
Vitamin E Can Ameliorate Oxidative Damage of Ovine Hepatocytes In Vitro by Regulating Genes Expression Associated with Apoptosis and Pyroptosis, but Not Ferroptosis


 and
and
Abstract
:1. Introduction
2. Results
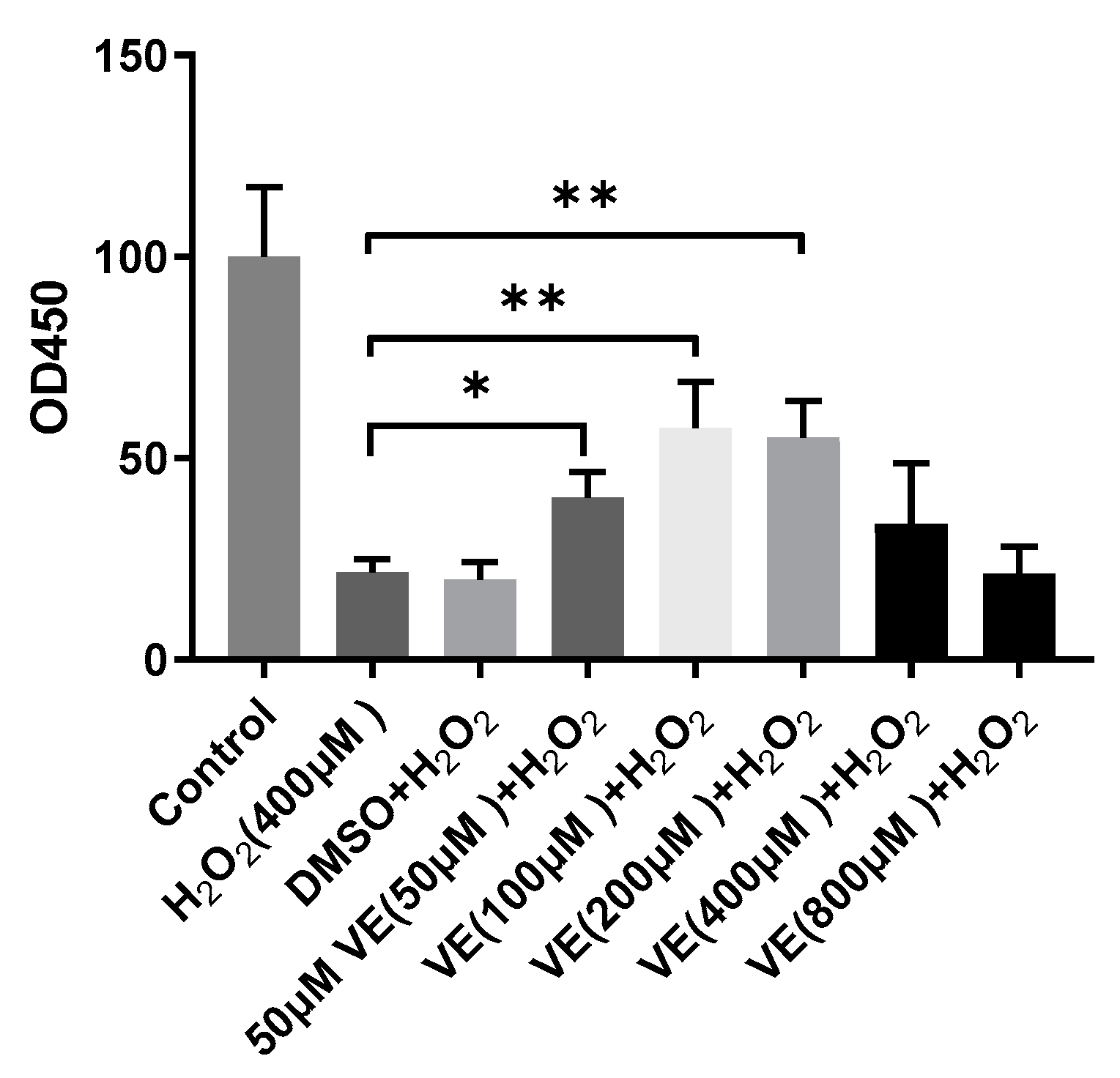
2.1. Vitamin E Inhibits H2O2-Induced Cell Death in Primary Sheep Hepatocytes
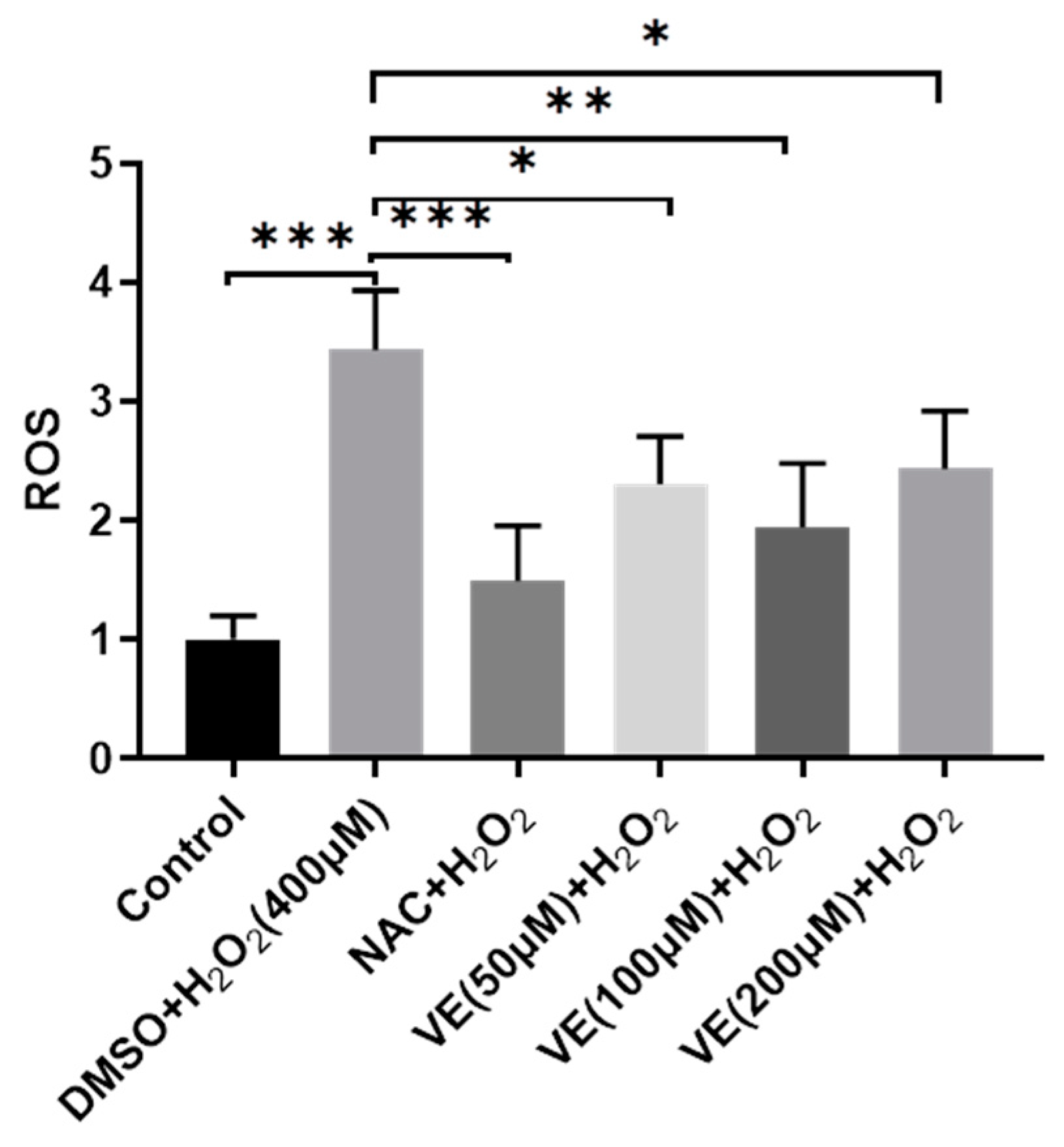
2.2. VE Ameliorates H2O2-Induced Oxidative Stress in Primary Sheep Hepatocytes
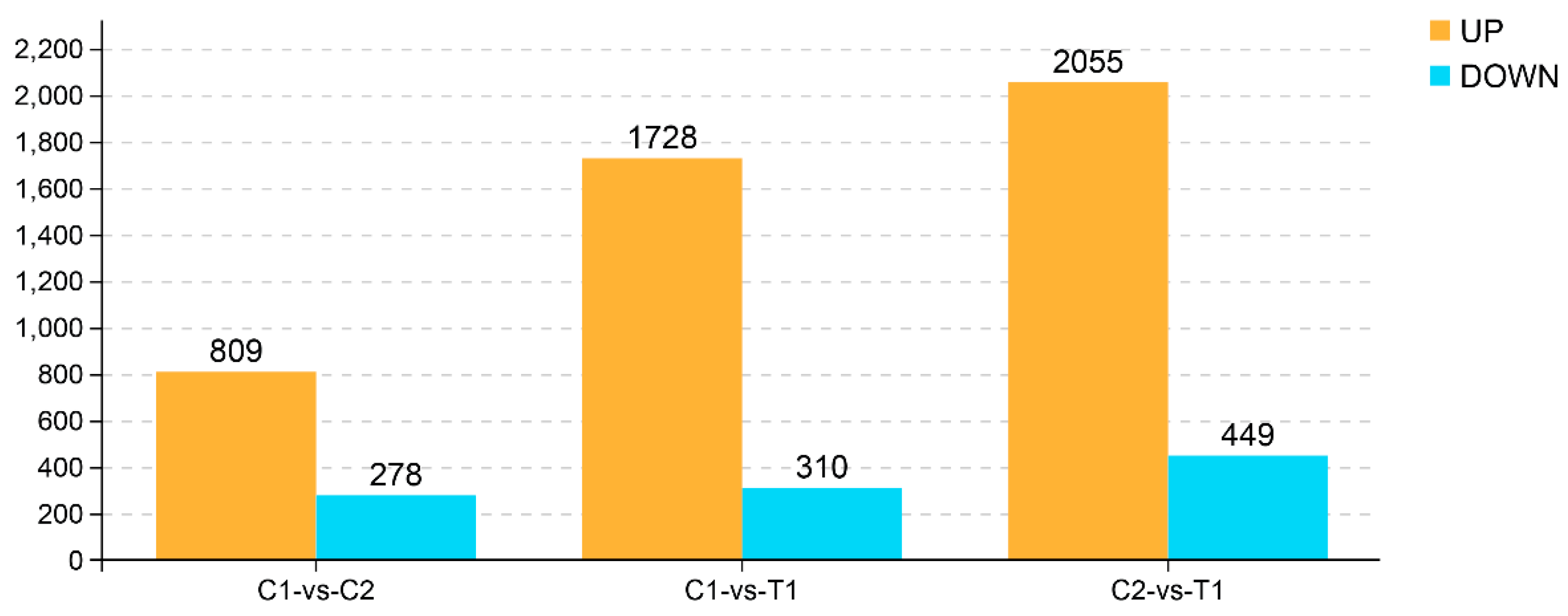
2.3. Number of Genes and Differentially Expressed Genes
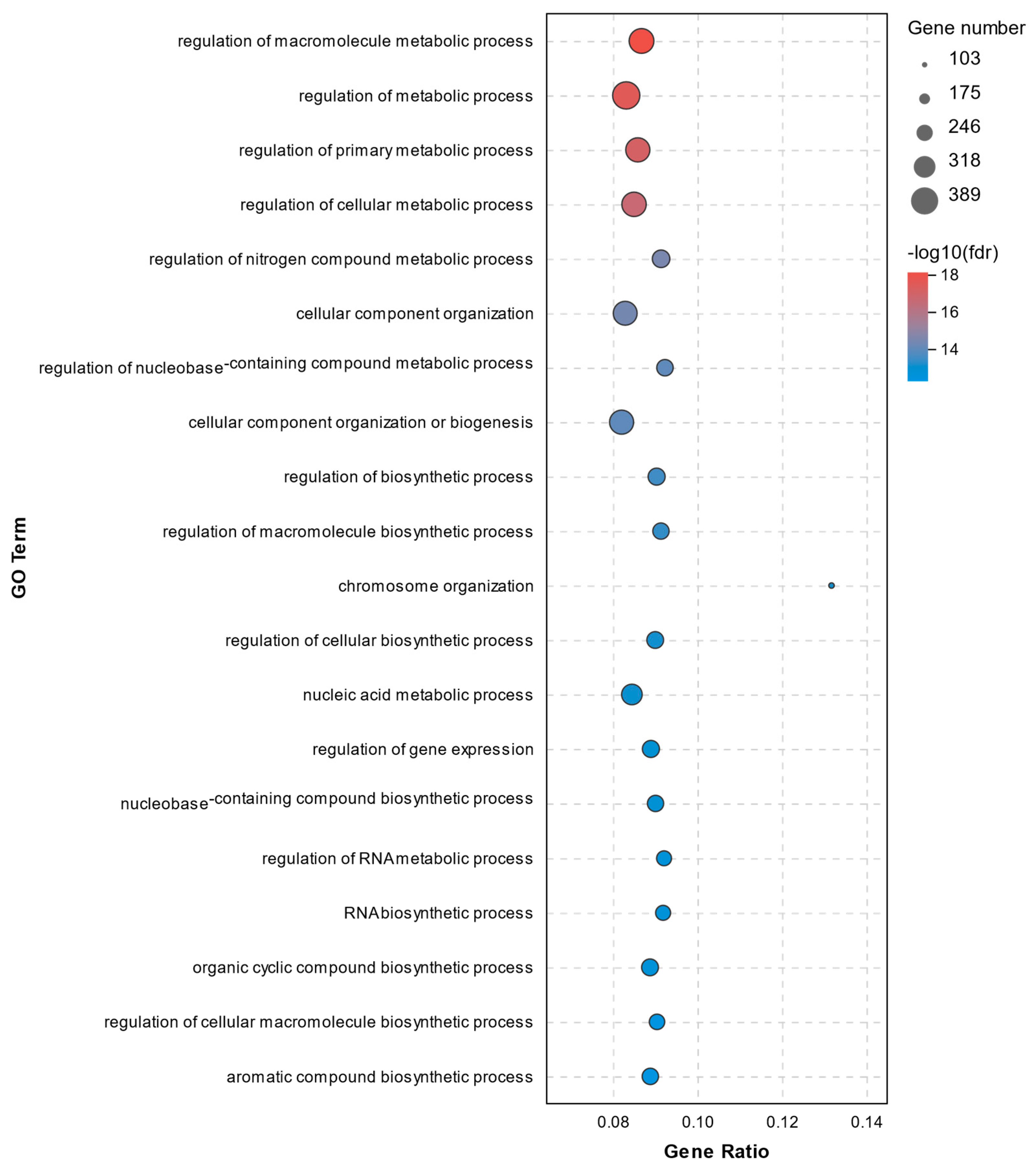
2.4. GO Analysis of DEGs
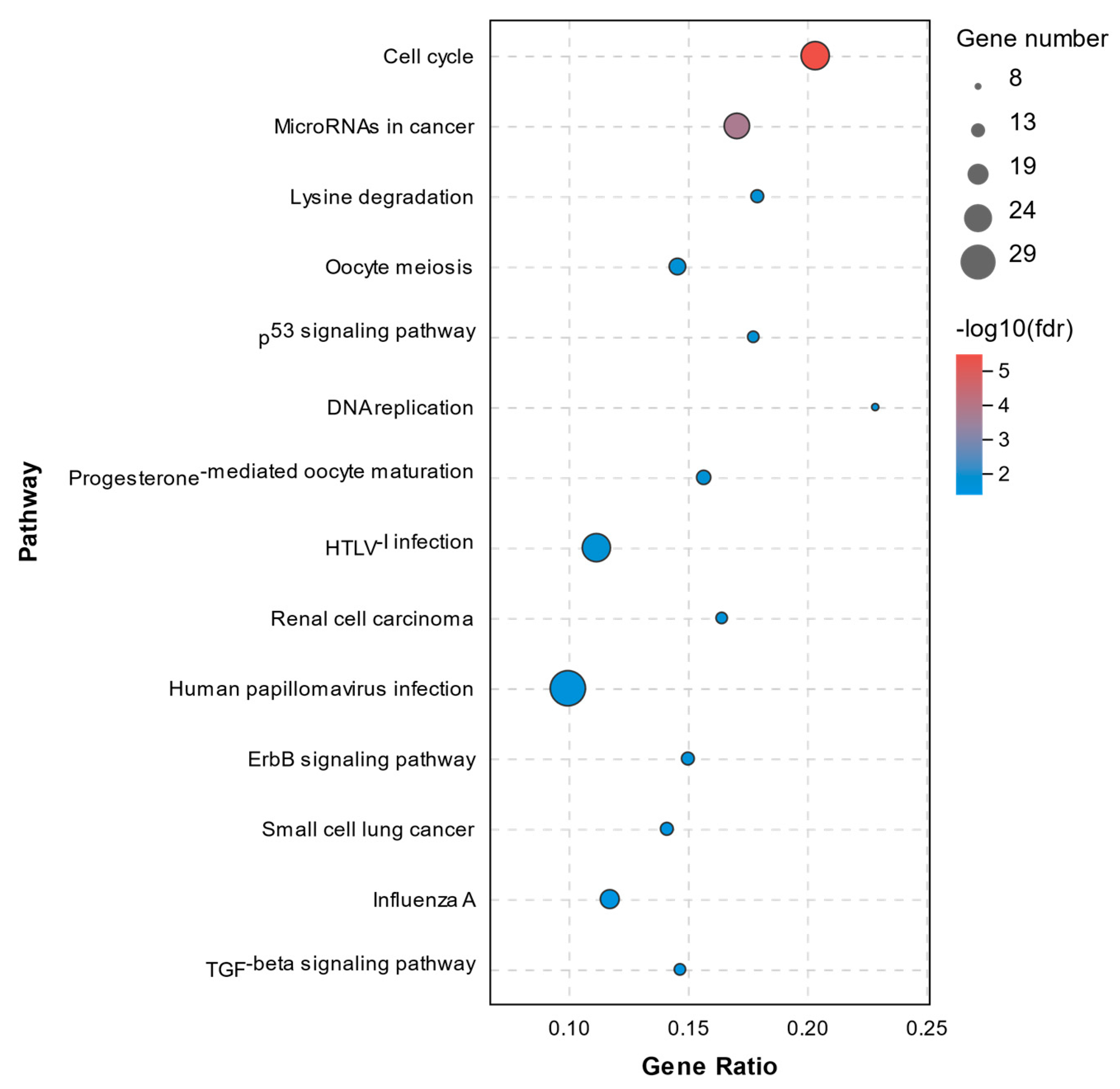
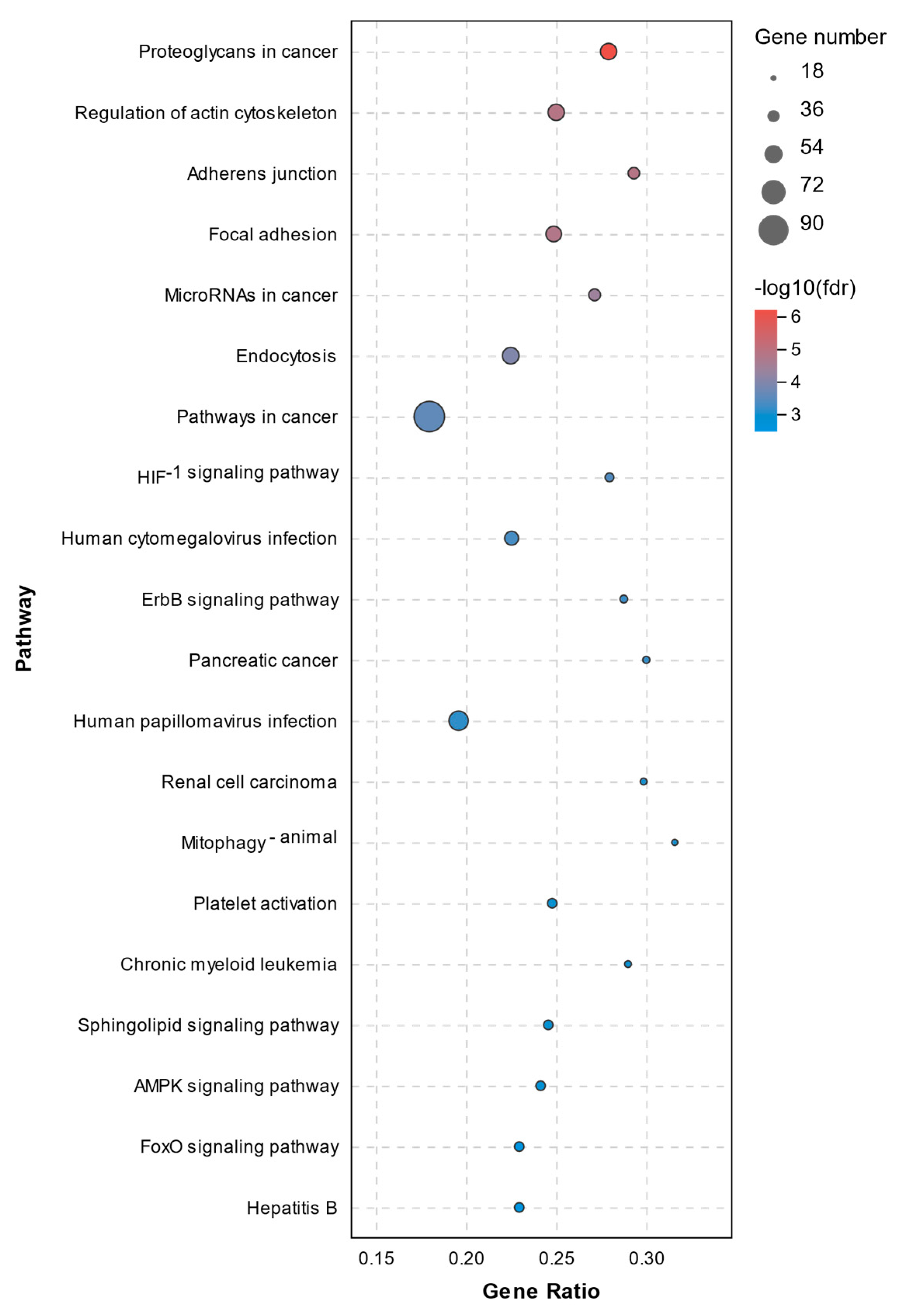
2.5. KEGG Pathway Analysis of DEGs
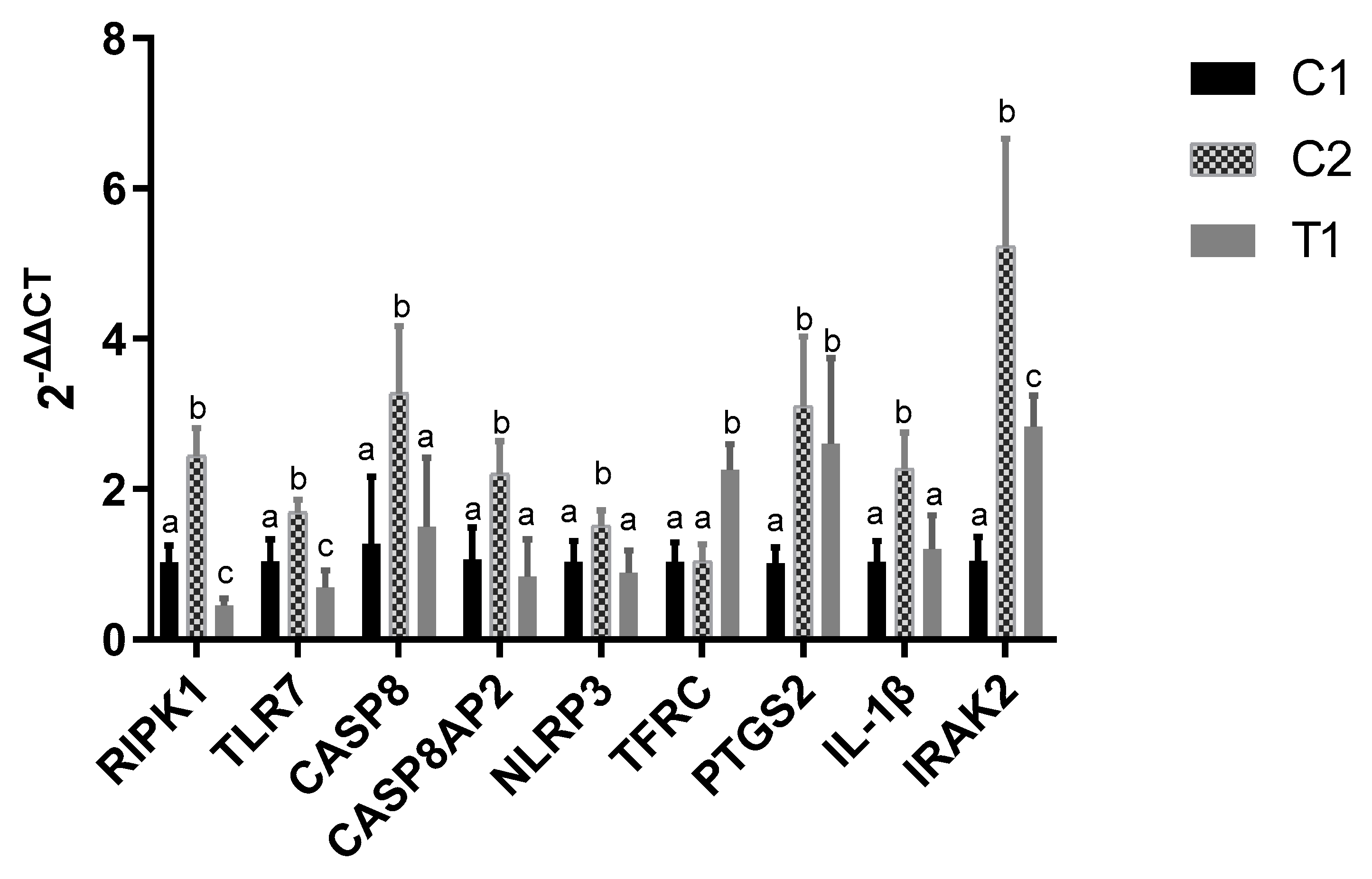
2.6. Validation of Genes by qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation and Characterization of Primary Sheep Hepatocytes (PSH)
4.3. Cell Viability Assay
4.4. Detection of Intracellular ROS
4.5. Sample Preparation, RNA Extraction, Library Construction, and Sequencing
4.6. Quality Control and Alignment with a Reference Genome
4.7. Transcript Assembly and Gene Abundance Quantification
4.8. Summary of the RNA-Seq Data
4.9. Identification of DEGs
4.10. Validation of RNA-Seq Data
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Evans, H.M.; Bishop, K.S. On the Existence of a Hitherto Unrecognized Dietary Factor Essential for Reproduction. Science 1922, 56, 650–651. [Google Scholar] [CrossRef] [Green Version]
- Rotzoll, D.E.; Scherling, R.; Etzl, R.; Stepan, H.; Horn, L.C.; Poschl, J.M. Immunohistochemical localization of alpha-tocopherol transfer protein and lipoperoxidation products in human first-trimester and term placenta. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 140, 183–191. [Google Scholar] [CrossRef]
- Colombo, M.L. An update on vitamin E, tocopherol and tocotrienol-perspectives. Molecules 2010, 15, 2103–2113. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Nallella, K.P.; Allamaneni, S.S.; Said, T.M. Role of antioxidants in treatment of male infertility: An overview of the literature. Reprod. Biomed. Online 2004, 8, 616–627. [Google Scholar] [CrossRef]
- Xu, C.; Zuo, Z.; Liu, K.; Jia, H.; Zhang, Y.; Luo, H. Transcriptome analysis of the Tan sheep testes: Differential expression of antioxidant enzyme-related genes and proteins in response to dietary vitamin E supplementation. Gene 2016, 579, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.Y.; Luo, H.L.; Liu, K.; Jia, H.N.; Zhang, Y.W.; Jiao, L.J.; Chang, Y.F. Dietary vitamin E affects alpha-TTP mRNA levels in different tissues of the Tan sheep. Gene 2014, 541, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.H.; Fu, J.C.; Liu, K.; Zuo, Z.Y.; Jia, H.N.; Ma, Y.; Luo, H.L. Screening of alpha-Tocopherol Transfer Protein Sensitive Genes in Human Hepatoma Cells (HepG2). Int. J. Mol. Sci. 2016, 17, 1016. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Jian, L.; Lu, W.; Xue, Y.; Luo, H. Vitamin E can promote spermatogenesis by regulating the expression of proteins associated with the plasma membranes and protamine biosynthesis. Gene 2021, 773, 145364. [Google Scholar] [CrossRef]
- Head, B.; Ramsey, S.A.; Kioussi, C.; Tanguay, R.L.; Traber, M.G. Vitamin E Deficiency Disrupts Gene Expression Networks during Zebrafish Development. Nutrients 2021, 13, 468. [Google Scholar] [CrossRef] [PubMed]
- Baquerre, C.; Montillet, G.; Pain, B. Liver organoids in domestic animals: An expected promise for metabolic studies. Vet. Res. 2021, 52, 47. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.P.; Pavlovic, I.; Poljsak, B.; Suput, D.; Milisav, I. Beneficial Role of ROS in Cell Survival: Moderate Increases in H2O2 Production Induced by Hepatocyte Isolation Mediate Stress Adaptation and Enhanced Survival. Antioxidants 2019, 8, 434. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei Khorramabadi, K.; Reza Talebi, A.; Abbasi Sarcheshmeh, A.; Mirjalili, A. Protective effect of vitamin E on oxidative stress and sperm apoptosis in diabetic Mice. Int. J. Reprod. Biomed. 2019, 17, 127. [Google Scholar] [CrossRef] [PubMed]
- Florean, C.; Song, S.; Dicato, M.; Diederich, M. Redox biology of regulated cell death in cancer: A focus on necroptosis and ferroptosis. Free Radic. Biol. Med. 2019, 134, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.S.; Gao, L.; Xie, X.L.; Ren, Y.L.; Shen, Z.Q.; Wang, F.; Shen, M.; Eyporsdottir, E.; Hallsson, J.H.; Kiseleva, T.; et al. Genome-Wide Association Analyses Highlight the Potential for Different Genetic Mechanisms for Litter Size Among Sheep Breeds. Front. Genet 2018, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Ito, Y.; Shi, L.; Amin, P.; Chu, J.; Ouchida, A.T.; Mookhtiar, A.K.; Zhao, H.; Xu, D.; Shan, B.; et al. Regulation of RIPK1 activation by TAK1-mediated phosphorylation dictates apoptosis and necroptosis. Nat. Commun. 2017, 8, 359. [Google Scholar] [CrossRef] [PubMed]
- Mihaly, S.R.; Ninomiya-Tsuji, J.; Morioka, S. TAK1 control of cell death. Cell Death Differ. 2014, 21, 1667–1676. [Google Scholar] [CrossRef] [Green Version]
- Morioka, S.; Broglie, P.; Omori, E.; Ikeda, Y.; Takaesu, G.; Matsumoto, K.; Ninomiya-Tsuji, J. TAK1 kinase switches cell fate from apoptosis to necrosis following TNF stimulation. J. Cell Biol. 2014, 204, 607–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tummers, B.; Green, D.R. Caspase-8: Regulating life and death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Muzio, M.; Chinnaiyan, A.M.; Kischkel, F.C.; O’Rourke, K.; Dixit, V.M. FLICE, a novel FADD-homologous ICE/CED-3-like protease, is recruited to the CD95 (Fas/APO-1) death--inducing signaling complex. Cell 1996, 85, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Dowling, J.P.; Zhang, J. RIPK1 can mediate apoptosis in addition to necroptosis during embryonic development. Cell Death. Dis. 2019, 10, 245. [Google Scholar] [CrossRef]
- Tao, P.; Sun, J.; Wu, Z.; Wang, S.; Wang, J.; Li, W.; Pan, H.; Bai, R.; Zhang, J.; Wang, Y.; et al. A dominant autoinflammatory disease caused by non-cleavable variants of RIPK1. Nature 2020, 577, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Wickliffe, K.E.; Dugger, D.L.; Maltzman, A.; Roose-Girma, M.; Dohse, M.; Kőműves, L.; Webster, J.D.; Dixit, V.M. Cleavage of RIPK1 by caspase-8 is crucial for limiting apoptosis and necroptosis. Nature 2019, 574, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Lalaoui, N.; Boyden, S.E.; Oda, H.; Wood, G.M.; Stone, D.L.; Chau, D.; Liu, L.; Stoffels, M.; Kratina, T.; Lawlor, K.E.; et al. Mutations that prevent caspase cleavage of RIPK1 cause autoinflammatory disease. Nature 2020, 577, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Dugger, D.L.; Maltzman, A.; Greve, J.M.; Hedehus, M.; Martin-Mcnulty, B.; Carano, R.; Cao, T.C.; Bruggen, N.V.; Bernstein, L. RIPK3 deficiency or catalytically inactive RIPK1 provides greater benefit than MLKL deficiency in mouse models of inflammation and tissue injury. Cell Death Differ. 2016, 23, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Gu, T.X.; Yu, F.M.; Zhang, G.W.; Zhao, Y. Overexpression of miR-210 promotes the potential of cardiac stem cells against hypoxia. Scand. Cardiovasc. J. 2018, 52, 367–371. [Google Scholar] [CrossRef]
- Zeng, C.; Duan, F.; Hu, J.; Luo, B.; Huang, B.; Lou, X.; Sun, X.; Li, H.; Zhang, X.; Yin, S.; et al. NLRP3 inflammasome-mediated pyroptosis contributes to the pathogenesis of non-ischemic dilated cardiomyopathy. Redox. Biol. 2020, 34, 101523. [Google Scholar] [CrossRef]
- Frank, D.; Vince, J.E. Pyroptosis versus necroptosis: Similarities, differences, and crosstalk. Cell Death Differ. 2019, 26, 99–114. [Google Scholar] [CrossRef]
- Hirschhorn, T.; Stockwell, B.R. The development of the concept of ferroptosis. Free Radic. Biol. Med. 2019, 133, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascon, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Fu, V.; Plouffe, S.W.; Guan, K.L. The Hippo pathway in organ development, homeostasis, and regeneration. Curr. Opin. Cell Biol. 2017, 49, 99–107. [Google Scholar] [CrossRef]
- Wu, J.; Minikes, A.M.; Gao, M.; Bian, H.; Li, Y.; Stockwell, B.R.; Chen, Z.N.; Jiang, X. Intercellular interaction dictates cancer cell ferroptosis via NF2-YAP signalling. Nature 2019, 572, 402–406. [Google Scholar] [CrossRef]
- Yang, W.H.; Ding, C.C.; Sun, T.; Rupprecht, G.; Lin, C.C.; Hsu, D.; Chi, J.T. The Hippo Pathway Effector TAZ Regulates Ferroptosis in Renal Cell Carcinoma. Cell Rep. 2019, 28, 2501–2508. [Google Scholar] [CrossRef]
- Dai, C.; Chen, X.; Li, J.; Comish, P.; Kang, R.; Tang, D. Transcription factors in ferroptotic cell death. Cancer Gene Ther. 2020, 27, 645–656. [Google Scholar] [CrossRef]
- Sampilvanjil, A.; Karasawa, T.; Yamada, N.; Komada, T.; Higashi, T.; Baatarjav, C.; Watanabe, S.; Kamata, R.; Ohno, N.; Takahashi, M. Cigarette smoke extract induces ferroptosis in vascular smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2020, 318, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Jiang, Y.; Liang, W.; Wang, Y.; Cao, S.; Yan, H.; Gao, L.; Zhang, L. miR-212-5p attenuates ferroptotic neuronal death after traumatic brain injury by targeting Ptgs2. Mol. Brain 2019, 12, 78. [Google Scholar] [CrossRef] [PubMed]
- Agmon, E.; Stockwell, B.R. Lipid homeostasis and regulated cell death. Curr. Opin. Chem. Biol. 2017, 39, 83–89. [Google Scholar] [CrossRef]
- Dalleau, S.; Baradat, M.; Gueraud, F.; Huc, L. Cell death and diseases related to oxidative stress: 4-hydroxynonenal (HNE) in the balance. Cell Death Differ. 2013, 20, 1615–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulatowski, L.M.; Manor, D. Vitamin E and neurodegeneration. Neurobiol. Dis. 2015, 84, 78–83. [Google Scholar] [CrossRef]
- Lee, S.M.; Schelcher, C.; Demmel, M.; Hauner, M.; Thasler, W.E. Isolation of human hepatocytes by a two-step collagenase perfusion procedure. J. Vis. Exp. 2013, 50615. [Google Scholar] [CrossRef] [Green Version]
- Xiang, C.; Du, Y.; Meng, G.; Soon Yi, L.; Sun, S.; Song, N.; Zhang, X.; Xiao, Y.; Wang, J.; Yi, Z.; et al. Long-term functional maintenance of primary human hepatocytes in vitro. Science 2019, 364, 399–402. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
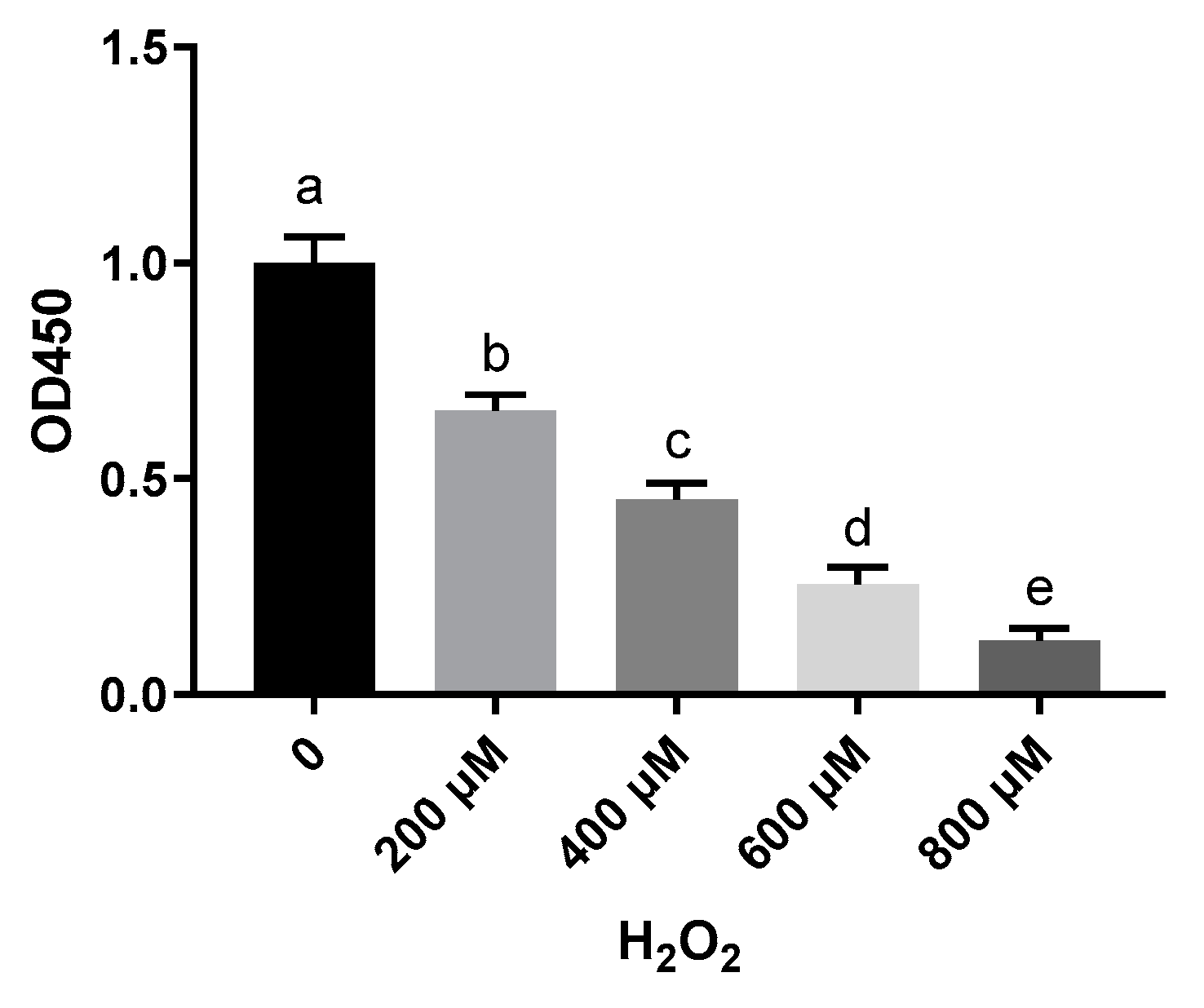


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Product Size (bp) |
|---|---|---|
| β-actin | 5′ CATCGTCCACCGCAAAT 3′ | 17 |
| 5′ GCCATGCCAATCTCATCTC 3′ | 19 | |
| NLRP3 | 5′ TGCAGCCTCACATCACA 3′ | 17 |
| 5′ ATCACCCAGGTCGTTGTT 3′ | 18 | |
| IL-1β | 5′ CAGCCGTGCAGTCAGTAA 3′ | 18 |
| 5′ TGTGAGAGGAGGTGGAGAG 3′ | 19 | |
| IRAK2 | 5′ TCACCCATGTCCTGTCAA 3′ | 18 |
| 5′ TGCCCCACTCTGATGAA 3′ | 17 | |
| CASP8 | 5′ AGGCAATGGTTTCACAGG 3′ | 18 |
| 5′ TCCACCAGGCTTTTATGC 3′ | 18 | |
| CASP8AP2 | 5′ AAGAGGACGCATCTGAACA 3′ | 19 |
| 5′ TACTGAAAGCCTGGAGCAA 3′ | 19 | |
| PTGS2 | 5′ CCAGGGAGACAATGCTTCT 3′ | 19 |
| 5′ TGCAGCCTTAAACCCAGT 3′ | 18 | |
| TFRC | 5′ AGAGTTTCCTTTCCGACACA 3′ | 20 |
| 5′ CAGCTCCCTGAATAGTCCAA 3′ | 20 | |
| RIPK1 | 5′ TCTCGGGTTGTGTGTTTCC 3′ | 19 |
| 5′ ACATGGCCGCTTTCCTT 3′ | 17 | |
| TLR7 | 5′ TCCATTTCCTTGCACACC 3′ | 18 |
| 5′ CCATCTTTGGGGCACAT 3′ | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jian, L.; Xue, Y.; Gao, Y.; Wang, B.; Qu, Y.; Li, S.; Li, H.; Li, Z.; Wang, B.; Luo, H. Vitamin E Can Ameliorate Oxidative Damage of Ovine Hepatocytes In Vitro by Regulating Genes Expression Associated with Apoptosis and Pyroptosis, but Not Ferroptosis. Molecules 2021, 26, 4520. https://doi.org/10.3390/molecules26154520
Jian L, Xue Y, Gao Y, Wang B, Qu Y, Li S, Li H, Li Z, Wang B, Luo H. Vitamin E Can Ameliorate Oxidative Damage of Ovine Hepatocytes In Vitro by Regulating Genes Expression Associated with Apoptosis and Pyroptosis, but Not Ferroptosis. Molecules. 2021; 26(15):4520. https://doi.org/10.3390/molecules26154520
Chicago/Turabian StyleJian, Luyang, Ying Xue, Yuefeng Gao, Bo Wang, Yanghua Qu, Shuanghong Li, Heqiong Li, Zhen Li, Bing Wang, and Hailing Luo. 2021. "Vitamin E Can Ameliorate Oxidative Damage of Ovine Hepatocytes In Vitro by Regulating Genes Expression Associated with Apoptosis and Pyroptosis, but Not Ferroptosis" Molecules 26, no. 15: 4520. https://doi.org/10.3390/molecules26154520
APA StyleJian, L., Xue, Y., Gao, Y., Wang, B., Qu, Y., Li, S., Li, H., Li, Z., Wang, B., & Luo, H. (2021). Vitamin E Can Ameliorate Oxidative Damage of Ovine Hepatocytes In Vitro by Regulating Genes Expression Associated with Apoptosis and Pyroptosis, but Not Ferroptosis. Molecules, 26(15), 4520. https://doi.org/10.3390/molecules26154520