
Trichoderma Enzymes for Degradation of Aflatoxin B1 and Ochratoxin A

 ,
, 
 ,
,  ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
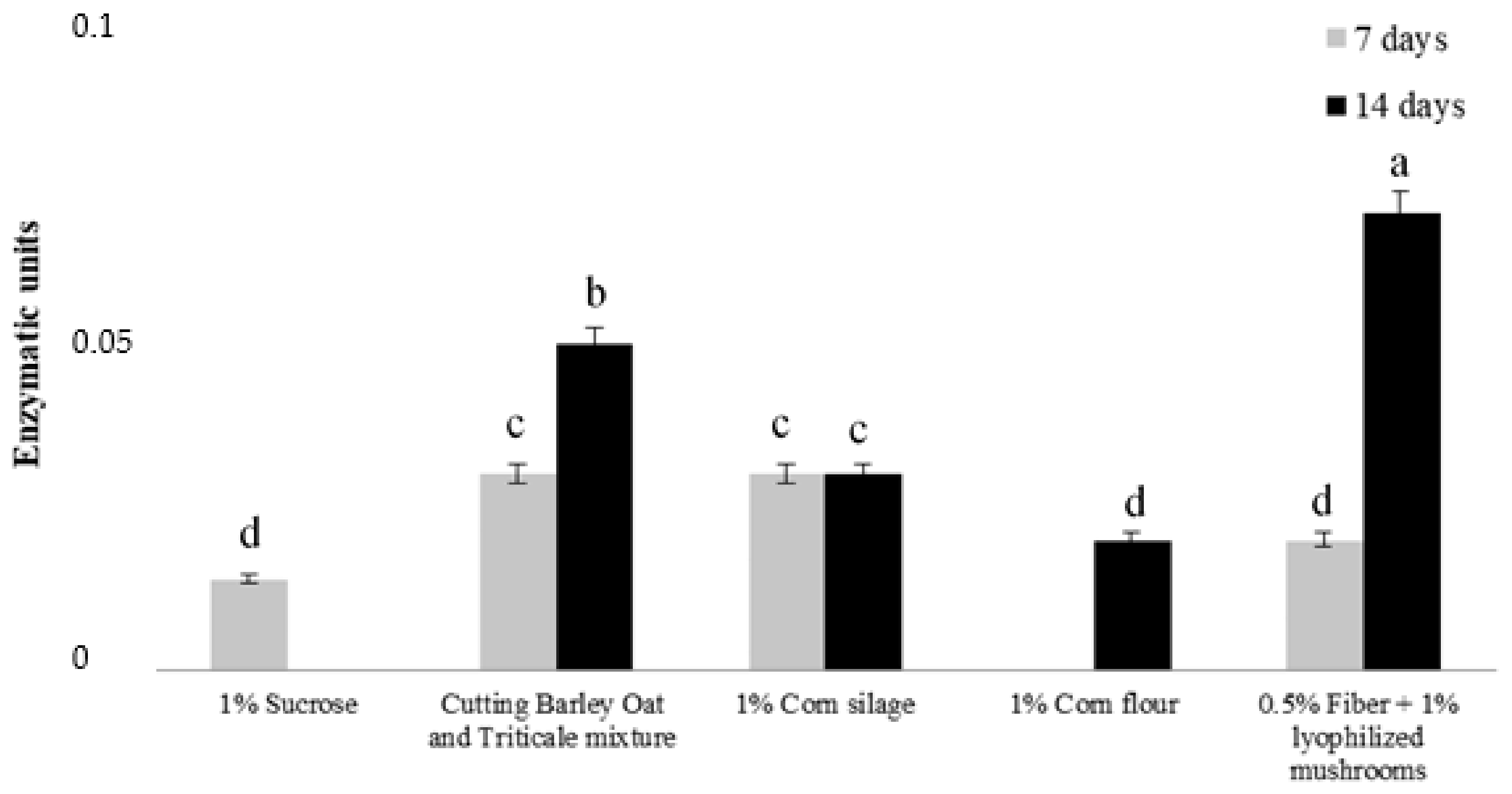
2.1. Peroxidases Production
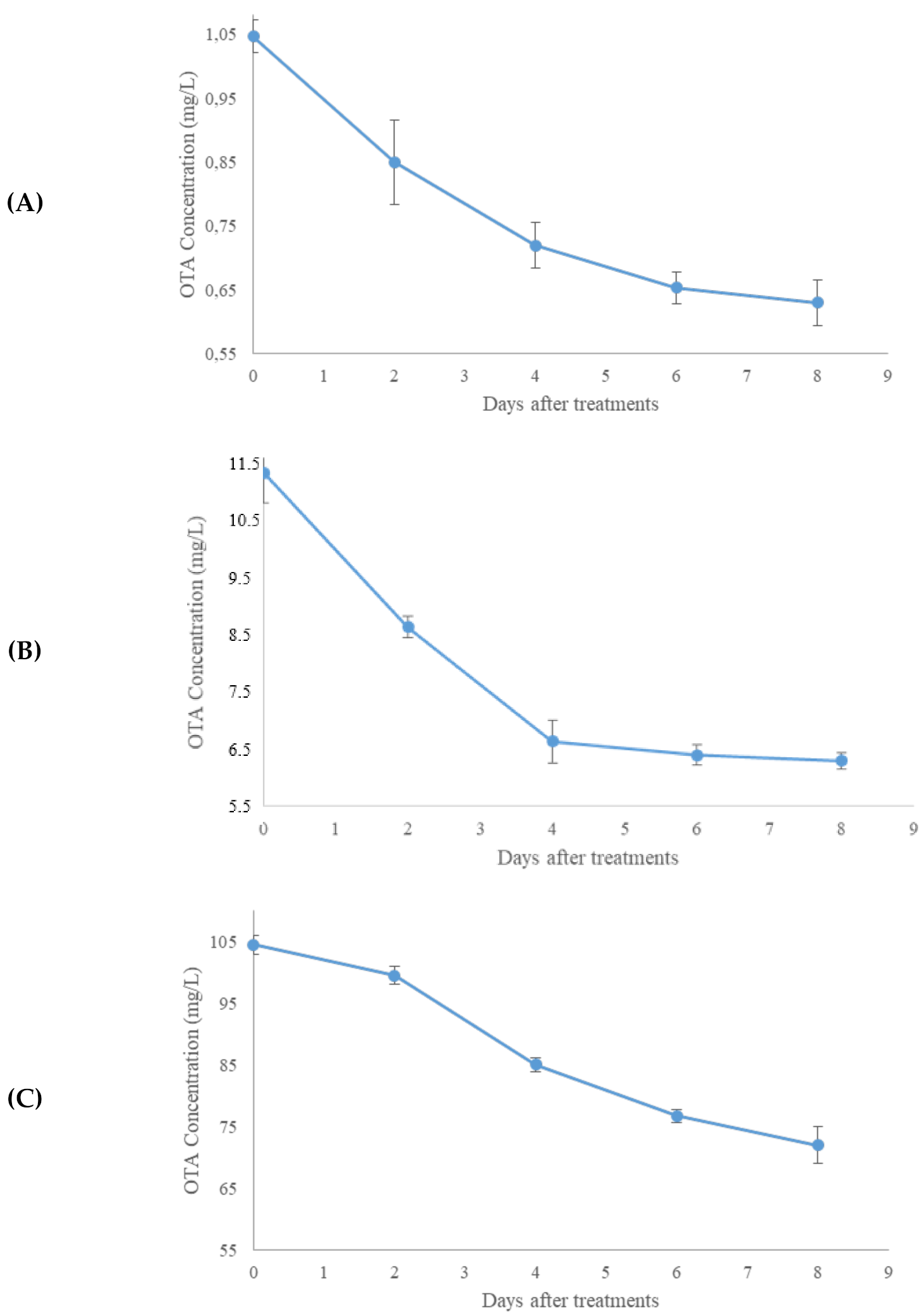
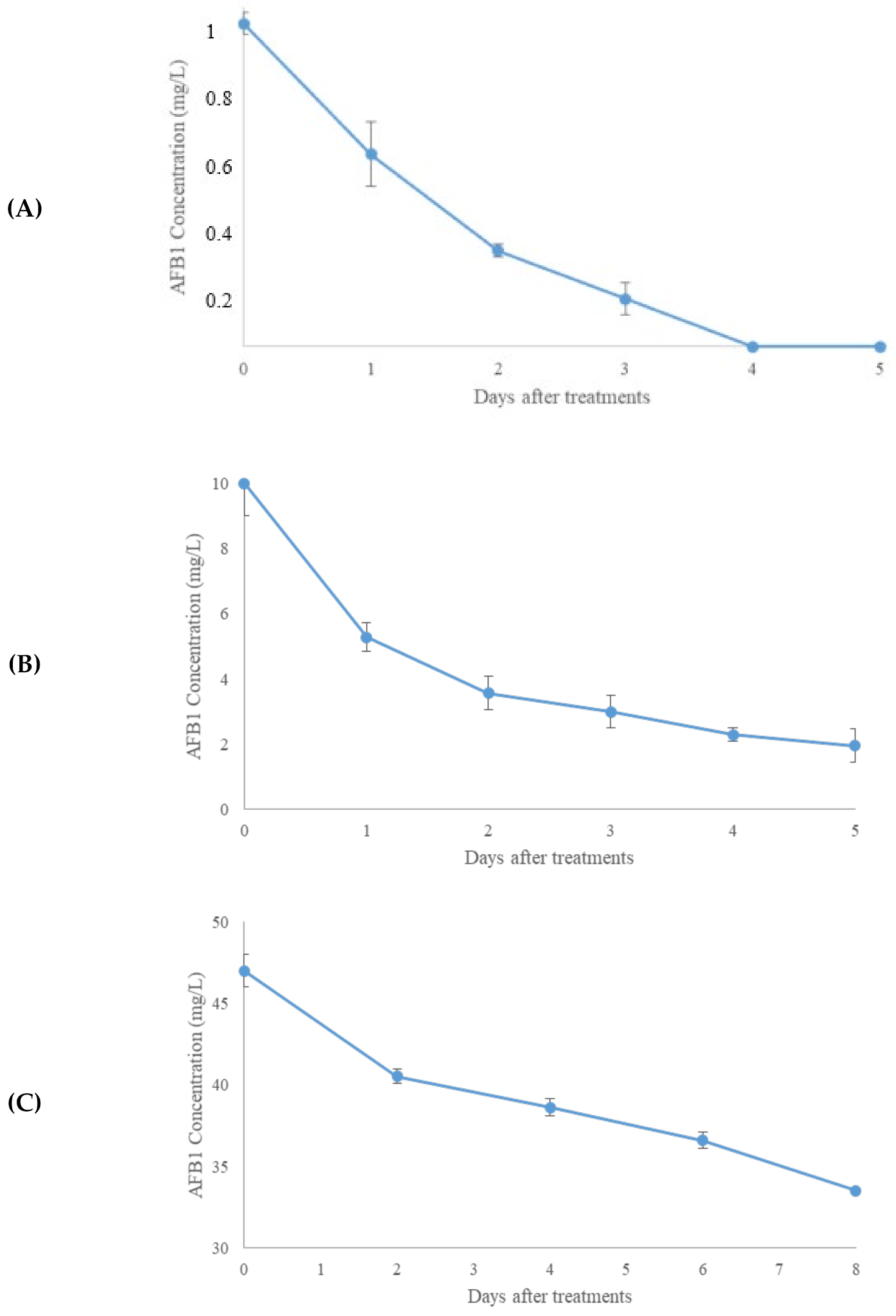
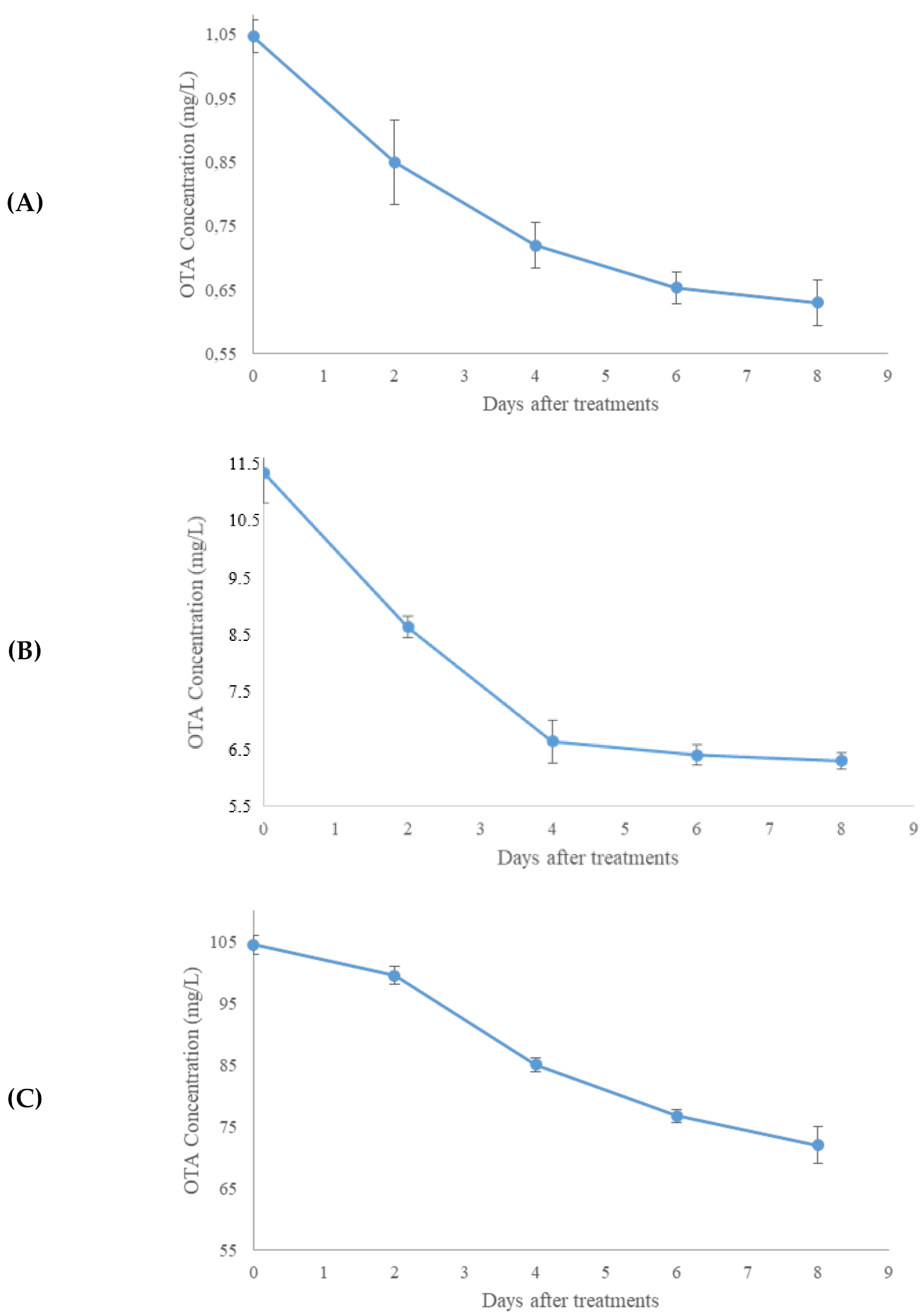
2.2. Aflatoxin B1 and OTA In Vitro Degradation
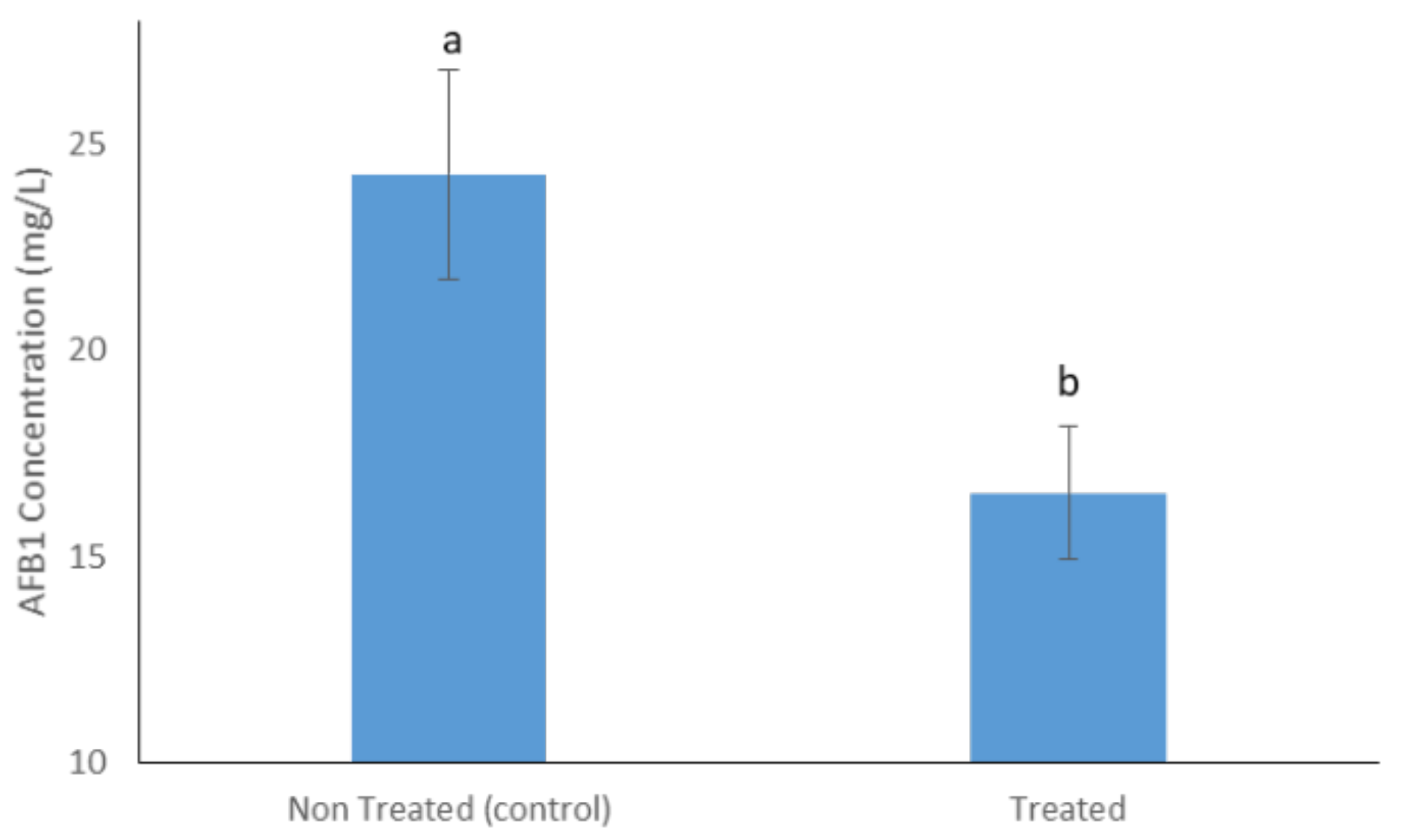
2.3. Aflatoxin B1 and OTA Degradation on Maize Flour and Enzyme Isolation
3. Material and Methods
3.1. Fungal Strains and Growing Conditions
3.2. Peroxidase Assay
3.3. Degradation of Aflatoxin B1 and Ochratoxin A Using Enzymatic Mixtures Produced by Trichoderma
3.3.1. Aflatoxin B1 Quantification
3.3.2. Ochratoxin A Quantification
3.4. Degradation of Aflatoxin B1 in Cornflour
3.5. Extraction of Aflatoxin B1 from Cornflour
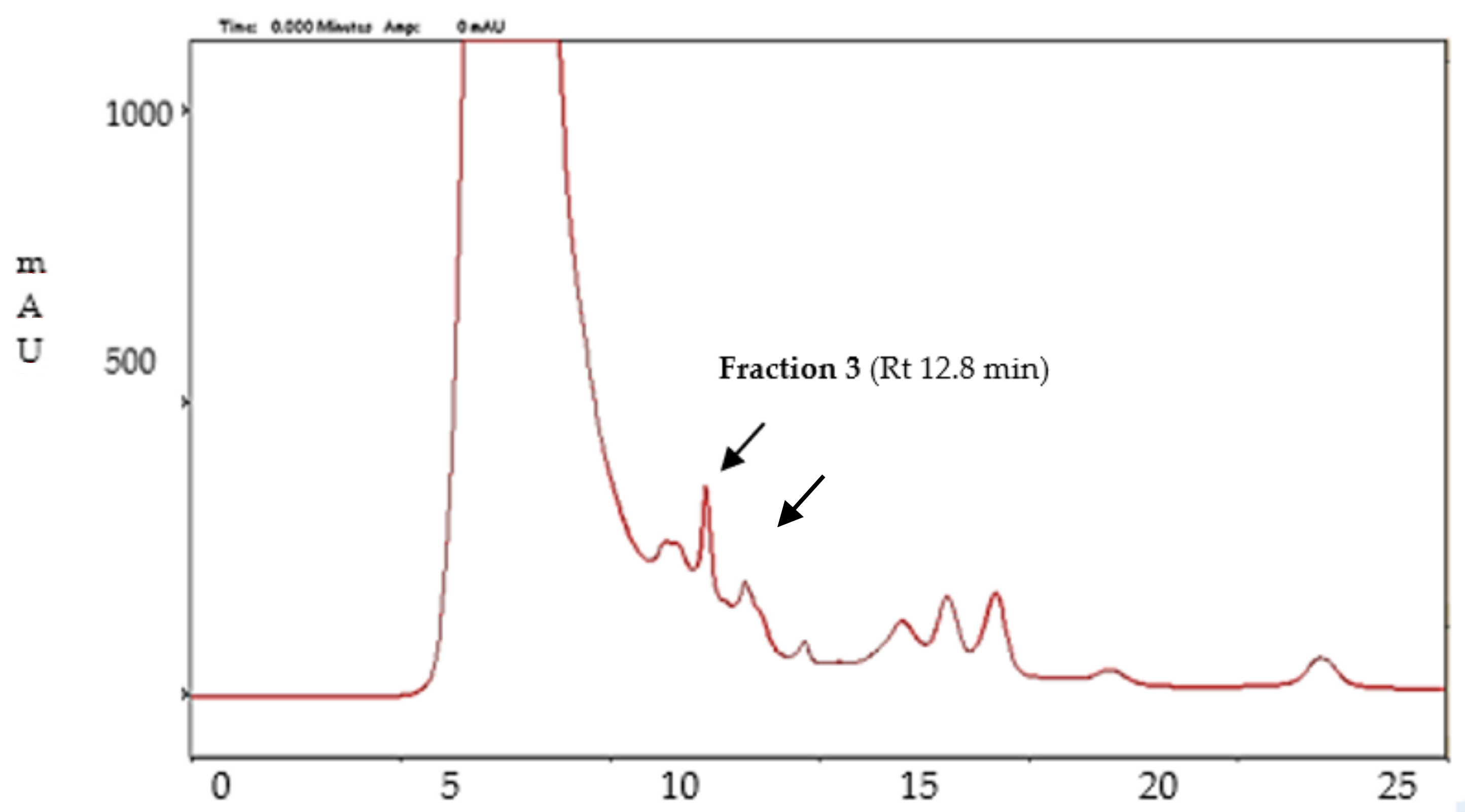
3.6. Fractionation of Enzymatic Mixtures Involved in Aflatoxin B1 Degradation
3.6.1. Gel Filtration
3.6.2. Electrophoresis
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in Analysis and Detection of Major Mycotoxins in Foods. Foods 2020, 9, 518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenier, B.; Applegate, T.J. Modulation of Intestinal Functions Following Mycotoxin Ingestion: Meta-Analysis of Published Experiments in Animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udovicki, B.; Audenaert, K.; De Saeger, S.; Rajkovic, A. Overview on the mycotoxins incidence in Serbia in the period 2004–2016. Toxins 2018, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef]
- Fanelli, F.; Cozzi, G.; Raiola, A.; Dini, I.; Mulè, G.; Logrieco, A.F.; Ritieni, A. Raisins and Currants as Conventional Nutraceuticals in Italian Market: Natural Occurrence of Ochratoxin A. Food Sci. J. 2017, 82, 2306–2312. [Google Scholar] [CrossRef] [PubMed]
- Enyiukwu, D.N.; Awurum, A.N.; Nwaneri, J.A. Mycotoxins in stored agricultural products: Implications to food safety and health and prospects of plant-derived pesticides as novel approach to their management. Greener J. Microbiol. Antimicrob. 2014, 2, 32–48. [Google Scholar] [CrossRef]
- Frangiamone, M.; Cimbalo, A.; Alonso-Garrido, M.; Vila-Donat, P.; Manyes, L. In vitro and in vivo evaluation of AFB1 and OTA-toxicity through immunofluorescence and flow cytometry techniques: A systematic review. Food Chem. Toxicol. 2021, 160, 112798. [Google Scholar] [CrossRef]
- Frangiamone, M.; Alonso-Garrido, M.; Font, G.; Cimbalo, A.; Manyes, L. Pumpkin extract and fermented whey individually and in combination alleviated AFB1-and OTA-induced alterations on neuronal differentiation in vitro. Food Chem. Toxicol. 2022, 164, 113011. [Google Scholar] [CrossRef]
- Čolović, R.; Puvača, N.; Cheli, F.; Avantaggiato, G.; Greco, D.; Đuragić, O.; Kos, J.; Pinotti, L. Decontamination of Mycotoxin-Contaminated Feedstuffs and Compound Feed. Toxins 2019, 11, 617. [Google Scholar] [CrossRef] [Green Version]
- Çelik, K. The efficacy of mycotoxin-detoxifying and biotransforming agents in animal nutrition. In Nanomycotoxicology; Rai, M., Abd-Elsalam, K.A., Eds.; Academic Press: London, UK, 2020; pp. 271–284. [Google Scholar]
- Conte, G.; Fontanelli, M.; Galli, F.; Cotrozzi, L.; Pagni, L.; Pellegrini, E. Mycotoxins in Feed and Food and the Role of Ozone in Their Detoxification and Degradation: An Update. Toxins 2020, 12, 486. [Google Scholar] [CrossRef]
- Haque, M.D.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef] [PubMed]
- Dini, I. Chapter 14—Use of Essential Oils in Food Packaging. In Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Cambridge, MA, USA, 2016; pp. 139–147. [Google Scholar]
- Moncini, L.; Sarrocco, S.; Pachetti, G.; Moretti, A.; Haidukowski, M.; Vannacci, G. N2 controlled atmosphere reduces postharvest mycotoxin risk and pests attack on cereal grains. Phytoparasitica 2020, 48, 555–565. [Google Scholar]
- Alberts, J.F.; Lilly, M.; Rheeder, J.P.; Burger, H.M.; Shephard, G.S.; Gelderblom, W.C.A. Technological and community-based methods to reduce mycotoxin exposure. Food Control 2017, 73, 101–109. [Google Scholar] [CrossRef]
- Dini, I.; Marra, R.; Cavallo, P.; Pironti, A.; Sepe, I.; Troisi, J.; Scala, G.; Lombari, P.; Vinale, F. Trichoderma Strains and Metabolites Selectively Increase the Production of Volatile Organic Compounds (VOCs) in Olive Trees. Metabolites 2021, 11, 213. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Marra, R.; Scala, F.; Ghisalberti, E.; Lorito, M.; Sivasithamparam, K. Major secondary metabolites produced by two commercial Trichoderma strains active against different phytopathogens. Lett. Appl. Microbiol. 2006, 43, 143–148. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Altomare, C.; Norvell, W.A.; Björkman, T.; Harman, G.E. Solubilization of Phosphates and Micronutrients by the Plant-Growth Promoting and Biocontrol Fungus Trichoderma harzianum Rifai 1295-22. Appl. Environ. Microbiol. 1999, 65, 2926–2933. [Google Scholar] [CrossRef] [Green Version]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as biostimulant: Exploiting the multilevel properties of a plant beneficial fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Hermosa, R.; Cardoza, R.E.; Rubio, M.B.; Gutiérrez, S.; Monte, E. Secondary metabolism and antimicrobial metabolites of Trichoderma. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 125–137. [Google Scholar]
- Ren, X.; Branà, M.T.; Haidukowski, M.; Gallo, A.; Zhang, Q.; Logrieco, A.F.; Li, P.; Zhao, S.; Altomare, C. Potential of Trichoderma spp. for Biocontrol of Aflatoxin-Producing Aspergillus flavus. Toxins 2022, 14, 86. [Google Scholar] [CrossRef]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant-pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Valenzuela, N.L.; Angel, D.N.; Ortiz, D.T.; Rosas, R.A.; García, C.F.O.; Santos, M.O. Biological Control of Anthracnose by Postharvest Application of Trichoderma spp. on Maradol Papaya Fruit. Biol. Control 2015, 91, 88–93. [Google Scholar] [CrossRef]
- Dal Bello, G.; Lampugnani, G.; Abramoff, C.; Fusé, C.; Perelló, A. Postharvest Control of Botrytis Gray Mould in Tomato by Antagonists and Biorational Compounds. IOBC-WPRS Bull. 2015, 111, 417–425. [Google Scholar]
- Quaglia, M.; Ederli, L.; Pasqualini, S.; Zazzerini, A. Biological Control Agents and Chemical Inducers of Resistance for Postharvest Control of Penicillium expansum Link. on Apple Fruit. Postharvest Biol. Technol. 2011, 59, 307–315. [Google Scholar] [CrossRef]
- Batta, Y.A. Effect of Treatment with Trichoderma harzianum Rifai Formulated in Invert Emulsion on Postharvest Decay of Apple Blue Mold. Int. J. Food Microbiol. 2004, 96, 281–288. [Google Scholar] [CrossRef]
- Mortuza, M.G.; Ilag, L.L. Potential for Biocontrol of Lasiodiplodia theobromae (Pat.) Griff. & Maubl. in Banana Fruits by Trichoderma Species. Biol. Control 1999, 15, 235–240. [Google Scholar]
- Sangeetha, G.; Usharani, S.; Muthukumar, A. Biocontrol with Trichoderma Species for the Management of Postharvest Crown Rot of Banana. Phytopathol. Mediterr. 2009, 48, 214–225. [Google Scholar]
- Dania, V.O. Bioefficacy of Trichoderma Species against Important Fungal Pathogens Causing Post-Harvest Rot in Sweet Potato (Ipomoea batatas (L.) Lam). J. Bangladesh Agric. Univ. 2019, 17, 446–453. [Google Scholar] [CrossRef] [Green Version]
- Prabakar, K.; Raguchander, T.; Saravanakumar, D.; Muthulakshmi, P.; Parthiban, V.K.; Prakasam, V. Management of Postharvest Disease of Mango Anthracnose Incited by Colletotrichum gleosporioides. Arch. Phytopathol. Plant Prot. 2008, 41, 333–339. [Google Scholar] [CrossRef]
- Nallathambi, P.; Umamaheswari, C.; Thakore, B.B.L.; More, T.A. Post-Harvest Management of Ber (Ziziphus mauritiana Lamk) Fruit Rot (Alternaria alternata Fr. Keissler) Using Trichoderma Species, Fungicides and Their Combinations. Crop Prot. 2009, 28, 525–532. [Google Scholar] [CrossRef]
- Harman, G.E.; Hayes, C.K.; Lorito, M.; Broadway, R.M.; Di Pietro, A.; Tronsmo, A. Chitinolytic enzymes of Trichoderma harzianum: Purification of chitobiosidase and endochitinase. Phytopathology 1993, 83, 313–318. [Google Scholar] [CrossRef]
- Jalili, M.; Jinap, S.; Adzahan, N. Survey of aflatoxins in retail samples of whole and ground black and white peppercorns. Food Addit. Contam. 2009, 2, 178–182. [Google Scholar]
- Jalili, M.; Jinap, S.; Radu, S. Natural occurrence of ochratoxin A contamination in commercial black and white pepper products. Mycopathologia 2010, 170, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shcherbakova, L.A. Fungicide resistance of plant pathogenic fungi and their chemosensitization as a tool to increase anti-disease effects of triazoles and strobilurines. Sel’skokhozyaistvennaya Biol. 2019, 54, 875–891. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.Q.; Wang, M.; Sun, J.; Chen, J. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium stalk rot. Sci. Rep. 2017, 7, 1771. [Google Scholar] [CrossRef]
- Eslahi, N.; Kowsari, M.; Zamani, M.; Motallebi, M. The profile change of defense pathways in Phaseouls vulgaris L. by biochemical and molecular interactions of Trichoderma harzianum transformants overexpressing a chimeric chitinase. Biol. Control 2021, 152, 104304. [Google Scholar] [CrossRef]
- Iannaccone, F.; Alborino, V.; Dini, I.; Balestrieri, A.; Marra, R.; Davino, R.; Di Francia, A.; Masucci, F.; Serrapica, F.; Vinale, F. In Vitro Application of Exogenous Fibrolytic Enzymes from Trichoderma spp. to Improve Feed Utilization by Ruminants. Agriculture 2022, 12, 573. [Google Scholar] [CrossRef]
- Ijoma, G.N.; Selvarajan, R.; Tekere, M. The potential of fungal co-cultures as biological inducers for increased ligninolytic enzymes on agricultural residues. Int. J. Environ. Sci. Technol. 2019, 16, 305–324. [Google Scholar] [CrossRef]
- Nora, N.S.; Feltrin, A.C.P.; Sibaja, K.V.M.; Furlog, E.B.; Garda-Buffon, J. Ochratoxin A reduction by peroxidase in a model system and grape juice. Braz. J. Microbiol. 2019, 50, 1075–1082. [Google Scholar] [CrossRef]
- Tripathi, S.; Mishra, H.N. Modeling and optimization of enzymatic degradation of aflatoxin B1 (AFB1) in red chili powder using response surface methodology. Food Bioprocess Technol. 2011, 4, 770–780. [Google Scholar] [CrossRef]
- Hackbart, H.C.S.; Machado, A.R.; Christ-Ribeiro, A.; Prietto, L.; Badiale-Furlong, E. Reduction of aflatoxins by Rhizopus oryzae and Trichoderma reesei. Mycotoxin Res. 2014, 30, 141–149. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, F.K.; Santos, L.O.; Buffon, J.G. Mechanism of action, sources, and application of peroxidases. Food Res. Int. 2021, 143, 110266. [Google Scholar] [CrossRef] [PubMed]
- Motamed, S.; Ghaemmaghami, F.; Alemzadeh, I. Turnip (Brassica rapa) peroxidase: Purification and characterization. Ind. Eng. Chem. Res. 2009, 48, 10614–10618. [Google Scholar] [CrossRef]
- Alberts, J.F.; Engelbrecht, Y.; Steyn, P.S.; Holzapfel, W.H.; Van Zyl, W.H. Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. Int. J. Food Microbiol. 2006, 109, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Teniola, O.D.; Addo, P.A.; Brost, I.M.; Farber, P.; Jany, K.D.; Alberts, J.F.; Van Zyl, W.H.; Steyn, P.S.; Holzapfel, W.H. Degradation of aflatoxin B1 by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp. nov. DSM44556. Int. J. Food Microbiol. 2005, 105, 11–117. [Google Scholar] [CrossRef]
- Cheng-An, H.; Draughan, F.A. Degradation of Ochratoxin A by Acinetobacter calcoaeticus. J. Food Prot. 1994, 57, 410–414. [Google Scholar]
- Braun, H.; Woitsch, L.; Hetzer, B.; Geisen, R.; Zange, B.; Schmidt-Heydt, M. Trichoderma harzianum: Inhibition of mycotoxin producing fungi and toxin biosynthesis. Int. J. Food Microbiol. 2018, 280, 10–16. [Google Scholar] [CrossRef]
- Denardi de Souza, T.; Caldas, S.S.; Primel, E.G.; Furlong, E.B. Exposure to deoxynivalenol, Ht-2 and T-2 toxins by consumption of wheat-based product in southern Brazil. Food Control 2015, 50, 789–793. [Google Scholar] [CrossRef]
- Guan, Y.; Chen, J.; Nepovimova, E.; Long, M.; Wu, W.; Kuca, K. Aflatoxin Detoxification Using Microorganisms and Enzymes. Toxins 2021, 13, 46. [Google Scholar] [CrossRef]
- Adebo, O.; Njobeh, P.; Gbashi, S.; Nwinyi, O.; Mavumengwana, V. Review on microbial degradation of aflatoxins. Crit. Rev. Food Sci. Nutr. 2017, 57, 3208–3217. [Google Scholar] [CrossRef] [PubMed]
- Magouz, F.; Abu-Ghanima, H.; Zaineldin, A.I.; Gewaily, M.S.; Soliman, A.; Amer, A.A.; Moustafa, E.M.; Younis, E.M.; Abdel-Warith, A.-W.A.; Davies, S.J.; et al. Dietary Bacillus subtilis relieved the growth retardation, hepatic failure, and antioxidative depression induced by ochratoxin A in Thinlip Mullet (Liza ramada). Aquac. Rep. 2022, 22, 100984. [Google Scholar] [CrossRef]
- Mei, L.; Guang–Shu, M.A.; Lian, H.; Su, X.; Tian, Y.; Huang, W.; Mei, J.; Jiang, X. The effects of Trichoderma on preventing cucumber fusarium wilt and regulating cucumber physiology. J. Integr. Agric. 2019, 18, 607–617. [Google Scholar]
- De Oliveira Garcia, S.; Sibaja, K.V.M.; Nogueira, W.V.; Feltrin, A.C.P.; Pinheiro, D.F.A.; Cerqueira, M.B.R.; Furlong, E.B.; Garda-Buffon, J. Peroxidase as a simultaneous degradation agent of ochratoxin A and zearalenone applied to model solution and beer. Food Res. Int. 2020, 131, 109039. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dini, I.; Alborino, V.; Lanzuise, S.; Lombardi, N.; Marra, R.; Balestrieri, A.; Ritieni, A.; Woo, S.L.; Vinale, F. Trichoderma Enzymes for Degradation of Aflatoxin B1 and Ochratoxin A. Molecules 2022, 27, 3959. https://doi.org/10.3390/molecules27123959
Dini I, Alborino V, Lanzuise S, Lombardi N, Marra R, Balestrieri A, Ritieni A, Woo SL, Vinale F. Trichoderma Enzymes for Degradation of Aflatoxin B1 and Ochratoxin A. Molecules. 2022; 27(12):3959. https://doi.org/10.3390/molecules27123959
Chicago/Turabian StyleDini, Irene, Vittoria Alborino, Stefania Lanzuise, Nadia Lombardi, Roberta Marra, Anna Balestrieri, Alberto Ritieni, Sheridan L. Woo, and Francesco Vinale. 2022. "Trichoderma Enzymes for Degradation of Aflatoxin B1 and Ochratoxin A" Molecules 27, no. 12: 3959. https://doi.org/10.3390/molecules27123959
APA StyleDini, I., Alborino, V., Lanzuise, S., Lombardi, N., Marra, R., Balestrieri, A., Ritieni, A., Woo, S. L., & Vinale, F. (2022). Trichoderma Enzymes for Degradation of Aflatoxin B1 and Ochratoxin A. Molecules, 27(12), 3959. https://doi.org/10.3390/molecules27123959