
In Vitro Determination of the Skin Anti-Aging Potential of Four-Component Plant-Based Ingredient


Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Experimental Product
2.3. Cell Culture
2.4. High-Performance Liquid Chromatography (HPLC) Analysis of EY
2.5. Human Fibroblast Proliferation by MTT Assay in Aging Conditioned Medium
2.6. Measurement of Intracellular ROS Formation in Aging Conditioned Medium
2.7. Telomere Length Measurement in Human Fibroblast: Real-Time PCR
2.8. Telomere Length Measurement in Fibroblast Submitted to Oxidative Stress: TAT® Technology
2.9. Relative Telomerase Activity
2.10. Antiglycation Study in Human Fibroblasts. AGE Quantification-ELISA Assay
2.11. Melanin Quantification in Human Melanocytes Irradiated with UVA
3. Results
3.1. Characterization of EY by High-Performance Liquid Chromatography (HPLC)
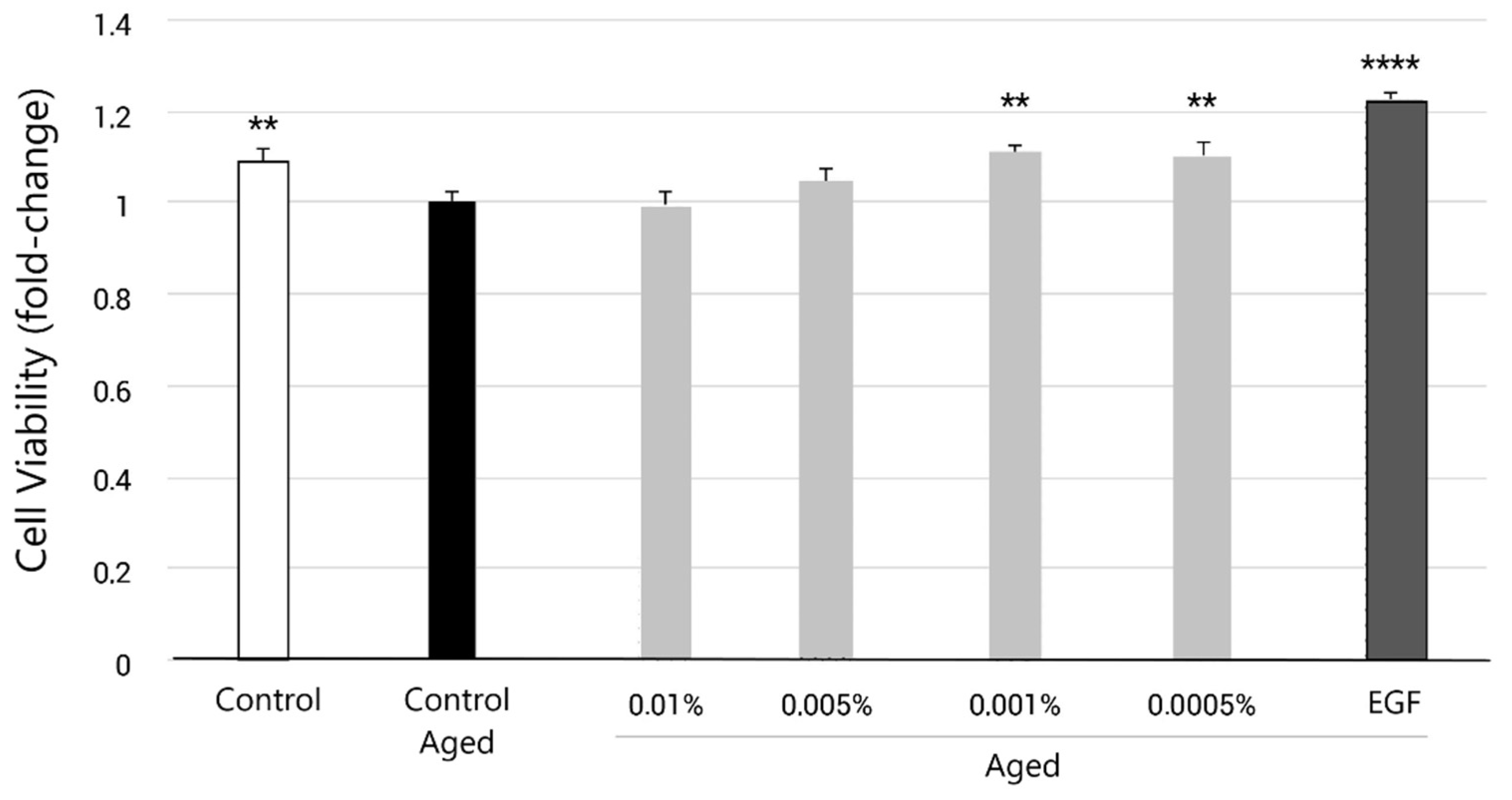
3.2. EY Inhibited H2O2-Induced Proliferation Reduction in Human Dermal Fibroblasts
3.3. EY Inhibited H2O2-ROS Production in Human Skin Fibroblasts
3.4. Effect of EY on Telomere Length in Human Fibroblasts
3.5. Effect of EY on Telomere Length in Human Fibroblasts under Oxidative Stress Conditions
3.6. Effect of EY on Telomerase Activity in Skin Fibroblasts
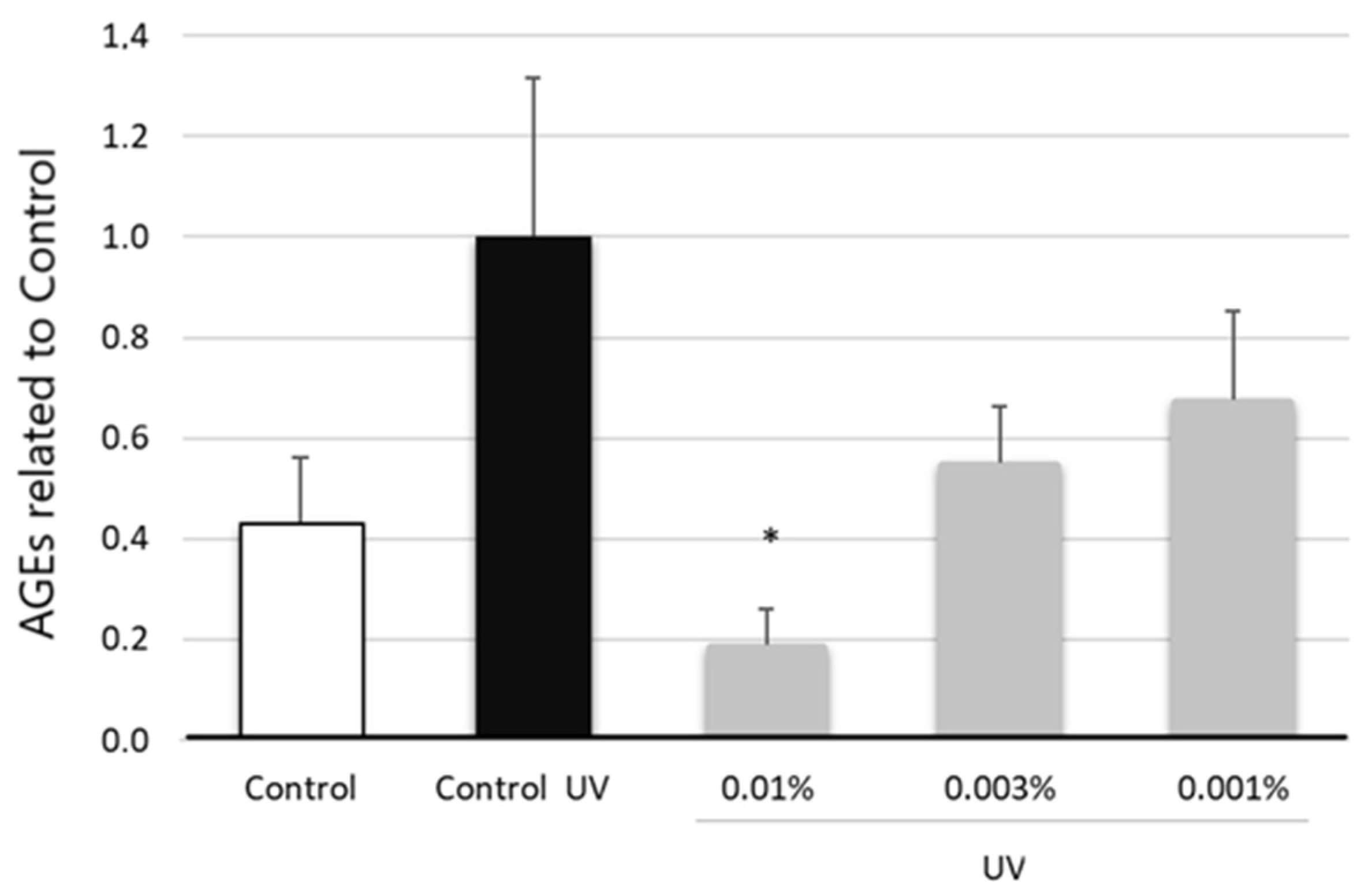
3.7. Antiglycation Assessment of EY
3.8. EY Inhibited Melanin Biosynthesis in Human Melanocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khavkin, J.; Ellis, D.A.F. Aging Skin: Histology, Physiology, and Pathology. Facial Plast. Surg. Clin. N. Am. 2011, 19, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L.; Moorhead, P.S. The Serial Cultivation of Human Diploid Cell Strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Campisi, J. Aging, Cellular Senescence, and Cancer. Annu. Rev. Physiol. 2013, 75, 685–705. [Google Scholar] [CrossRef]
- Franco, A.C.; Aveleira, C.; Cavadas, C. Skin Senescence: Mechanisms and Impact on Whole-Body Aging. Trends Mol. Med. 2022, 28, 97–109. [Google Scholar] [CrossRef]
- Csekes, E.; Račková, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef]
- Cuollo, L.; Antonangeli, F.; Santoni, A.; Soriani, A. The Senescence-Associated Secretory Phenotype (SASP) in the Challenging Future of Cancer Therapy and Age-Related Diseases. Biology 2020, 9, 485. [Google Scholar] [CrossRef]
- Bernadotte, A.; Mikhelson, V.M.; Spivak, I.M. Markers of Cellular Senescence. Telomere Shortening as a Marker of Cellular Senescence. Aging 2016, 8, 3–11. [Google Scholar] [CrossRef]
- Victorelli, S.; Passos, J.F. Telomeres and Cell Senescence—Size Matters Not. EBioMedicine 2017, 21, 14–20. [Google Scholar] [CrossRef]
- Sherratt, D.J.; West, S.C.; Chan, S.R.W.L.; Blackburn, E.H. Telomeres and Telomerase. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 109–122. [Google Scholar] [CrossRef]
- Allsopp, R.C.; Vaziri, H.; Patterson, C.; Goldstein, S.; Younglai, E.V.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere Length Predicts Replicative Capacity of Human Fibroblasts. Proc. Natl. Acad. Sci. USA 1992, 89, 10114–10118. [Google Scholar] [CrossRef] [PubMed]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The Impact of Oxidative DNA Damage and Stress on Telomere Homeostasis. Mech. Ageing Dev. 2019, 177, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, E.M.; Klingelhutz, A.J. The Role of Telomeres in the Ageing of Human Skin. Exp. Dermatol. 2011, 20, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Ohta, T.; Horie, N.; Naru, E.; Hasegawa, M.; Kanda, N. Donor Age Reflects the Replicative Lifespan of Human Fibroblasts in Culture. Hum. Cell 2009, 22, 38–42. [Google Scholar] [CrossRef]
- Brun, C.; Jean-Louis, F.; Oddos, T.; Bagot, M.; Bensussan, A.; Michel, L. Phenotypic and Functional Changes in Dermal Primary Fibroblasts Isolated from Intrinsically Aged Human Skin. Exp. Dermatol. 2016, 25, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Lago, J.C.; Puzzi, M.B. The Effect of Aging in Primary Human Dermal Fibroblasts. PLoS ONE 2019, 14, e0219165. [Google Scholar] [CrossRef] [PubMed]
- Rorteau, J.; Chevalier, F.P.; Bonnet, S.; Barthélemy, T.; Lopez-Gaydon, A.; Martin, L.S.; Bechetoille, N.; Lamartine, J. Maintenance of Chronological Aging Features in Culture of Normal Human Dermal Fibroblasts from Old Donors. Cells 2022, 11, 858. [Google Scholar] [CrossRef]
- Nutrition Business Journal (NBC). Condition Specific Report © 2022; Market Study; Informa PLC: London, UK, 2022. [Google Scholar]
- Hernandez, D.F.; Cervantes, E.L.; Luna-Vital, D.A.; Mojica, L. Food-Derived Bioactive Compounds with Anti-Aging Potential for Nutricosmetic and Cosmeceutical Products. Crit. Rev. Food Sci. Nutr. 2021, 61, 3740–3755. [Google Scholar] [CrossRef]
- Nobile, V.; Schiano, I.; Peral, A.; Giardina, S.; Spartà, E.; Caturla, N. Antioxidant and Reduced Skin-Ageing Effects of a Polyphenol-Enriched Dietary Supplement in Response to Air Pollution: A Randomized, Double-Blind, Placebo-Controlled Study. Food Nutr. Res. 2021, 65. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, A.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Nutraceuticals for Skin Care: A Comprehensive Review of Human Clinical Studies. Nutrients 2018, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- Domaszewska-Szostek, A.; Puzianowska-Kuźnicka, M.; Kuryłowicz, A. Flavonoids in Skin Senescence Prevention and Treatment. Int. J. Mol. Sci. 2021, 22, 6814. [Google Scholar] [CrossRef] [PubMed]
- Jacczak, B.; Rubiś, B.; Totoń, E. Potential of Naturally Derived Compounds in Telomerase and Telomere Modulation in Skin Senescence and Aging. Int. J. Mol. Sci. 2021, 22, 6381. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Bartosz, G. Prevention of Protein Glycation by Natural Compounds. Molecules 2015, 20, 3309–3334. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wu, L.; Wu, Y.; Zhang, C.; Qin, L.; Hayashi, M.; Kudo, M.; Gao, M.; Liu, T. Therapeutic Potential of Centella Asiatica and Its Triterpenes: A Review. Front. Pharmacol. 2020, 11, 568032. [Google Scholar] [CrossRef]
- Kim, Y.J.; Cha, H.J.; Nam, K.H.; Yoon, Y.; Lee, H.; An, S. Centella Asiatica Extracts Modulate Hydrogen Peroxide-Induced Senescence in Human Dermal Fibroblasts. Exp. Dermatol. 2011, 20, 998–1003. [Google Scholar] [CrossRef]
- Eghbali, S.; Askari, S.F.; Avan, R.; Sahebkar, A. Therapeutic Effects of Punica Granatum (Pomegranate): An Updated Review of Clinical Trials. J. Nutr. Metab. 2021, 2021, 5297162. [Google Scholar] [CrossRef]
- Pacheco-Palencia, L.A.; Noratto, G.; Hingorani, L.; Talcott, S.T.; Mertens-Talcott, S.U. Protective Effects of Standardized Pomegranate (Punica granatum L.) Polyphenolic Extract in Ultraviolet-Irradiated Human Skin Fibroblasts. J. Agric. Food Chem. 2008, 56, 8434–8441. [Google Scholar] [CrossRef]
- Wang, N.; Ji, S.; Zhang, H.; Mei, S.; Qiao, L.; Jin, X. Herba Cistanches: Anti-Aging. Aging Dis. 2017, 8, 740. [Google Scholar] [CrossRef]
- Garg, A.; Garg, S.; Zaneveld, L.J.; Singla, A.K. Chemistry and Pharmacology of the Citrus Bioflavonoid Hesperidin. Phytother. Res. PTR 2001, 15, 655–669. [Google Scholar] [CrossRef]
- Man, M.-Q.; Yang, B.; Elias, P.M. Benefits of Hesperidin for Cutaneous Functions. Evid.-Based Complement. Altern. Med. ECAM 2019, 2019, 2676307. [Google Scholar] [CrossRef]
- Bylka, W.; Znajdek-Awiżeń, P.; Studzińska-Sroka, E.; Brzezińska, M. Centella Asiatica in Cosmetology. Adv. Dermatol. Allergol. Dermatol. Alergol. 2013, 30, 46–49. [Google Scholar] [CrossRef]
- Aslam, M.N.; Lansky, E.P.; Varani, J. Pomegranate as a Cosmeceutical Source: Pomegranate Fractions Promote Prolifera-tion and Procollagen Synthesis and Inhibit Matrix Metalloproteinase-1 Production in Human Skin Cells. J. Ethnopharmacol. 2006, 103, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Farris, P.K. Skin Aging, Glycation and Glycation Inhibitors. In Cosmeceuticals and Cosmetic Practice; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 173–183. [Google Scholar] [CrossRef]
- Barltrop, J.A.; Owen, T.C.; Cory, A.H.; Cory, J.G. 5-(3-Carboxymethoxyphenyl)-2-(4,5-Dimethylthiazolyl)-3-(4-Sulfophenyl) Tetrazolium, Inner Salt (MTS) and Related Analogs of 3-(4,5-Dimethylthiazolyl)-2,5-Diphenyltetrazolium Bromide (MTT) Reducing to Purple Water-Soluble Formazans As Cell-Viability Indicators. Bioorg. Med. Chem. Lett. 1991, 1, 611–614. [Google Scholar] [CrossRef]
- van Meerloo, J.; Kaspers, G.J.L.; Cloos, J. Cell Sensitivity Assays: The MTT Assay. Methods Mol. Biol. Clifton NJ 2011, 731, 237–245. [Google Scholar] [CrossRef]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an Aqueous Soluble Tetrazolium/Formazan Assay for Cell Growth Assays in Culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef]
- Bladier, C.; Wolvetang, E.J.; Hutchinson, P.; de Haan, J.B.; Kola, I. Response of a Primary Human Fibroblast Cell Line to H2O2: Senescence-like Growth Arrest or Apoptosis? Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1997, 8, 589–598. [Google Scholar]
- Hahn, H.J.; Kim, K.B.; An, I.-S.; Ahn, K.J.; Han, H.J. Protective Effects of Rosmarinic Acid against Hydrogen Peroxide-induced Cellular Senescence and the Inflammatory Response in Normal Human Dermal Fibroblasts. Mol. Med. Rep. 2017, 16, 9763–9769. [Google Scholar] [CrossRef]
- MTT Assay Protocol 17. Available online: http://cidportal.jrc.ec.europa.eu/ftp/jrc-opendata/EURL-ECVAM/datasets/DBALM/LATEST/online/DBALM_docs/17_P_MTT%20Assay.pdf (accessed on 15 November 2022).
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- de Pedro, N.; Díez, M.; García, I.; García, J.; Otero, L.; Fernández, L.; García, B.; González, R.; Rincón, S.; Pérez, D.; et al. Analytical Validation of Telomere Analysis Technology® for the High-Throughput Analysis of Multiple Telomere-Associated Variables. Biol. Proced. Online 2020, 22, 2. [Google Scholar] [CrossRef]
- Samuel, P.; Tsapekos, M.; de Pedro, N.; Liu, A.G.; Casey Lippmeier, J.; Chen, S. Ergothioneine Mitigates Telomere Shortening under Oxidative Stress Conditions. J. Diet. Suppl. 2022, 19, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Herbert, B.-S.; Hochreiter, A.E.; Wright, W.E.; Shay, J.W. Nonradioactive Detection of Telomerase Activity Using the Telomeric Repeat Amplification Protocol. Nat. Protoc. 2006, 1, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Xu, D.; Björkholm, M.; Gruber, A. Real-Time Quantitative Telomeric Repeat Amplification Protocol Assay for the Detection of Telomerase Activity. Clin. Chem. 2001, 47, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Wege, H.; Chui, M.S.; Le, H.T.; Tran, J.M.; Zern, M.A. SYBR Green Real-Time Telomeric Repeat Amplification Protocol for the Rapid Quantification of Telomerase Activity. Nucleic Acids Res. 2003, 31, e3. [Google Scholar] [CrossRef]
- Dwyer, T.; Muller, H.K.; Blizzard, L.; Ashbolt, R.; Phillips, G. The Use of Spectrophotometry to Estimate Melanin Density in Caucasians. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 1998, 7, 203–206. [Google Scholar] [CrossRef][Green Version]
- Ou-Yang, H.; Stamatas, G.; Kollias, N. Spectral Responses of Melanin to Ultraviolet A Irradiation. J. Investig. Dermatol. 2004, 122, 492–496. [Google Scholar] [CrossRef]
- Sorg, O. Oxidative Stress: A Theoretical Model or a Biological Reality? Comptes Rendus. Biol. 2004, 327, 649–662. [Google Scholar] [CrossRef]
- Tobin, D.J. Introduction to Skin Aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef]
- Pineda-Pampliega, J.; Herrera-Dueñas, A.; Mulder, E.; Aguirre, J.I.; Höfle, U.; Verhulst, S. Antioxidant Supplementation Slows Telomere Shortening in Free-Living White Stork Chicks. Proc. R. Soc. B Biol. Sci. 2020, 287, 20191917. [Google Scholar] [CrossRef]
- Zgheib, N.K.; Sleiman, F.; Nasreddine, L.; Nasrallah, M.; Nakhoul, N.; Isma’eel, H.; Tamim, H. Short Telomere Length Is Associated with Aging, Central Obesity, Poor Sleep and Hypertension in Lebanese Individuals. Aging Dis. 2018, 9, 77–89. [Google Scholar] [CrossRef]
- Shammas, M.A. Telomeres, Lifestyle, Cancer, and Aging. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Tang, M.; Xin, H.; Huang, J. Assessing Telomerase Activities in Mammalian Cells Using the Quantitative PCR-Based Telomeric Repeat Amplification Protocol (qTRAP). Methods Mol. Biol. 2017, 1587, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, R.L.; Ziegler, T.D.; Supakorndej, T.; Terns, R.M.; Terns, M.P. Cell Cycle-Regulated Trafficking of Human Telomerase to Telomeres. Mol. Biol. Cell 2006, 17, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, Z.-P.; Dong, J.; Liao, T.-T.; Wang, Y.-P.; Sun, X.-P.; Yan, Z.-J.; Qian, X.-Q.; Cui, Y.-G.; Xue, Z.; et al. Evaluation of X-Inactivation Status and Cytogenetic Stability of Human Dermal Fibroblasts after Long-Term Culture. Int. J. Cell Biol. 2010, 2010, 289653. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Farrar, M. Advanced Glycation End Products in Skin Ageing and Photoageing: What Are the Implications for Epidermal Function? Exp. Dermatol. 2016, 25, 947–948. [Google Scholar] [CrossRef]
- Kang, H.Y.; Lee, J.W.; Papaccio, F.; Bellei, B.; Picardo, M. Alterations of the Pigmentation System in the Aging Process. Pigment Cell Melanoma Res. 2021, 34, 800–813. [Google Scholar] [CrossRef]
- Brenner, M.; Hearing, V.J. The Protective Role of Melanin Against UV Damage in Human Skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef]
- Marrot, L.; Meunier, J.-R. Skin DNA Photodamage and Its Biological Consequences. J. Am. Acad. Dermatol. 2008, 58 (Suppl. 2), S139–S148. [Google Scholar] [CrossRef]
- Panich, U.; Onkoksoong, T.; Limsaengurai, S.; Akarasereenont, P.; Wongkajornsilp, A. UVA-Induced Melanogenesis and Modulation of Glutathione Redox System in Different Melanoma Cell Lines: The Protective Effect of Gallic Acid. J. Photochem. Photobiol. B 2012, 108, 16–22. [Google Scholar] [CrossRef]
- Naylor, E.C.; Watson, R.E.B.; Sherratt, M.J. Molecular Aspects of Skin Ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- Spagnuolo, L.; Della Posta, S.; Fanali, C.; Dugo, L.; De Gara, L. Antioxidant and Antiglycation Effects of Polyphenol Compounds Extracted from Hazelnut Skin on Advanced Glycation End-Products (AGEs) Formation. Antioxidants 2021, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Papaccio, F.; D’arino, A.; Caputo, S.; Bellei, B. Focus on the Contribution of Oxidative Stress in Skin Aging. Antioxidants 2022, 11, 1121. [Google Scholar] [CrossRef] [PubMed]
- Varesi, A.; Chirumbolo, S.; Campagnoli, L.I.M.; Pierella, E.; Piccini, G.B.; Carrara, A.; Ricevuti, G.; Scassellati, C.; Bonvicini, C.; Pascale, A. The Role of Antioxidants in the Interplay between Oxidative Stress and Senescence. Antioxidants 2022, 11, 1224. [Google Scholar] [CrossRef] [PubMed]
- Dunaway, S.; Odin, R.; Zhou, L.; Ji, L.; Zhang, Y.; Kadekaro, A.L. Natural Antioxidants: Multiple Mechanisms to Protect Skin From Solar Radiation. Front. Pharmacol. 2018, 9, 392. [Google Scholar] [CrossRef]
- Williamson, E.M. Synergy and Other Interactions in Phytomedicines. Phytomed. Int. J. Phytother. Phytopharm. 2001, 8, 401–409. [Google Scholar] [CrossRef]
- Tsoukalas, D.; Fragkiadaki, P.; Docea, A.O.; Alegakis, A.K.; Sarandi, E.; Thanasoula, M.; Spandidos, D.A.; Tsatsakis, A.; Razgonova, M.P.; Calina, D. Discovery of Potent Telomerase Activators: Unfolding New Therapeutic and Anti-Aging Perspectives. Mol. Med. Rep. 2019, 20, 3701–3708. [Google Scholar] [CrossRef]
- Bruno, E.J.; Simpson, G.D.; Martin, R.L. Extending Telomere Length with a Multivitamin: A Pilot Study. J. Health Educ. Res. Dev. 2017, 5, 1–3. [Google Scholar] [CrossRef]
- Khotimah, H.; Ismail, D.D.L.; Widasmara, D.; Riawan, W.; Retnaningtyas, E.; Nugraheni, R.W.; Puspita, O.E.; Adianingsih, O.R.; Mardiyah, M.; Setiawan, A. Ameliorative Effect of Gel Combination of Centella Asiatica Extract Transfersomes and Rosemary Essential Oil Nanoemulsion against UVB-Induced Skin Aging in Balb/c Mice. F1000Research 2022, 11, 288. [Google Scholar] [CrossRef]
- Guo, C.; Zhang, H.; Guan, X.; Zhou, Z. The Anti-Aging Potential of Neohesperidin and Its Synergistic Effects with Other Citrus Flavonoids in Extending Chronological Lifespan of Saccharomyces Cerevisiae BY4742. Molecules 2019, 24, 4093. [Google Scholar] [CrossRef]
- Li, Z.; Lin, H.; Gu, L.; Gao, J.; Tzeng, C.-M. Herba Cistanche (Rou Cong-Rong): One of the Best Pharmaceutical Gifts of Traditional Chinese Medicine. Front. Pharmacol. 2016, 7, 41. [Google Scholar] [CrossRef]
- Yang, W.T.; Kim, K.S.; Kwon, Y.S.; Kim, D.H.; Kim, D.H. Whitening and anti-aging effects of Cistanche deserticola extract. J. Plant Biotechnol. 2016, 43, 492–499. [Google Scholar] [CrossRef]
- Vertuani, S.; Beghelli, E.; Scalambra, E.; Malisardi, G.; Copetti, S.; Dal Toso, R.; Baldisserotto, A.; Manfredini, S. Activity and Stability Studies of Verbascoside, a Novel Antioxidant, in Dermo-Cosmetic and Pharmaceutical Topical Formulations. Molecules 2011, 16, 7068–7080. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.-O.; Lee, S.-A.; Kim, S.-S.; Jang, Y.-S.; Chun, J.-C.; Lee, J.-C. Acteoside Inhibits Melanogenesis in B16F10 Cells through ERK Activation and Tyrosinase Down-Regulation. J. Pharm. Pharmacol. 2011, 63, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.G.; Mikhal’chik, E.; Suprun, M.V.; Pastore, S.; Dal Toso, R. Molecular Mechanisms Underlying Wound Healing and Anti-Inflammatory Properties of Naturally Occurring Biotechnologically Produced Phenylpropanoid Glycosides. Cell. Mol. Biol. 2007, 53, 84–91. [Google Scholar] [PubMed]
- Zhang, H.; Weng, X.; Chen, L.; Chin, L. Effect of Cistanche Tubulosa (Scheuk) Whight Acteoside on Telomerase Activity and Immunity of Aging Mice. Chin. J. Pharmacol. Toxicol. 2008, 22, 270–273. [Google Scholar] [CrossRef]
- Xie, H.; Zhu, H.; Cheng, C.; Liang, Y.; Wang, Z. Echinacoside Retards Cellular Senescence of Human Fibroblastic Cells MRC-5. Pharm. 2009, 64, 752–754. [Google Scholar]
- Yuan, L.I.; Yuanyuan, S.; Chunming, C.H.U.; Hongquan, Z. Study on the Anti-Aging Effect of Echinacoside. Zhongguo Yaoxue Zazhi 2011, 46, 1077–1080. [Google Scholar]
- Li, C.; Schluesener, H. Health-Promoting Effects of the Citrus Flavanone Hesperidin. Crit. Rev. Food Sci. Nutr. 2017, 57, 613–631. [Google Scholar] [CrossRef]
- Lee, H.J.; Im, A.-R.; Kim, S.-M.; Kang, H.-S.; Lee, J.D.; Chae, S. The Flavonoid Hesperidin Exerts Anti-Photoaging Effect by Downregulating Matrix Metalloproteinase (MMP)-9 Expression via Mitogen Activated Protein Kinase (MAPK)-Dependent Signaling Pathways. BMC Complement. Altern. Med. 2018, 18, 39. [Google Scholar] [CrossRef]
- Sun, K.; Xiang, L.; Ishihara, S.; Matsuura, A.; Sakagami, Y.; Qi, J. Anti-Aging Effects of Hesperidin on Saccharomyces Cerevisiae via Inhibition of Reactive Oxygen Species and UTH1 Gene Expression. Biosci. Biotechnol. Biochem. 2012, 76, 640–645. [Google Scholar] [CrossRef]
- Herath, H.M.U.L.; Piao, M.J.; Kang, K.A.; Zhen, A.X.; Fernando, P.D.S.M.; Kang, H.K.; Yi, J.M.; Hyun, J.W. Hesperidin Exhibits Protective Effects against PM2.5-Mediated Mitochondrial Damage, Cell Cycle Arrest, and Cellular Senescence in Human HaCaT Keratinocytes. Molecules 2022, 27, 4800. [Google Scholar] [CrossRef] [PubMed]
- Lorzadeh, E.; Heidary, Z.; Mohammadi, M.; Nadjarzadeh, A.; Ramezani-Jolfaie, N.; Salehi-Abargouei, A. Does Pomegranate Consumption Improve Oxidative Stress? A Systematic Review and Meta-Analysis of Randomized Controlled Clinical Trials. Clin. Nutr. ESPEN 2022, 47, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Afaq, F.; Malik, A.; Syed, D.; Maes, D.; Matsui, M.S.; Mukhtar, H. Pomegranate Fruit Extract Modulates UV-B-Mediated Phosphorylation of Mitogen-Activated Protein Kinases and Activation of Nuclear Factor Kappa B in Normal Human Epidermal Keratinocytes Paragraph Sign. Photochem. Photobiol. 2005, 81, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.K.; Lee, J.-W.; Kim, Y.M.; Boo, Y.C. Punicalagin and (-)-Epigallocatechin-3-Gallate Rescue Cell Viability and Attenuate Inflammatory Responses of Human Epidermal Keratinocytes Exposed to Airborne Particulate Matter PM10. Skin Pharmacol. Physiol. 2018, 31, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.; Dadmohammadi, Y.; Abbaspourrad, A. Nutritional and Bioactive Components of Pomegranate Waste Used in Food and Cosmetic Applications: A Review. Foods 2021, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.-Y.; Xu, K.; Wang, X.; Wen, Y.-T.; Wang, L.-J.; Huang, D.-Q.; Chen, X.-X.; Chai, W.-M. Punicalagin as a Novel Tyrosinase and Melanin Inhibitor: Inhibitory Activity and Mechanism. LWT 2022, 161, 113318. [Google Scholar] [CrossRef]
- Toussaint, O.; Royer, V.; Salmon, M.; Remacle, J. Stress-Induced Premature Senescence and Tissue Ageing. Biochem. Pharmacol. 2002, 64, 1007–1009. [Google Scholar] [CrossRef]
- Ortiz-Espín, A.; Morel, E.; Juarranz, Á.; Guerrero, A.; González, S.; Jiménez, A.; Sevilla, F. An Extract from the Plant Deschampsia Antarctica Protects Fibroblasts from Senescence Induced by Hydrogen Peroxide. Oxid. Med. Cell. Longev. 2017, 2017, 2694945. [Google Scholar] [CrossRef]
- Toussaint, O.; Medrano, E.E.; von Zglinicki, T. Cellular and Molecular Mechanisms of Stress-Induced Premature Senescence (SIPS) of Human Diploid Fibroblasts and Melanocytes. Exp. Gerontol. 2000, 35, 927–945. [Google Scholar] [CrossRef]
- Choi, M.J.; Kim, B.K.; Park, K.Y.; Yokozawa, T.; Song, Y.O.; Cho, E.J. Anti-Aging Effects of Cyanidin under a Stress-Induced Premature Senescence Cellular System. Biol. Pharm. Bull. 2010, 33, 421–426. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Effect of Antioxidants on the Fibroblast Replicative Lifespan In Vitro. Oxid. Med. Cell. Longev. 2020, 2020, 6423783. [Google Scholar] [CrossRef] [PubMed]
- Koliada, A.K.; Krasnenkov, D.S.; Vaiserman, A.M. Telomeric Aging: Mitotic Clock or Stress Indicator? Front. Genet. 2015, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Peng, S.-F.; Chen, L.; Chen, H.-M.; Cheng, X.-E.; Tang, Y.-H. Association between the Oxidative Balance Score and Telomere Length from the National Health and Nutrition Examination Survey 1999–2002. Oxid. Med. Cell. Longev. 2022, 2022, 1345071. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Smith, D.L.; Esteves, K.; Drury, S. Telomere Length Measurement by QPCR—Summary of Critical Factors and Recommendations for Assay Design. Psychoneuroendocrinology 2019, 99, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G.; Hills, M.; Lansdorp, P.M. Telomere Length Measurement-Caveats and a Critical Assessment of the Available Technologies and Tools. Mutat. Res. 2012, 730, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Hemann, M.T.; Strong, M.A.; Hao, L.Y.; Greider, C.W. The Shortest Telomere, Not Average Telomere Length, Is Critical for Cell Viability and Chromosome Stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef]
- Vaiserman, A.; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front. Genet. 2021, 11, 630186. [Google Scholar] [CrossRef]
- Mehkri, S.; Perez, D.; Najarro, P.; Tsapekos, M.; Bopanna, K.N. Effect of Lutein (Lute-Gen®) on Proliferation Rate and Telomere Length in Vitro and Possible Mechanism of Action. Int. J. Biol. Res. 2019, 4, 84–91. [Google Scholar]
- Guillon, C.; Ferraro, S.; Clément, S.; Bouschbacher, M.; Sigaudo-Roussel, D.; Bonod, C. Glycation by Glyoxal Leads to Profound Changes in the Behavior of Dermal Fibroblasts. BMJ Open Diabetes Res. Care 2021, 9, e002091. [Google Scholar] [CrossRef]
- Gkogkolou, P.; Böhm, M. Advanced Glycation End Products. Dermato-Endocrinology 2012, 4, 259–270. [Google Scholar] [CrossRef]
- Hettwer, S.; Suter, B.; Bänziger, S.; Obermayer, B. Anti-Glycation Skin Care—A Promising Approach for Young, Mature and Diabetic Skin. 4. Available online: https://www.rahn-group.com/en/rahn/download-document/a6e3b8d7-dbfc-4775-b04d-a4fb033a02a2/proteoela_techpaper_ec_032016_en.pdf (accessed on 15 November 2022).
- Chen, C.; Zhang, J.-Q.; Li, L.; Guo, M.; He, Y.; Dong, Y.; Meng, H.; Yi, F. Advanced Glycation End Products in the Skin: Molecular Mechanisms, Methods of Measurement, and Inhibitory Pathways. Front. Med. 2022, 9, 837222. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, W.J.; Chang, S.E.; Lee, G.-Y. Hesperidin, A Popular Antioxidant Inhibits Melanogenesis via Erk1/2 Mediated MITF Degradation. Int. J. Mol. Sci. 2015, 16, 18384–18395. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Compound | RT (min) | Wavelength (nm) | Method |
|---|---|---|---|---|
| A 1 | Hesperidin | 19.5 | 280 | 1 |
| B 1 | Echinacoside | 13.7 | 330 | 1 |
| B 2 | Verbascoside | 16.8 | 330 | 1 |
| C 1 | Punicalagin a | 13.7 | 280 | 2 |
| C 2 | Punicalagin b | 19.3 | 280 | 2 |
| D 1 | Asiaticoside B | 21.2 | 200 | 3 |
| D 2 | Madecassoside | 21.5 | 200 | 3 |
| D3 | Asiaticoside | 23 | 200 | 3 |
| Median Length (bp) | Variation from Untreated Control (%) | 20th Percentile Length (bp) | Variation from Untreated Control (%) | Telomeres <3 kbp (%) | Variation from Untreated Control (%) | ||
|---|---|---|---|---|---|---|---|
| Start (Week 0) | Unaged Control | 9955 | 6704 | 3.9 | |||
| Week 6 | Control H2O2 | 8292 | 5356 | 5.8 | |||
| EY 0.001% | 8854 | 6.78 | 5819 | 8.64 | 4.6 | −20.69 | |
| EY 0.0005% | 9356 | 12.83 | 6385 | 19.21 | 3.6 | −37.93 | |
| EY 0.0001% | 9523 | 14.85 | 6530 | 21.92 | 3.5 | −39.66 | |
| Week 8 | Control H2O2 | 6813 | 3621 | 16.4 | |||
| EY 0.001% | 7088 | 4.04 | 3765 | 3.98 | 16 | −2.44 | |
| EY 0.0005% | 8013 | 17.61 | 4627 | 27.78 | 11.6 | −29.27 | |
| EY 0.0001% | 7626 | 11.93 | 4718 | 30.30 | 9 | −45.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiles, J.; Cabrera, M.; Jones, J.; Tsapekos, M.; Caturla, N. In Vitro Determination of the Skin Anti-Aging Potential of Four-Component Plant-Based Ingredient. Molecules 2022, 27, 8101. https://doi.org/10.3390/molecules27228101
Quiles J, Cabrera M, Jones J, Tsapekos M, Caturla N. In Vitro Determination of the Skin Anti-Aging Potential of Four-Component Plant-Based Ingredient. Molecules. 2022; 27(22):8101. https://doi.org/10.3390/molecules27228101
Chicago/Turabian StyleQuiles, José, Maria Cabrera, Jonathan Jones, Menelaos Tsapekos, and Nuria Caturla. 2022. "In Vitro Determination of the Skin Anti-Aging Potential of Four-Component Plant-Based Ingredient" Molecules 27, no. 22: 8101. https://doi.org/10.3390/molecules27228101
APA StyleQuiles, J., Cabrera, M., Jones, J., Tsapekos, M., & Caturla, N. (2022). In Vitro Determination of the Skin Anti-Aging Potential of Four-Component Plant-Based Ingredient. Molecules, 27(22), 8101. https://doi.org/10.3390/molecules27228101