
Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods





Abstract
:1. Introduction
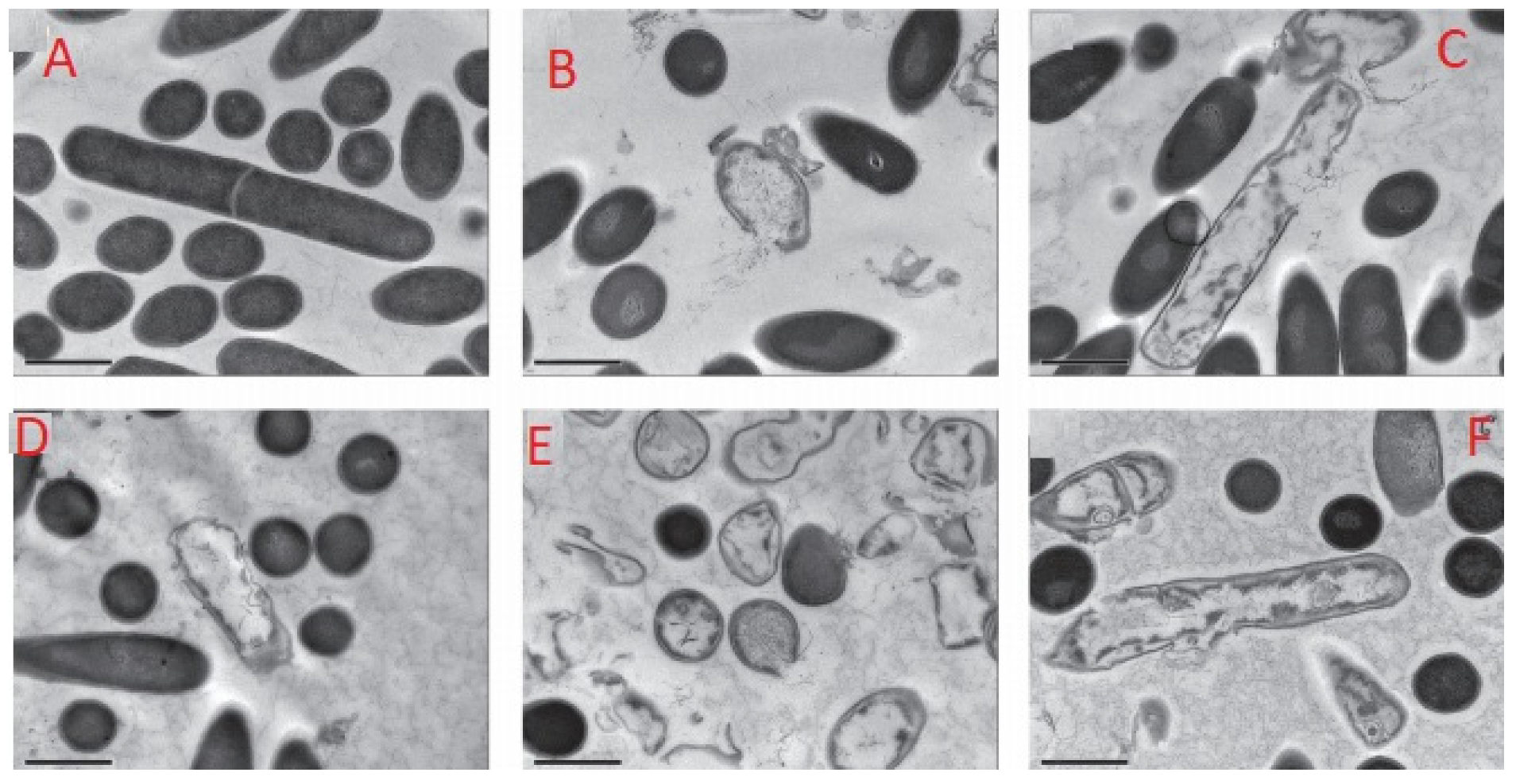
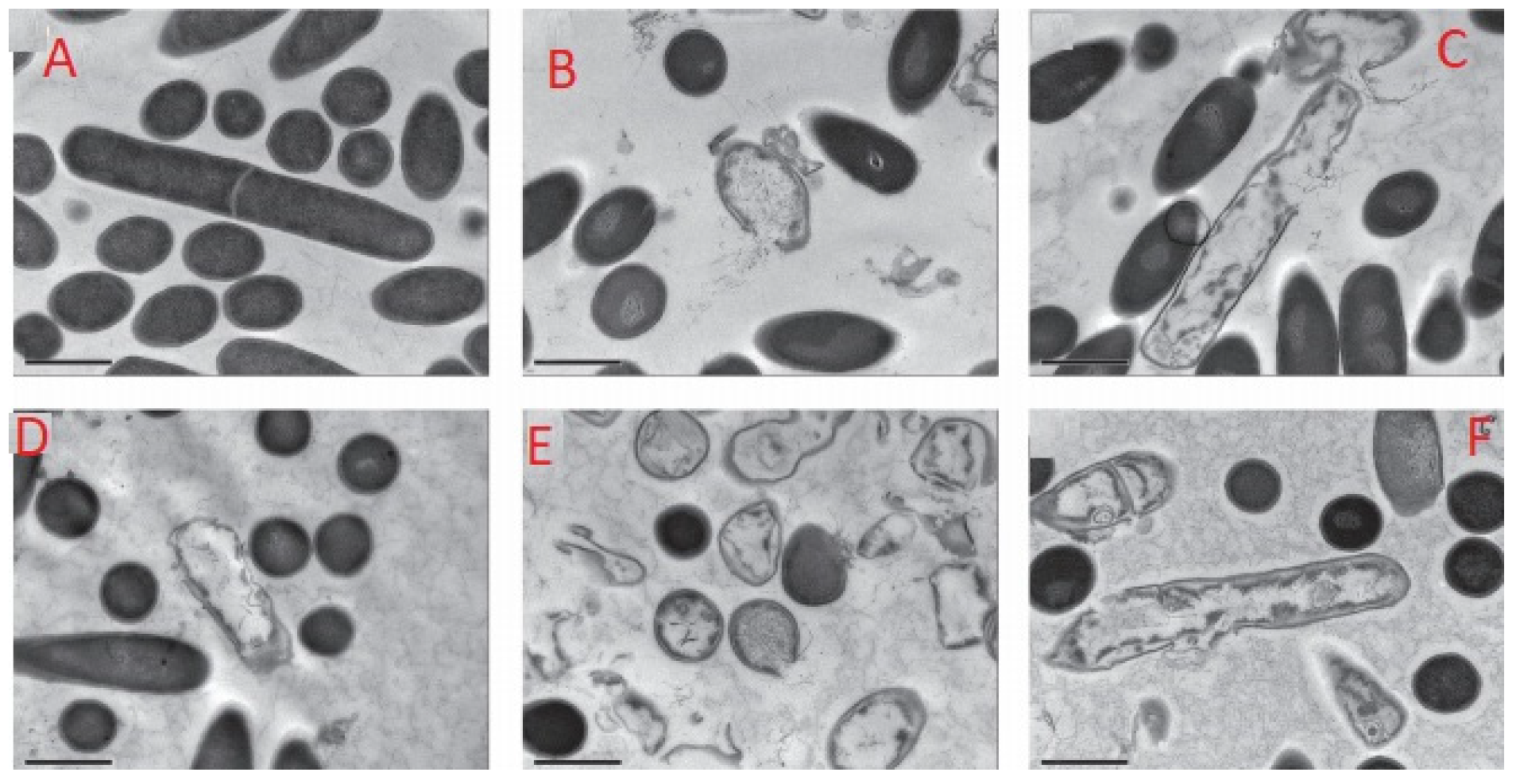
2. Inactivation of Bacteria Spores by AMPs
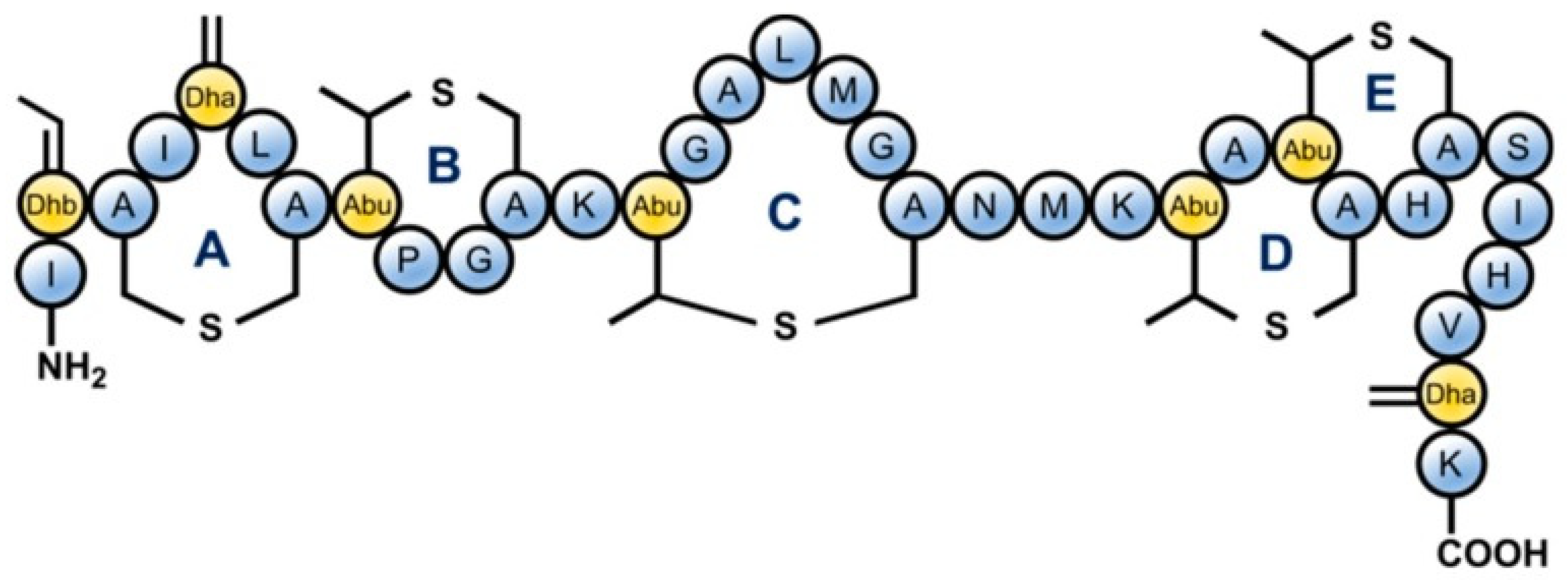
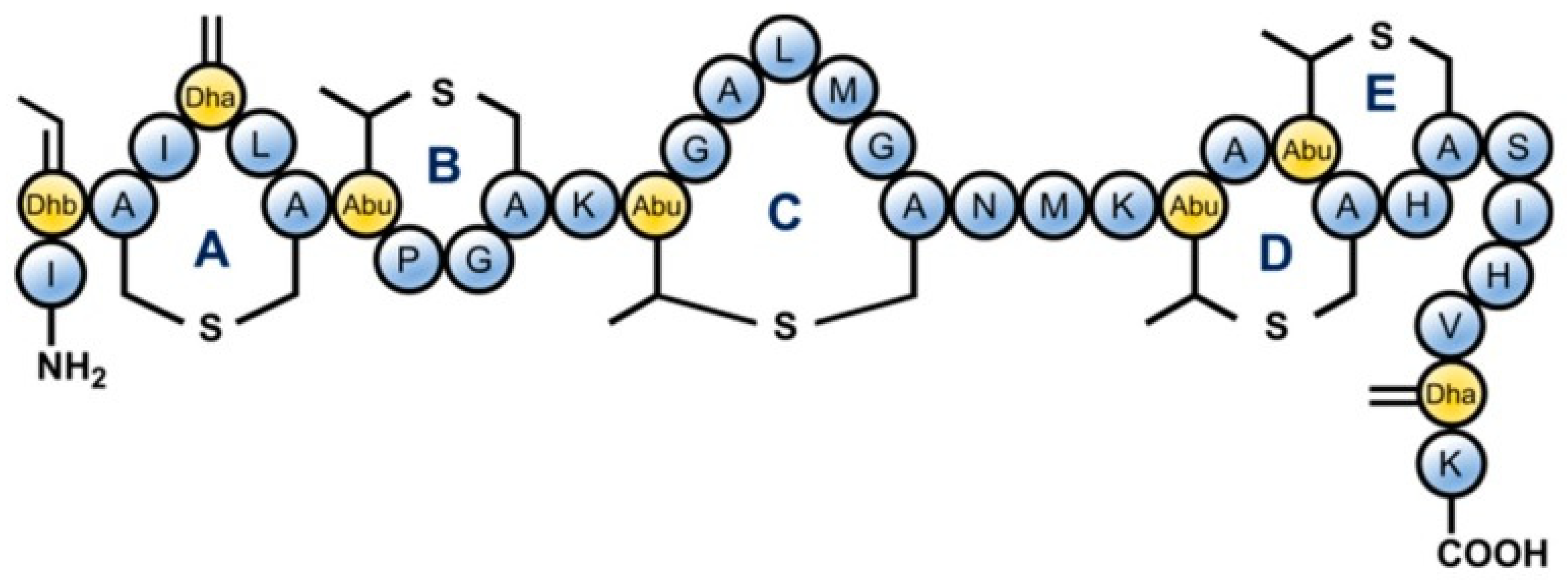
3. Mechanism of Microbial Inactivation by AMP Nisin
4. Impact of AMPs on Food Quality, Advantages and Challenges in Food Processing
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reineke, K.; Mathys, A. Endospore Inactivation by Emerging Technologies: A Review of Target Structures and Inactivation Mechanisms. Annu. Rev. Food Sci. Technol. 2020, 11, 255–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojgani, N.; Amirnia, C. Kinetics of Growth and Bacteriocin Production in L. casei RN 78 Isolated from a Dairy Sample in IR Iran. Int. J. Dairy Sci. 2017, 2, 1–12. [Google Scholar]
- Preciado, G.M.; Michel, M.M.; Villarreal-Morales, S.L.; Flores-Gallgos, A.C.; Aguirre-Joyal, J.; Morlett-Chavez, J.; Aguilar, C.N.; Rodriguez-Herrera, R. Bacteriocins and Its Use for Multidrug-Resistant Bacteria Control. In Antibiotic Resistance; Academic Press: Cambridge, MA, USA, 2016; Chapter 16. [Google Scholar] [CrossRef]
- Chandrakasan, G.; Rodríguez-Hernández, A.I.; Del Rocío López-Cuellar, M.; Palma-Rodrı’guez, H.-M.; Chavarrı’a-Herna’ndez, N. Bacteriocin encapsulation for food and pharmaceutical applications: Advances in the past 20 years. Biotechnol. Lett. 2019, 41, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Pandit, R.; Gaikwad, S.; Ko¨vics, G. Antimicrobial peptides as natural bio-preservative to enhance the shelf-life of food. J. Food Sci. Technol. 2016, 53, 3381–3394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, F.R.; Roque-Borda, A.C.; Vicente, F.E. Antimicrobial peptides as a feed additive alternative to animal production; food safety and public health implications: An overview. Anim. Nutr. 2021. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, H.D.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Alakomi, H.L.; Skytta, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.C.; Lin, C.H.; Sung, C.T.; Fang, J.Y. Antibacterial activities of bacteriocins: Application in foods and pharmaceuticals. Front. Microbiol. 2014, 5, 241. [Google Scholar] [PubMed] [Green Version]
- Jamaluddin, N.; Stuckey, D.C.; Ariff, A.B.; Wong, F.W.F. Novel approaches to purifying bacteriocin: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2453–2465. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.E.; Smid, E.J.; Kok, J.; De Kruijff, B.; Kuipers, O.P.; Breukink, E. Resistance of Gram-positive bacteria to nisin is not determined by lipid II levels. FEMS Microbiol. Lett. 2004, 239, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, N.I.; Breukink, E. The expanding role of lipid II as a target for lantibiotics. Future Microbiol. 2007, 2, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Marriott, B.M. Heat as a Factor in the Perception of Taste, Smell, and Oral Sensation. In Nutritional Needs in Hot Environments: Applications for Military Personnel in Field Operations; National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- Hart, A.; Anumudu, C.; Onyeaka, H.; Miri, T. Application of supercritical fluid carbon dioxide in improving food shelf-life and safety by inactivating spores: A review. J. Food Sci. Technol. 2021, 1–12. [Google Scholar] [CrossRef]
- Kaskoniene, V.; Stankevicius, M.; Bimbiraite-Surviliene, K.; Naujokaityte, G.; Serniene, L.; Mulkyte, K.; Malakauskas, M.; Maruska, A. Current state of purification; isolation and analysis of bacteriocins produced by lactic acid bacteria. Appl. Microbiol. Biotechnol. 2017, 101, 1323–1335. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalban-Lopez, M.; Mu, D.D.; Kuipers, O.P. Bacteriocins of lactic acid bacteria: Extending the family. Appl Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [Green Version]
- Ogunbanwo, S.T.; Sanni, A.I.; Onilude, A.A. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol. 2003, 2, 219–227. [Google Scholar]
- Ghrairi, T.; Chaftar, N.; Hani, K. Bacteriocins: Recent Advances and Opportunities. Prog. Food Preserv. 2012, 485–511. [Google Scholar] [CrossRef]
- Rao, L.; Wang, Y.; Chen, F.; Liao, X. The Synergistic Effect of High Pressure CO2 and Nisin on Inactivation of Bacillus subtilis Spores in Aqueous Solutions. Front. Microbiol. 2016, 7, 1507. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xu, Z.; Zhao, F.; Wang, Y.; Liao, X. Synergetic effects of high- pressure carbon dioxide and nisin on the inactivation of Escherichia coli and Staphylococcus aureus. Innov. Food Sci. Emerg Technol. 2016, 33, 180–186. [Google Scholar] [CrossRef]
- Bolt, H.L.; Kleijn, L.H.; Martin, N.I.; Cobb, S.L. Synthesis of Antibacterial Nisin–Peptoid Hybrids Using Click Methodology. Molecules 2018, 23, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudêncio, C.V.; Dos Santos, M.T.; Vanetti, M.C.D. Strategies for the use of bacteriocins in Gram-negative bacteria: Relevance in food microbiology. J. Food Sci. Technol. 2015, 52, 5408–5417. [Google Scholar] [CrossRef] [Green Version]
- Mills, S.; Serrano, L.M.; Griffin, C.; O’Connor, P.M.; Schaad, G.; Bruining, C.; Hill, C.; Ross, R.P.; Meijer, W.C. Inhibitory activity of Lactobacillus plantarum LMG P-26358 against Listeria innocua when used as an adjunct starter in the manufacture of cheese. Microb. Cell Fact. 2011, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Gough, R.; O’Connor, M.P.; Rea, C.M.; Gomez-Sala, B.; Miao, S.; Hill, C.; Brodkorb, A. Simulated gastrointestinal digestion of nisin and interaction between nisin and bile. LWT 2017, 86, 530–537. [Google Scholar] [CrossRef] [Green Version]
- Belgacem, Z.B.; Rehaiem, A.; Bernárdez, P.F.; Manai, M.; Castro, L.P. Interactive effects of pH and temperature on the bacteriocin stability by response surface analysis. Microbiology 2012, 81, 195–200. [Google Scholar] [CrossRef]
- Julien, M.C.; Dion, P.; Lafreniere, C.; Antoun, H.; Drouin, P. Sources of clostridia in raw milk on farms. Appl. Environ. Microbiol. 2008, 74, 6348–6357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.R.; Moss, M.O. Food Microbiology; RSC Publishing: Cambridge, UK, 2008. [Google Scholar]
- Vicente, E.F.; Basso, L.G.M.; Cespedes, G.F.; Lorenzon, E.N.; Castro, M.S.; Mendes-Giannini, M.J.S.; Cilli, E.M. Dynamics and conformational studies of TOAC spin labeled analogues of Ctx(Ile21)-Ha peptide from Hypsiboas albopunctatus. PLoS ONE 2013, 8, e60818. [Google Scholar] [CrossRef] [PubMed]
- Oman, T.J.; Van der Donk, W.A. Insights into the mode of action of the two-peptide lantibiotic haloduracin. ACS Chem. Biol. 2009, 4, 865–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Cuesta, M.C.; Bengoechea, J.; Bustos, I.; Rodríguez, B.; Requena, T.; Pelaez, C. Control of late blowing in cheese by adding lacticin 3147-producing Lactococcus lactis IFPL 3593 to the starter. Int. Dairy J. 2010, 20, 18–24. [Google Scholar] [CrossRef]
- Bogovic Matijasic, B.; Koman Rajsp, M.; Perko, B.; Rogelj, I. Inhibition of Clostridium tyrobutyricum in cheese by Lactobacillus gasseri. Int. Dairy J. 2007, 17, 157–166. [Google Scholar] [CrossRef]
- Garde, S.; Rodríguez, E.; Gaya, P.; Medina, M.; Nuñez, M. PCR detection of the structural genes of nisin Z and lacticin 481 in Lactococcus lactis subsp. lactis INIA 415; a strain isolated from raw milk Manchego cheese. Biotechnol. Lett. 2001, 23, 85–89. [Google Scholar] [CrossRef]
- Garde, S.; Carbonell, M.; Fernández-García, E.; Medina, M.; Nuñez, M. Volatile compounds in Hispánico cheese manufactured using a mesophilic starter; a thermophilic starter and bacteriocin-producing Lactococcus lactis subsp. lactis INIA 415. J. Agric. Food Chem. 2002, 50, 6752–6757. [Google Scholar] [CrossRef] [PubMed]
- Garde, S.; Avila, M.; Arias, R.; Gaya, P.; Nunez, M. Outgrowth inhibition of Clostridium beijerinckii spores by a bacteriocin-producing lactic culture in ovine milk cheese. Int. J. Food Microbiol. 2011, 150, 59–65. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Vandamme, E.J. Nisin; A Lantibiotic Produced by Lactococcus Lactis Subsp. Lactis: Properties; Biosynthesis; Fermentation and Applications. In Bacteriocins of Lactic Acid Bacteria; De Vuyst, L., Vandamme, E.J., Eds.; Blackie Academic & Professional: London, UK, 1994; pp. 151–221. [Google Scholar]
- Maria-Neto, S.; De Almeida, K.C.; Macedo, M.L.R.; Franco, O.L. Understanding bacterial resistance to antimicrobial peptides: From the surface to deep inside. Biochim. Biophys. Acta 2015, 1848, 3078–3088. [Google Scholar] [CrossRef] [Green Version]
- Hofstetter, S.; Gebhardt, D.; Ho, L.; Ganzle, M.; McMullen, M. Effect of nisin and reutericyclin on resistanceof endospores of Clostridium spp. to heat and high pressure. Food Microbiol. 2013, 34, 46–51. [Google Scholar] [CrossRef]
- Nerandzic, M.M.; Donskey, C.J. Activate to Eradicate: Inhibition of Clostridium difficile Spore Outgrowth by the Synergistic Effects of Osmotic Activation and Nisin. PLoS ONE 2013, 8, e54740. [Google Scholar] [CrossRef] [Green Version]
- Aouadhi, C.; Mejri, S.; Maaroufi, A. Inhibitory effects of nisin and potassium sorbate alone or in combination on vegetative cells growth and spore germination of Bacillus sporothermodurans in milk. Food Microbiol. 2015, 46, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Abriouel, H.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Valdivia, E. Inhibition of Bacterial Growth; Enterotoxin Production; and Spore Outgrowth in Strains of Bacillus cereus by Bacteriocin AS-48. Appl. Environ. Microbiol. 2002, 68, 1473–1477. [Google Scholar] [CrossRef] [Green Version]
- Lucas, R.; Grande, M.J.; Abriouel, H.; Maqueda, M.; Omar, N.B.; Valdivia, E.; Gálvez, A. Application of the broad-spectrum bacteriocin enterocin AS-48 to inhibit Bacillus coagulans in canned fruit and vegetable foods. Food. Chem Toxicol. 2006, 44, 1774–1781. [Google Scholar] [CrossRef]
- Pei, J.; Yue, T.; Jin, W. Application of bacteriocin RC20975 in apple juice. Food Sci. Technol. Int. 2016, 23, 166–173. [Google Scholar] [CrossRef]
- González, L.; Zárate, V. Inhibitory activity of Lactobacillus plantarum TF711 against Clostridium sporogenes when used as adjunct culture in cheese manufacture. J. Dairy Res. 2015, 82, 236–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Manns, D.C.; Guron, G.K.; Churey, J.J.; Worobo, R.W. Large-scale purification; characterization; and spore outgrowth inhibitory effect of Thurincin H, a bacteriocin produced by Bacillus thuringiensis SF361. Probiotics Antimicrob. Proteins 2014, 6, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Fang, J.; Tian, Y.; Lu, Y.X. Mechanisms of nisin resistance in Gram-positive bacteria. Ann. Microbiol. 2014, 64, 413–420. [Google Scholar] [CrossRef]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, 216–230. [Google Scholar] [CrossRef]
- Barbosa, A.A.T.; De Melo, M.R.; Da Silva, C.M.R.; Jain, S.; Dolabella, S.S. Nisin resistance in Gram-positive bacteria and approaches to circumvent resistance for successful therapeutic use. Crit. Rev. Microbiol. 2021, 47, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2012, 11, 95–105. [Google Scholar] [CrossRef]
- Aouadhi, C.; Simonin, H.; Mejri, S.; Maaroufi, A. The combined effect of nisin; moderate heating and high hydrostatic pressure on the inactivation of Bacillus sporothermodurans spores. J. Appl. Microbiol. 2013, 115, 147–155. [Google Scholar] [CrossRef]
- Aouadhi, C.; Rouissi, Z.; Mejri, S.; Maarouf, A. Inactivation of Bacillus sporothermodurans spores by nisin and temperature studied by design of experiments in water and milk. Food Microbiol. 2014, 38, 270–275. [Google Scholar] [CrossRef]
- Udompijitkul, P.; Paredes-Sabja, D.; Sarker, M.R. (2012). Inhibitory effects of nisin against Clostridium perfringens food poisoning and nonfood-borne isolates. J. Food Sci. 2012, 77, M51–M56. [Google Scholar] [CrossRef]
- Gut, I.M.; Blanke, S.R.; Van der Donk, W.A. Mechanism of inhibition of Bacillus anthracis spore outgrowth by the lantibiotic nisin. ACS Chem. Biol. 2011, 6, 744–752. [Google Scholar] [CrossRef]
- Le Lay, C.; Dridi, L.; Bergeron, M.G.; Ouellette, M.; Fliss, I. Nisin is an effective inhibitor of Clostridium difficile vegetative cells and spore germination. J. Med. Microbiol. 2016, 65, 169–175. [Google Scholar] [CrossRef]
- Setlow, B.; Atluri, S.; Kitchel, R.; Koziol-Dube, K.; Setlow, P. Role of dipicolinic acid in resistance and stability of spores of Bacillus subtilis with or without DNA-protective α/β-type small acid-soluble proteins. J. Bacteriol. 2006, 188, 3740–3747. [Google Scholar] [CrossRef] [Green Version]
- Pol, I.E.; Van Arendonk, W.G.C.; Mastwijk, H.C.; Krommer, J.; Smid, E.J.; Moezelaar, R. Sensitivities of Germinating Spores and Carvacrol-Adapted Vegetative Cells and Spores of Bacillus cereus to Nisin and Pulsed-Electric-Field Treatment. Appl. Environ. Microbiol. 2001, 67, 1693–1699. [Google Scholar] [CrossRef] [Green Version]
- Chai, C.; Lee, K.-S.; Oh, S.-W. Synergistic inhibition of Clostridium difficile with nisin-lysozyme combination treatment. Anaerobe 2015, 34, 24–26. [Google Scholar] [CrossRef]
- Chai, C.; Lee, K.S.; Imm, G.S.; Kim, Y.S.; Oh, S.W. Inactivation of Clostridium difficile spore outgrowth by synergistic effects of nisin and lysozyme. Can. J. Microbiol. 2017, 63, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Galvagno, M.A.; Gil, G.R.; Iannone, L.J.; Cerrutti, P. Exploring the use of natural antimicrobial agents and pulsed electric fields to control spoilage bacteria during a beer production process. Rev. Argent Microbiol. 2007, 39, 170–176. [Google Scholar] [PubMed]
- Grande, M.J.; Lucas, R.; Abriouel, H.; Valdivia, E.; Omar, N.B.; Maqueda, M.; Gálvez, A. Inhibition of toxicogenic Bacillus cereus in rice-based foods by enterocin AS-48. Int. J. Food Microbiol. 2006, 106, 185–194. [Google Scholar] [CrossRef]
- Lo’pez-Pedemonte, J.T.; Roig-Sague’s, X.A.; Trujillo, J.A.; Capellas, M.; Guamis, B. Inactivation of Spores of Bacillus cereus in Cheese by High Hydrostatic Pressure with the Addition of Nisin or Lysozyme. J. Dairy Sci. 2003, 86, 3075–3081. [Google Scholar] [CrossRef] [Green Version]
- Kmiha, S.; Modugno, C.; Aouadhi, C.; Simonin, H.; Mejri, S.; Perrier-Cornet, J.; Maaroufi, A. Inhibitory effect of high hydrostatic pressure; nisin; and moderate heating on the inactivation of Paenibacillus sp. and Terribacillus aidingensis spores isolated from UHT milk. High Press. Res. 2021, 1–13. [Google Scholar] [CrossRef]
- Modugno, C.; Kmiha, S.; Simonin, H.; Aouadhi, C.; Cañizares, E.D.; Lang, E.; André, S.; Mejri, S.; Maaroufi, A.; Perrier-Cornet, J.M. High pressure sensitization of heat-resistant and pathogenic foodborne spores to nisin. Food Microbiol. 2019, 84, 103244. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Ismail, B.B.; Hou, F.; Muhammad, A.I.; Ding, T.; Liu, D. Ultrasound pretreatment enhances the inhibitory effects of nisin/carvacrol against germination; outgrowth and vegetative growth of spores of Bacillus subtilis ATCC6633 in laboratory medium and milk: Population and single-cell analysis. Int. J. Food Microbiol. 2019, 311, 108329. [Google Scholar] [CrossRef]
- Gao, Y.; Qiu, W.; Wu, D.; Fu, Q. Assessment of Clostridium perfringens Spore Response to High Hydrostatic Pressure and Heat with Nisin. Appl. Biochem. Biotechnol. 2011, 164, 1083–1095. [Google Scholar] [CrossRef]
- Rodgers, S.; Peiris, P.; Casadei, G. Inhibition of Nonproteolytic Clostridium botulinum with Lactic Acid Bacteria and Their Bacteriocins at Refrigeration Temperatures. J. Food Prot. 2003, 66, 674–678. [Google Scholar] [CrossRef]
- Gut, I.M.; Prouty, A.M.; Ballard, J.D.; Van der Donk, W.A.; Blanke, S.R. Inhibition of Bacillus anthracis Spore Outgrowth by Nisin. Antimicrob. Agents Chemother. 2008, 52, 4281–4288. [Google Scholar] [CrossRef] [Green Version]
- Naim, F.; Zareifard, M.R.; Zhu, S.; Huizing, R.H.; Grabowski, S.; Marcotte, M. Combined effects of heat; nisin and acidification on the inactivation of Clostridium sporogenes spores in carrot-alginate particles: From kinetics to process validation. Food Microbiol. 2008, 25, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-L.; Ju, X.-R. Exploiting the combined effects of high pressure and moderate heat with nisin on inactivation of Clostridium botulinum spores. J. Microbiol. Methods 2008, 72, 20–28. [Google Scholar] [CrossRef]
- Ros-Chumillas, M.; Esteban, M.-D.; Huertas, J.-P.; Palop, A. Effect of Nisin and Thermal Treatments on the Heat Resistance of Clostridium sporogenes Spores. J. Food Prot. 2015, 78, 2019–2023. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Novel bacteriocins from lactic acid bacteria (LAB): Various structures and applications. Microb. Cell Fact. 2014, 13, S3. [Google Scholar] [CrossRef] [Green Version]
- Leroy, F.; De Vuyst, L. Bacteriocins of lactic acid bacteria to combat undesirable bacteria in dairy products. Aust. J. Dairy Technol. 2010, 65, 143–149. [Google Scholar]
- Oumer, A.; Garde, S.; Gaya, P.; Medina, M.; Nuñez, M. The effects of cultivating lactic starter cultures with bacteriocin-producing lactic acid bacteria. J. Food Prot. 2001, 64, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Oshima, S.; Hirano, A.; Kamikado, H.; Nishimura, J.; Kawai, Y.; Saito, T. Nisin A extends the shelf life of high-fat chilled dairy dessert; a milk-based pudding. J. Appl. Microbiol. 2014, 116, 1218–1228. [Google Scholar] [CrossRef]
- Thomas, L.V.; Ingram, R.E.; Bevis, H.E.; Davies, E.A.; Milne, C.F.; Delves-Broughton, J. Effective Use of Nisin to Control Bacillus and Clostridium Spoilage of a Pasteurized Mashed Potato Product. J. Food Prot. 2002, 65, 1580–1585. [Google Scholar] [CrossRef]
- Da Costa, R.J.; Voloski, F.L.; Mondadori, R.G.; Duval, E.H.; Fiorentini, Â.M. Preservation of meat products with bacteriocins produced by lactic acid bacteria isolated from meat. J. Food Qual. 2019, 4726510. [Google Scholar] [CrossRef] [Green Version]
- Wandling, L.R.; Sheldon, B.W.; Foegeding, P.M. Nisin in Milk Sensitizes Bacillus Spores to Heat and Prevents Recovery of Survivors. J. Food Prot. 1999, 62, 492–498. [Google Scholar] [CrossRef]
- Cruz, J.; Montville, T.J. Influence of Nisin on the Resistance of Bacillus anthracis Sterne Spores to Heat and Hydrostatic Pressure. J. Food Prot. 2008, 71, 196–199. [Google Scholar] [CrossRef]
- Mansour, M.; Amri, D.; Bouttefroy, A.; Linder, M.; Milliere, J.B. Inhibition of Bacillus licheniformis spore growth in milk by nisin; monolaurin; and pH combinations. J. Appl. Microbiol. 1999, 86, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.L., Jr.; Sniff, E.E. Effect of Subtilin and Nisin on the Spores of Bacillus coagulans; Pullman, P., Ed.; Scientific Paper No. 1795; Project Nos. 1222 and 1290; Washington Agricultural Experiment Stations: Washington, DC, USA, 1959; Volume 77, pp. 766–770. [Google Scholar]

{kind=link}
{kind=link}
| AMPs | Spore Former | References |
|---|---|---|
| Nisin | C. perfringens, C. sporogenes, C. botulinum, C. difficile, C. beijerinckii | [38,39,40] |
| Enterocin | A. acidoterrestris, B. cereus, B. licheniformis, G. stearothermophilus | [41,42] |
| Bificin | A. acidoterrestris | [43] |
| Lacticin | C. tyrobutyricum | [31] |
| Plantaricin | C. sporogenes | [44] |
| Thurincin | B. cereus | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anumudu, C.; Hart, A.; Miri, T.; Onyeaka, H. Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods. Molecules 2021, 26, 5552. https://doi.org/10.3390/molecules26185552
Anumudu C, Hart A, Miri T, Onyeaka H. Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods. Molecules. 2021; 26(18):5552. https://doi.org/10.3390/molecules26185552
Chicago/Turabian StyleAnumudu, Christian, Abarasi Hart, Taghi Miri, and Helen Onyeaka. 2021. "Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods" Molecules 26, no. 18: 5552. https://doi.org/10.3390/molecules26185552
APA StyleAnumudu, C., Hart, A., Miri, T., & Onyeaka, H. (2021). Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods. Molecules, 26(18), 5552. https://doi.org/10.3390/molecules26185552