
Cannabinoid Receptor Modulation of Neurogenesis: ST14A Striatal Neural Progenitor Cells as a Simplified In Vitro Model

 ,
,  ,
,
Abstract
1. Introduction
2. Results
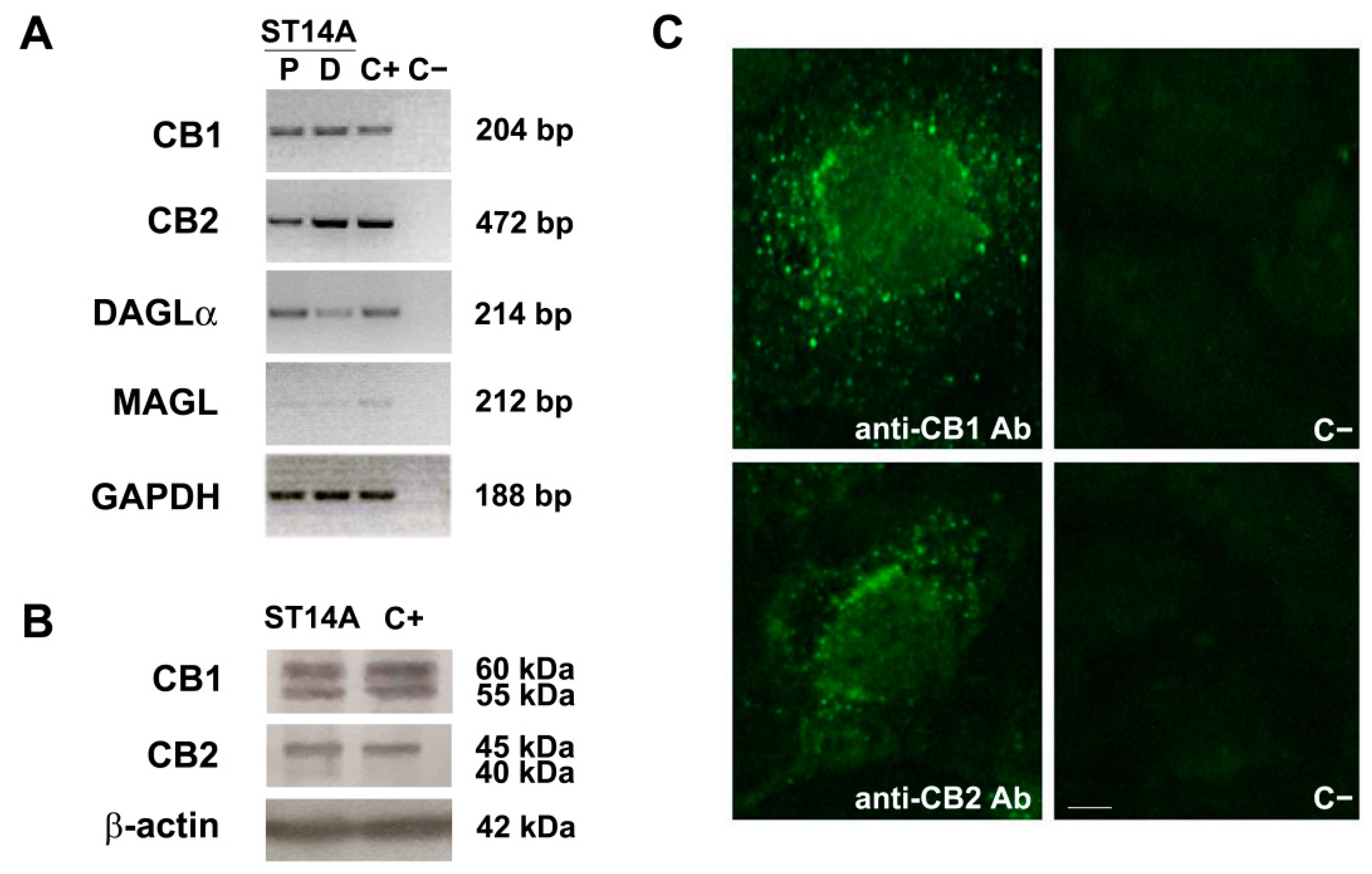
2.1. The Endocannabinoid System Is Expressed in ST14A Striatal Neural Progenitor Cells
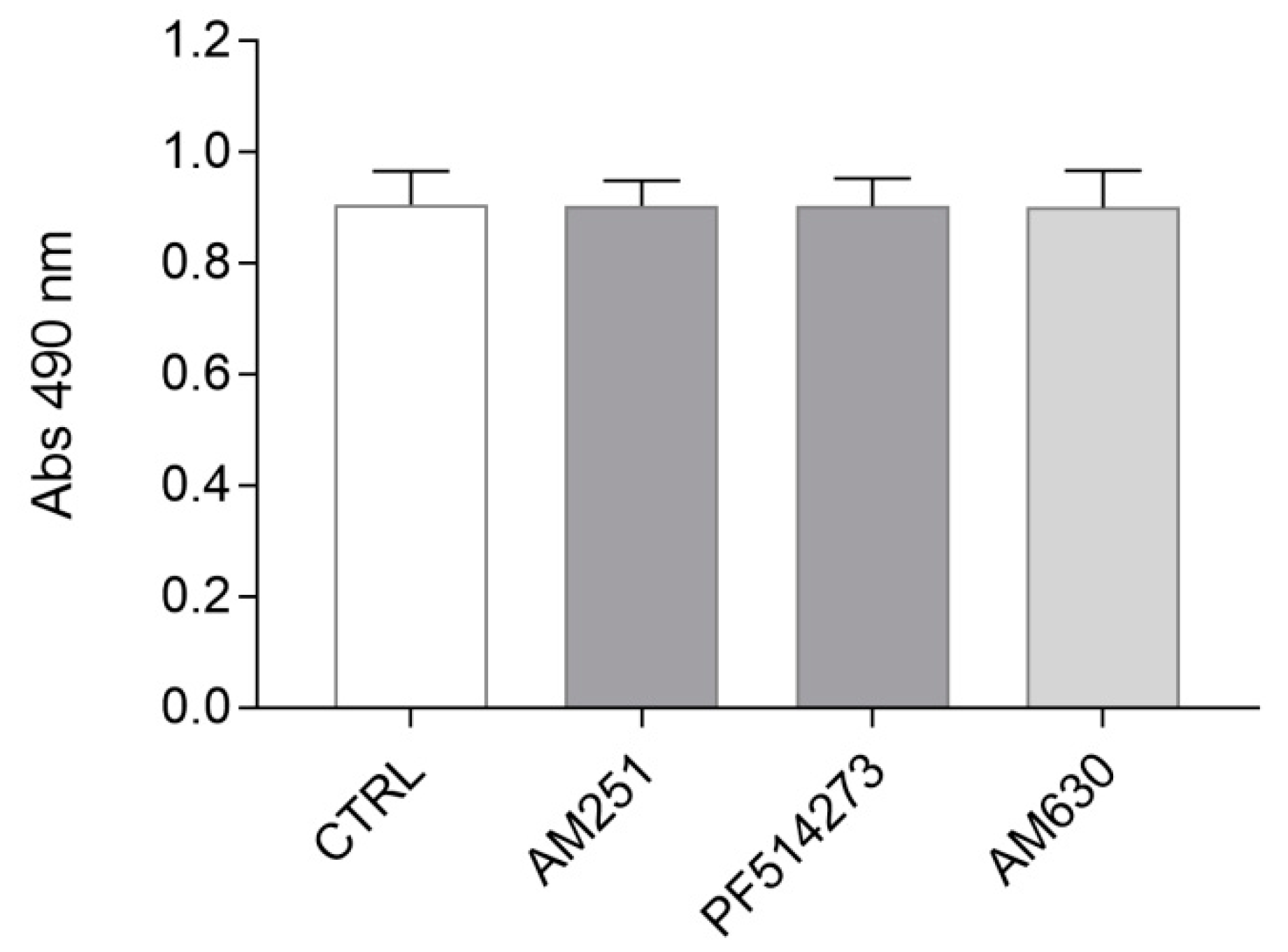
2.2. The Pharmacological Blockade of Cannabinoid Receptors Does Not Affect ST14A Cell Number
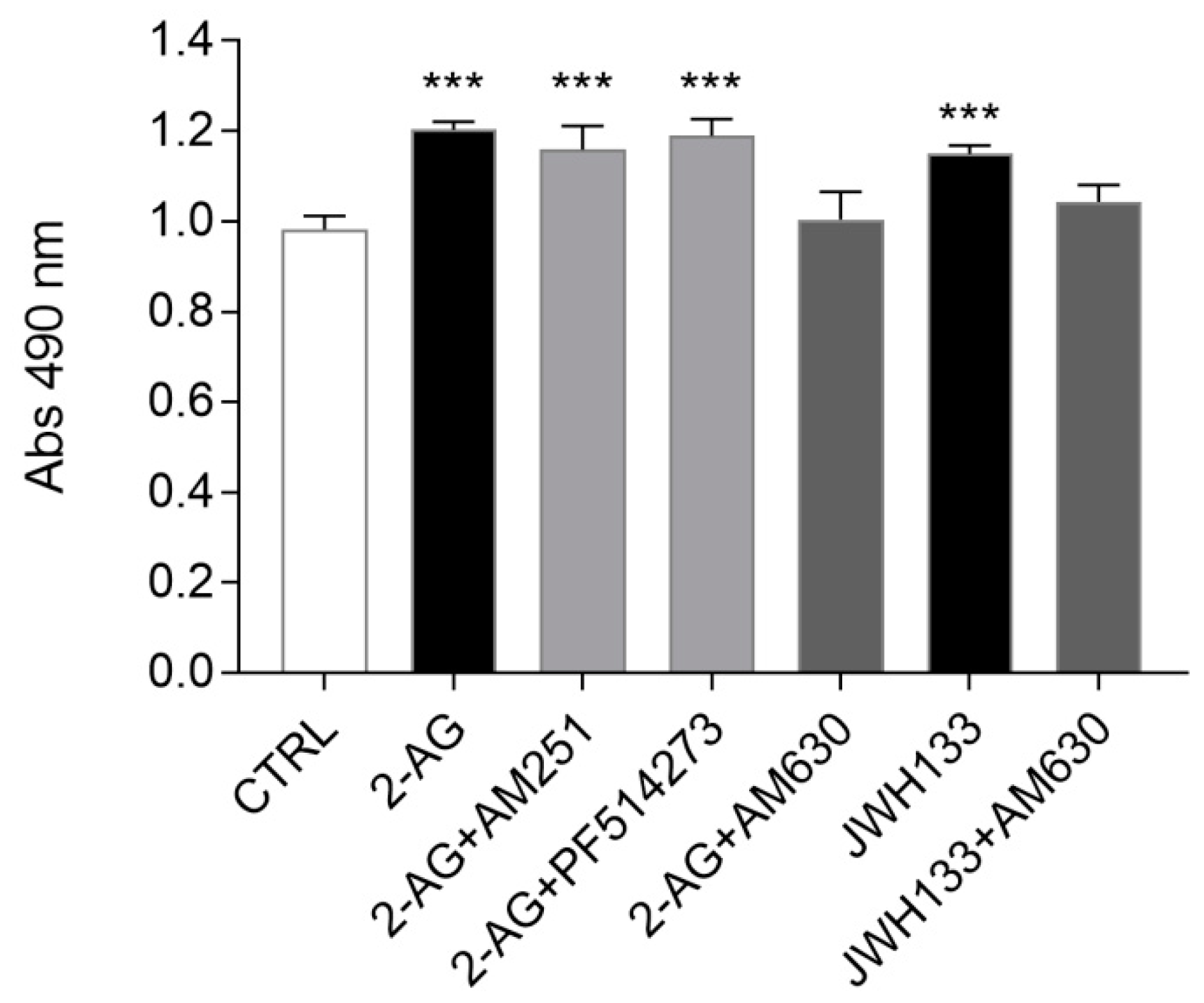
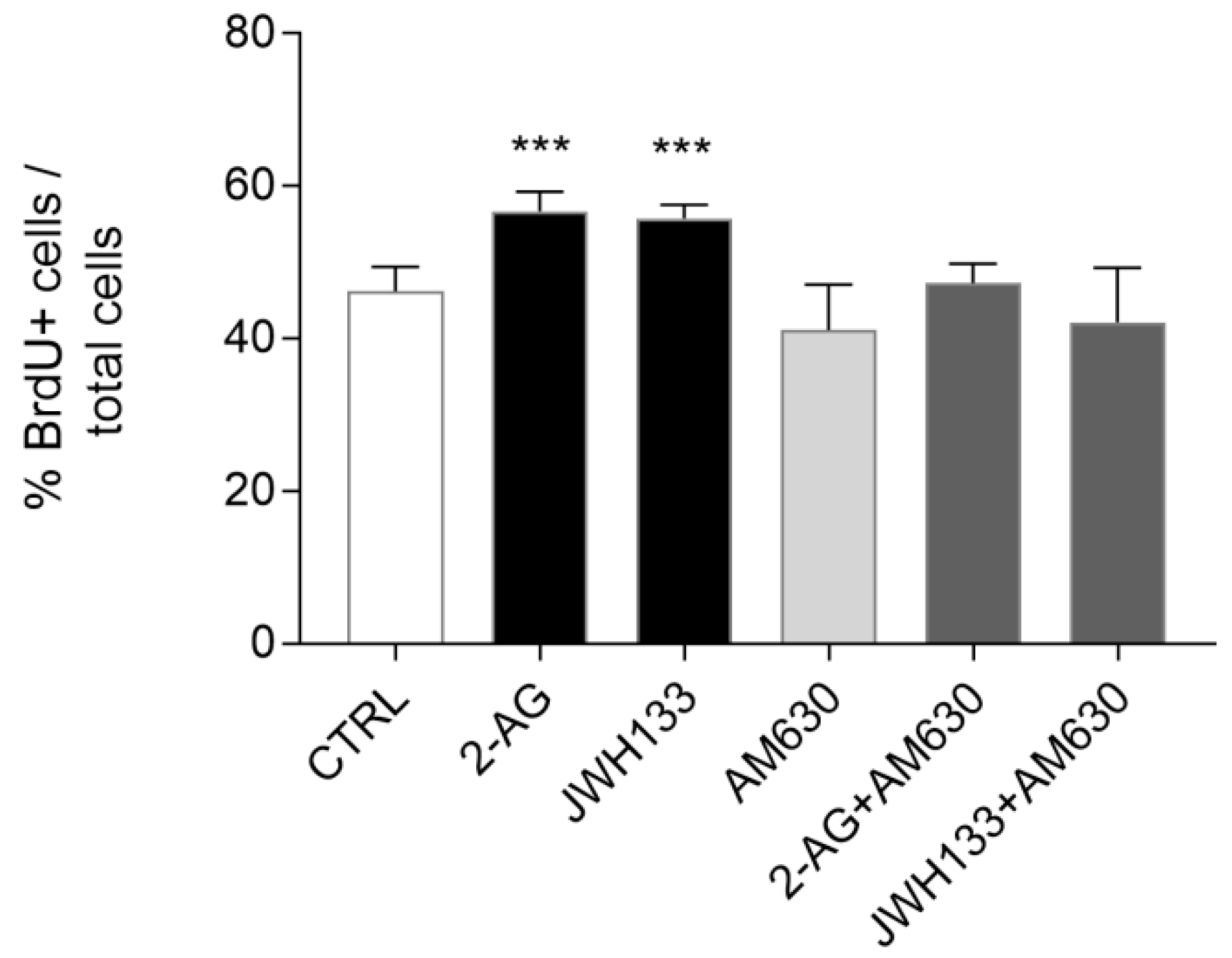
2.3. Exogenous Administration of the Endocannabinoid 2-AG and the CB2 Agonist JWH133 Induces ST14A Cell Proliferation through CB2 Receptor Activation
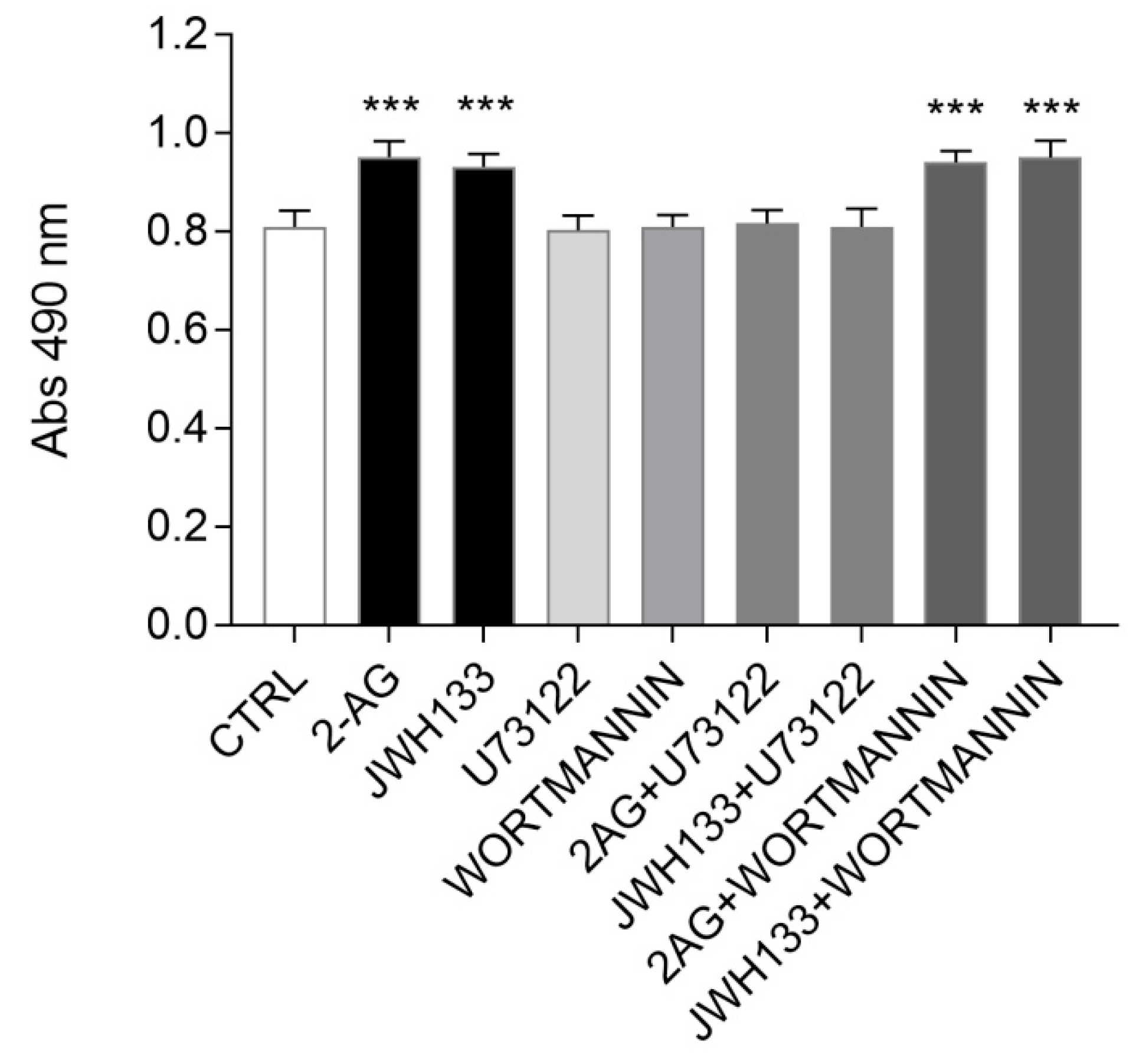
2.4. PLC Pharmacological Blockade Impairs CB2-Mediated ST14A Cell Proliferation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Extraction and RT-PCR
4.3. Western Blot
4.4. Immunofluorescence
4.5. Cell Count Assays
4.6. Cell Proliferation Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statements
Conflicts of Interest
Sample Availability
References
- Lu, H.-C.; Mackie, K. Review of the Endocannabinoid System. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020. [Google Scholar] [CrossRef]
- Di Marzo, V. New approaches and challenges to targeting the endocannabinoid system. Nat. Rev. Drug Discov. 2018, 17, 623–639. [Google Scholar] [CrossRef]
- Shahbazi, F.; Grandi, V.; Banerjee, A.; Trant, J.F. Cannabinoids and Cannabinoid Receptors: The Story so Far. iScience 2020. [Google Scholar] [CrossRef]
- Mechoulam, R.; Gaoni, Y. A Total Synthesis of dl-Δ1-Tetrahydrocannabinol, the Active Constituent of Hashish1. J. Am. Chem. Soc. 1965, 87, 3273–3275. [Google Scholar] [CrossRef]
- Amal, H.; Fridman-Rozevich, L.; Senn, R.; Strelnikov, A.; Gafni, M.; Keren, O.; Sarne, Y. Long-term consequences of a single treatment of mice with an ultra-low dose of Δ9-tetrahydrocannabinol (THC). Behav. Brain Res. 2010, 206, 245–253. [Google Scholar] [CrossRef]
- Bossong, M.G.; Van Berckel, B.N.M.; Boellaard, R.; Zuurman, L.; Schuit, R.C.; Windhorst, A.D.; Van Gerven, J.M.A.; Ramsey, N.F.; Lammertsma, A.A.; Kahn, R.S. Δ9-Tetrahydrocannabinol Induces Dopamine Release in the Human Striatum. Neuropsychopharmacology 2008, 34, 759–766. [Google Scholar] [CrossRef]
- Di Marzo, V.; Piscitelli, F. The Endocannabinoid System and its Modulation by Phytocannabinoids. Neurotherapeutics 2015, 12, 692–698. [Google Scholar] [CrossRef]
- Haspula, D.; Clark, M.A. Cannabinoid Receptors: An Update on Cell Signaling, Pathophysiological Roles and Therapeutic Opportunities in Neurological, Cardiovascular, and Inflammatory Diseases. Int. J. Mol. Sci. 2020, 21, 7693. [Google Scholar] [CrossRef]
- Pertwee, R.G. Endocannabinoids and their pharmacological actions. In Endocannabinoids; Pertwee, R.G., Ed.; Springer: Berlin, Germany, 2015; Volume 231, pp. 1–37. [Google Scholar] [CrossRef]
- De Oliveira, R.W.; Oliveira, C.L.; Guimarães, F.S.; Campos, A.C. Cannabinoid signalling in embryonic and adult neurogenesis: Possible implications for psychiatric and neurological disorders. Acta Neuropsychiatr. 2018, 31, 1–16. [Google Scholar] [CrossRef]
- Lutz, B. Neurobiology of cannabinoid receptor signaling. Dialog Clin. Neurosci. 2020, 22, 207–222. [Google Scholar] [CrossRef]
- Oddi, S.; Scipioni, L.; Maccarrone, M. Endocannabinoid system and adult neurogenesis: A focused review. Curr. Opin. Pharmacol. 2020, 50, 25–32. [Google Scholar] [CrossRef]
- Galiegue, S.; Mary, S.; Marchand, J.; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Fur, G.; Casellas, P. Expression of Central and Peripheral Cannabinoid Receptors in Human Immune Tissues and Leukocyte Subpopulations. JBIC J. Biol. Inorg. Chem. 1995, 232, 54–61. [Google Scholar] [CrossRef]
- Gong, J.-P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.-R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Aymerich, M.S.; Aso, E.; Abellanas, M.A.; Tolon, R.M.; Ramos, J.A.; Ferrer, I.; Romero, J.; Fernández-Ruiz, J. Cannabinoid pharmacology/therapeutics in chronic degenerative disorders affecting the central nervous system. Biochem. Pharmacol. 2018, 157, 67–84. [Google Scholar] [CrossRef]
- Palazuelos, J.; Aguado, T.; Egia, A.; Mechoulam, R.; Guzmán, M.; Galve-Roperh, I. Non-psychoactive CB 2 cannabinoid agonists stimulate neural progenitor proliferation. FASEB J. 2006, 20, 2405–2407. [Google Scholar] [CrossRef] [PubMed]
- Palazuelos, J.; Ortega, Z.; Díaz-Alonso, J.; Guzmán, M.; Galve-Roperh, I. CB2 Cannabinoid Receptors Promote Neural Progenitor Cell Proliferation via mTORC1 Signaling. J. Biol. Chem. 2012, 287, 1198–1209. [Google Scholar] [CrossRef] [PubMed]
- Molina-Holgado, F.; Rubio-Araiz, A.; García-Ovejero, D.; Williams, R.J.; Moore, J.D.; Arévalo-Martín, Á.; Gómez-Torres, O. CB2 cannabinoid receptors promote mouse neural stem cell proliferation. Eur. J. Neurosci. 2007, 25, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Anavi-Goffer, S.; Mulder, J. The Polarised Life of the Endocannabinoid System in CNS Development. ChemBioChem 2009, 10, 1591–1598. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Romero, J.; Velasco, G.; Tolón, R.M.; Ramos, J.A.; Guzmán, M. Cannabinoid CB2 receptor: A new target for controlling neural cell survival? Trends Pharmacol. Sci. 2007, 28, 39–45. [Google Scholar] [CrossRef]
- Galve-Roperh, I.; Chiurchiù, V.; Díaz-Alonso, J.; Bari, M.; Guzmán, M.; Maccarrone, M. Cannabinoid receptor signaling in progenitor/stem cell proliferation and differentiation. Prog. Lipid Res. 2013, 52, 633–650. [Google Scholar] [CrossRef]
- Duff, G.; Argaw, A.; Cecyre, B.; Cherif, H.; Tea, N.; Zabouri, N.; Casanova, C.; Ptito, M.; Bouchard, J.-F. Cannabinoid Receptor CB2 Modulates Axon Guidance. PLoS ONE 2013, 8, e70849. [Google Scholar] [CrossRef]
- Goncalves, M.B.; Suetterlin, P.; Yip, P.; Molina-Holgado, F.; Walker, D.J.; Oudin, M.J.; Zentar, M.P.; Pollard, S.; Yáñez-Muñoz, R.J.; Williams, G.; et al. A diacylglycerol lipase-CB2 cannabinoid pathway regulates adult subventricular zone neurogenesis in an age-dependent manner. Mol. Cell. Neurosci. 2008, 38, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Oudin, M.J.; Gajendra, S.; Williams, G.; Hobbs, C.; Lalli, G.; Doherty, P. Endocannabinoids Regulate the Migration of Subventricular Zone-Derived Neuroblasts in the Postnatal Brain. J. Neurosci. 2011, 31, 4000–4011. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Xie, L.; Kim, S.H.; Parmentier-Batteur, S.; Sun, Y.; Mao, X.O.; Childs, J.; Greenberg, D.A. Defective Adult Neurogenesis in CB1 Cannabinoid Receptor Knockout Mice. Mol. Pharmacol. 2004, 66, 204–208. [Google Scholar] [CrossRef]
- Cattaneo, E.; Conti, L. Generation and characterization of embryonic striatal conditionally immortalized ST14A cells. J. Neurosci. Res. 1998, 53, 223–234. [Google Scholar] [CrossRef]
- Hovakimyan, M.; Weinreich, K.; Haas, S.J.-P.; Cattaneo, E.; Rolfs, A.; Wree, A. In Vitro Characterization of Embryionic ST14A-Cells. Int. J. Neurosci. 2008, 118, 1489–1501. [Google Scholar] [CrossRef]
- Ehrlich, M.E.; Conti, L.; Tosellic, M.; Tagliettic, L.; Fiorilloc, E.; Tagliettic, V.; Ivkovica, S.; Guineaa, B.; Tranberga, A.; Sipioneb, S.; et al. ST14A Cells Have Properties of a Medium-Size Spiny Neuron. Exp. Neurol. 2001, 167, 215–226. [Google Scholar] [CrossRef]
- Rigamonti, D.; Bauer, J.H.; De-Fraja, C.; Conti, L.; Sipione, S.; Sciorati, C.; Clementi, E.; Hackam, A.; Hayden, M.R.; Li, Y.; et al. Wild-Type Huntingtin Protects from Apoptosis Upstream of Caspase-3. J. Neurosci. 2000, 20, 3705–3713. [Google Scholar] [CrossRef]
- Cacci, E.; Salani, M.; Anastasi, S.; Perroteau, I.; Poiana, G.; Biagioni, S.; Augusti-Tocco, G. Hepatocyte growth factor stimulates cell motility in cultures of the striatal progenitor cells ST14A. J. Neurosci. Res. 2003, 74, 760–768. [Google Scholar] [CrossRef]
- Gambarotta, G.; Garzotto, D.; Destro, E.; Mautino, B.; Giampietro, C.; Cutrupi, S.; Dati, C.; Cattaneo, E.; Fasolo, A.; Perroteau, I. ErbB4 Expression in Neural Progenitor Cells (ST14A) Is Necessary to Mediate Neuregulin-1β1-induced Migration. J. Biol. Chem. 2004, 279, 48808–48816. [Google Scholar] [CrossRef]
- Pregno, G.; Zamburlin, P.; Gambarotta, G.; Farcito, S.; Licheri, V.; Fregnan, F.; Perroteau, I.; Lovisolo, D.; Bovolin, P. Neuregulin1/ErbB4-induced migration in ST14A striatal progenitors: Calcium-dependent mechanisms and modulation by NMDA receptor activation. BMC Neurosci. 2011, 12, 103. [Google Scholar] [CrossRef]
- Beyer, S.; Mix, E.; Hoffrogge, R.; Lünser, K.; Völker, U.; Rolfs, A. Neuroproteomics in stem cell differentiation. Proteom. Clin. Appl. 2007, 1, 1513–1523. [Google Scholar] [CrossRef]
- Bari, M.; Battista, N.; Valenza, M.; Mastrangelo, N.; Malaponti, M.; Catanzaro, G.; Centonze, D.; Finazzi-Agrò, A.; Cattaneo, E.; Maccarrone, M. In vitro and in vivo models of Huntington’s disease show alterations in the endocannabinoid system. FEBS J. 2013, 280, 3376–3388. [Google Scholar] [CrossRef]
- Saba, J.; Couselo, F.L.; Turati, J.; Carniglia, L.; Durand, D.; De Laurentiis, A.; Lasaga, M.; Caruso, C. Astrocytes from cortex and striatum show differential responses to mitochondrial toxin and BDNF: Implications for protection of striatal neurons expressing mutant huntingtin. J. Neuroinflam. 2020, 17, 1–15. [Google Scholar] [CrossRef]
- Avraham, H.K.; Jiang, S.; Fu, Y.; Rockenstein, E.; Makriyannis, A.; Zvonok, A.; Masliah, E.; Avraham, S. The cannabinoid CB2receptor agonist AM1241 enhances neurogenesis in GFAP/Gp120 transgenic mice displaying deficits in neurogenesis. Br. J. Pharmacol. 2014, 171, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Aguado, T.; Monory, K.; Palazuelos, J.; Stella‡, N.; Cravatt, B.; Lutz, B.; Marsicano, G.; Kokaia, Z.; Guzmán, M.; Galve-Roperh, I. The endocannabinoid system drives neural progenitor proliferation. FASEB J. 2005, 19, 1704–1706. [Google Scholar] [CrossRef]
- Atwood, B.K.; Lee, D.; Straiker, A.; Widlanski, T.S.; Mackie, K. CP47,497-C8 and JWH073, commonly found in ‘Spice’ herbal blends, are potent and efficacious CB1 cannabinoid receptor agonists. Eur. J. Pharmacol. 2011, 659, 139–145. [Google Scholar] [CrossRef]
- Boon, F.S.D.; Chameau, P.; Schaafsma-Zhao, Q.; Van Aken, W.; Bari, M.; Oddi, S.; Kruse, C.G.; Maccarrone, M.; Wadman, W.J.; Werkman, T.R. Excitability of prefrontal cortical pyramidal neurons is modulated by activation of intracellular type-2 cannabinoid receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 3534–3539. [Google Scholar] [CrossRef] [PubMed]
- Brailoiu, G.C.; Deliu, E.; Marcu, J.; Hoffman, N.E.; Console-Bram, L.; Zhao, P.; Madesh, M.; Abood, M.E.; Brailoiu, E. Differential Activation of Intracellular versus Plasmalemmal CB2Cannabinoid Receptors. Biochem. 2014, 53, 4990–4999. [Google Scholar] [CrossRef] [PubMed]
- Bénard, G.; Massa, F.; Puente, N.; Lourenço, J.; Bellocchio, L.; Soria-Gómez, E.; Matias, I.; Delamarre, A.; Metna-Laurent, M.; Cannich, A.; et al. Mitochondrial CB1 receptors regulate neuronal energy metabolism. Nat. Neurosci. 2012, 15, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Rozenfeld, R. Type I Cannabinoid Receptor Trafficking: All Roads Lead to Lysosome. Traffic 2010, 12, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Augustin, S.M.; Lovinger, D.M. Functional Relevance of Endocannabinoid-Dependent Synaptic Plasticity in the Central Nervous System. ACS Chem. Neurosci. 2018, 9, 2146–2161. [Google Scholar] [CrossRef] [PubMed]
- Soldati, C.; Biagioni, S.; Poiana, G.; Augusti-Tocco, G. β-Catenin and actin reorganization in HGF/SF response of ST14A cells. J. Neurosci. Res. 2008, 86, 1044–1052. [Google Scholar] [CrossRef]
- Lange, C.; Mix, E.; Rateitschak, K.; Rolfs, A. Wnt Signal Pathways and Neural Stem Cell Differentiation. Neurodegener. Dis. 2006, 3, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, J.L.; Ruckle, M.B.; Mayeux, P.R.; Prather, P.L. Agonist-Directed Trafficking of Response by Endocannabinoids Acting at CB2 Receptors. J. Pharmacol. Exp. Ther. 2005, 315, 828–838. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Primers | Annealing T (°C) | |
|---|---|---|
| GAPDH | Fw: 5′-TGGCATTGTGGAAGGGCTCATGAC-3 Rev: 5′-ATGCCAGTGAGCTTCCCGTTCAGC-3′ | 60 |
| CB1 | Fw: 5′-GGGTTACAGCCTCCTTCACA-3′ Rev: 5′-CAGATTGCAGCTTCTTGCAG-3′ | 55 |
| CB2 | Fw: 5′-GGAGTACATGATCTTGAGTGAT-3′ Rev: 5′-AGAACAGGGACTAGGACAAC-3′ | 50 |
| DAGLα | Fw: 5′-GGCAAGACCCTGTAGAGCTG-3′ Rev: 5′-TAAAACAGGTGGCCCTCATC-3′ | 60 |
| MAGL | Fw: 5′-TAGCAGCTGCAGAGAGACCA-3′ Rev: 5′-GATGAGTGGGTCGGAGTTGT-3′ | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cottone, E.; Pomatto, V.; Rapelli, S.; Scandiffio, R.; Mackie, K.; Bovolin, P. Cannabinoid Receptor Modulation of Neurogenesis: ST14A Striatal Neural Progenitor Cells as a Simplified In Vitro Model. Molecules 2021, 26, 1448. https://doi.org/10.3390/molecules26051448
Cottone E, Pomatto V, Rapelli S, Scandiffio R, Mackie K, Bovolin P. Cannabinoid Receptor Modulation of Neurogenesis: ST14A Striatal Neural Progenitor Cells as a Simplified In Vitro Model. Molecules. 2021; 26(5):1448. https://doi.org/10.3390/molecules26051448
Chicago/Turabian StyleCottone, Erika, Valentina Pomatto, Stefania Rapelli, Rosaria Scandiffio, Ken Mackie, and Patrizia Bovolin. 2021. "Cannabinoid Receptor Modulation of Neurogenesis: ST14A Striatal Neural Progenitor Cells as a Simplified In Vitro Model" Molecules 26, no. 5: 1448. https://doi.org/10.3390/molecules26051448
APA StyleCottone, E., Pomatto, V., Rapelli, S., Scandiffio, R., Mackie, K., & Bovolin, P. (2021). Cannabinoid Receptor Modulation of Neurogenesis: ST14A Striatal Neural Progenitor Cells as a Simplified In Vitro Model. Molecules, 26(5), 1448. https://doi.org/10.3390/molecules26051448