
Structural Design, Synthesis and Antioxidant, Antileishmania, Anti-Inflammatory and Anticancer Activities of a Novel Quercetin Acetylated Derivative


 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Synthesis and Characterization
2.2. Antioxidant ABTS•+ Radical Activity
2.3. Antileishmania Activity
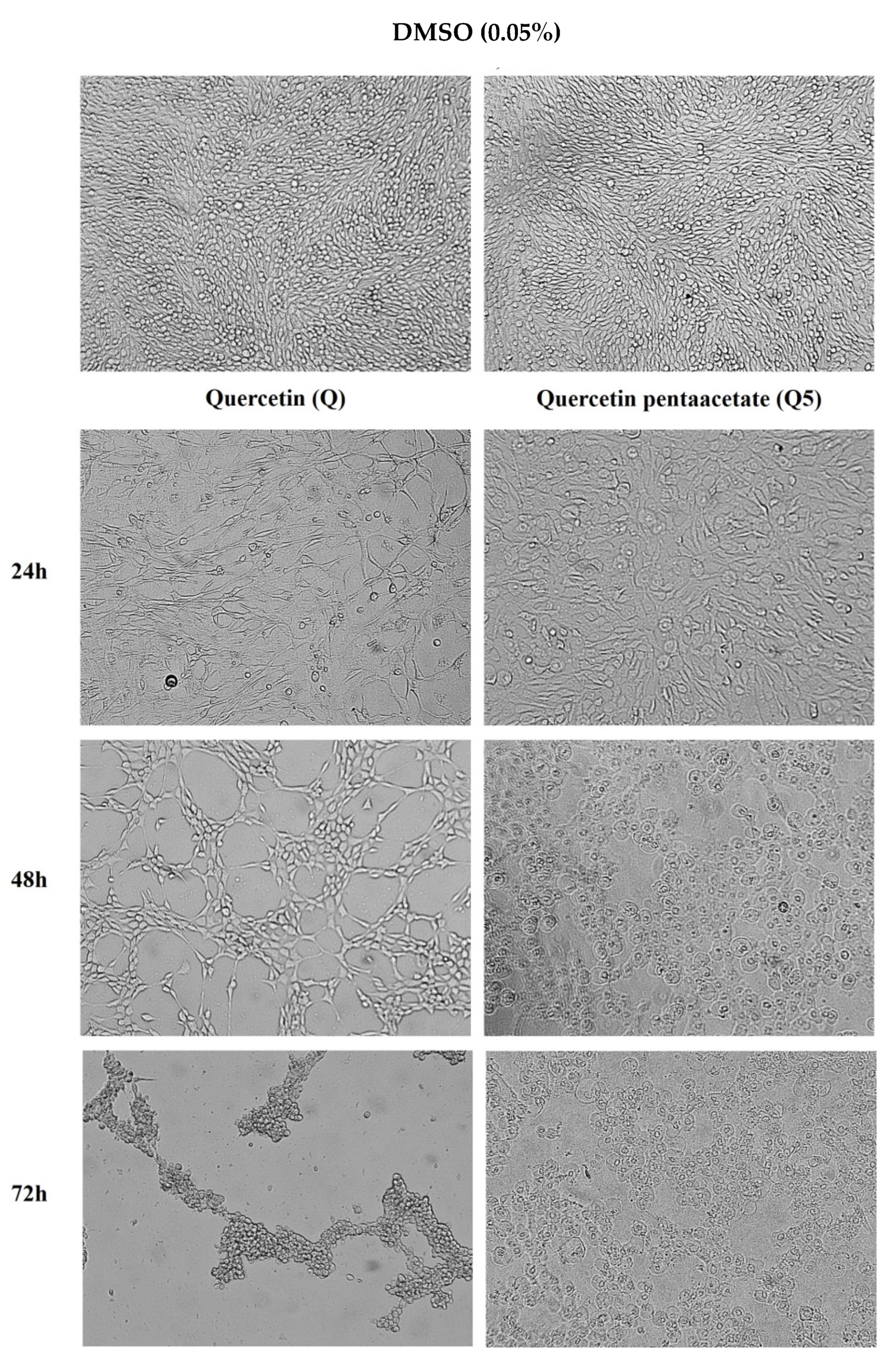
2.4. Anti-Inflammatory and Cytotoxicity Activities
3. Materials and Methods
3.1. Reagents and Materials
3.2. Synthesis and Characterization
3.3. Antioxidant ABTS•+ Radical Activity
3.4. Antileishmania Activity
3.5. Anti-Inflammatory and Cytotoxicity Activities
3.5.1. Drugs
3.5.2. Animals
3.5.3. Cells
3.5.4. Cytotoxic Activity Assay
3.5.5. Macrophage Culture
3.5.6. TNF Dosage
3.5.7. Nitric Oxide Dosage
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Formica, J.V.; Regelson, W. Review of the biology of quercetin and related bioflavonoids. Food Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Materska, M. Quercetin and its derivatives: Chemical structure and bioactivity—A review. Pol. J. Food Nutr. Sci. 2008, 58, 407–413. [Google Scholar]
- Wang, T.Y.; Li, Q.; Bi, K.S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef]
- Araújo, M.V.; Queiroz, A.C.; Silva, J.F.M.; Silva, A.E.; Silva, J.K.S.; Silva, G.R.; Silva, E.C.O.; Souza, S.T.; Fonseca, E.J.S.; Camara, C.A.; et al. Flavonoids induce cell death in: Leishmania amazonensis: In vitro characterization by flow cytometry and Raman spectroscopy. Analyst 2019, 144, 5232–5244. [Google Scholar] [CrossRef] [PubMed]
- Faixová, D.; Hrčková, G.; Kubašková, T.M.; Mudroňová, D. Antiparasitic effects of selected isoflavones on flatworms. Helminthologia 2021, 58, 1–16. [Google Scholar] [CrossRef]
- Manjolin, L.C.; Reis, M.B.G.; Maquiaveli, C.C.; Santos-Filho, O.A.; Silva, E.R. Dietary flavonoids fisetin, luteolin and their derived compounds inhibit arginase, a central enzyme in Leishmania (Leishmania) amazonensis infection. Food Chem. 2013, 141, 2253–2262. [Google Scholar] [CrossRef] [Green Version]
- Tasdemir, D.; Kaiser, M.; Brun, R.; Yardley, V.; Schmidt, T.J.; Tosun, F.; Rüedi1, P. Antitrypanosomal and antileishmanial activities of flavonoids and their analogues: In vitro, in vivo, structure-activity relationship, and quantitative structure-activity relationship studies. Antimicrob. Agents Chemother. 2006, 50, 1352–1364. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.J.; Hsiao, M.; Chang, J.L.; Yang, S.F.; Tseng, T.H.; Cheng, C.W.; Chow, J.M.; Lin, K.H.; Lin, Y.W.; Liu, C.C.; et al. Quercetin induces mitochondrial-derived apoptosis via reactive oxygen species-mediated ERK activation in HL-60 leukemia cells and xenograft. Arch. Toxicol. 2015, 89, 1103–1117. [Google Scholar] [CrossRef]
- Drummond, E.M.; Harbourne, N.; Marete, E.; Martyn, D.; Jacquier, J.; O’Riordan, D.; Gibney, E.R. Inhibition of proinflammatory biomarkers in THP1 macrophages by polyphenols derived from chamomile, meadowsweet and willow bark. Phytother Res. 2013, 27, 588–594. [Google Scholar] [CrossRef]
- Thon, N.; Thorsteinsdottir, J.; Eigenbrod, S.; Schüller, U.; Lutz, J.; Kreth, S.; Belka, C.; Tonn, J.C.; Niyazi, M.; Kreth, F.W. Outcome in unresectable glioblastoma: MGMT promoter methylation makes the difference. J. Neurol. 2016, 264, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Perzelova, A.; Tardy, M. Effects of a novel synthetic retinoid on malignant glioma in vitro inhibition of cell proliferation, induction of apoptosis and differentiation. Eur. J. Cancer 2001, 37, 520–530. [Google Scholar]
- Wang, G.; Wang, J.; Du, L.; Li, F. Effect and mechanism of total flavonoids extracted from Cotinus coggygria against Glioblastoma cancer in vitro and in vivo. Biomed. Res. Int. 2015, 2015, 856349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, B.L.; Oliveira, M.N.; Coelho, P.L.C.; Pitanga, B.P.S.; Silva, A.B.; Adelita, T.; Silva, V.D.A.; Costa, M.F.D.; El-Bachá, R.S.; Tardy, M.; et al. Flavonoids suppress human glioblastoma cell growth by inhibiting cell metabolism, migration, and by regulating extracellular matrix proteins and metalloproteinases expression. Chem.-Biol. Interact. 2015, 242, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, L.; Mohammadi-Motlagh, H.R.; Seyfi, P.; Mostafaie, A. Low concentrations of flavonoid-rich fraction of shallot extract induce delayed-type hypersensitivity and TH1 cytokine IFNγ expression in BALB/c mice. Int. J. Mol. Cell. Med. 2014, 3, 16–25. [Google Scholar]
- Nair, M.P.N.; Kandaswami, C.; Mahajan, S.; Chadha, K.C.; Chawda, R.; Nair, H.; Kumar, N.; Nair, R.E.; Schwartz, S.A. The flavonoid, quercetin, differentially regulates Th-1 (IFNγ) and Th-2 (IL4) cytokine gene expression by normal peripheral blood mononuclear cells. Biochim. Biophys. Acta 2002, 1593, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Massi, A.; Bortolini, O.; Ragno, D.; Bernardi, T.; Sacchetti, G.; Tacchini, M.; De Risi, C. Research progress in the modification of quercetin leading to anticancer agents. Molecules 2017, 22, 1270. [Google Scholar] [CrossRef]
- Foti, M.C.; Rocco, C. Unveiling the chemistry behind bromination of quercetin: The “violet chromogen”. Tetrahedron Lett. 2014, 55, 1602–1607. [Google Scholar] [CrossRef]
- Helgren, T.R.; Sciotti, R.J.; Lee, P.; Duffy, S.; Avery, V.M.; Igbinoba, O.; Akoto, M.; Hagen, T.J. The synthesis, antimalarial activity and CoMFA analysis of novel aminoalkylated quercetin analogs. Bioorg. Med. Chem. Lett. 2015, 25, 327–332. [Google Scholar] [CrossRef]
- Özyürek, M.; Akpınar, D.; Bener, M.; Türkkan, B.; Güçlü, K.; Apak, R. Novel oxime based flavanone, naringin-oxime: Synthesis, characterization and screening for antioxidant activity. Chem.-Biol. Interact. 2014, 212, 40–46. [Google Scholar] [CrossRef]
- Mattarei, A.; Biasutto, L.; Rastrelli, F.; Garbisa, S.; Marotta, E.; Zoratti, M.; Paradisi, C. Regioselective O-derivatization of quercetin via ester intermediates. An improved synthesis of rhamnetin and development of a new mitochondriotropic derivative. Molecules 2010, 15, 4722–4736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danihelová, M.; Veverka, M.; Šturdík, E.; Jantová, S. Antioxidant action and cytotoxicity on HeLa and NIH-3T3 cells of new quercetin derivatives. Interdiscip. Toxicol. 2013, 6, 209–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasutto, L.; Marotta, E.; De Marchi, U.; Zoratti, M.; Paradisi, C. Ester-based precursors to increase the bioavailability of quercetin. J. Med. Chem. 2007, 50, 241–253. [Google Scholar] [CrossRef]
- Mohajeri, M.; Saghaei, L.; Ghanadian, M.; Saberi, S.; Pestechian, N.; Ostadhusseini, E. Synthesis and in vitro leishmanicidal activities of six quercetin derivatives. Adv. Biomed. Res. 2018, 7, 64. [Google Scholar] [PubMed]
- Dehghan, G.; Khoshkam, Z. Tin(II)-quercetin complex: Synthesis, spectral characterisation and antioxidant activity. Food Chem. 2012, 131, 422–426. [Google Scholar] [CrossRef]
- Oh, W.Y.; Ambigaipalan, P.; Shahidi, F. Quercetin and its ester derivatives inhibit oxidation of food, LDL and DNA. Food Chem. 2021, 364, 130394. [Google Scholar] [CrossRef]
- Fonseca-Silva, F.; Inacio, J.D.F.; Canto-Cavalheiro, M.M.; Almeida-Amaral, E.E. Reactive oxygen species production and mitochondrial dysfunction contribute to quercetin induced death in Leishmania amazonensis. PLoS ONE 2011, 8, e14666. [Google Scholar] [CrossRef] [PubMed]
- Cataneo, A.H.D.; Tomiotto-Pellissier, F.; Miranda-Sapla, M.M.; Assolini, J.P.; Panis, C.; Kian, D.; Yamauchi, L.M.; Simão, A.N.C.; Casagrande, R.; Pinge-Filho, P.; et al. Quercetin promotes antipromastigote effect by increasing the ROS production and anti-amastigote by upregulating Nrf2/HO-1 expression, affecting iron availability. Biomed. Pharmacother. 2019, 113, 108745. [Google Scholar] [CrossRef]
- Sen, G.; Mukhopadhyay, S.; Ray, M.; Biswas, T. Quercetin interferes with iron metabolism in Leishmania donovani and targets ribonucleotide reductase to exert leishmanicidal activity. J. Antimicrob. Chemother. 2008, 61, 1066–1075. [Google Scholar] [CrossRef] [Green Version]
- Mittra, B.; Saha, A.; Chowdhury, A.R.; Pal, C.; Mandal, S.; Mukhopadhyay, S.; Bandyopadhyay, S.; Majumder, H.K. Luteolin, an abundant dietary component is a potent anti-leishmanial agent that acts by inducing topoisomerase II-mediated kinetoplast DNA cleavage leading to apoptosis. Mol. Med. 2000, 6, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Suganthy, N.; Devi, K.P.; Nabavi, S.F.; Braidy, N.; Nabavi, S.M. Bioactive effects of quercetin in the central nervous system: Focusing on the mechanisms of actions. Biomed. Pharmacother. 2016, 84, 892–908. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.L.; Silva, V.D.A.; Souza, C.S.; Santos, C.C.; Paris, I.; Muñoz, P.; Segura-Aguilar, J. Impact of plant-derived flavonoids on neurodegenerative diseases. Neurotox. Res. 2016, 30, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzade, A.; Sadeghi, O.; Biregani, A.N.; Soukhtehzari, S.; Brandt, G.S.; Esmaillzadeh, A. Immunomodulatory effects of flavonoids: Possible induction of T CD4+ regulatory cells through suppression of mTOR pathway signaling activity. Front. Immunol. 2019, 10, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Shen, S.C.; Lee, W.R.; Hou, W.C.; Yang, L.L.; Lee, T.J. Inhibition of nitric oxide synthase inhibitors and lipopolysaccharide induced inducible NOS and cyclooxygenase-2 gene expressions by rutin, quercetin, and quercetin pentaacetate in RAW 264.7 macrophages. J. Cell Biochem. 2001, 82, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Sakao, K.; Fujii, M.; Hou, D.X. Clarification of the role of quercetin hydroxyl groups in superoxide generation and cell apoptosis by chemical modification. Biosci. Biotechnol. Biochem. 2009, 73, 2048–2053. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.B.; Coelho, P.L.C.; Amparo, J.A.O.; Carneiro, M.M.A.A.; Borges, J.M.P.; Souza, C.S.; Costa, M.F.D.; Mecha, M.; Rodriguez, C.G.; Silva, V.D.A.; et al. The flavonoid rutin modulates microglial/macrophage activation to a CD150/CD206 M2 phenotype. Chem.-Biol. Interact. 2017, 274, 89–99. [Google Scholar] [CrossRef]
- Kim, M.; Park, Y.; Cho, S.; Burapan, S.; Han, J. Synthesis of alkyl quercetin derivatives. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 343–348. [Google Scholar] [CrossRef]
- Bao, D.; Wang, J.; Pang, X.; Liu, H. Protective effect of quercetin against oxidative stress-induced cytotoxicity in rat pheochromocytoma (PC-12) Cells. Molecules 2017, 22, 1122. [Google Scholar] [CrossRef]
- Bispo da Silva, A.; Coelho, P.L.C.; Oliveira, M.N.; Oliveira, J.L.; Amparo, J.A.O.; Silva, K.C.; Soares, J.R.P.; Pitanga, B.P.S.; Souza, C.S.; Lopes, G.P.F.; et al. The flavonoid rutin and its aglycone quercetin modulate the micróglia inflammatory profile improving antiglioma activity. Brain Behav. Immun. 2020, 85, 170–185. [Google Scholar] [CrossRef]
- Zamin, L.L.; Filippi-Chiela, E.C.; Dillenburg-Pilla, P.; Horn, F.; Salbego, C.; Lenz, G. Resveratrol and quercetin cooperate to induce senescence-like growth arrest in C6 rat glioma cells. Cancer Sci. 2009, 100, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Usman, U.Z.; Ofutet, E.O.; Owu, D.U. Quercetin exerts preventive, ameliorative and prophylactic effects on cadmium chloride–induced oxidative stress in the uterus and ovaries of female Wistar rats. Food Chem. Toxicol. 2017, 102, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Shen, C.; Li, C.; Liu, Y.; Gao, D.; Shi, C.; Peng, F.; Liu, Z.; Zhao, B.; Zheng, Z.; et al. Inhibition of autophagy induced by quercetin at a late stage enhances cytotoxic effects on glioma cells. Tumour Biol. 2016, 37, 3549–3560. [Google Scholar] [CrossRef] [PubMed]
- Dell’Albani, P.; Di Marco, B.; Grasso, S.; Rocco, C.; Foti, M.C. Quercetin derivatives as potent inducers of selective cytotoxicity in glioma cells. Eur. J. Pharm. Sci. 2017, 101, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.J.; Jeng, J.Y.; Lin, C.W.; Wu, C.Y.; Chen, Y.C. Quercetin inhibition of ROS-dependent and -independent apoptosis in rat glioma C6 cells. Toxicology 2006, 223, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Coelho, P.L.C.; Freitas, S.R.V.B.; Pitanga, B.P.S.; Silva, V.D.A.; Oliveira, M.N.; Grangeiro, M.S.; Souza, C.S.; El-Bachá, R.S.; Costa, M.F.D.; Barbosa, P.R.; et al. Flavonoids from the Brazilian plant croton betulaster inhibit the growth of human glioblastoma cells and induce apoptosis. Braz. J. Pharmacog. 2016, 26, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.L. Antiglioma potential of flavonoids. J. Bioanal. Biomed. 2014, 6, 1–3. [Google Scholar]
- Dorman, H.J.D.; Hiltunen, R. Fe(III) reductive and free radical-scavenging properties of summer savory (Satureja hortensis L.) extract and subfractions. Food Chem. 2004, 88, 193–199. [Google Scholar] [CrossRef]
- Guedes, C.E.S.; Dias, B.R.S.; Petersen, A.L.D.O.A.; Cruz, K.P.; Almeida, N.J.; Andrade, D.R.; Menezes, J.P.B.; Borges, V.M.; Veras, P.S.T. In vitro evaluation of the anti-leishmanial activity and toxicity of PK11195. Mem. Inst. Oswaldo Cruz 2018, 113, e170345. [Google Scholar] [CrossRef] [Green Version]
- Benda, P.; Lightbody, J.; Sato, G.; Levine, L.; Sweet, W. Differentiated rat glial cell strain in tissue culture. Science 1968, 161, 370–371. [Google Scholar] [CrossRef]
- Oliveira, D.M.; Lima, R.M.F.; Clarencio, J.; Velozo, E.S.; de Amorim, I.A.; Da Mota, T.H.A.; Costa, S.L.; Silva, F.P.; El-Bachá, R.S. The classical photoactivated drug 8-methoxypsoralen and related compounds are effective without UV light irradiation against glioma cells. Neurochem. Int. 2016, 99, 33–41. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Gogal, R.M.; Walsh, J.E. A new rapid and simple non-radioactive assay to monitor and determine the proliferation of lymphocytes an alternative to [3H] thymidine incorporation assay. J. Immunol. Methods 1994, 170, 211–224. [Google Scholar] [CrossRef]
- Hansen, M.B.; Nielsen, S.E.; Berg, K. Re-examination, and further development of a precise and rapid dye method for measuring cell growth/cell kill. J. Immunol. Methods 1989, 119, 203–210. [Google Scholar] [CrossRef]
- Hora, N.R.S.; Santana, L.F.; Silva, V.D.A.; Costa, S.L.; Zambotti-Villela, L.; Colepicolo, P.; Ferraz, C.G.; Ribeiro, P.R. Identification of bioactive metabolites from corn silk extracts by a combination of metabolite profiling, univariate statistical analysis and chemometrics. Food Chem. 2021, 365, 130479. [Google Scholar] [CrossRef] [PubMed]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15 N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Compounds (IC50 µM) | |||
|---|---|---|---|
| Quercetin (Q) | Q5 | Doxorrubicin | |
| Cancer cells | |||
| IC50 (µM) a HL-60 | 51.3 (±0.4) | 33.6 (±2.6) | 0.2 (±0.0) |
| IC50 (µM) b HepG2 | >80 | 53.9 (±11.3) | 0.1 (±0.0) |
| Non-cancer cells | |||
| CC50 MRC-5 | >80 | >80 | 0.9 (±0.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, S.V.S.; Barboza, O.M.; Souza, J.T.; Soares, É.N.; dos Santos, C.C.; Pacheco, L.V.; Santos, I.P.; Magalhães, T.B.d.S.; Soares, M.B.P.; Guimarães, E.T.; et al. Structural Design, Synthesis and Antioxidant, Antileishmania, Anti-Inflammatory and Anticancer Activities of a Novel Quercetin Acetylated Derivative. Molecules 2021, 26, 6923. https://doi.org/10.3390/molecules26226923
da Silva SVS, Barboza OM, Souza JT, Soares ÉN, dos Santos CC, Pacheco LV, Santos IP, Magalhães TBdS, Soares MBP, Guimarães ET, et al. Structural Design, Synthesis and Antioxidant, Antileishmania, Anti-Inflammatory and Anticancer Activities of a Novel Quercetin Acetylated Derivative. Molecules. 2021; 26(22):6923. https://doi.org/10.3390/molecules26226923
Chicago/Turabian Styleda Silva, Saul Vislei Simões, Orlando Maia Barboza, Jéssica Teles Souza, Érica Novaes Soares, Cleonice Creusa dos Santos, Luciano Vasconcellos Pacheco, Ivanilson Pimenta Santos, Tatiana Barbosa dos Santos Magalhães, Milena Botelho Pereira Soares, Elisalva Teixeira Guimarães, and et al. 2021. "Structural Design, Synthesis and Antioxidant, Antileishmania, Anti-Inflammatory and Anticancer Activities of a Novel Quercetin Acetylated Derivative" Molecules 26, no. 22: 6923. https://doi.org/10.3390/molecules26226923
APA Styleda Silva, S. V. S., Barboza, O. M., Souza, J. T., Soares, É. N., dos Santos, C. C., Pacheco, L. V., Santos, I. P., Magalhães, T. B. d. S., Soares, M. B. P., Guimarães, E. T., Meira, C. S., Costa, S. L., da Silva, V. D. A., de Santana, L. L. B., & de Freitas Santos Júnior, A. (2021). Structural Design, Synthesis and Antioxidant, Antileishmania, Anti-Inflammatory and Anticancer Activities of a Novel Quercetin Acetylated Derivative. Molecules, 26(22), 6923. https://doi.org/10.3390/molecules26226923