
Lipidomics of the Edible Brown Alga Wakame (Undaria pinnatifida) by Liquid Chromatography Coupled to Electrospray Ionization and Tandem Mass Spectrometry

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
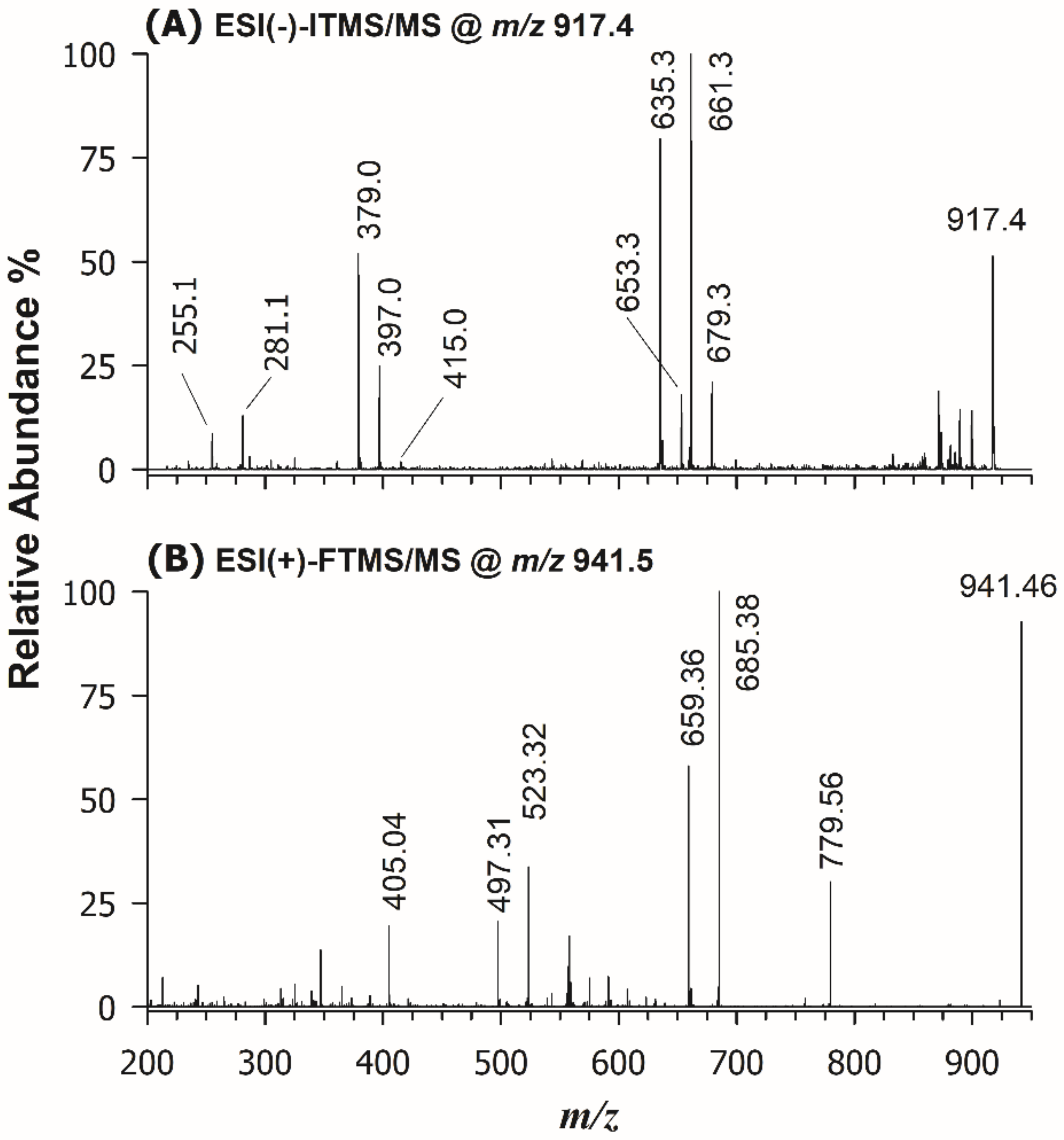
2. Results and Discussion
2.1. Lipidomics of U. pinnatifida by HILIC-ESI(-)-FTMS
2.2. SQDG and SQMG in the Lipid Extract of U. pinnatifida by RPLC-ESI(-)-MS and Tandem MS Analysis
2.3. HILIC-ESI(-)-MS(/MS) and DI-ESI(+)-MS/MS Analysis of DGDG and DGMG in the Lipid Extract of U. pinnatifida
2.4. PL and lyso-PL in the Lipid Extract of U. pinnatifida by HILIC-ESI(-)-MS(/MS) Analysis
2.5. Relative Abundance of PL and GL in the Lipid Extract of U. pinnatifida
3. Materials and Methods
3.1. Chemicals and Samples
3.2. LC-MS Instrumentation and Operating Conditions
3.3. Extraction of Lipids
3.4. Solid-Phase Extraction of Lipid Extracts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Llorénsc, L.J. World cuisine of seaweeds: Science meets gastronomy. Int. J. Gastron. Food Sci. 2018, 14, 55–65. [Google Scholar] [CrossRef]
- Taboada, M.C.; Millán, R.; Miguez, M.I. Nutritional value of the marine algae wakame (Undaria pinnatifida) and nori (Porphyra purpurea) as food supplements. J. Appl. Phycol. 2013, 25, 1271–1276. [Google Scholar] [CrossRef]
- Terasaki, M.; Kubota, A.; Kojima, H.; Maeda, H.; Miyashita, K.; Kawagoe, C.; Mutoh, M.; Tanaka, T. Fucoxanthin and Colorectal Cancer Prevention. Cancers 2021, 13, 2379. [Google Scholar] [CrossRef] [PubMed]
- Burtin, P. Nutritional value of seaweeds. Electron. J. Environ. Agric. Food Chem. 2003, 2, 498–503. [Google Scholar]
- Watanabe, F.; Yabuta, Y.; Bito, T.; Teng, F. Vitamin B12-containing plant food sources for vegetarians. Nutrients 2014, 6, 1861–1873. [Google Scholar] [CrossRef]
- Martínez–Hernández, G.B.; Castillejo, N.; Carrión–Monteagudo, M.d.M.; Artés, F.; Artés-Hernández, F. Nutritional and bioactive compounds of commercialized algae powders used as food supplements. Food Sci. Technol. Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.C.; Gallagher, E.; Tasdemir, D.; Hayes, M. Heart Health Peptides from Macroalgae and Their Potential Use in Functional Foods. J. Agric. Food Chem. 2011, 59, 6829–6836. [Google Scholar] [CrossRef]
- Pereira, L. Seaweeds as Source of Bioactive Substances and Skin Care Therapy-cosmeceuticals, Algotheraphy and Thalassotherapy. Cosmetics 2018, 5, 68. [Google Scholar] [CrossRef]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from Marine Macroalgae for Human and Animal Health Applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar] [CrossRef]
- Smit, A.J. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Tascon, M.; Gagliardi, L.G.; Benavente, F. Parts-per-trillion detection of harmala alkaloids in Undaria pinnatifida algae by on-line solid phase extraction capillary electrophoresis mass spectrometry. Anal. Chim. Acta 2017, 954, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Generalić Mekinić, I.; Skroza, D.; Šimat, V.; Hamed, I.; Čagalj, M.; Popović Perković, Z. Phenolic Content of Brown Algae (Pheophyceae) Species: Extraction, Identification, and Quantification. Biomolecules 2019, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Badocco, D.; Di Marco, V.; Piovan, A.; Caniato, R.; Pastore, P. A procedure for the quantification of total iodine by inductively coupled plasma mass spectrometry, and its application to the determination of iodine in algae sampled in the lagoon of Venice. Anal. Methods 2016, 8, 7545–7551. [Google Scholar] [CrossRef]
- Zhao, Y.-F.; Wu, J.-F.; Shang, D.-R.; Ning, J.-S.; Ding, H.-Y.; Zhai, Y.-X. Arsenic Species in Edible Seaweeds Using In Vitro Biomimetic Digestion Determined by High-Performance Liquid Chromatography Inductively Coupled Plasma Mass Spectrometry. Int. J. Food Sci. 2014, 2014, 436347. [Google Scholar] [CrossRef]
- García-Salgado, S.; Raber, G.; Raml, R.; Magnes, C.; Francesconi, K.A. Arsenosugar phospholipids and arsenic hydrocarbons in two species of brown macroalgae. Environ. Chem. 2012, 9, 63–66. [Google Scholar] [CrossRef]
- Viczek, S.A.; Jensen, K.B.; Francesconi, K.A. Arsenic-Containing Phosphatidylcholines: A New Group of Arsenolipids Discovered in Herring Caviar. Angew. Chem. 2016, 128, 5345–5348. [Google Scholar] [CrossRef]
- Coniglio, D.; Calvano, C.D.; Ventura, G.; Losito, I.; Cataldi, T.R.I.I. Arsenosugar Phospholipids (As-PL) in Edible Marine Algae: An Interplay between Liquid Chromatography with Electrospray Ionization Multistage Mass Spectrometry and Phospholipases A 1 and A 2 for Regiochemical Assignment. J. Am. Soc. Mass Spectrom. 2020, 31, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Francesconi, K.A. Complete extraction of arsenic species: A worthwhile goal? Appl. Organomet. Chem. 2003, 17, 682–683. [Google Scholar] [CrossRef]
- Francesconi, K.A. Arsenic species in seafood: Origin and human health implications. Pure Appl. Chem. 2010, 82, 373–381. [Google Scholar] [CrossRef]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Schlotterbeck, J.; Kolb, A.; Lämmerhofer, M. Free fatty acid profiling in marine algae extract by LC-MS/MS and isolation as well as quantification of the ω-3 fatty acid hexadeca-4,7,10,13-tetraenoic acid. J. Sep. Sci. 2018, 41, 4286–4295. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Rozentsvet, O.A. Phospholipid composition of some marine red algae. Phytochemistry 1990, 29, 3149–3152. [Google Scholar] [CrossRef]
- Vyssotski, M.; Lagutin, K.; MacKenzie, A.; Mitchell, K.; Scott, D. Phospholipids of New Zealand Edible Brown Algae. Lipids 2017, 52, 629–639. [Google Scholar] [CrossRef]
- Řezanka, T.; Lukavský, J.; Vítová, M.; Nedbalová, L.; Sigler, K. Lipidomic analysis of Botryococcus (Trebouxiophyceae, Chlorophyta)—Identification of lipid classes containing very long chain fatty acids by offline two-dimensional LC-tandem MS. Phytochemistry 2018, 148, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Peralta-García, E.; Caamal-Fuentes, E.; Robledo, D.; Hernández-Núñez, E.; Freile-Pelegrín, Y. Lipid characterization of red alga Rhodymenia pseudopalmata (Rhodymeniales, Rhodophyta). Phycol. Res. 2017, 65, 58–68. [Google Scholar] [CrossRef]
- Murphy, R.C. Tandem Mass Spectrometry of Lipids; Royal Society of Chemistry: London, UK, 2015. [Google Scholar]
- Buszewski, B.; Noga, S. Hydrophilic interaction liquid chromatography (HILIC)—A powerful separation technique. Anal. Bioanal. Chem. 2012, 402, 231–247. [Google Scholar] [CrossRef]
- Cajka, T.; Fiehn, O. Comprehensive analysis of lipids in biological systems by liquid chromatography-mass spectrometry. Trends Anal. Chem. 2014, 61, 192–206. [Google Scholar] [CrossRef]
- Granafei, S.; Azzone, P.; Spinelli, V.A.; Losito, I.; Palmisano, F.; Cataldi, T.R.I. Hydrophilic interaction and reversed phase mixed-mode liquid chromatography coupled to high resolution tandem mass spectrometry for polar lipids analysis. J. Chromatogr. A 2016, 1477, 47–55. [Google Scholar] [CrossRef]
- Zianni, R.; Bianco, G.; Lelario, F.; Losito, I.; Palmisano, F.; Cataldi, T.R.I. Fatty acid neutral losses observed in tandem mass spectrometry with collision-induced dissociation allows regiochemical assignment of sulfoquinovosyl-diacylglycerols. J. Mass Spectrom. 2013, 48, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Granafei, S.; Losito, I.; Palmisano, F.; Cataldi, T.R.I. Unambiguous regiochemical assignment of sulfoquinovosyl mono- and diacylglycerols in parsley and spinach leaves by liquid chromatography/electrospray ionization sequential mass spectrometry assisted by regioselective enzymatic hydrolysis. Rapid Commun. Mass Spectrom. 2017, 31, 1499–1509. [Google Scholar] [CrossRef]
- Han, X. Lipidomics-Comprehensive Mass Spectrometry of Lipids, 2016th ed.; Wiley: New York, NY, USA, 2016. [Google Scholar]
- Liebisch, G.; Vizcaíno, J.A.; Köfeler, H.; Trötzmüller, M.; Griffi, W.J.; Schmitz, G.; Spener, F.; Wakelam, M.J.O. Shorthand notation for lipid structures derived from mass spectrometry. J. Lipid Res. 2013, 54, 1523–1530. [Google Scholar] [CrossRef]
- da Costa, E.; Azevedo, V.; Melo, T.; Rego, A.M.; Evtuguin, D.V.; Domingues, P.; Calado, R.; Pereira, R.; Abreu, M.H.; Domingues, M.R.; et al. High-Resolution Lipidomics of the Early Life Stages of the Red Seaweed Porphyra dioica. Molecules 2018, 23, 187. [Google Scholar] [CrossRef]
- Carter, J.D.; Leblond, J.D. Red (hot) algae: Modulation of mono- and digalactosyldiacylglycerol-associated fatty acids of Polysiphonia sp. and Porphyridium sp. in response to growth temperature. Eur. J. Phycol. 2018, 53, 460–470. [Google Scholar] [CrossRef]
- Guella, G.; Frassanito, R.; Mancini, I. A new solution for an old problem: The regiochemical distribution of the acyl chains in galactolipids can be established by electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 1982–1994. [Google Scholar] [CrossRef]
- Hsu, F.-F.; Turk, J. Studies of phosphatidylglycerol with triple quadrupole tandem mass spectrometry with electrospray ionization: Fragmentation processes and structural characterization. J. Am. Soc. Mass Spectrom. 2001, 12, 1036–1043. [Google Scholar] [CrossRef]
- Hsu, F.-F.F.; Turk, J. Characterization of phosphatidylinositol, phosphatidylinositol-4-phosphate, and phosphatidylinositol-4,5-bisphosphate by electrospray ionization tandem mass spectrometry: A mechanistic study. J. Am. Soc. Mass Spectrom. 2000, 11, 986–999. [Google Scholar] [CrossRef]
- Hsu, F.-F.; Turk, J. Charge-Driven Fragmentation Processes in Diacyl Glycerophosphatidic Acids Upon LowEnergy Collisional Activation. A Mechanistic Proposal. J. Am. Soc. Mass Spectrom. 2000, 11, 797–803. [Google Scholar] [CrossRef]
- Hsu, F.-F.F.; Turk, J. Charge-Remote and Charge-Driven Fragmentation Processes in Diacyl Glycerophosphoethanolamine upon Low-Energy Collisional Activation: A Mechanistic Proposal. J. Am. Soc. Mass Spectrom. 2000, 11, 892–899. [Google Scholar] [CrossRef]
- Calvano, C.D.; Bianco, M.; Ventura, G.; Losito, I.; Palmisano, F.; Cataldi, T.R.I. Analysis of Phospholipids, Lysophospholipids, and Their Linked Fatty Acyl Chains in Yellow Lupin Seeds (Lupinus luteus L.) by Liquid Chromatography and Tandem Mass Spectrometry. Molecules 2020, 25, 805. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.C.; Axelsen, P.H. Mass spectrometric analysis of long-chain lipids. Mass Spectrom. Rev. 2011, 30, 579–599. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yoo, J.S.; Kim, M.S. Structural characterization of sulfoquinovosyl, monogalactosyl and digalactosyl diacylglycerols by FAB-CID-MS/MS. J. Mass Spectrom. 1997, 32, 968–977. [Google Scholar] [CrossRef]
- Kim, Y.H.; Choi, J.-S.; Yoo, J.S.; Park, Y.-M.; Kim, M.S. Structural Identification of Glycerolipid Molecular Species Isolated from CyanobacteriumSynechocystissp. PCC 6803 Using Fast Atom Bombardment Tandem Mass Spectrometry. Anal. Biochem. 1999, 267, 260–270. [Google Scholar] [CrossRef]
- Ventura, G.; Bianco, M.; Calvano, C.D.; Losito, I.; Cataldi, T.R.I. HILIC-ESI-FTMS with all ion fragmentation (AIF) scans as a tool for fast lipidome investigations. Molecules 2020, 25, 2310. [Google Scholar] [CrossRef] [PubMed]
- Hvattum, E.; Hagelin, G.; Larsen, Å. Study of mechanisms involved in the collision-induced dissociation of carboxylate anions from glycerophospholipids using negative ion electrospray tandem quadrupole mass spectrometry. Rapid Commun. Mass Spectrom. 1998, 12, 1405–1409. [Google Scholar] [CrossRef]
- Bianco, M.; Calvano, C.D.; Huseynli, L.; Ventura, G.; Losito, I.; Cataldi, T.R.I. Identification and quantification of phospholipids in strawberry seeds and pulp (Fragaria x ananassa cv San Andreas) by liquid chromatography with electrospray ionization and tandem mass spectrometry. J. Mass Spectrom. 2020, 55, e4523. [Google Scholar] [CrossRef] [PubMed]
- Losito, I.; Patruno, R.; Conte, E.; Cataldi, T.R.I.I.; Megli, F.M.; Palmisano, F. Phospholipidomics of Human Blood Microparticles. Anal. Chem. 2013, 85, 6405–6413. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Ishimaru, T.; Itabashi, Y. Lipid Classes, Fatty Acid Composition, and Glycerolipid Molecular Species of the Red Alga Gracilaria vermiculophylla, a Prostaglandin-Producing Seaweed. J. Oleo Sci. 2016, 65, 723–732. [Google Scholar] [CrossRef]
- Parker, P.L.; Van Baalen, C.; Maurer, L. Fatty Acids in Eleven Species of Blue-Green Algae: Geochemical Significance. Science 1967, 155, 707–708. [Google Scholar] [CrossRef]
- Shanmugam, A.; Palpandi, C. Biochemical Composition and Fatty Acid Profile of the Green Alga Ulva reticulata. Asian J. Biochem. 2008, 3, 26–31. [Google Scholar] [CrossRef][Green Version]
- Takagi, T.; Asahi, M.; Itabashi, Y. Fatty Acid Composition of Twelve Algae from Japanese Waters. J. Japan Oil Chem. Soc. 1985, 34, 1008–1012. [Google Scholar] [CrossRef]
- Ishihara, K.; Murata, M.; Kaneiwa, M.; Saito, H.; Komatsu, W.; Shinohara, K. Purification of Stearidonic Acid (18:4(n-3)) and Hexadecatetraenoic Acid (16:4(n-3)) from Algal Fatty Acid with Lipase and Medium Pressure Liquid Cromatography. Biosci. Biotechnol. Biochem. 2000, 64, 2454–2457. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Ventura, G.; Sardanelli, A.M.M.; Savino, L.; Losito, I.; De Michele, G.; Palmisano, F.; Cataldi, T.R.I. Searching for potential lipid biomarkers of parkinson’s disease in parkin-mutant human skin fibroblasts by HILIC-ESI-MS/MS: Preliminary findings. Int. J. Mol. Sci. 2019, 20, 3341. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Glaciale, M.; Palmisano, F.; Cataldi, T.R.I. Glycosphingolipidomics of donkey milk by hydrophilic interaction liquid chromatography coupled to ESI and multistage MS. Electrophoresis 2018, 39, 1634–1644. [Google Scholar] [CrossRef]
- Calvano, C.D.; Ventura, G.; Sardanelli, A.M.; Losito, I.; Palmisano, F.; Cataldi, T.R.I. Identification of neutral and acidic glycosphingolipids in the human dermal fibroblasts. Anal. Biochem. 2019, 581, 113348. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Coniglio, D.; D’Alesio, P.E.; Losito, I.; Cataldi, T.R.I. The occurrence of inositolphosphoceramides in spirulina microalgae. Electrophoresis 2020, 41, 1760–1767. [Google Scholar] [CrossRef]
- Pulfer, M.; Murphy, R.C. Electrospray Mass Spectrometry of Phospholipids. Mass Spectrom. Rev. 2003, 22, 332–364. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Accurate m/z | Mass Error (ppm.) | SQDG (sn-1/sn-2) | Adduct | Molecular Formula | Relative Abundance (%) |
|---|---|---|---|---|---|---|
| 1 | 815.4978 | −0.9 | 18:3/16:0 | [M-H]− | [C43H75O12S]− | 22.4 |
| 2 | 817.5143 | 0.2 | 18:2/16:0 | [C43H77O12S]− | 16.2 | |
| 3 | 819.5322 | 2.9 | 18:1/16:0 | [C43H79O12S]− | 48.2 | |
| SQMG (sn-1/sn-2) | ||||||
| 4 | 555.2820 | −4.5 | 16:0/0:0 | [M-H]− | [C25H47O11S]− | 65.0 |
| 5 | 0:0/16:0 | 12.0 | ||||
| DGDG (sn-1/sn-2) | ||||||
| 6 | 967.6330 | 0.2 | 18:1/18:1; 16:0/20:2 | [M+Na]+ | [C51H92O15Na]+ | 70 |
| 7 | 969.6483 | −0.2 | 18:0/18:1 | [C51H94O15Na]+ | 15 | |
| DGMGb | ||||||
| 8 | 673.3455 | 2.1 | 18:4 | [M-H]− | [C33H53O14]− | 93 |
| 709.3223 | 2.1 | [M+35Cl]− | [C33H54O14Cl]− | |||
| 711.3170 | −1.1 | [M+37Cl]− | [C33H54O14Cl]− | |||
| 9 | 685.3219 | 1.6 | 16:2 | [M+35Cl]− | [C31H54O14Cl]− | 1.2 |
| PG (sn-1/sn-2) | ||||||
| 10 | 719.4895 | 3.6 | 16:0/16:1; 18:1_14:0 | [M-H]− | [C38H72O10P]− | 13.67 |
| 11 | 745.5030 | 0.7 | 16:0/18:2; 18:1/16:1 | [C40H74O10P]− | 20.47 | |
| 12 | 747.5177 | −0.7 | 16:0/18:1 | [C40H76O10P]− | 38.09 | |
| LPGb | ||||||
| 13 | 481.2571 | −0.2 | 16:1 | [M-H]− | [C22H42O9P]− | 28.89 |
| 14 | 483.2730 | 0.4 | 16:0 | [C22H44O9P]− | 20.80 | |
| 15 | 507.2729 | 0.2 | 18:2 | [C24H44O9P]− | 16.87 | |
| 16 | 509.2883 | −0.4 | 18:1 | [C24H46O9P]− | 24.34 | |
| PI (sn-1/sn-2) | ||||||
| 17 | 833.5197 | 1.3 | 16:0/18:2 | [M-H]− | [C43H78O13P]− | 18.78 |
| 18 | 835.5349 | 0.8 | 16:0/18:1 | [C43H80O13P]− | 69.69 | |
| PA (sn-1/sn-2) | ||||||
| 19 | 645.4516 | 2.3 | 16:0/16:1; 14:0/18:1 | [M-H]− | [C35H66O8P]− | 12.80 |
| 20 | 695.4668 | 1.6 | 16:0/20:4; 18:2/18:2 | [C39H68O8P]− | 12.88 | |
| 21 | 743.4671 | 1.9 | 20:4/20:4 | [C43H68O8P]− | 47.85 | |
| PE (sn-1/sn-2) | ||||||
| 22 | 686.4792 | 3.8 | 16:1/16:1 | [M-H]− | [C37H69NO8P]− | 7.11 |
| 23 | 688.4929 | 0.9 | 16:0/16:1; 18:1/14:0 | [C37H71NO8P]− | 63.02 | |
| 24 | 714.5069 | −1.4 | 18:1/16:1 | [C39H73NO8P]− | 9.68 | |
| 25 | 738.5092 | 1.8 | 16:0/20:4 | [C41H73NO8P]− | 6.06 | |
| 26 | 786.5100 | 2.7 | 20:4/20:4 | [C45H73NO8P]− | 5.52 | |
| LPE (sn-1/sn-2) | ||||||
| 27 | 500.2797 | 2.8 | 20:4/0:0 | [M-H]− | [C25H43NO7P]− | 65.07 |
| PC (sn-1/sn-2) | ||||||
| 28 | 774.5302 | 1.4 | 14:0/18:2; 16:0/16:2; | [M+HCOO]− | [C41H77NO10P]− | 9.45 |
| 16:1/16:1; 19:2_13:0 | ||||||
| 29 | 802.5614 | 1.2 | 16:0/18:2 | [C43H81NO10P]− | 34.05 | |
| 30 | 826.5598 | −0.7 | 16:0/20:4; 18:2/18:2 | [C45H81NO10P]− | 17.73 | |
| 31 | 850.5580 | −2.8 | 20:4/18:2 | [C47H81NO10P]− | 12.05 | |
| LPC (sn-1/sn-2) | ||||||
| 32 | 504.3113 | 3.4 | 18:2/0:0 | [M-CH3]− | [C25H47NO7P]− | 11.17 |
| 33 | 564.3323 | 2.8 | 18:2/0:0 | [M+HCOO]− | [C27H51NO9P]− | 23.71 |
| 34 | 588.3323 | 2.7 | 20:4/0:0 | [C29H51NO9P]− | 22.32 | |
| Lipid Class | Precursor Ion | Diagnostic Product Ion for Regiochemical Assignment: sn-1/sn-2 | Comparison between Peak Signal Intensity | Ref. |
|---|---|---|---|---|
| SQDG | [M-H]− | [M-H-R1COOH]−; [M-H-R2COOH]− | [M-H-R1COOH]− > [M-H-R2COOH]− | [31] |
| SQMG | [M-H]− | m/z 243.0, [C6H11O8S]− | Product ion is generated if the FA is at sn-2 | [32] |
| DGDG | [M+Na]+ | [M+Na-R1COOH]+; [M+Na-R2COOH]+ | [M+Na-R1COOH]+ > [M+Na-R2COOH]+ | [37] |
| PG | [M-H]− | [M-H-R1COOH]−; [M-H-R2COOH]− | [M-H-R2COOH]− > [M-H-R1COOH]− | [38] |
| PI | [M-H]− | [M-H-R1COOH]−; [M-H-R2COOH]− | [M-H-R2COOH]− > [M-H-R1COOH]− | [39] |
| PA | [M-H]− | [M-H-R1COOH]−; [M-H-R2COOH]− | [M-H-R2COOH]− > [M-H-R1COOH]− | [40] |
| PE | [M-H]− | [M-H-R’1=C=O]−; [M-H-R’2=C=O]− | [M-H-R’2=C=O]− > [M-H-R’1=C=O]− | [41] |
| LPE | [M-H]− | m/z 196.1, [M-H-RCOOH]−; | If RCOOH is in sn-1: [M-H-RCOOH]− > [M-H-R’=C=O]− else viceversa | [42] |
| m/z 214.1, [M-H-R’=C=O]− | ||||
| PC | [M-CH3]−, | [M-CH3-R’1=C=O]−; [M-CH3-R’2=C=O]− | [M-CH3-R’2=C=O]− > [M-CH3-R’1=C=O]− | [43] |
| [M+HCOO]− | ||||
| LPC | [M-CH3]−, | m/z 224.1, [M-CH3-RCOOH]− | If RCOOH is in sn-1: [M-CH3-RCOOH]− > [M-CH3-R’=C=O]− else viceversa | [42] |
| [M+HCOO]− | m/z 242.1, [M-CH3-R’=C=O]− |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coniglio, D.; Bianco, M.; Ventura, G.; Calvano, C.D.; Losito, I.; Cataldi, T.R.I. Lipidomics of the Edible Brown Alga Wakame (Undaria pinnatifida) by Liquid Chromatography Coupled to Electrospray Ionization and Tandem Mass Spectrometry. Molecules 2021, 26, 4480. https://doi.org/10.3390/molecules26154480
Coniglio D, Bianco M, Ventura G, Calvano CD, Losito I, Cataldi TRI. Lipidomics of the Edible Brown Alga Wakame (Undaria pinnatifida) by Liquid Chromatography Coupled to Electrospray Ionization and Tandem Mass Spectrometry. Molecules. 2021; 26(15):4480. https://doi.org/10.3390/molecules26154480
Chicago/Turabian StyleConiglio, Davide, Mariachiara Bianco, Giovanni Ventura, Cosima D. Calvano, Ilario Losito, and Tommaso R. I. Cataldi. 2021. "Lipidomics of the Edible Brown Alga Wakame (Undaria pinnatifida) by Liquid Chromatography Coupled to Electrospray Ionization and Tandem Mass Spectrometry" Molecules 26, no. 15: 4480. https://doi.org/10.3390/molecules26154480
APA StyleConiglio, D., Bianco, M., Ventura, G., Calvano, C. D., Losito, I., & Cataldi, T. R. I. (2021). Lipidomics of the Edible Brown Alga Wakame (Undaria pinnatifida) by Liquid Chromatography Coupled to Electrospray Ionization and Tandem Mass Spectrometry. Molecules, 26(15), 4480. https://doi.org/10.3390/molecules26154480