
Glycosidically-Bound Volatile Phenols Linked to Smoke Taint: Stability during Fermentation with Different Yeasts and in Finished Wine


 , , ,
, , ,
Abstract
:1. Introduction
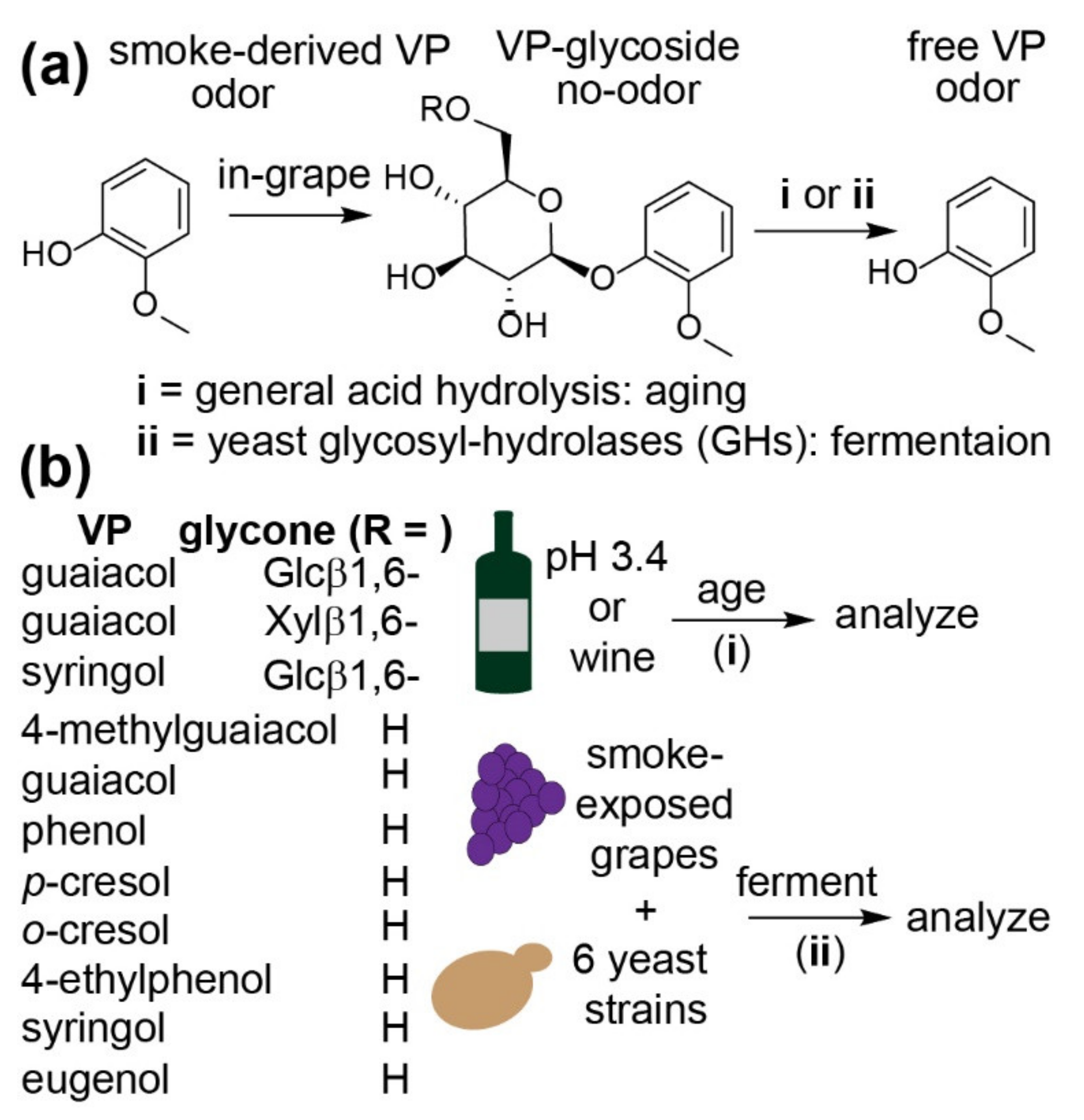
2. Results and Discussion
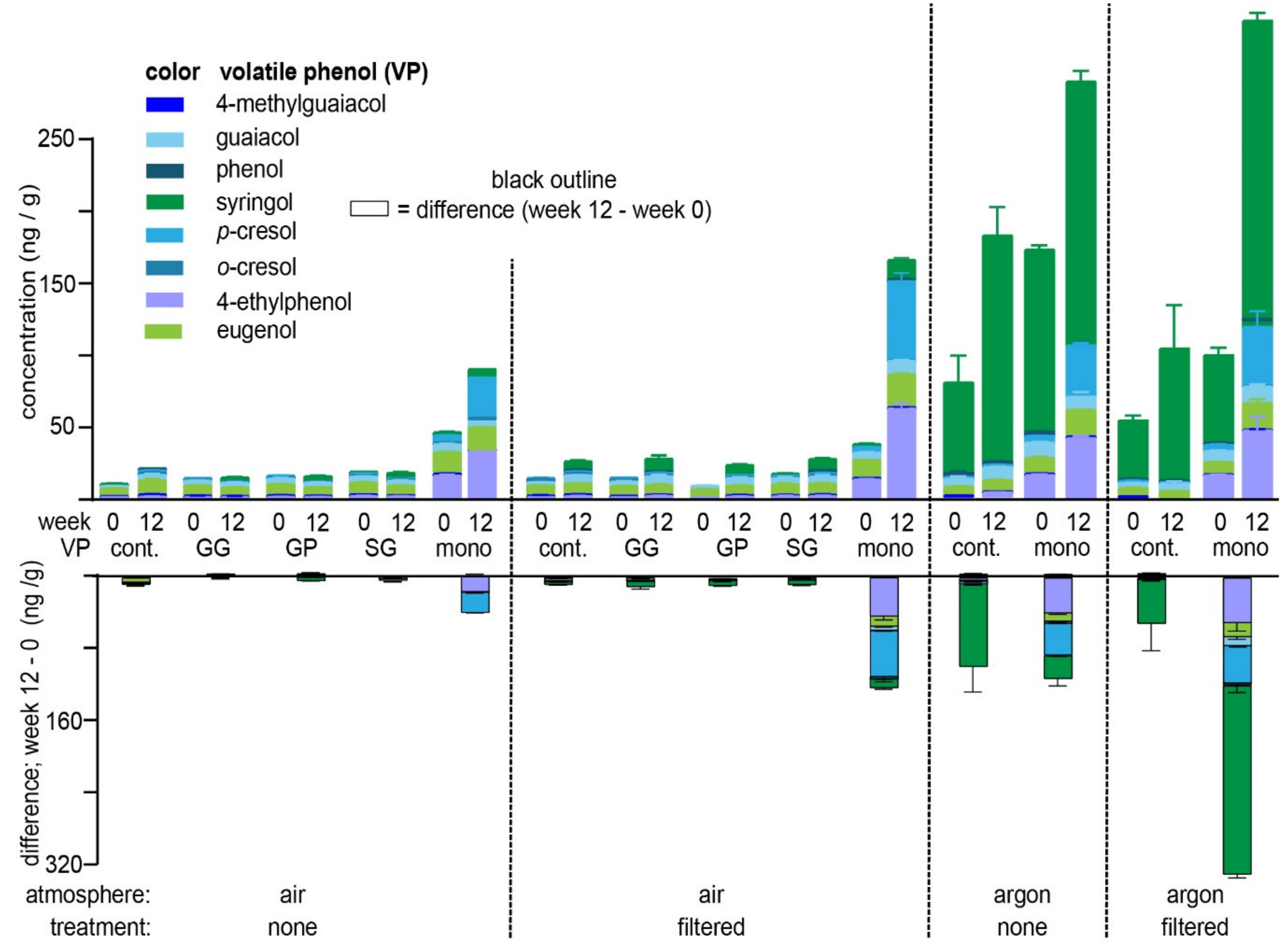
2.1. Hydrolysis of VP-Glycosides during Aging in a Model Wine
2.2. Hydrolysis of VP-Glycosides during Aging in a Commercial Wine
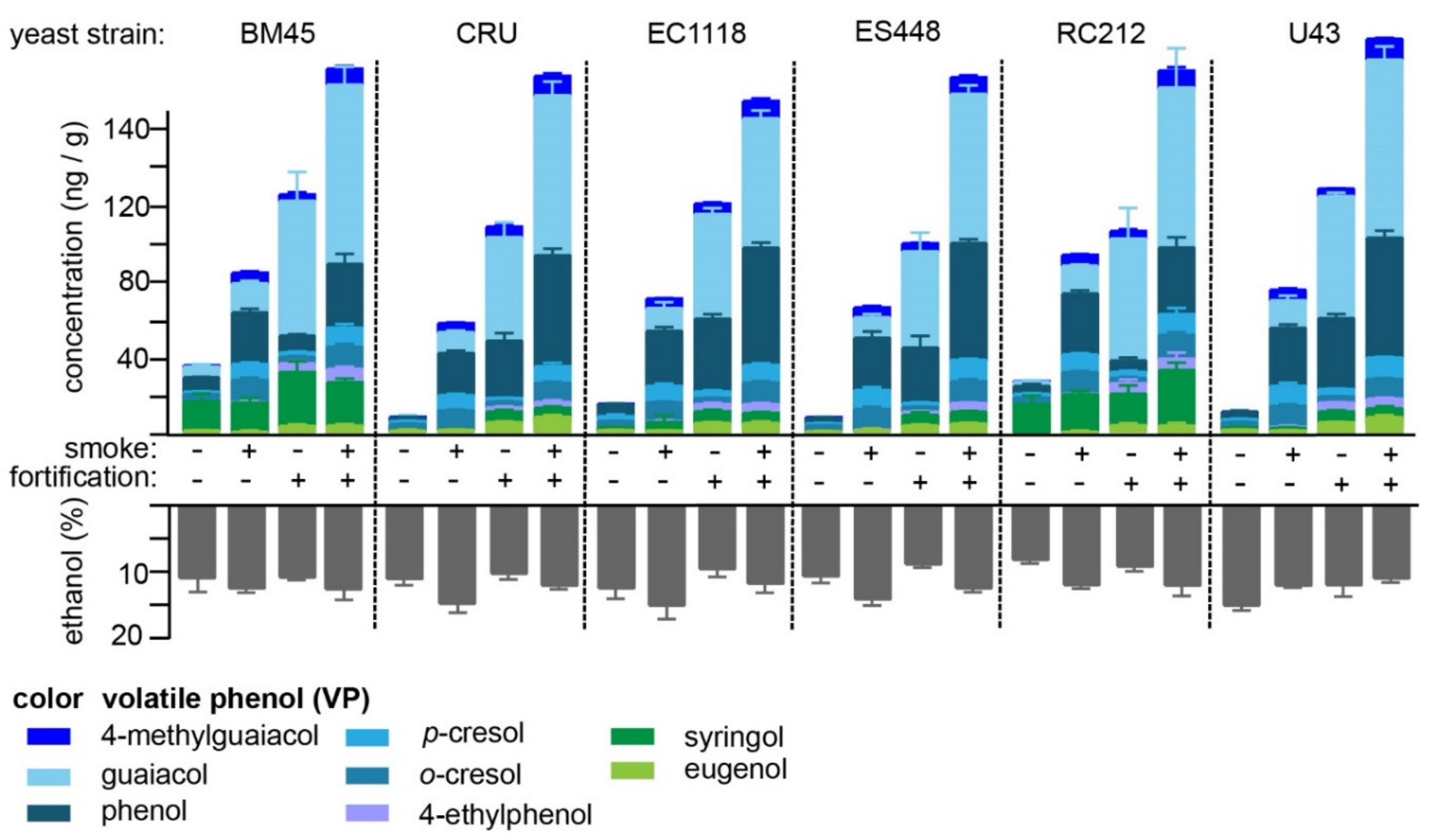
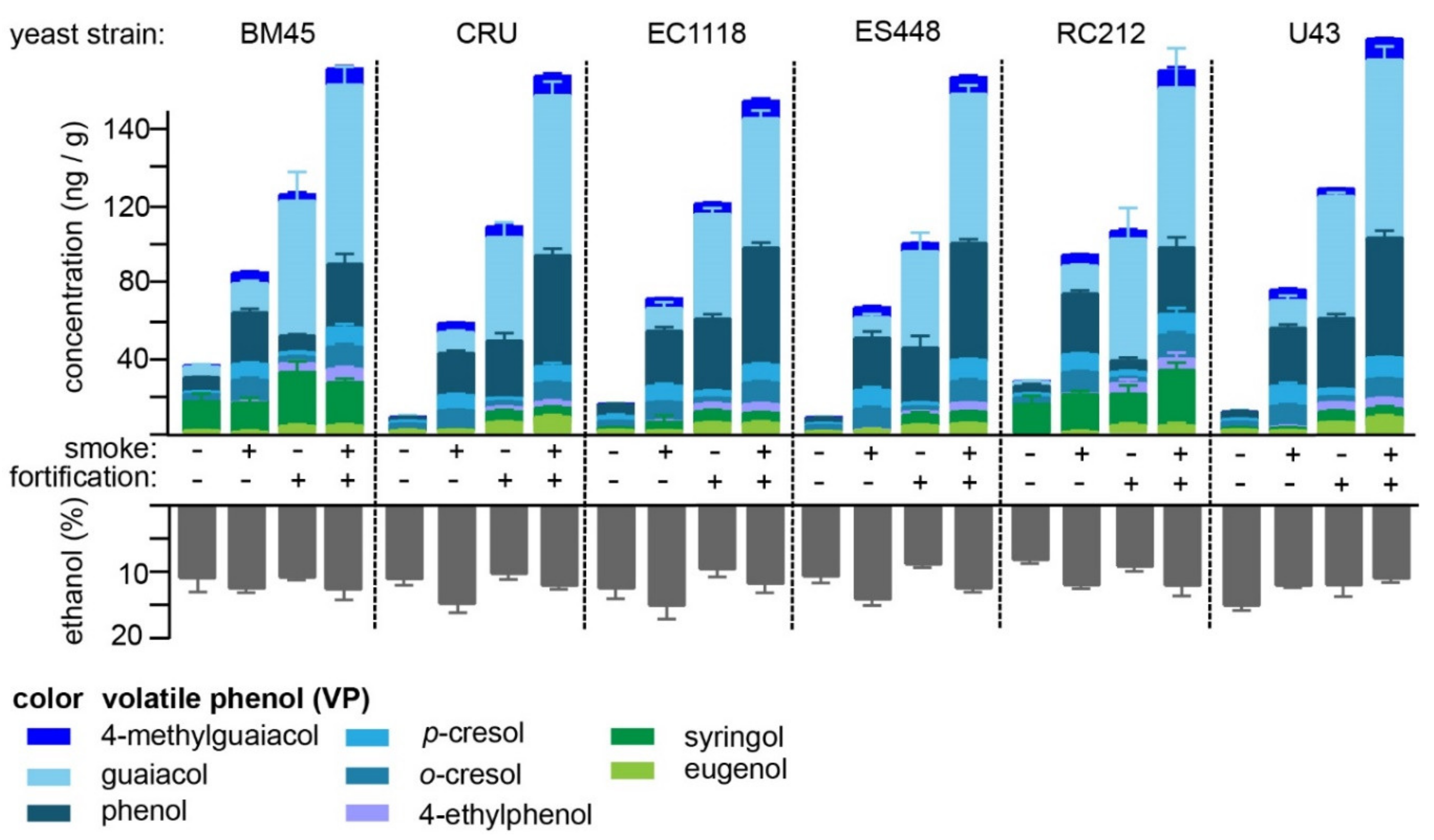
2.3. Hydrolysis of VP-Glucosides during Primary Fermentation with Different Yeast Strains
3. Materials and Methods
3.1. Chemicals and General Details
3.2. Stock and Calibration Solutions
3.3. Spontaneous Hydrolysis
3.4. Vinification
3.5. Sample Preparation and Analysis
3.6. Data Acquisition and Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Krstic, M.P.; Johnson, D.L.; Herderich, M.J. Review of smoke taint in wine: Smoke-derived volatile phenols and their glycosidic metabolites in grapes and vines as biomarkers for smoke exposure and their role in the sensory perception of smoke taint. Aust. J. Grape Wine Res. 2015, 21, 537–553. [Google Scholar] [CrossRef]
- Parker, M.; Osidacz, P.; Baldock, G.A.; Hayasaka, Y.; Black, C.A.; Pardon, K.H.; Jeffery, D.W.; Geue, J.P.; Herderich, M.J.; Francis, I.L. Contribution of several volatile phenols and their glycoconjugates to smoke-related sensory properties of red wine. J. Agric. Food Chem. 2012, 60, 2629–2637. [Google Scholar] [CrossRef]
- Kennison, K.R.; Gibberd, M.R.; Pollnitz, A.P.; Wilkinson, K.L. Smoke-derived taint in wine: The release of smoke-derived volatile phenols during fermentation of Merlot juice following grapevine exposure to smoke. J. Agric. Food Chem. 2008, 56, 7379–7383. [Google Scholar] [CrossRef] [PubMed]
- Dungey, K.A.; Hayasaka, Y.; Wilkinson, K.L. Quantitative analysis of glycoconjugate precursors of guaiacol in smoke-affected grapes using liquid chromatography-tandem mass spectrometry based stable isotope dilution analysis. Food Chem. 2011, 126, 801–806. [Google Scholar] [CrossRef]
- Hayasaka, Y.; Baldock, G.A.; Parker, M.; Pardon, K.H.; Black, C.A.; Herderich, M.J.; Jeffery, D.W. Glycosylation of smoke-derived volatile phenols in grapes as a consequence of grapevine exposure to bushfire smoke. J. Agric. Food Chem. 2010, 58, 10989–10998. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Härtl, K.; McGraphery, K.; Hoffmann, T.; Schwab, W. Attractive but toxic: Emerging roles of glycosidically bound volatiles and glycosyltransferases involved in their formation. Mol. Plant 2018, 11, 1225–1236. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, X.L.; Ullah, N.; Tao, Y.S. Aroma glycosides in grapes and wine. J. Food Sci. 2017, 82, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Caffrey, A.; Lerno, L.; Rumbaugh, A.; Girardello, R.; Zweigenbaum, J.; Oberholster, A.; Ebeler, S.E. Changes in smoke-taint volatile-phenol glycosides in wildfire smoke-exposed Cabernet Sauvignon grapes throughout winemaking. Am. J. Enol. Vitic. 2019, 70, 373–381. [Google Scholar] [CrossRef]
- Mayr, C.M.; Parker, M.; Baldock, G.A.; Black, C.A.; Pardon, K.H.; Williamson, P.O.; Herderich, M.J.; Francis, I.L. Determination of the importance of in-mouth release of volatile phenol glycoconjugates to the flavor of smoke-tainted wines. J. Agric. Food Chem. 2014, 62, 2327–2336. [Google Scholar] [CrossRef]
- Szeto, C.; Ristic, R.; Capone, D.; Puglisi, C.; Pagay, V.; Culbert, J.; Jiang, W.W.; Herderich, M.; Tuke, J.; Wilkinson, K. Uptake and glycosylation of smoke-derived volatile phenols by Cabernet Sauvignon grapes and their subsequent fate during winemaking. Molecules 2020, 25, 3720. [Google Scholar] [CrossRef]
- Noestheden, M.; Thiessen, K.; Dennis, E.G.; Tiet, B.; Zandberg, W.F. Quantitating organoleptic volatile phenols in smoke-exposed Vitis vinifera berries. J. Agric. Food Chem. 2017, 65, 8418–8425. [Google Scholar] [CrossRef] [PubMed]
- Favell, J.W.; Noestheden, M.; Lyons, S.M.; Zandberg, W.F. Development and evaluation of a vineyard-based strategy to mitigate smoke-taint in wine grapes. J. Agric. Food Chem. 2019, 67, 14137–14142. [Google Scholar] [CrossRef] [PubMed]
- Noestheden, M.; Noyovitz, B.; Riordan-Short, S.; Dennis, E.G.; Zandberg, W.F. Smoke from simulated forest fire alters secondary metabolites in Vitis vinifera L. berries and wine. Planta 2018, 248, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Sarry, J.E.; Günata, Z. Plant and microbial glycoside hydrolases: Volatile release from glycosidic aroma precursors. Food Chem. 2004, 87, 509–521. [Google Scholar] [CrossRef]
- Hopkinson, S.M. The chemistry and biochemistry of phenolic glycosides. Q. Rev. Chem. Soc. 1969, 23, 98–124. [Google Scholar] [CrossRef]
- Kelly, D.; Zerihun, A.; Hayasaka, Y.; Gibberd, M. Winemaking practice affects the extraction of smoke-borne phenols from grapes into wines. Aust. J. Grape Wine Res. 2014. [Google Scholar] [CrossRef]
- Fudge, A.L.; Ristic, R.; Wollan, D.; Wilkinson, K.L. Amelioration of smoke taint in wine by reverse osmosis and solid phase adsorption. Aust. J. Grape Wine Res. 2011, 17, 41–48. [Google Scholar] [CrossRef]
- Milheiro, J.; Filipe-Ribeiro, L.; Cosme, F.; Nunes, F.M. A simple, cheap and reliable method for control of 4-ethylphenol and 4-ethylguaiacol in red wines. Screening of fining agents for reducing volatile phenols levels in red wines. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1041–1042, 183–190. [Google Scholar] [CrossRef]
- Gil-Muñoz, R.; Jiménez-Martínez, M.D.; Bautista-Ortín, A.B.; Gómez-Plaza, E. Effect of the use of purified grape pomace as a fining agent on the volatile composition of monastrell wines. Molecules 2019, 24, 2423. [Google Scholar] [CrossRef] [Green Version]
- Fudge, A.L.; Schiettecatte, M.; Ristic, R.; Hayasaka, Y.; Wilkinson, K.L. Amelioration of smoke taint in wine by treatment with commercial fining agents. Aust. J. Grape Wine Res. 2012, 18, 302–307. [Google Scholar] [CrossRef]
- Ristic, R.; Van Der Hulst, L.; Capone, D.L.; Wilkinson, K.L. Impact of bottle aging on smoke-tainted wines from different grape cultivars. J. Agric. Food Chem. 2017, 65, 4146–4152. [Google Scholar] [CrossRef]
- Ristic, R.; Osidacz, P.; Pinchbeck, K.A.; Hayasaka, Y.; Fudge, A.L.; Wilkinson, K.L. The effect of winemaking techniques on the intensity of smoke taint in wine. Aust. J. Grape Wine Res. 2011, 17. [Google Scholar] [CrossRef]
- Izquierdo-Cañas, P.M.; González Viñas, M.A.; Mena-Morales, A.; Pérez Navarro, J.; García-Romero, E.; Marchante-Cuevas, L.; Gómez-Alonso, S.; Sánchez-Palomo, E. Effect of fermentation temperature on volatile compounds of Petit Verdot red wines from the Spanish region of La Mancha (central-southeastern Spain). Eur. Food Res. Technol. 2020, 246, 1153–1165. [Google Scholar] [CrossRef]
- Botondi, R.; De Sanctis, F.; Moscatelli, N.; Vettraino, A.M.; Catelli, C.; Mencarelli, F. Ozone fumigation postharvest treatment for the quality of wine grape. Food Chem. 2015, 188, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Antolini, A.; Forniti, R.; Modesti, M.; Bellincontro, A.; Catelli, C.; Mencarelli, F. First application of ozone postharvest fumigation to remove smoke taint from grapes. Ozone Sci. Eng. 2020. [Google Scholar] [CrossRef]
- Modesti, M.; Szeto, C.; Ristic, R.; Jiang, W.W.; Culbert, J.; Bindon, K.; Catelli, C.; Mencarelli, F.; Tonutti, P.; Wilkinson, K. Potential mitigation of smoke taint in wines by post-harvest ozone treatment of grapes. Molecules 2021, 26, 1798. [Google Scholar] [CrossRef]
- Maicas, S.; Mateo, J.J. Hydrolysis of terpenyl glycosides in grape juice and other fruit juices: A review. Appl. Microbiol. Biotechnol. 2005, 67, 322–335. [Google Scholar] [CrossRef]
- Gunata, Z.; Bitteur, S.; Brillouet, J.M.; Bayonove, C.; Cordonnier, R. Sequential enzymic hydrolysis of potentially aromatic glycosides from grape. Carbohydr. Res. 1988, 184, 139–149. [Google Scholar] [CrossRef]
- Hernández, L.F.; Espinosa, J.C.; Fernández-González, M.; Briones, A. β-glucosidase activity in a Saccharomyces cerevisiae wine strain. Int. J. Food Microbiol. 2003, 80, 171–176. [Google Scholar] [CrossRef]
- Fia, G.; Giovani, G.; Rosi, I. Study of beta-glucosidase production by wine-related yeasts during alcoholic fermentation. A new rapid fluorimetric method to determine enzymatic activity. J. Appl. Microbiol. 2005, 99, 509–517. [Google Scholar] [CrossRef]
- Bisotto, A.; Julien, A.; Rigou, P.; Schneider, R.; Salmon, J.M. Evaluation of the inherent capacity of commercial yeast strains to release glycosidic aroma precursors from Muscat grape must. Aust. J. Grape Wine Res. 2015, 21, 194–199. [Google Scholar] [CrossRef]
- Romero-Cascales, I.; Ros-García, J.M.; López-Roca, J.M.; Gómez-Plaza, E. The effect of a commercial pectolytic enzyme on grape skin cell wall degradation and colour evolution during the maceration process. Food Chem. 2012, 130, 626–631. [Google Scholar] [CrossRef]
- Wanapu, C.; Sripunya, P.; Boonkerd, N. Selection of yeast strains containing β-glucosidase for improving wine aroma. J. Agric. Sci. Technol. B 2012, 2, 691–702. [Google Scholar]
- Noestheden, M.; Dennis, E.G.; Romero-Montalvo, E.; Dilabio, G.A.; Zandberg, W.F. Detailed characterization of glycosylated sensory-active volatile phenols in smoke-exposed grapes and wine. Food Chem. 2018, 259, 147–156. [Google Scholar] [CrossRef]
- Danilewicz, J.C. Mechanism of autoxidation of polyphenols and participation of sulfite in wine: Key role of iron. Am. J. Enol. Vitic. 2011, 62, 319–328. [Google Scholar] [CrossRef]
- De, K.K.; Timell, T.E. The acid hydrolysis of glycosides III: Hydrolysis of O-methylated glucosides and disaccharides. Carbohydr. Res. 1967, 4, 72–77. [Google Scholar] [CrossRef]
- Morgan, S.C.; Scholl, C.M.; Benson, N.L.; Stone, M.L.; Durall, D.M. Sulfur dioxide addition at crush alters Saccharomyces cerevisiae strain composition in spontaneous fermentations at two Canadian wineries. Int. J. Food Microbiol. 2017, 244, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Boulesteix, A.L.; Strimmer, K. Partial least squares: A versatile tool for the analysis of high-dimensional genomic data. Brief. Bioinform. 2007, 8, 32–44. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| VP-Glycoside | Atmosphere/Treatment 1,2,3 | |||||||
|---|---|---|---|---|---|---|---|---|
| Air/Not Filtered | Air/Filtered | Argon/Not Filtered | Argon/Filtered | |||||
| Week 0 | Week 12 | Week 0 | Week 12 | Week 0 | Week 12 | Week 0 | Week 12 | |
| 4-EP-Glc | 15.2 ± 0.70 | 17.3 ± 0.65 | 14.4 ± 0.77 | 16.6 ± 0.69 | 11.3 ± 0.23 | 13.5 ± 1.10 | 9.0 ± 3.39 | 14.6 ± 1.27 |
| 4-MG-Glc | - | - | - | - | - | - | - | - |
| Eug-Glc | 6.2 ± 0.61 | 9.2 ± 0.95 | 7.1 ± 0.60 | 7.6 ± 0.69 | 4.3 ± 0.24 | 4.8 ± 0.24 | 4.9 ± 3.06 | 8.0 ± 0.70 |
| Gua-Glc | 1.0 | 1.4 ± 0.19 | - | 1.2 | 1.1 | 1.6 ± 0.14 | - | 1.3 ± 0.18 |
| o-Cre-Glc | - | - | 1.2 | 1.3 ± 0.39 | - | - | - | - |
| p-Cre-Glc | 2.8 ± 0.27 | 4.4 ± 0.32 | 2.7 ± 0.17 | 3.1 ± 0.17 | 2.4 ± 0.06 | 3.1 ± 0.13 | 2.5 ± 1.08 | 3.3 ± 0.23 |
| Phe-Glc | - | - | - | - | - | - | - | - |
| Syr-Glc | 20.4 ± 0.92 | 23.9 ± 1.67 | 18.6 ± 0.72 | 28.5 ± 3.65 | 22.3 ± 0.47 | 24.1 ± 0.97 | 13.9 ± 3.01 | 29.3 ± 3.16 |
| Gua-Gent | - | - | - | - | n.a. | n.a. | ||
| Gua-Prim | - | - | - | - | n.a. | n.a. | ||
| Syr-Gent | 2.1 ± 0.44 | 3.1 ± 0.35 | 2.3 ± 0.13 | 3.3 ± 0.56 | n.a. | n.a. | ||
| VP-Glycoside | Atmosphere/Treatment 1,2 | |||
|---|---|---|---|---|
| Air/Not Filtered | Air/Filtered | Argon/Not Filtered | Argon/Filtered | |
| 4-EP-Glc | 70.3 ± 8.24 | 76.0 ± 3.64 | 77.1 ± 2.83 | 99.8 ± 4.88 |
| 4-MG-Glc | 84.8 ± 3.31 | 120.7 ± 7.65 | 121.3 ± 9.82 | 89.4 ± 12.45 |
| Eug-Glc | 84.8 ± 4.28 | 99.5 ± 8.89 | 109.4 ± 5.22 | 103.5 ± 8.46 |
| Gua-Glc | 93.9 ± 5.98 | 82.3 ± 4.89 | 112.0 ± 5.58 | 92.7 ± 7.72 |
| p-Cre-Glc | 72.5 ± 7.57 | 84.0 ± 4.23 | 75.0 ± 4.51 | 105.5 ± 5.54 |
| Syr-Glc | 90.9 ± 5.40 | 88.3 ± 4.13 | 80.3 ± 5.30 | 95.8 ± 5.74 |
| Gua-Gent | 102.2 ± 7.24 | 104.9 ± 7.07 | n.a. | n.a. |
| Gua-Prim | 106.9 ± 17.82 | 102.3 ± 6.21 | n.a. | n.a. |
| Syr-Gent | 102.4 ± 6.05 | 118.4 ± 3.72 | n.a. | n.a. |
| Strain | Fortified | Grapes | Concentration (ng/g) 1,2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4-MG | Gua | Phe | p-Cre | o-Cre | 4-EP | Syr | Eug | |||
| EC1118 | N | ctrl | - | 1.3 ± 0.02 c | 6.45 ± 0.49 ab | 3.0 ± 0.44 a | 3.5 ± 0.07 ab | - | 1.3 ± 0.03 b | 1.4 ± 0.04 ab |
| RC212 | N | ctrl | - | 4.4 ± 0.60 b | 5.1 ± 0.66 bc | 2.1 ± 0.14 ab | 3.3 ± 0.05 abc | - | 15.3 ± 2.36 a | - |
| BM45 | N | ctrl | - | 6.9 ± 0.54 a | 8.3 ± 0.16 a | 1.8 ± 0.01 b | 3.6 ± 0.17 a | - | 15.8 ± 2.20 a | 1.2 ± 0.03 ab |
| U43 | N | ctrl | - | 1.4 ± 0.02 c | 4.4 ± 0.48 cd | 1.7 ± 0.14 b | 3.4 ± 0.10 abc | - | 1.2 ± 0.10 b | 1.6 ± 0.04 a |
| ES488 | N | ctrl | - | 1.1 ± 0.04 c | 3.8 ± 0.03 cd | 1.5 ± 0.12 b | 3.0 ± 0.03 c | - | - | 1.1 ± 0.06 ab |
| Cru | N | ctrl | - | 1.2 ± 0.48 c | 2.8 ± 0.27 d | 1.5 ± 0.08 b | 3.1 ± 0.09 bc | - | - | 0.9 ± 0.33 bc |
| EC1118 | Y | ctrl | 5.1 ± 0.32 a | 58.1 ± 1.50 a | 39.4 ± 1.41 a | 3.5 ± 0.33 ab | 3.3 ± 0.09 a | 4.2 ± 0.34 ab | 7.2 ± 0.28 c | 5.2 ± 0.25 abc |
| RC212 | Y | ctrl | 3.6 ± 0.50 a | 67.4 ± 9.71 a | 6.9 ± 0.93 c | 3.1 ± 0.31 abc | 3.3 ± 0.23 a | 6.2 ± 1.09 a | 16.7 ± 2.42 b | 3.6 ± 0.56 cd |
| BM45 | Y | ctrl | 3.4 ± 0.59 a | 74.5 ± 9.2 a | 9.6 ± 0.48 c | 3.0 ± 0.16 abc | 3.4 ± 0.24 a | 5.4 ± 0.67 a | 28.6 ± 3.13 a | 3.5 ± 0.50 d |
| U43 | Y | ctrl | 4.4 ± 0.07 a | 67.5 ± 0.87 a | 37.5 ± 1.30 ab | 3.9 ± 0.05 a | 3.2 ± 0.05 a | 5.1 ± 0.09 a | 7.5 ± 0.15 c | 5.9 ± 0.04 a |
| ES488 | Y | ctrl | 3.7 ± 0.11 a | 53.8 ± 5.67 a | 29.9 ± 3.59 b | 2.5 ± 0.13 bc | 3.0 ± 0.12 a | 2.2 ± 0.19 b | 5.7 ± 0.64 c | 4.3 ± 0.23 bcd |
| Cru | Y | ctrl | 5.2 ± 0.49 a | 57.4 ± 4.43 a | 30.3 ± 2.34 ab | 2.5 ± 0.02 c | 3.1 ± 0.14 a | 2.4 ± 0.32 b | 5.8 ± 0.35 c | 5.8 ± 0.16 ab |
| EC1118 | N | smoked | 4.7 ± 0.13 a | 13.6 ± 1.60 ab | 29.67 ± 1.22 a | 8.4 ± 0.62 ab | 10.0 ± 0.21 a | 0.5 ± 0.27 a | 4.8 ± 1.90 b | 1.2 ± 0.04 a |
| RC212 | N | smoked | 5.4 ± 0.23 a | 16.3 ± 0.5 ab | 32.9 ± 1.01 a | 9.8 ± 0.29 ab | 11.3 ± 0.35 a | 0.5 ± 0.33 a | 19.7 ± 0.85 a | 0.8 ± 0.30 a |
| BM45 | N | smoked | 5.3 ± 0.39 a | 17.0 ± 0.54 a | 28.3 ± 1.20 ab | 8.7 ± 0.46 ab | 11.5 ± 0.67 a | 0.5 ± 0.30 a | 15.7 ± 1.30 a | 0.5 ± 0.33 a |
| U43 | N | smoked | 5.9 ± 0.42 a | 16.3 ± 1.23 ab | 31.1 ± 1.13 a | 10.9 ± 0.82 a | 11.2 ± 0.50 a | 0.8 ± 0.29 a | 1.8 ± 0.05 b | 1.6 ± 0.09 a |
| ES488 | N | smoked | 5.2 ± 0.39 a | 12.5 ± 0.77 b | 27.9 ± 2.02 ab | 10.0 ± 0.83 ab | 10.8 ± 0.67 a | 0.5 ± 0.30 a | - | 1.0 ± 0.41 a |
| Cru | N | smoked | 4.8 ± 0.10 a | 12.6 ± 0.18 ab | 23.1 ± 0.84 b | 7.9 ± 0.34 b | 9.5 ± 0.22 a | - | - | 1.3 ± 0.05 a |
| EC1118 | Y | smoked | 9.34 ± 0.72 a | 71.6 ± 2.15 a | 62.3 ± 1.60 a | 9.7 ± 0.37 a | 11.2 ± 0.47 a | 5.0 ± 0.29 ab | 6.4 ± 0.09 c | 5.3 ± 0.41 b |
| RC212 | Y | smoked | 9.1 ± 0.92 a | 88.2 ±12.49 a | 35.5 ± 3.23 b | 11.5 ± 1.73 a | 12.6 ± 0.82 a | 6.7 ± 1.92 ab | 29.3 ± 2.12 a | 4.2 ± 0.81 b |
| BM45 | Y | smoked | 8.1 ± 0.77 a | 98.7 ± 5.71 a | 34.7 ± 3.10 b | 10.9 ± 0.98 a | 11.6 ± 0.83 a | 7.9 ± 0.59 a | 22.9 ± 1.01 b | 4.2 ± 0.41 b |
| U43 | Y | smoked | 10.7 ± 0.26 a | 99.4 ± 3.97 a | 63.6 ± 2.19 a | 11.5 ± 0.07 a | 10.3 ± 0.26 a | 5.9 ± 0.27 ab | 9.0 ± 0.54 c | 8.7 ± 0.58 a |
| ES488 | Y | smoked | 8.9 ± 0.47 a | 82.8 ± 2.34 a | 62.8 ± 1.20 a | 12.2 ± 0.26 a | 11.7 ± 0.57 a | 4.8 ± 0.17 ab | 7.2 ± 0.10 c | 5.5 ± 0.29 b |
| Cru | Y | smoked | 10.1 ± 0.71 a | 88.1 ± 4.07 a | 58.9 ± 2.11 a | 8.8 ± 0.91 a | 10.2 ± 0.30 a | 3.5 ± 0.46 b | 7.5 ± 0.33 c | 8.1 ± 0.56 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitmore, B.A.; McCann, S.E.; Noestheden, M.; Dennis, E.G.; Lyons, S.M.; Durall, D.M.; Zandberg, W.F. Glycosidically-Bound Volatile Phenols Linked to Smoke Taint: Stability during Fermentation with Different Yeasts and in Finished Wine. Molecules 2021, 26, 4519. https://doi.org/10.3390/molecules26154519
Whitmore BA, McCann SE, Noestheden M, Dennis EG, Lyons SM, Durall DM, Zandberg WF. Glycosidically-Bound Volatile Phenols Linked to Smoke Taint: Stability during Fermentation with Different Yeasts and in Finished Wine. Molecules. 2021; 26(15):4519. https://doi.org/10.3390/molecules26154519
Chicago/Turabian StyleWhitmore, Brandon A., Stephanie E. McCann, Matthew Noestheden, Eric G. Dennis, Sarah M. Lyons, Daniel M. Durall, and Wesley F. Zandberg. 2021. "Glycosidically-Bound Volatile Phenols Linked to Smoke Taint: Stability during Fermentation with Different Yeasts and in Finished Wine" Molecules 26, no. 15: 4519. https://doi.org/10.3390/molecules26154519
APA StyleWhitmore, B. A., McCann, S. E., Noestheden, M., Dennis, E. G., Lyons, S. M., Durall, D. M., & Zandberg, W. F. (2021). Glycosidically-Bound Volatile Phenols Linked to Smoke Taint: Stability during Fermentation with Different Yeasts and in Finished Wine. Molecules, 26(15), 4519. https://doi.org/10.3390/molecules26154519