


Phytochemical Characterization of Chamomile (Matricaria recutita L.) Roots and Evaluation of Their Antioxidant and Antibacterial Potential

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results and Discussion
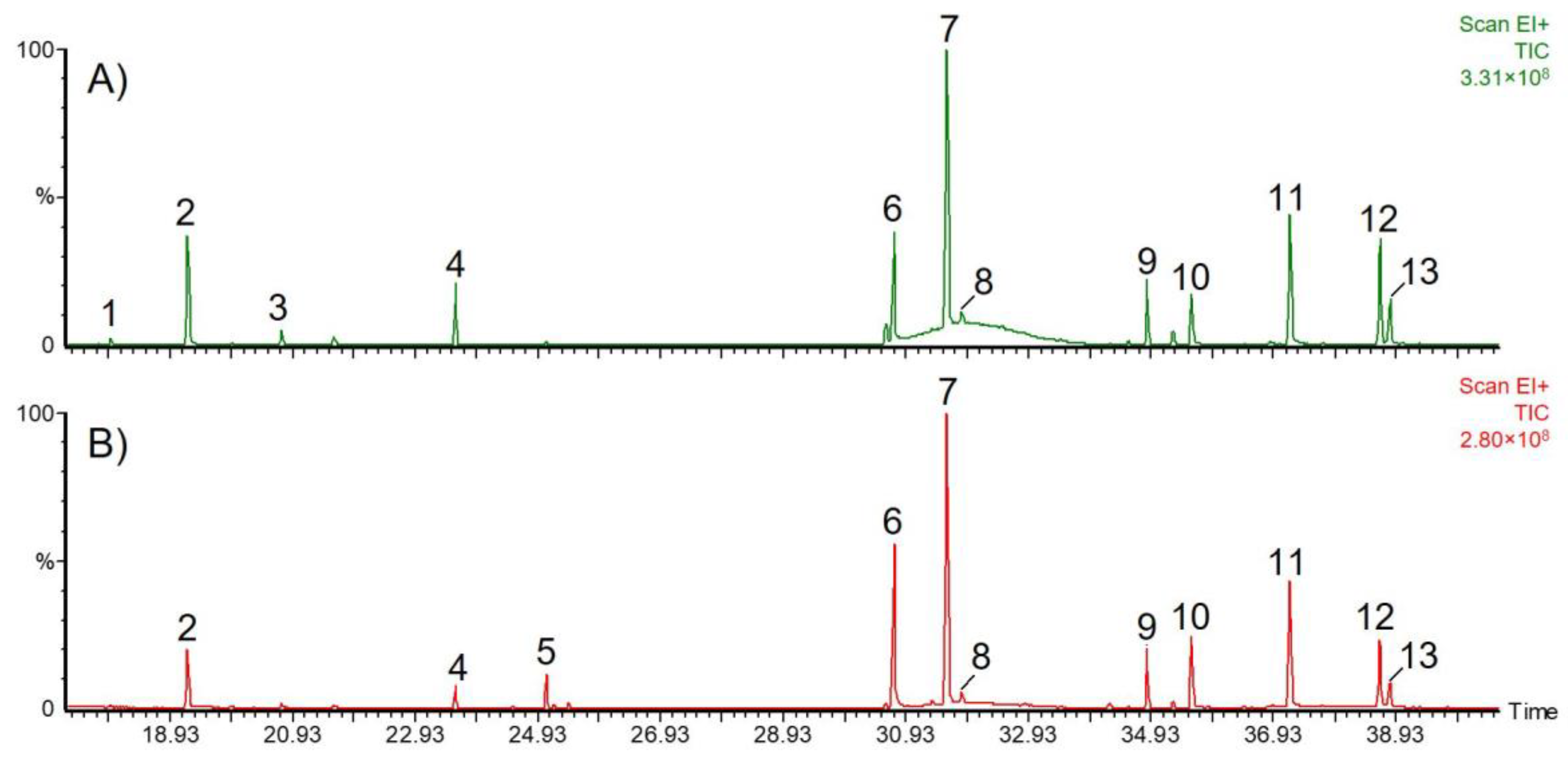
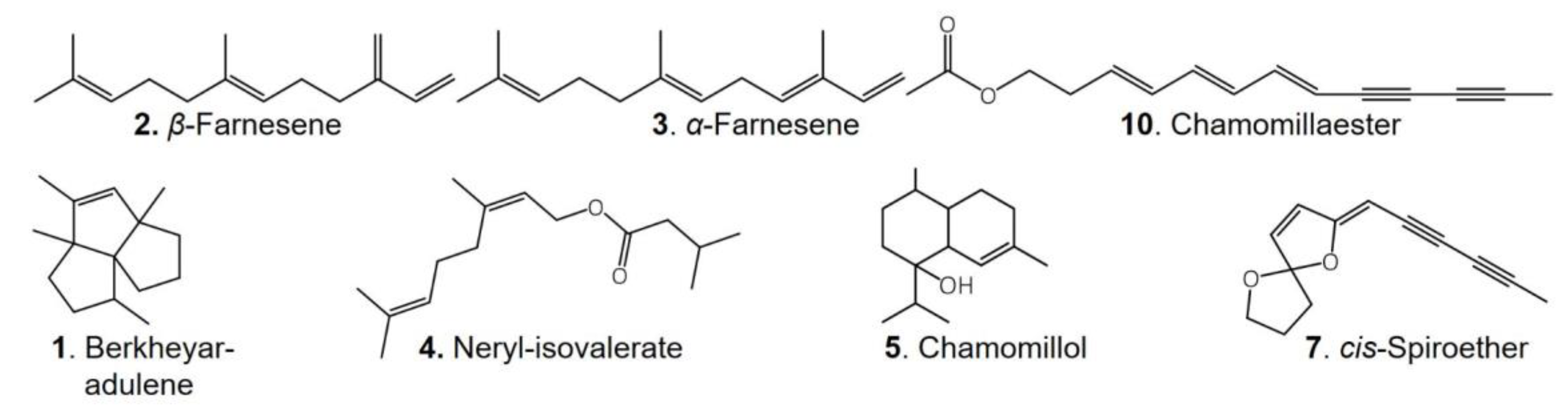
2.1. Secondary Metabolites in M. recutita Roots at Different Developmental Stages
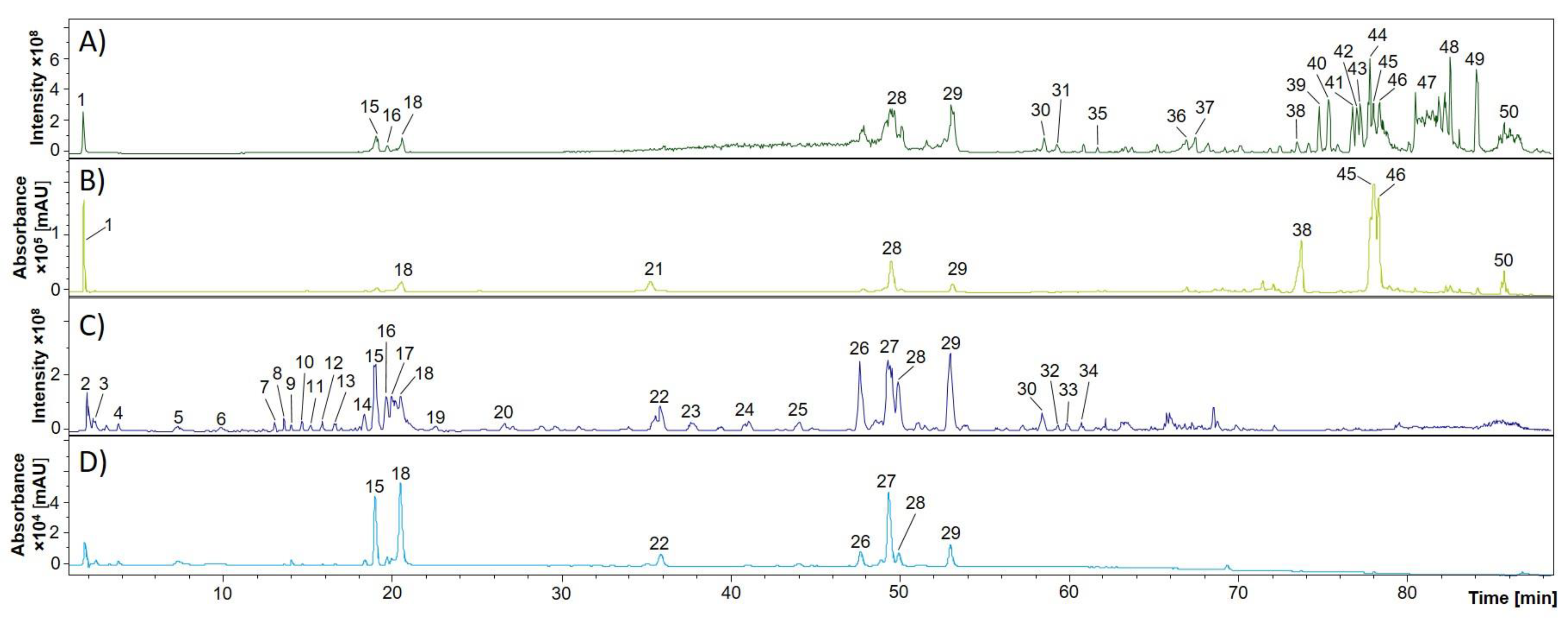
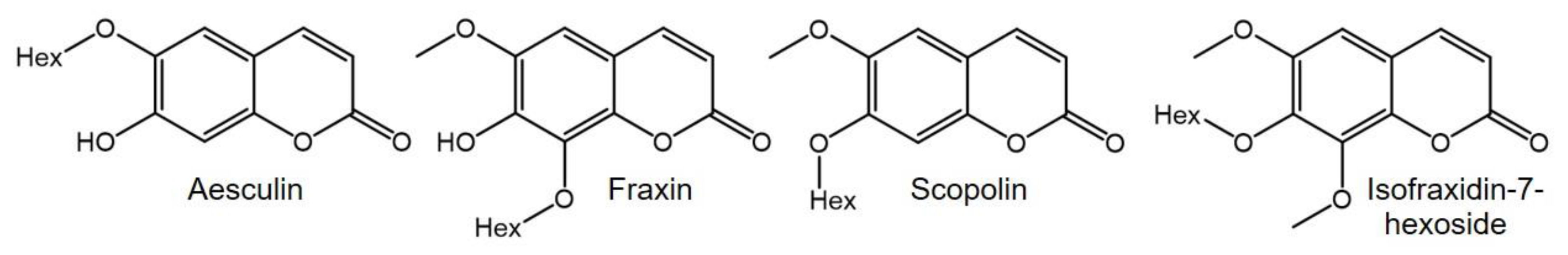
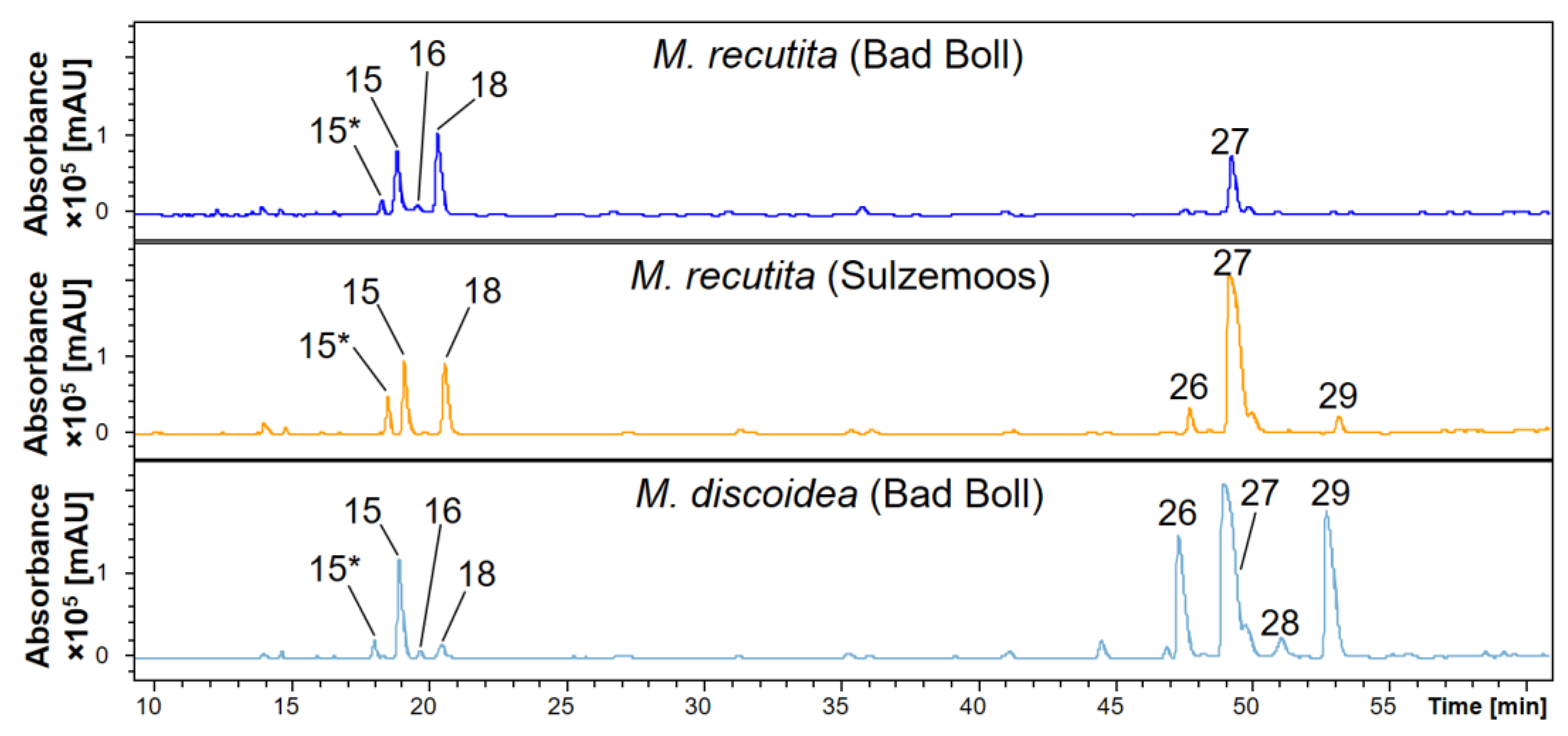
2.2. Phytochemical Comparison of Different Chamomile Varieties
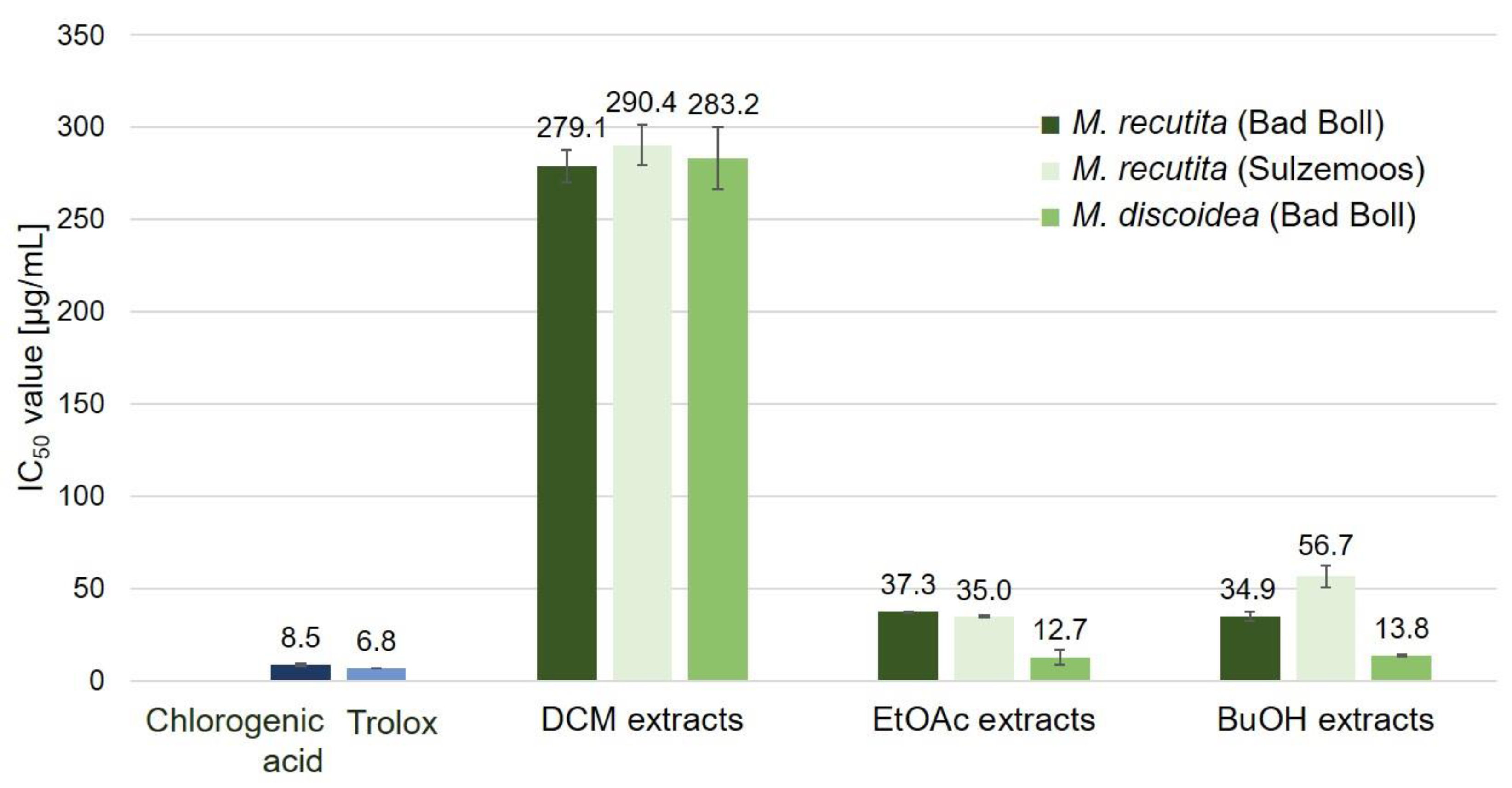
2.3. Antioxidant Potential of Chamomile Root Extracts
2.3.1. DPPH Assay
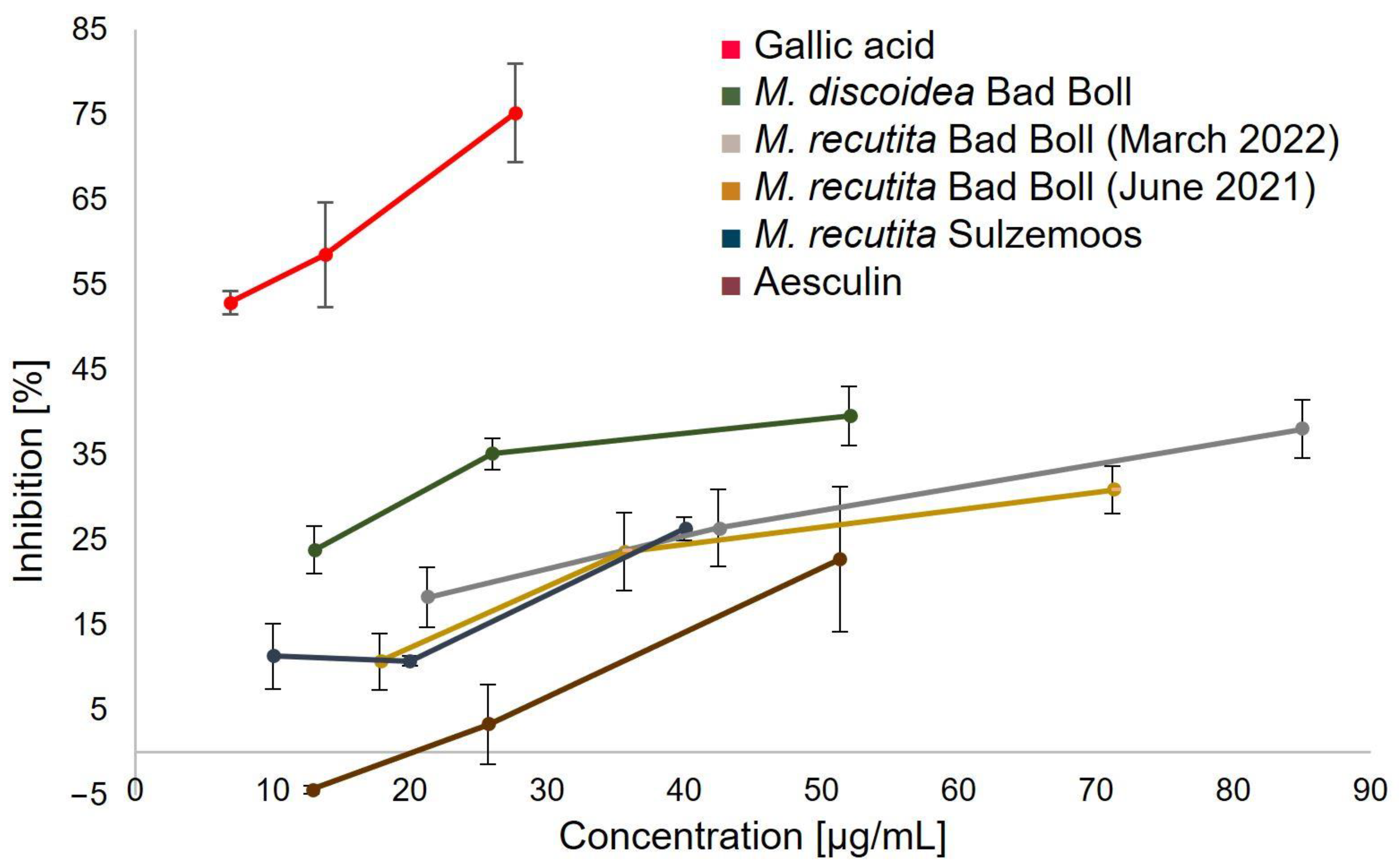
2.3.2. Superoxide Assay
2.4. Antibacterial Potential of Chamomile Roots
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material and Extraction
3.3. GC-MS Analysis of Volatile Constituents
3.4. RP-HPLC-DAD-ESI-MSn Analysis
3.5. 2,2- Diphenyl-1-picrylhydrazyl (DPPH) Assay
3.6. Superoxide Assay
3.7. Antimicrobial Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Singh, O.; Khanam, Z.; Misra, N.; Srivastava, M.K. Chamomile (Matricaria chamomilla L.): An overview. Pharmacogn. Rev. 2011, 5, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Hannig, H.-J. Kamille (Matricaria recutita L.). Z. Arznei Gewürzpfla. 2020, 25, 171–174. [Google Scholar]
- Srivastava, J.K.; Shankar, E.; Gupta, S. Chamomile: A herbal medicine of the past with bright future. Mol. Med. Rep. 2010, 3, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Höferl, M.; Wanner, J.; Tabanca, N.; Ali, A.; Gochev, V.; Schmidt, E.; Kaul, V.K.; Singh, V.; Jirovetz, L. Biological activity of Matricaria chamomilla essential oils of various chemotypes. Planta Med. Int. 2020, 7, e114–e121. [Google Scholar] [CrossRef]
- El Joumaa, M.M.; Borjac, J.M. Matricaria chamomilla: A valuable insight into recent advances in medicinal uses and pharmacological activities. Phytochem. Rev. 2022, 21, 1913–1940. [Google Scholar] [CrossRef]
- Sotiropoulou, N.S.; Megremi, S.F.; Tarantilis, P. Evaluation of antioxidant activity, toxicity, and phenolic profile of aqueous extracts of chamomile (Matricaria chamomilla L.) and Sage (Salvia officinalis L.) prepared at different temperatures. Appl. Sci. 2020, 10, 2270. [Google Scholar] [CrossRef]
- European Medicines Agency (EMA). European Union herbal monograph on Matricaria recutita L., flos. In Committee on Herbal Medicinal Products (HMPC); EMA/HMPC/55843/2011; European Medicines Agency (EMA): Amsterdam, The Netherlands, 2015. [Google Scholar]
- McKay, D.L.; Blumberg, J.B. A review of the bioactivity and potential health benefits of chamomile tea (Matricaria recutita L.). Phytother. Res. 2006, 20, 519–530. [Google Scholar] [CrossRef]
- Schilcher, H. Neuere Erkenntnisse bei der Qualitätsbeurteilung von Kamillenblüten bzw. Kamillenöl. 2. Qualitative Beurteilung des ätherischen Öles in flores chamomillae. Aufteilung der Handelskamillen in vier bzw. fünf chemische Typen. Planta Med. 1973, 23, 132–144. [Google Scholar] [CrossRef]
- Raal, A.; Püssa, T.; Sepp, J.; Malmiste, B.; Arak, E. Content of phenolic compounds in aerial parts of Chamomilla suaveolens from Estonia. Nat. Prod. Comm. 2011, 6, 1934578X1100600. [Google Scholar] [CrossRef]
- Orav, A.; Sepp, J.; Kailas, T.; Müürisepp, M.; Arak, E.; Raal, A. Composition of essential oil of aerial parts of Chamomilla suaveolens from Estonia. Nat. Prod. Comm. 2010, 5, 133–136. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Ali, A.; Jones, A.M.P. Isolation and identification of mosquito biting deterrents from the North American mosquito repelling folk remedy plant, Matricaria discoidea DC. PLoS ONE 2018, 13, e0206594. [Google Scholar] [CrossRef]
- Yousefbeyk, F.; Hemmati, G.; Gholipour, Z.; Ghasemi, S.; Evazalipour, M.; Schubert, C.; Koohi, D.E.; Böhm, V. Phytochemical analysis, antioxidant, cytotoxic, and antimicrobial activities of golden chamomile (Matricaria aurea (Loefl.) Schultz Bip). Z. Naturforsch. C J. Biosci. 2022, 77, 331–342. [Google Scholar] [CrossRef]
- Ignatiadou, M.-E.; Kostaki, M.; Kabouche, Z.; Chatzopoulou, P.; Rallis, M.C.; Karioti, A. HPLC–NMR-based chemical profiling of Matricaria pubescens (Desf.) Schultz and Matricaria recutita and their protective effects on UVA-exposed fibroblasts. Sci 2022, 4, 14. [Google Scholar] [CrossRef]
- Dioscorides Pedanius, T.; Osbaldeston, T.A.; Wood, R.P.A. De Materia Medica: Being an Herbal with Many Other Medicinal Materials Written in Greek in the First Century of the Common Era; Ibidis: Johannesburg, South Africa, 2000; ISBN 9780620234351. [Google Scholar]
- Reichling, J.; Becker, H. Ätherisches Öl in Radix Chamomillae (Matricaria chamomilla L.)/Essential oil of radix chamomillae. Z. Naturforsch. C J. Biosci. 1978, 33, 589–591. [Google Scholar] [CrossRef]
- Sommer, M. Heilpflanzen: Ihr Wesen, Ihre Wirkung, Ihre Anwendung (aethera), 1. Auflage; Urachhaus: Stuttgart, Germany, 2013; ISBN 978-3-8251-8001-0. [Google Scholar]
- Reichling, J.; Bisson, W.; Becker, H.; Schilling, G. Zusammensetzung und Akkumulation des ätherischen Öls in Matricariae Radix (2. Mitteilung)/Composition and Accumulation of Essential Oil in Matricariae Radix (2. Communication). Z. Naturforsch. C 1983, 38, 159–164. [Google Scholar] [CrossRef]
- Das, M.; Ram, G.; Singh, A.; Mallavarapu, G.R.; Ramesh, S.; Ram, M.; Kumar, S. Volatile constituents of different plant parts of Chamomilla recutita L. Rausch grown in the Indo-Gangetic plains. Flavour Fragr. J. 2002, 17, 9–12. [Google Scholar] [CrossRef]
- Szoke, E.; Máday, E.; Tyihák, E.; Kuzovkina, I.N.; Lemberkovics, E. New terpenoids in cultivated and wild chamomile (in vivo and in vitro). J. Chromatogr. B 2004, 800, 231–238. [Google Scholar] [CrossRef]
- Kováčik, J.; Klejdus, B. Dynamics of phenolic acids and lignin accumulation in metal-treated Matricaria chamomilla roots. Plant Cell Rep. 2008, 27, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Bačkor, M. Changes of phenolic metabolism and oxidative status in nitrogen-deficient Matricaria chamomilla plants. Plant Soil 2007, 297, 255–265. [Google Scholar] [CrossRef]
- Kováčik, J.; Bačkor, M. Oxidative status of Matricaria chamomilla plants related to cadmium and copper uptake. Ecotoxicology 2008, 17, 471–479. [Google Scholar] [CrossRef]
- Kováčik, J.; Grúz, J.; Bačkor, M.; Tomko, J.; Strnad, M.; Repčák, M. Phenolic compounds composition and physiological attributes of Matricaria chamomilla grown in copper excess. Environ. Exp. Bot. 2008, 62, 145–152. [Google Scholar] [CrossRef]
- Kováčik, J.; Tomko, J.; Bačkor, M.; Repčák, M. Matricaria chamomilla is not a hyperaccumulator, but tolerant to cadmium stress. Plant Growth Regul. 2006, 50, 239–247. [Google Scholar] [CrossRef]
- Faehnrich, B.; Franz, C.; Nemaz, P.; Kaul, H.-P. Medicinal plants and their secondary metabolites—State of the art and trends in breeding, analytics and use in feed supplementation—With special focus on German chamomile. J. Appl. Bot. Food Qual. 2021, 94, 61–74. [Google Scholar] [CrossRef]
- Máday, E.; Szöke, E.; Muskáth, Z.; Lemberkovics, E. A study of the production of essential oils in chamomile hairy root cultures. Eur. J. Drug Metab. Pharmacokinet. 1999, 24, 303–308. [Google Scholar] [CrossRef]
- Bohlmann, F.; Herbst, P.; Arndt, C.; Schönowsky, H.; Gleinig, H. Polyacetylenverbindungen, XXXIV. Über einen neuen Typ von Polyacetylenverbindungen aus verschiedenen Vertretern des Tribus Anthemideae L. Chem. Ber. 1961, 94, 3193–3216. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Extensive characterisation of bioactive phenolic constituents from globe artichoke (Cynara scolymus L.) by HPLC-DAD-ESI-QTOF-MS. Food Chem. 2013, 141, 2269–2277. [Google Scholar] [CrossRef]
- Jin, J.; Lao, J.; Zhou, R.; He, W.; Qin, Y.; Zhong, C.; Xie, J.; Liu, H.; Wan, D.; Zhang, S.; et al. Simultaneous identification and dynamic analysis of saccharides during steam processing of rhizomes of Polygonatum cyrtonema by HPLC-QTOF-MS/MS. Molecules 2018, 23, 2855. [Google Scholar] [CrossRef] [PubMed]
- Claereboudt, J.; Esmans, E.L.; Claeys, M. Mass spectral behaviour of (M-H)—Ions of some pyrimidine nucleosides. Biol. Mass Spectrom. 1993, 22, 419–421. [Google Scholar] [CrossRef]
- Rodrigues da Silva, M.; Sanchez Bragagnolo, F.; Lajarim Carneiro, R.; de Oliveira Carvalho Pereira, I.; Aquino Ribeiro, J.A.; Martins Rodrigues, C.; Jelley, R.E.; Fedrizzi, B.; Soleo Funari, C. Metabolite characterization of fifteen by-products of the coffee production chain: From farm to factory. Food Chem. 2022, 369, 130753. [Google Scholar] [CrossRef] [PubMed]
- Bunse, M.; Lorenz, P.; Stintzing, F.C.; Kammerer, D.R. Characterization of secondary metabolites in flowers of Sanguisorba officinalis L. by HPLC-DAD-MSn and GC/MS. Chem. Biodivers. 2020, 17, e1900724. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, Q.; Ma, J.; Wu, B.; Zeng, X. Chemical characterization of phenolic compounds in Erigeron injection by rapid-resolution LC coupled with multi-stage and quadrupole-TOF-MS. Chroma 2010, 72, 651–658. [Google Scholar] [CrossRef]
- Jiménez-Sánchez, C.; Lozano-Sánchez, J.; Rodríguez-Pérez, C.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Comprehensive, untargeted, and qualitative RP-HPLC-ESI-QTOF/MS2 metabolite profiling of green asparagus (Asparagus officinalis). J. Food Comp. Anal. 2016, 46, 78–87. [Google Scholar] [CrossRef]
- Rolnik, A.; Soluch, A.; Kowalska, I.; Olas, B. Antioxidant and hemostatic properties of preparations from Asteraceae family and their chemical composition—Comparative studies. Biomed. Pharmacother. 2021, 142, 111982. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liu, S.; Song, W.; Wang, Y.; Li, Y.; Qiao, X.; Liang, H.; Ye, M. Chemical analysis of the Tibetan herbal medicine Carduus acanthoides by UPLC/DAD/qTOF-MS and simultaneous determination of nine major compounds. Anal. Methods 2014, 6, 7181. [Google Scholar] [CrossRef]
- Kolodziej, H.; Kayser, O.; Tan, N. Novel coumarin sulfates from Pelargonium sidoides. In Natural Products in the New Millennium: Prospects and Industrial Application; Rauter, A.P., Palma, F.B., Justino, J., Araújo, M.E., Santos, S.P., Eds.; Imprint; Springer: Dordrecht, The Netherlands, 2002; pp. 59–64. ISBN 9789401598767. [Google Scholar]
- Jaiswal, R.; Müller, H.; Müller, A.; Karar, M.G.E.; Kuhnert, N. Identification and characterization of chlorogenic acids, chlorogenic acid glycosides and flavonoids from Lonicera henryi L. (Caprifoliaceae) leaves by LC-MSn. Phytochemistry 2014, 108, 252–263. [Google Scholar] [CrossRef]
- Cunha, A.G.; Brito, E.S.; Moura, C.F.H.; Ribeiro, P.R.V.; Miranda, M.R.A. UPLC-qTOF-MS/MS-based phenolic profile and their biosynthetic enzyme activity used to discriminate between cashew apple (Anacardium occidentale L.) maturation stages. J. Chromatogr. B 2017, 1051, 24–32. [Google Scholar] [CrossRef]
- Sisó-Terraza, P.; Luis-Villarroya, A.; Fourcroy, P.; Briat, J.-F.; Abadía, A.; Gaymard, F.; Abadía, J.; Álvarez-Fernández, A. Accumulation and secretion of coumarinolignans and other coumarins in Arabidopsis thaliana roots in response to iron deficiency at high pH. Front. Plant Sci. 2016, 7, 1711. [Google Scholar] [CrossRef]
- Fiorentino, M.; Gravina, C.; Piccolella, S.; Pecoraro, M.T.; Formato, M.; Stinca, A.; Pacifico, S.; Esposito, A. Calendula arvensis (Vaill.) L.: A systematic plant analysis of the polar extracts from its organs by UHPLC-HRMS. Foods 2022, 11, 247. [Google Scholar] [CrossRef]
- Clifford, M.N.; Knight, S.; Kuhnert, N. Discriminating between the six isomers of dicaffeoylquinic acid by LC-MS(n). J. Agric. Food Chem. 2005, 53, 3821–3832. [Google Scholar] [CrossRef]
- Guimarães, R.; Barros, L.; Dueñas, M.; Calhelha, R.C.; Carvalho, A.M.; Santos-Buelga, C.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Infusion and decoction of wild German chamomile: Bioactivity and characterization of organic acids and phenolic compounds. Food Chem. 2013, 136, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Tsivelika, N.; Irakli, M.; Mavromatis, A.; Chatzopoulou, P.; Karioti, A. Phenolic profile by HPLC-PDA-MS of Greek chamomile populations and commercial varieties and their antioxidant activity. Foods 2021, 10, 2345. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Xiao, H.; Liu, J.; Liang, X. Identification of phenolic constituents in Radix Salvia miltiorrhizae by liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2006, 20, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Kiprotich, J.; Kuhnert, N. Determination of the hydroxycinnamate profile of 12 members of the Asteraceae family. Phytochemistry 2011, 72, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Marks, S.; Knight, S.; Kuhnert, N. Characterization by LC-MS(n) of four new classes of p-coumaric acid-containing diacyl chlorogenic acids in green coffee beans. J. Agric. Food Chem. 2006, 54, 4095–4101. [Google Scholar] [CrossRef]
- Schütz, K.; Kammerer, D.R.; Carle, R.; Schieber, A. Characterization of phenolic acids and flavonoids in dandelion (Taraxacum officinale WEB. ex WIGG.) root and herb by high-performance liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2005, 19, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.; Sovdat, T.; Vivan, F.; Kuhnert, N. Profiling and characterization by LC-MSn of the chlorogenic acids and hydroxycinnamoylshikimate esters in maté (Ilex paraguariensis). J. Agric. Food Chem. 2010, 58, 5471–5484. [Google Scholar] [CrossRef]
- Herrero, M.; Vicente, M.J.; Cifuentes, A.; Ibáñez, E. Characterization by high-performance liquid chromatography/electrospray ionization quadrupole time-of-flight mass spectrometry of the lipid fraction of Spirulina platensis pressurized ethanol extract. Rapid Commun. Mass Spectrom. 2007, 21, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lee, C.; Kim, Y.H.; Ma, J.Y.; Shim, S.H. Chemical constituents of the aerial part of Taraxacum mongolicum and their chemotaxonomic significance. Nat. Prod. Res. 2017, 31, 2303–2307. [Google Scholar] [CrossRef]
- Della Corte, A.; Chitarrini, G.; Di Gangi, I.M.; Masuero, D.; Soini, E.; Mattivi, F.; Vrhovsek, U. A rapid LC-MS/MS method for quantitative profiling of fatty acids, sterols, glycerolipids, glycerophospholipids and sphingolipids in grapes. Talanta 2015, 140, 52–61. [Google Scholar] [CrossRef]
- Kolodziej, H. Fascinating metabolic pools of Pelargonium sidoides and Pelargonium reniforme, traditional and phytomedicinal sources of the herbal medicine Umckaloabo. Phytomedicine 2007, 14 (Suppl. S6), 9–17. [Google Scholar] [CrossRef] [PubMed]
- Price, R.J.; Renwick, A.B.; Beamand, J.A.; Esclangon, F.; Wield, P.T.; Walters, D.G.; Lake, B.G. Comparison of the metabolism of 7-ethoxycoumarin and coumarin in precision-cut rat liver and lung slices. Food Chem. Toxicol. 1995, 33, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Kotov, A.G.; Khvorost, P.P.; Komissarenko, N.F. Coumarins of Matricaria recutita. Chem. Nat. Compd. 1991, 27, 753. [Google Scholar] [CrossRef]
- Petruľová-Poracká, V.; Repčák, M.; Vilková, M.; Imrich, J. Coumarins of Matricaria chamomilla L.: Aglycones and glycosides. Food Chem. 2013, 141, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; de Jonge, R.; Pieterse, C.M.J. The Age of Coumarins in Plant-Microbe Interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, P.; Ma, Y.; Wang, K.; Chang, X.; Bai, Y.; Zhang, D.; Yang, L. Simultaneous quantitative determination of six caffeoylquinic acids in Matricaria chamomilla L. with high-performance liquid chromatography. J. Chem. 2019, 2019, 1–7. [Google Scholar] [CrossRef]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Roby, M.H.H.; Sarhan, M.A.; Selim, K.A.-H.; Khalel, K.I. Antioxidant and antimicrobial activities of essential oil and extracts of fennel (Foeniculum vulgare L.) and chamomile (Matricaria chamomilla L.). Ind. Crops Prod. 2013, 44, 437–445. [Google Scholar] [CrossRef]
- Móricz, Á.M.; Ott, P.G.; Morlock, G.E. Discovered acetylcholinesterase inhibition and antibacterial activity of polyacetylenes in tansy root extract via effect-directed chromatographic fingerprints. J. Chromatogr. A 2018, 1543, 73–80. [Google Scholar] [CrossRef]
- Abdoul-Latif, F.; Mohamed, N.; Edou, P.; Ali, A.; Djama, S.; Obame, L.-C.; Bassolé, I.; Dicko, M. Antimicrobial and antioxidant activities of essential oil and methanol extract of Matricaria chamomilla L. from Djibouti. J. Med. Plants Res. 2011, 9, 1512–1517. [Google Scholar]
- Al-Dabbagh, B.; Elhaty, I.A.; Elhaw, M.; Murali, C.; Al Mansoori, A.; Awad, B.; Amin, A. Antioxidant and anticancer activities of chamomile (Matricaria recutita L.). BMC Res. Notes 2019, 12, 3. [Google Scholar] [CrossRef]
- Cvetanović, A.; Švarc-Gajić, J.; Zeković, Z.; Jerković, J.; Zengin, G.; Gašić, U.; Tešić, Ž.; Mašković, P.; Soares, C.; Fatima Barroso, M.; et al. The influence of the extraction temperature on polyphenolic profiles and bioactivity of chamomile (Matricaria chamomilla L.) subcritical water extracts. Food Chem. 2019, 271, 328–337. [Google Scholar] [CrossRef]
- Li, X.; Li, K.; Xie, H.; Xie, Y.; Li, Y.; Zhao, X.; Jiang, X.; Chen, D. Antioxidant and cytoprotective effects of the Di-O-caffeoylquinic acid family: The mechanism, structure–activity relationship, and conformational effect. Molecules 2018, 23, 222. [Google Scholar] [CrossRef]
- Indo, H.P.; Yen, H.-C.; Nakanishi, I.; Matsumoto, K.-I.; Tamura, M.; Nagano, Y.; Matsui, H.; Gusev, O.; Cornette, R.; Okuda, T.; et al. A mitochondrial superoxide theory for oxidative stress diseases and aging. J. Clin. Biochem. Nutr. 2015, 56, 1–7. [Google Scholar] [CrossRef]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Furuno, K.; Akasako, T.; Sugihara, N. The contribution of the pyrogallol moiety to the superoxide radical scavenging activity of flavonoids. Biol. Pharm. Bull. 2002, 25, 19–23. [Google Scholar] [CrossRef]
- Cvetanović, A.; Švarc-Gajić, J.; Zeković, Z.; Savić, S.; Vulić, J.; Mašković, P.; Ćetković, G. Comparative analysis of antioxidant, antimicrobiological and cytotoxic activities of native and fermented chamomile ligulate flower extracts. Planta 2015, 242, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Sebai, H.; Jabri, M.-A.; Souli, A.; Rtibi, K.; Selmi, S.; Tebourbi, O.; El-Benna, J.; Sakly, M. Antidiarrheal and antioxidant activities of chamomile (Matricaria recutita L.) decoction extract in rats. J. Ethnopharmacol. 2014, 152, 327–332. [Google Scholar] [CrossRef]
- Sebai, H.; Jabri, M.-A.; Souli, A.; Hosni, K.; Rtibi, K.; Tebourbi, O.; El-Benna, J.; Sakly, M. Chemical composition, antioxidant properties and hepatoprotective effects of chamomile (Matricaria recutita L.) decoction extract against alcohol-induced oxidative stress in rat. Gen. Physiol. Biophys. 2015, 34, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Barreiro, M.F.; Morales, P.; Ferreira, I.C.F.R. Adding molecules to food, pros and cons: A review on synthetic and natural food additives. Compr. Rev. Food Sci. Food Saf. 2014, 13, 377–399. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Del-Río, I.; López-Ibáñez, S.; Magadán-Corpas, P.; Fernández-Calleja, L.; Pérez-Valero, Á.; Tuñón-Granda, M.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Terpenoids and polyphenols as natural antioxidant agents in food preservation. Antioxidants 2021, 10, 1264. [Google Scholar] [CrossRef] [PubMed]
- Hoang, H.T.; Moon, J.-Y.; Lee, Y.-C. Natural antioxidants from plant extracts in skincare cosmetics: Recent applications, challenges and perspectives. Cosmetics 2021, 8, 106. [Google Scholar] [CrossRef]
- Caleja, C.; Barros, L.; Antonio, A.L.; Ciric, A.; Barreira, J.C.; Sokovic, M.; Oliveira, M.B.P.; Santos-Buelga, C.; Ferreira, I.C. Development of a functional dairy food: Exploring bioactive and preservation effects of chamomile (Matricaria recutita L.). J. Funct. Foods 2015, 16, 114–124. [Google Scholar] [CrossRef]
- Bunse, M.; Daniels, R.; Gründemann, C.; Heilmann, J.; Kammerer, D.R.; Keusgen, M.; Lindequist, U.; Melzig, M.F.; Morlock, G.E.; Schulz, H.; et al. Essential oils as multicomponent mixtures and their potential for human health and well-being. Front. Pharmacol. 2022, 13, 2645. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-X.; Erhunmwunsee, F.; Liu, M.; Yang, K.; Zheng, W.; Tian, J. Antimicrobial mechanisms of spice essential oils and application in food industry. Food Chem. 2022, 382, 132312. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Miao, M.; Yue, T.; Zhang, Y.; Cui, L.; Wang, Z.; Yuan, Y. Antibacterial activity and mechanism of cinnamic acid and chlorogenic acid against Alicyclobacillus acidoterrestris vegetative cells in apple juice. Int. J. Food Sci. Technol. 2019, 54, 1697–1705. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Fazly Bazzaz, B.S. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef]
- Móricz, A.M.; Szarka, S.; Ott, P.G.; Héthelyi, E.B.; Szoke, E.; Tyihák, E. Separation and identification of antibacterial chamomile components using OPLC, bioautography and GC-MS. Med. Chem. 2012, 8, 85–94. [Google Scholar] [CrossRef]
- Kenny, O.; Brunton, N.P.; Walsh, D.; Hewage, C.M.; McLoughlin, P.; Smyth, T.J. Characterisation of antimicrobial extracts from dandelion root (Taraxacum officinale) using LC-SPE-NMR. Phytother. Res. 2015, 29, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Schoss, K.; Kočevar Glavač, N.; Dolenc Koce, J.; Anžlovar, S. Supercritical CO2 plant extracts show antifungal activities against crop-borne fungi. Molecules 2022, 27, 1132. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Nazaruk, J.; Polito, L.; Morais-Braga, M.F.B.; Rocha, J.E.; Coutinho, H.D.M.; Salehi, B.; Tabanelli, G.; Montanari, C.; Del Mar Contreras, M.; et al. Matricaria genus as a source of antimicrobial agents: From farm to pharmacy and food applications. Microbiol. Res. 2018, 215, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, P.; Heinrich, M.; Conrad, J.; Heller, A.; Stintzing, F.C.; Kammerer, D.R. Comprehensive characterisation of n-alkylresorcinols and other lipid constituents of Mercurialis tomentosa L. from Alicante, Spain. Chem. Biodivers. 2017, 14, 255. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, P.; Zeh, M.; Martens-Lobenhoffer, J.; Schmidt, H.; Wolf, G.; Horn, T.F.W. Natural and newly synthesized hydroxy-1-aryl-isochromans: A class of potential antioxidants and radical scavengers. Free Radic. Res. 2005, 39, 535–545. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | tR (min) | MW (g/mol) | m/z (M+ Int. %) |
|---|---|---|---|---|
| 1 | Berkheyaradulene | 17.9 | 204 | 204 (15%), 189, 162, 147, 134, 119 |
| 2 | β-Farnesene | 19.2 | 204 | 204 (10%), 161, 133, 120, 107, 93, 79, 69, 55 |
| 3 | α-Farnesene | 20.8 | 204 | 204 (1%), 161, 119, 107, 93, 79, 69, 55 |
| 4 | Neryl-isovalerate | 23.6 | 238 | 238 (1%), 136, 121, 107, 93, 85, 69, 57 |
| 5 | Chamomillol | 25.1 | 222 | 222 (10%), 204, 179, 161, 119, 105, 81 |
| 6 | Not identified | 30.7 | 220 | 220 (100%), 190, 178, 136 |
| 7 | cis-Spiroether | 31.6 | 200 | 200 (100%), 170, 157, 128, 115,76 |
| 8 | trans-Spiroether | 31.8 | 200 | 200 (100%), 170, 157, 128, 115,76 |
| 9 | Palmitic acid * | 34.9 | 328 | 328 (20%), 313, 145, 161, 117, 73, 55 |
| 10 | Chamomillaester I | 35.6 | 228 | 228 (20%), 168, 153, 141, 128, 115, 91, 77 |
| 11 | Chamomillaester II | 37.2 | 228 | 228 (25%), 168, 152, 141, 128, 115, 91, 77 |
| 12 | Linoleic acid * | 38.7 | 352 | 352 (10%), 337, 262, 220, 129, 81, 73, 67 |
| 13 | Linolenic acid * | 38.8 | 350 | 350 (10%), 335, 157, 129, 108, 95, 73, 55 |
| EtOAc Extract (A) a | BuOH Extract (C) a | tR (min) | Substance | UV Maxima (nm) b | Mass Spectrometric Data (m/z) c | Reference | ||
|---|---|---|---|---|---|---|---|---|
| MS1 | MS2 | MS3 | ||||||
| 1 | 1.7 | Chlorogenic acid hexoside | 234, 324 | 515 | 353 | 191, 135 | [29] | |
| 2 | 1.9 | Sucrose | - | 683, 533, 439, 404 | 341, 179 | 143 | [30] | |
| 3 | 2.4 | 1-Kestose | - | 637, 549, 503 | 503, 464, 323 | [30] | ||
| 4 | 3.7 | Uridine | 202, 262 | 243 | 200, 152 | 138, 110 | [31] | |
| 5 | 7.3 | trans-Zeatin riboside | 204, 258 | 533, 312 | 266, 134 | 134 | MassBank PR100614 | |
| 6 | 9.9 | Ellagic acid | ND d | 347 | 301 | 223, 161, 139 | [32] | |
| 7 | 12.8 | Galloyl hexoside | ND d | 331 | 169, 161 | 152, 139 | [33] | |
| 8 | 13.5 | Galloyl-3-O-β-D-glucuronide | ND d | 391 | 345, 207, 183 | 331, 183 | [34] | |
| 9 | 14.0 | L-Tryptophan | 220, 278 | 203 | 159, 158 | [35,36], standard | ||
| 10 | 14.6 | 3-O-Caffeoylquinic acid | 324 | 353 | 191, 179, 135 | 85 | [37,38] | |
| 11 | 15.1 | Fraxin sulfate | 206, 230, 288 | 449 | 369, 241 | 207, 192 | [39] | |
| 12 | 15.6 | Aesculin | 290 sh, 342 | 339 | 177 | 133 | [40,41], standard | |
| 13 | 16.4 | Caffeoyl-Fraxetin | 259, 305 | 387, 339 | 207, 179 | 164, 161, 146 | Tentative | |
| 14 | 18.4 | Scopolin | 205, 226, 288 sh, 338 | 443, 419, 399 | 353, 237, 191, 176 | 176 | [42] | |
| 15 | 15 | 18.9 | 5-O-Caffeoylquinic acid | 218, 235 sh, 290 sh, 324 | 707 * | 353 | 191, 173, 135 | [43], standard |
| 16 | 16 | 19.7 | Fraxin | 208, 230, 300 | 369, 221 | 207 | 192 | [42], standard |
| 17 | 19.9 | Fraxetin sulfate | 206, 230, 338 | 287 | 207 | 192 | [39] | |
| 18 | 18 | 20.6 | Isofraxidin-7-hexoside | 208, 228 sh, 294, 334 sh | 429, 383, 287, 221 | 221 | 206, 191 | [42] |
| 19 | 22.6 | 4-O-Caffeoylquinic acid | 324 | 353 | 191 | 173, 93 | [44] | |
| 20 | 26.7 | Fraxetin derivative | ND d | 585 | 377 | 329, 314 | Tentative | |
| 21 | 35.4 | 3,5-Dicaffeoylquinic acid (3,5-diCQA) | 218, 236, 322 | 533, 515 | 353, 335 | 191, 179, 135 | [44] | |
| 22 | 35.9 | Ferulic acid hexoside | 223 sh, 236, 295 sh, 318 | 711 | 355 | 193, 149 | [45,46] | |
| 23 | 37.7 | Acetylquinic acid | ND d | 489, 233 | 171, 143, 127 | [40] | ||
| 24 | 41.1 | Dimethyl lithospermate | 226, 276 | 565, 467 | 339, 327 | 323, 309, 294 | [47] | |
| 25 | 43.9 | Tricaffeoyl-quinic acid | 322 | 677 | 515, 353 | 191, 179, 135 | [48] | |
| 26 | 47.4 | 1,4-diCQA | 218, 242, 300sh, 324 | 515 | 353, 335 | 191, 179, 173, 135 | [44,46] | |
| 27 | 49.1 | 1,3-diCQA | 218, 236 sh, 300 sh, 326 | 515 | 353, 191 | 191, 179, 135 | [44] | |
| 28 | 28 | 49.5 | 1,5-diCQA | 218, 242, 300 sh, 326 | 515 | 353, 335, 191 | 191, 179, 135 | [44] |
| 29 | 29 | 52.8 | 4,5-diCQA | 220, 242, 300 sh, 326 | 515 | 353, 203 | 191, 179, 173, 135 | [44] |
| 30 | 30 | 58.5 | 3,4-diCQA | 280, 322 | 515, 439, 345 | 353, 191, 173 | 191, 179, 173, 135 | [6,44] |
| 31 | 59.2 | Caffeoyl-feruloylquinic acid | 328 | 529, 439 | 367, 349 | 334, 191, 179, 161 | [40,49] | |
| 32 | 59.3 | unknown | ND d | 439 | 393, 379 | 349, 235, 217 | ||
| 33 | 59.9 | Chicoric acid (acetyl derivative) | ND d | 515, 455 | 473, 353, 311, 263, 221, 179 | 203, 179, 161, 143 | [36,50] | |
| 34 | 60.7 | Caffeic acid derivative | ND d | 707, 519 | 477 | 263, 221, 179, 161 | Tentative | |
| 35 | 61.8 | Sinapoyl-feruloyl-caffeoylquinic acid | 242, 328 | 735 | 559 | 517, 337, 235, 193 | [32,51] | |
| 36 | 66.9 | Coumaroyl-feruloylquinic acid | 238, 324 | 707, 427 | 513, 367 | 367, 173 | [48] | |
| 37 | 69.9 | Diferuloylquinic acid | 242, 318 | 707, 645 | 543 | 367 | [48,49] | |
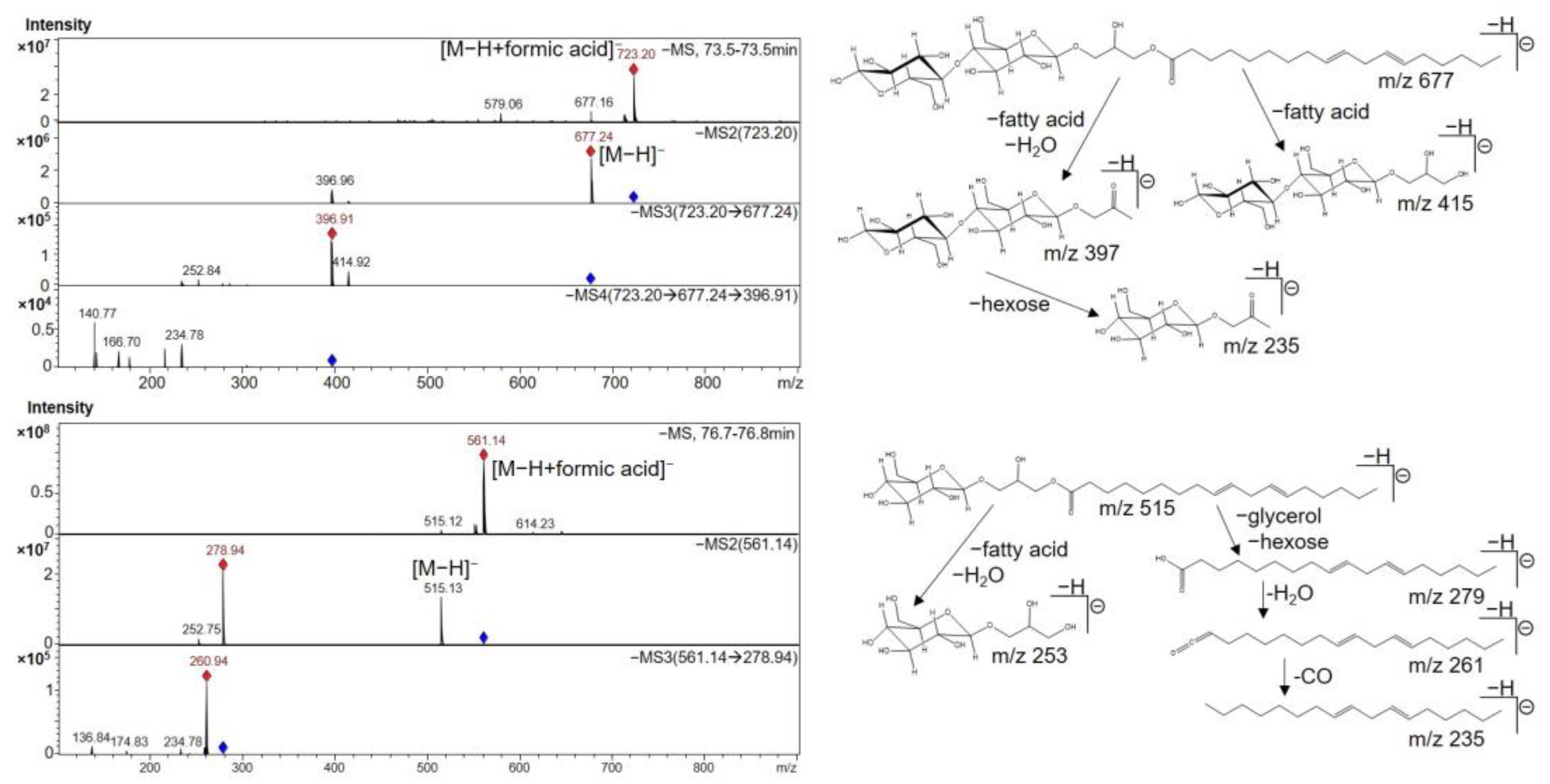
| 38 | 73.5 | Linoleic acid diglycosyl monoglyceride | 228, 238, 316 | 723 | 677, 397 | 415, 397, 235 | [35] | |
| 39, 40 | 74.8 75.4 | Linolenic acid monoglycosyl monoglyceride isomers | 240, 313 | 559 | 513, 277, 253 | 259, 233 | [35,52] | |
| 41 | 76.8 | Linoleic acid monoglycosyl monoglyceride | 238, 250, 314 | 561 | 515, 279 | 261, 205 | [35,53] | |
| 42 | 77.0 | Linoleic acid derivative | 238, 314 | 529 | 511, 279, 249 | 261, 205 | Tentative | |
| 43 | 77.3 | Linoleic acid monoglycosyl monoglyceride | 240, 316 | 561 | 515, 279 | 261, 205 | [35,53] | |
| 44 | 77.8 | Linoleic acid derivative | 242, 254, 324 | 529 | 511, 279, 249 | 261, 205 | Tentative | |
| 45, 46 | 78.0 78.4 | Phosphoglyceride isomers | 242, 250, 324 | 431 | 171, 153 | 97, 79 | [52] | |
| 47 | 82.1 | Phosphoglyceride | 314 | 433, 399 | 171, 153 | 79 | [52] | |
| 48 | 82.5 | Linolenic acid | <200, 242 | 311, 277 | 259, 233, 205 | 191, 179 | [35,54] | |
| 49 | 84.2 | Linoleic acid | <200 | 279 | 261 | 243 | [35,54] | |
| 50 | 85.7 | Dihydroxy-linolenic acid | 226 | 325, 281 | 183 | [35], tentative | ||
| S. aureus | B. subtilis | |||||
|---|---|---|---|---|---|---|
| Extract | 0.8 mg/Disk | 1.6 mg/Disk | 3.2 mg/Disk | 0.8 mg/Disk | 1.6 mg/Disk | 3.2 mg/Disk |
| M. recutita Bad Boll | ||||||
| DCM | 8 ± 0 | 9 ± 0 | 9 ± 2 | 6 ± 0 | 7 ± 0 | 8 ± 0 |
| EtOAc | 8 ± 0 | 7 ± 1 | 9 ± 1 | - | 8 ± 1 | 10 ± 1 |
| BuOH | 7 ± 0 | 8 ± 1 | 10 ± 1 | 7 ± 1 | 8 ± 0 | 9 ± 1 |
| M. recutita Sulzemoos | ||||||
| DCM | 7 ± 1 | 9 ± 1 | 10 ± 1 | 8 ± 0 | 9 ± 0 | 9 ± 1 |
| BuOH | - | 8 ± 1 | 9 ± 1 | - | 7 ± 0 | 8 ± 1 |
| M. discoidea | ||||||
| DCM | 9 ± 1 | 10 ± 1 | 11 ± 1 | 9 ± 2 | 9 ± 2 | 9 ± 1 |
| EtOAc | 7 ± 0 | 9 ± 1 | 10 ± 0 | 8 ± 0 | 9 ± 0 | 10 ± 0 |
| BuOH | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mailänder, L.K.; Lorenz, P.; Bitterling, H.; Stintzing, F.C.; Daniels, R.; Kammerer, D.R. Phytochemical Characterization of Chamomile (Matricaria recutita L.) Roots and Evaluation of Their Antioxidant and Antibacterial Potential. Molecules 2022, 27, 8508. https://doi.org/10.3390/molecules27238508
Mailänder LK, Lorenz P, Bitterling H, Stintzing FC, Daniels R, Kammerer DR. Phytochemical Characterization of Chamomile (Matricaria recutita L.) Roots and Evaluation of Their Antioxidant and Antibacterial Potential. Molecules. 2022; 27(23):8508. https://doi.org/10.3390/molecules27238508
Chicago/Turabian StyleMailänder, Lilo K., Peter Lorenz, Hannes Bitterling, Florian C. Stintzing, Rolf Daniels, and Dietmar R. Kammerer. 2022. "Phytochemical Characterization of Chamomile (Matricaria recutita L.) Roots and Evaluation of Their Antioxidant and Antibacterial Potential" Molecules 27, no. 23: 8508. https://doi.org/10.3390/molecules27238508
APA StyleMailänder, L. K., Lorenz, P., Bitterling, H., Stintzing, F. C., Daniels, R., & Kammerer, D. R. (2022). Phytochemical Characterization of Chamomile (Matricaria recutita L.) Roots and Evaluation of Their Antioxidant and Antibacterial Potential. Molecules, 27(23), 8508. https://doi.org/10.3390/molecules27238508