2O (Adriatica S.p.A. Loreo, Rovigo, Italy).
Mechanical weed control was applied in post-emergence at the end of January. Aphicide treatment with 50 mL/hL of Imidacloprid (Afidane, Chimiberg-Diachem S.p.A. Caravaggio, Bergamo, Italy) was applied on 3 March 2014.
The crop was harvested on 15 July 2014.
3.2. Chemicals
Formic acid (FA), dithiothreitol (DTT), iodoacetamide (IAA), Tris-HCl, urea, glycerol, sodium dodecyl sulfate (SDS), bromophenol blue, phosphoric acid, and ammonium sulfate were obtained from Sigma-Aldrich (Milan, Italy). Colloidal Coomassie Brilliant Blue G-250 was purchased from Amresco, Solon (OH), USA. Modified porcine trypsin was purchased from Promega (Milan, Italy). Water and acetonitrile for LC/MS analyses were provided from Carlo Erba (Milan, Italy).
3.3. Milling of Lupin Seeds
Lupin seeds were first broken by an experimental mill, Cyclotec type 120 (Falling Number, Huddinge, Sweden), with a sieve of 0.5 mm, and then the broken seeds were finely ground by a Bimby TM6 Vorwerk Thermomix Robot.
3.4. Multielemental Micro- (Fe, Zn, Mn, Cu) and Macronutrient (P, K, Ca, Mg, Na) Profiles
The seeds were milled and homogenized (final particle size ≤ 200 µm). For sample preparation for ICP-OES analysis, 0.5 g of milled lupine seeds to the nearest 1 mg was accurately weighed in a microwave vessel. For trace-element analysis, 2 mL of deionized water, 8 mL of nitric acid (concentration ≥ 69.0%), and 2 mL of extra-pure hydrogen peroxide (30% w/w) were added, and the samples were left for 12 h for reaction stabilization until the formation of bubbles had finished. The vessel was sealed and heated in the microwave system. A CEM MARS6 instrument was used for microwave digestion. The following thermal conditions were applied: 150 °C was reached in approx. 20 min and maintained for 30 min, and then 200 °C was reached in approx. 20 min and maintained for 30 min for the accomplishment of specific reactions. After cooling, the prepared solution was filtered through a filter with a 0.45 μm pore size. The solution was transferred to a 25 mL volumetric flask and supplemented with water to reach a 25 mL volume. Optima 2100 DV ICP/OES (PerkinElmer) and WinLab32 software for ICP/OES version 3.4.1.0271 (PerkinElmer) were used for analysis. The following operating conditions in Optima 2100 DV ICP/OES were used for the analysis of the samples: spray chamber type—cyclonic; plasma aerosol type—wet; sample flow rate—1.50 mL/min; plasma—15 L/min; aux—0.2 L/min; nebulizer—0.4 L/min; power—1300 Watts.
3.5. Protein Content
Nitrogen content was measured using the Kjeldahl method with a factor of 5.7 to determine the protein content. For sample preparation, 1.0 g of milled lupine seeds to the nearest 1 mg was accurately weighed in a sample tube with a size of 300 mL. The digestion of the samples was carried out using 15 mL sulfuric acid (concentration 95–98%) and a tablet of copper (II) sulfate as a catalyst, which oxidized organic matter to carbonic acid. The following thermal conditions were applied: 390 °C was reached in approx. 60 min and maintained for 180 min. A SpeedDigester K-439 (Buchi) instrument was used for the digestion of samples. Noxious fumes generated during digestion were captured by a sealed suction system and neutralized by the optionally available K-415 Scrubber (Buchi). After cooling, the solution in the digestion tube was then made alkaline by the addition of 90 mL of sodium hydroxide (32% w/w), which converted ammonium sulfate to ammonia gas. Ammonia was released and distilled into an excess (50 mL) of boric acid solution (4% w/w) adjusted to pH 4.65. The nitrogen content was then estimated by the titration of the ammonium borate formed with a standardized sulfuric acid solution (0.2 N) using a suitable indicator to determine the end point of the reaction. A KjelFlex K-360 distillation unit (Buchi) and KjelFlex K-360 ver 01.03 software were used to distil ammonia.
3.6. SC-CO2 Defatting Process
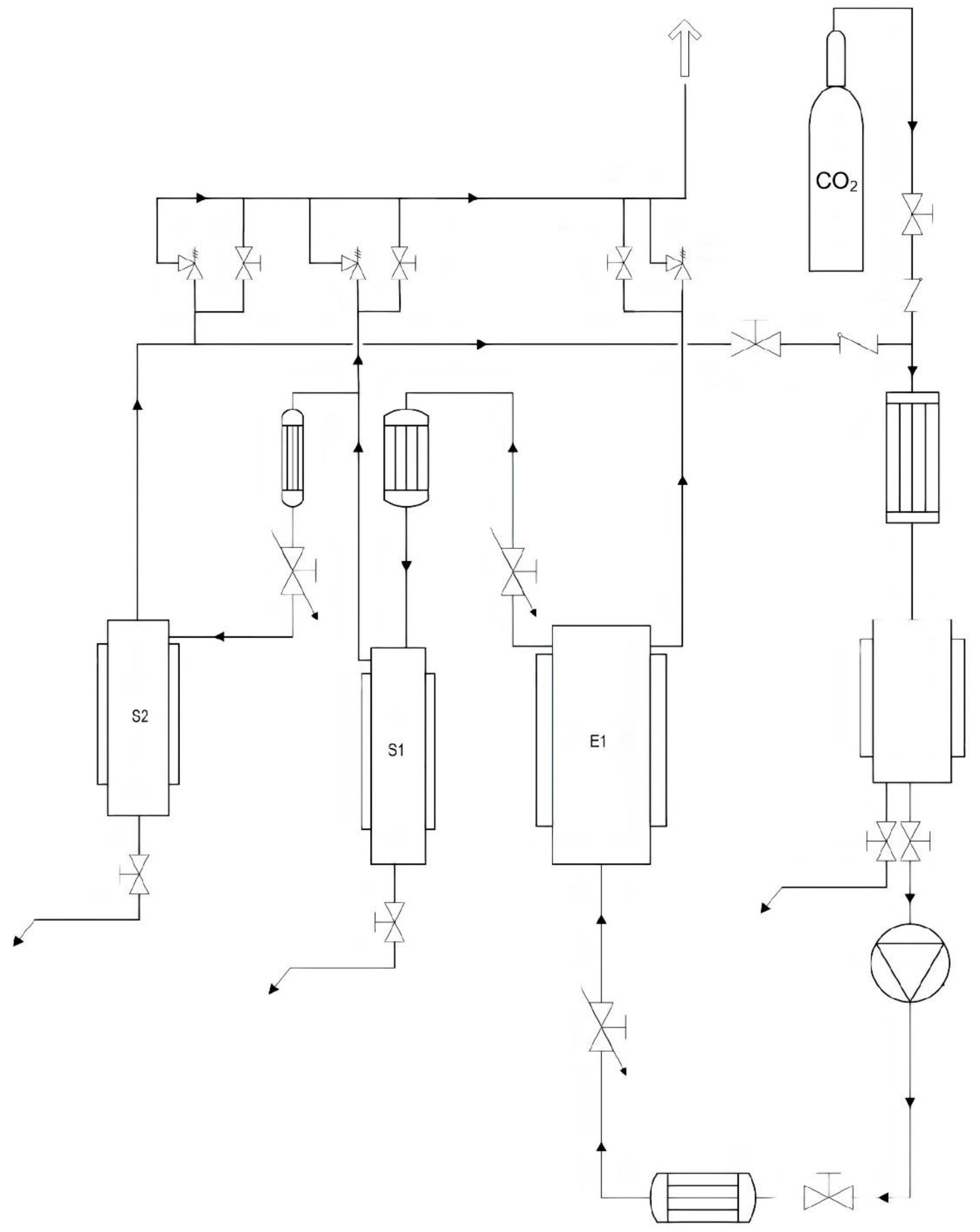
The employed supercritical fluid extraction (SFE) equipment is located at the technological pavilion of the CREA Research Center for Olive, Fruit, and Citrus Crops (Acireale, CT, Italy) and is entirely composed of AISI 316 L steel (Separeco s.r.l., Pinerolo, TO, Italy) according to the European Commission regulations for pressure systems (PED—Pressure Equipment Directory).
Figure 4 shows a scheme of the plant. It consists of an extraction reactor (E1) and two separators (S1 and S2). The reactor has a volume of 1 L, while the separators have a volume of about 0.2 L each. The CO
2 pump has a max flow rate of 5.78 L/h. The operating pressures can vary from 60 to 300 bar. The safety valves are therefore calibrated for opening above 330 bar. The system is thermostated using boiler and chiller integrated systems. Full-fat lupin seeds belonging to the different genotypes were previously oven dried at a mild temperature (<45 °C) and further milled to a fine powder by a domestic grinder so that the final particle size was ≤150 µm. Prior to the SC-CO
2 defatting process, an aliquot of 40 g of dried full-fat lupin seed flour of the different genotypes was used to determine the oil fraction by Soxhlet extraction for 5 h using hexane as the solvent. Then, the obtained lupin seed flour samples were lyophilized before being subjected to the defatting process by SC-CO
2. In order to improve the oil extraction yield, about 300 g of the lyophilized samples was manually blended with ethanol as an entrainer at a rate of 15% (
w/
w), before being placed in E1, previously set at the desired temperature (T = 40 ± 1 °C). CO
2 (99.95% grade purity) was subcooled in a cooling system in order to be pumped as a liquid by the pump (Lewa, max flow 5.78 L/h) and then directed into E1 until the desired pressure was reached (P = 300 bar). SC-CO
2 was allowed to flow continuously through E1 for 6 h at a rate of 5.176 kg/h. The oil was collected in a flask immersed in an ice bath at around 5 °C.
3.7. Fatty Acid Analysis
Fatty acid profile of lupin oil samples was determined by direct base-catalyzed transesterification and by gas-chromatography (GC). In brief, 50 mg of oil were converted to fatty acid methyl esters (FAME) by base catalyzed transesterification using 1 mL of freshly prepared 0.5 M sodium methoxide in methanol for 3 min at room temperature followed by 5 min of rest. Then, 2 mL of hexane were added and the supernatant was collected and placed in a 2 mL GC vial. The FAME were separated and quantified using a gas chromatograph (TRACE GC, Thermo Finnigan, San Jose, CA, USA) equipped with a flame-ionization detector and a 100-m fused silica capillary column (25-mm i.d., 0.25-μm film thickness; SP-2560; Supelco Inc., Bellefonte, PA, USA) and helium as the carrier gas (1 mL/min). Gas-chromatography conditions and identification of FAME was performed as reported by Natalello et al. [
50].
3.8. Tocopherol Analysis
Tocopherol concentrations in lupin oil samples were determined as described in Gliszczyńska-Świgło and Sikorska (2004) [
38] with some modifications. Briefly, 100 mg of oil was dissolved in 4 mL of 2-propanol and vortexed thoroughly for 1 min. The solution was then filtered using 0.2 µm/13 mm PTFE syringe filters, and an aliquot was placed into a 2 mL amber vial. Tocopherols were separated and quantified through ultra-high-performance liquid chromatograph (UHPLC) as detailed by Natalello et al. [
51]. A sample volume of 10 µL was injected into a Nexera (Shimadzu Corporation, Kyoto, Japan) UHPLC system equipped with a C18 phase column (Zorbax ODS; 25 cm × 4.6 mm, 5 µm; Supelco, Bellefonte, PA, USA). The mobile phase was methanol with a flow rate of 1.3 mL/min. The temperature of the samples and the column were adjusted to 25 °C and 40 °C, respectively. Tocopherols were detected using a spectrofluorometric detector at a 295 nm excitation wavelength and a 330 nm emission wavelength. The tocopherols were identified by the comparison of their retention times with those of pure standards (Merck Life Science S.r.l., Milano, Italy). The quantification was achieved by external calibration curves made with pure standards.
3.9. Protein Extraction
For two-dimensional (2D) electrophoresis, 50 mg/mL of defatted lupin flour was extracted in 8 M urea, 4% (
w/
v) CHAPS, 60 mM DTT, and 2% (
v/
v) immobilized 3–10 pH gradient (IPG) buffer (Bio Rad) at room temperature for 3 h. Extracts were centrifuged at 8000×
g for 30 min at 4 °C, and the supernatant was stored at −20 °C until use. The protein concentration was measured using the Qubit™ Protein Assay kit with the Qubit 1.0 Fluorometer (ThermoFisher Scientific, Milan, Italy) [
52].
3.10. Two-Dimensional Electrophoresis
IPG strips (pH 3, 10 NL, 11 cm (Ready Strip Bio-Rad)) were passively rehydrated o/n with 200 μg sample protein in the IPG Box (GE Healthcare, Milano, Italy). Isoelectric focusing (IEF) was performed using an Ettan IPGphor 3 Isoelectric Focusing Unit (Ettan IPGphor Manifold; GE Healthcare) for a total of 13.5 kVh, with a maximum of 6 kV, at 20 °C, 65 μA/strip. Focused IPG strips were equilibrated twice in SDS equilibration buffer (50 mM Tris-HCl pH 8.8), 6 M urea, 30% (v/v) glycerol, 2% (w/v) SDS, and 0.002% (w/v) bromophenol blue) containing 65 mM DTT for the first equilibration and 135 mM iodoacetamide for the second. SDS-PAGE was conducted on a Midi-Protean Cell (Bio-Rad) and Any kD™ Mini-Protean® TGX™ Precast gel (13.3 × 8.7 cm) (Bio-Rad, Hercules, CA, USA). Four microliters of protein molecular marker (Page Ruler unstained protein ladder, Thermo Scientific, Waltham, MA, USA) were loaded. Gels were stained using colloidal Coomassie Brilliant Blue G-250 (Amresco) in 10% phosphoric acid and 10% ammonium sulfate. Gels were fixed in ethanol, water, and acetic acid (4:5:1) for half an hour and then stained overnight in colloidal Coomassie. Gels were stored in 7% acetic acid until used for spot excision. A total of 21 2-DE gels, resulting from three technical replicates for each lupin cultivar, were analyzed. The gel images were acquired using an Epson 1680 Pro scanner and uploaded onto the Image Master 2D Platinum ver. 7 (GE Healthcare). Spots in the gels were detected automatically. Only statistically significant protein spots (p ≤ 0.01 ANOVA) were taken into consideration when evaluating the differential expression, with a two-fold change between gels considered an increase in relative abundance and a ratio of at least 0.5 considered a decrease in relative abundance. These spots were manually excised and subjected to mass spectrometric analyses. Only spots observed in all three gel sets were considered for protein identification.
3.11. In-Gel Digestion and Mass Spectrometric Analysis
Selected protein spots from the 2-DE gels were manually excised and transferred to 1.5 mL microcentrifuge tubes. Low-abundance protein spots were removed from all the replicate gels and pooled. According to the previously reported procedure, the spots were washed and subjected to in-gel trypsin digestion [
53,
54]. After soaking the gel pieces in trypsin, the supernatant containing excess trypsin was removed, and the gel pieces were covered with 50 μL of 50 mM NH
4HCO
3 pH 8.3 and incubated at 37 °C overnight. The enzymatic reaction was stopped by cooling the gel pieces and the supernatant solution at −20 °C. After in-gel digestion, the supernatant solution was transferred into a clean 1.5 mL tube. The peptides were extracted from gel pieces with 50 µL of 5% aqueous FA and subsequently with 50 μL of CH
3CN. This extraction procedure was repeated three times. The total extracts were pooled, combined with the first supernatant, lyophilized, and redissolved in 15 µL of 5% FA aqueous solution. Capillary RP-HPLC/nESI-MS/MS was performed using an Ultimate 3000 LC system combined with an autosampler and a flow splitter 1:100 (Dionex Corporation, Sunnyvale, CA, USA) coupled online with a linear ion trap nano-electrospray mass spectrometer (LTQ, Thermo Fischer Scientific, San Jose, CA, USA). Ionization was performed with liquid junction using an uncoated capillary probe (30 ± 2 µm i.d.; New Objective, Woburn, MA, USA). Ten microliters of enzymatic digest solution for each spot were loaded onto a C18 μ-precolumn cartridge (0.3 mm × 5 mm, 100 Ǻ, 5 μm, PepMap, Dionex) and equilibrated with 0.5% aqueous FA at a flow rate of 20 μL/min for 4 min. Subsequently, peptides were applied onto a C18 capillary column (0.18 mm × 150 mm, 300 Ǻ, 5 μm, Thermo Electron, Waltham, MA, USA) and eluted at room temperature with a linear gradient of CH
3CN + 0.5% FA/H
2O + 0.5% FA from 10 to 50% in 50 min at a flow rate of 1.5 μL/min. Repetitive mass spectra were scanned using the following electrospray ion source parameters: capillary temperature, 220 °C; spray voltage, 1.9 kV. Peptide ions were analyzed by the data-dependent method as follows: (1) full-scan MS in the m/z range 350–2000; (2) zoom scan of the three most intense ions (isolation width: 2 Da); and (3) MS/MS analysis of the three most intense ions (Q 0.250, collision energy 29 a.u.). Mass calibration was conducted using a standard mixture of caffeine (Mr 194.1 Da), MRFA peptide (Mr 523.6 Da), and Ultramark (Mr 1621 Da). Data acquisition was performed using Excalibur v. 1.4 software (Thermo Fisher Scientific).
3.12. Database Search and Protein Identification
MS/MS data were used to perform protein identifications by searching in the UniProt “Viridiplantae” database compiled by SwissProt with 25,438 sequences and TrEMBL with 5,724,529 sequences (January 2015 release). The identifications were achieved using the MOWSE algorithm as implemented in the Mascot search engine version 2.5 (Matrix Science:
www.matrixscience.com, accessed on 26 January 2015 ). The following parameters were used for database searches: cleavage specificity—trypsin with two missed cleavages allowed; mass tolerance of 1.2 Da for the precursor ions and a tolerance of 0.6 Da for the fragment ions; allowed modifications—Cys carboxyamidomethylation (fixed), oxidation of Met (variable), transformation of N-terminal Gln, and N-terminal Glu residue in the pyroglutamic acid form (variable). In MASCOT searches, protein scores were derived from ion scores as a non-probabilistic basis for ranking protein hits. Only proteins that met the following criteria were accepted as unambiguously identified: (1) at least two identified peptides fully tryptic; (2) MASCOT score > 57 (probability-based MOWSE score: −10*log(P), where P is the probability that an observed match is a random event.; score > 57 indicates identity or extensive homology (
p ≤ 0.05)). All identifications obtained through MASCOT software were subjected to a manual interpretation of the MS/MS spectra assisted by PepNovo software (
http://proteomics.ucsd.edu/Software/PepNovo.html, accessed on 26 January 2015 ) to confirm or disregard peptide sequence assignation and therefore protein identification. Multiple-sequence alignment was performed with ClustalOmega (
http://www.ebi.ac.uk/Tools/msa/clustalo/, accessed on 26 January 2015 ).
4. Conclusions
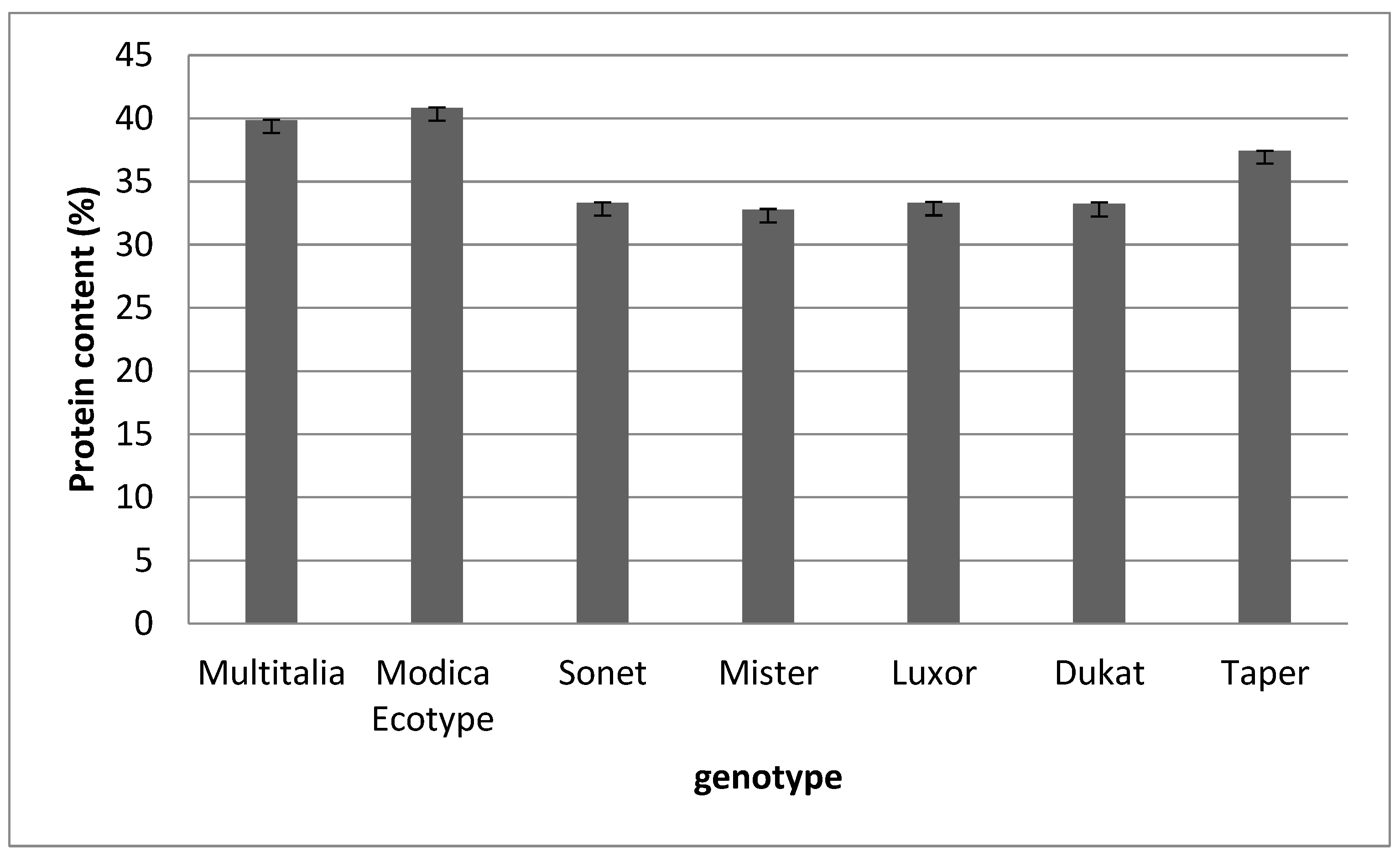
The results for the multielemental composition showed wide variability between genotypes. The amounts of mineral elements presented in this study indicate an excellent nutritional profile, suggesting the suitability of lupin as a raw material for consumption. The protein content results were similar for the different species and genotypes.
The employment of innovative green technologies that do not involve the use of organic solvents or other potentially toxic chemicals to defat lupin seed flour is a matter of growing interest, since consumers are increasingly attracted to natural proteins isolated from protein-rich crops. Moreover, lupin seed oil is a product of much relevance due to its fatty acid and tocopherol profiles, which encourage its use as a valuable ingredient in the food and feed industries. The present work, for the first time, reported useful data on the effects of the SC-CO2 defatting process on lupin oil yield and quality. The results showed that, on average, the SC-CO2 defatting process prevented the total oil recovery from dipping below 60%, with significantly higher percentages for the two white lupin genotypes, namely Luxor and the Modica ecotype, in comparison to Multitalia and the other yellow and narrow-leafed genotypes. Based on the obtained results, the SC-CO2 technology can be proposed as a useful and chemical-free process for defatting lupin seed flour, allowing the acquisition of solvent-free lupin protein isolates and lupin oils to be conveniently used in the food and feed industries. The oil yield differed according to the species and genotype.
The total saturated fatty acid quantities were very similar between the samples. The most abundant saturated fatty acids were palmitic, stearic, and beric. The predominant monounsaturated fatty acid was oleic acid. Among the main polyunsaturated fatty acids, the most abundant were linoleic and alpha linoleic acid.
The total tocopherol quantities averaged about 1500.00 µg/g FW. This total amount of tocopherols was not sufficient to define their quality; nevertheless, the benefits of alpha-tocopherols should be considered, as they provide greater amounts of vitamin E, as well as gamma-tocopherols, which have high antioxidant activity in food. It can be concluded that lupin, in addition to being a good supply of mineral elements, also contributes vitamin E and powerful antioxidant activity, thanks to the very high content of gamma-tocopherols.
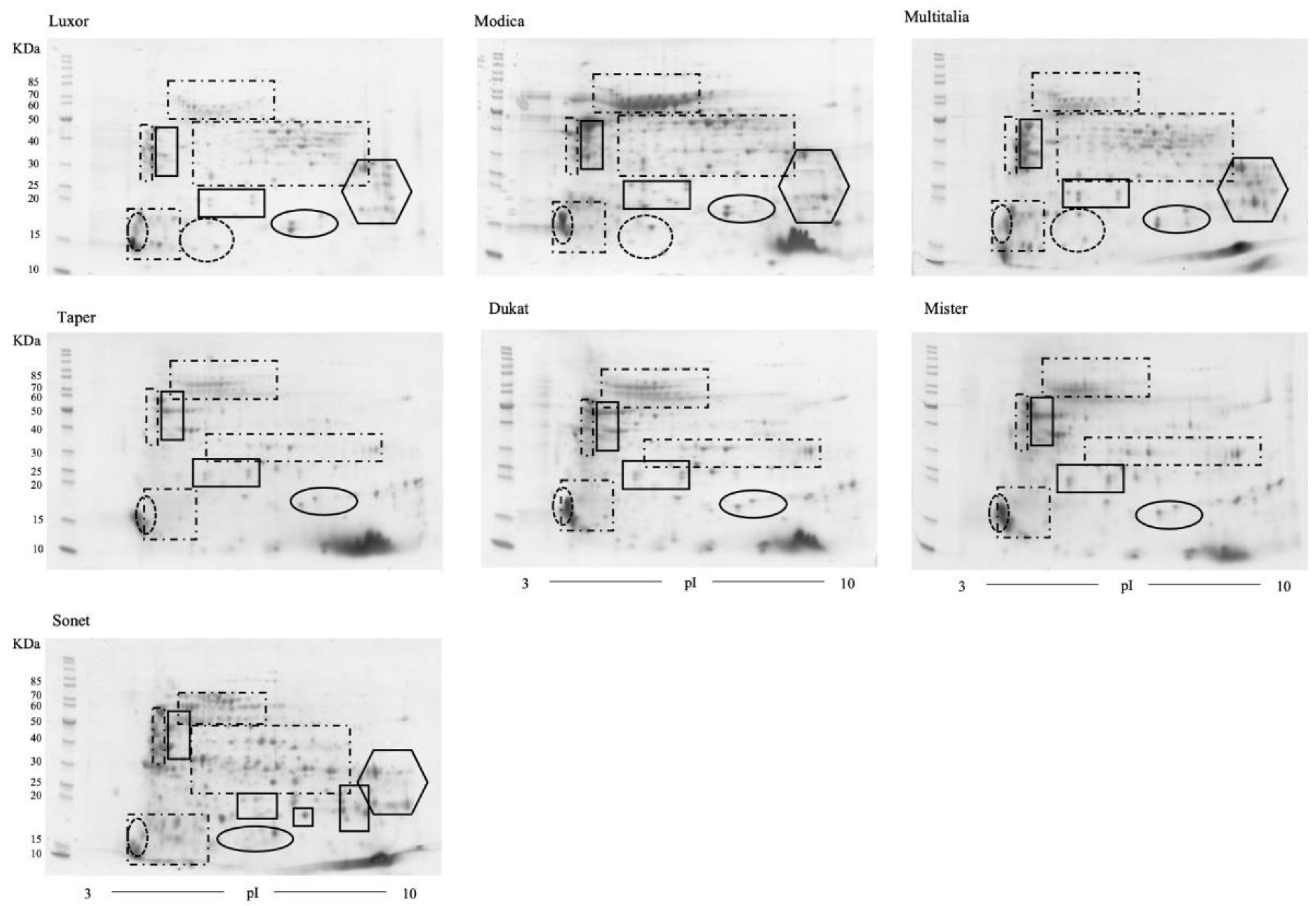
A detailed proteomic analysis of Lupinus albus was performed. The 2D gel analysis showed substantial equivalence between the cultivars of the same species (Lupinus albus and L. luteus). Moreover, the L. angustifolius proteome map showed the presence of additional spots in comparison to L. albus, corresponding to α-conglutins.



 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}