
Honeybee Pollen Extracts Reduce Oxidative Stress and Steatosis in Hepatic Cells

 ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Botanical Origin, Content of Phenolic Compounds, Flavonoid Compounds, Quercetin Concentration, ORAC-PGR, and FRAP Values of Honeybee Pollen
2.2. Phenolic Acids of Honeybee Pollen Extracts (BPEs) Determined by HPLC-DAD
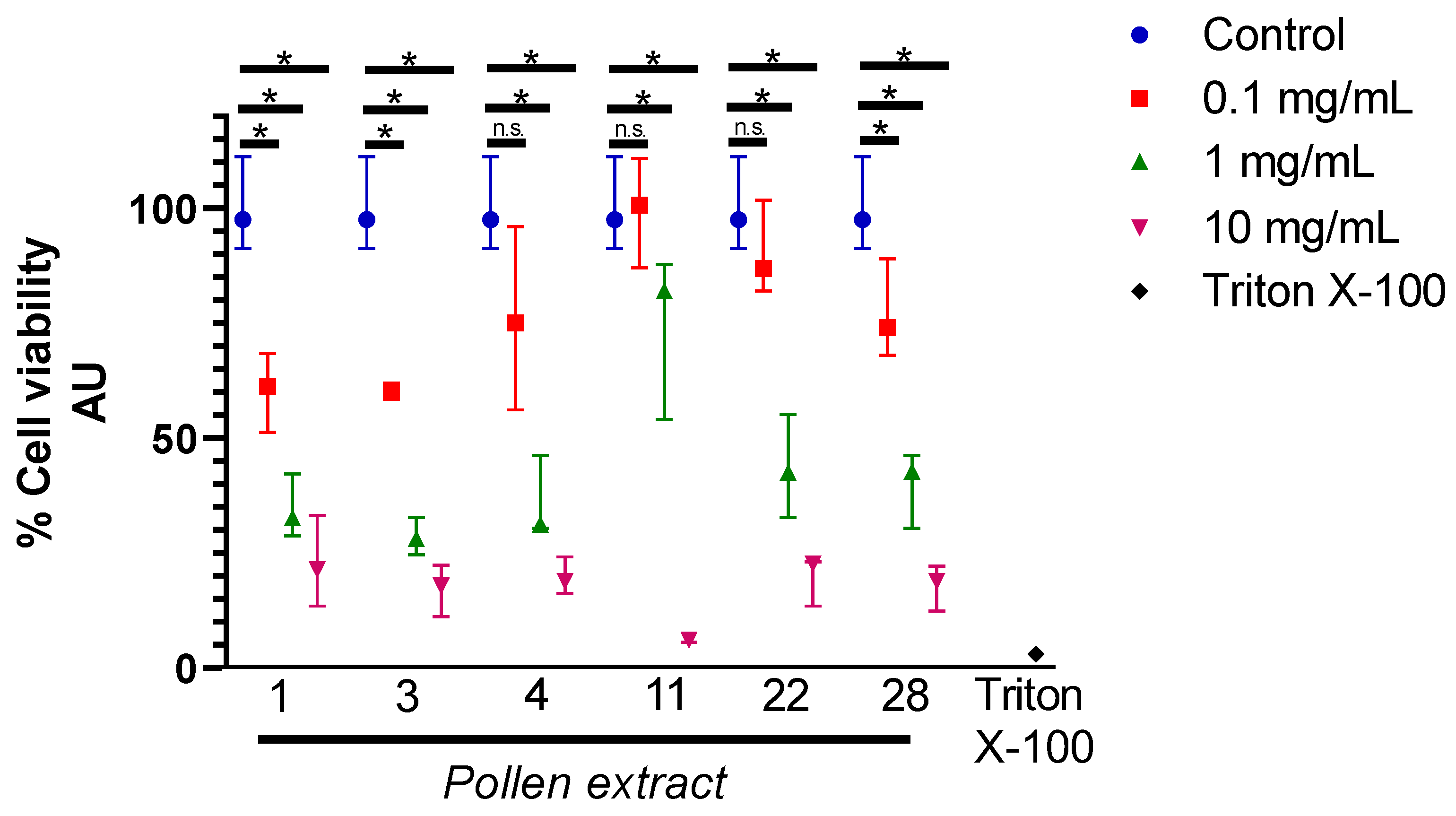
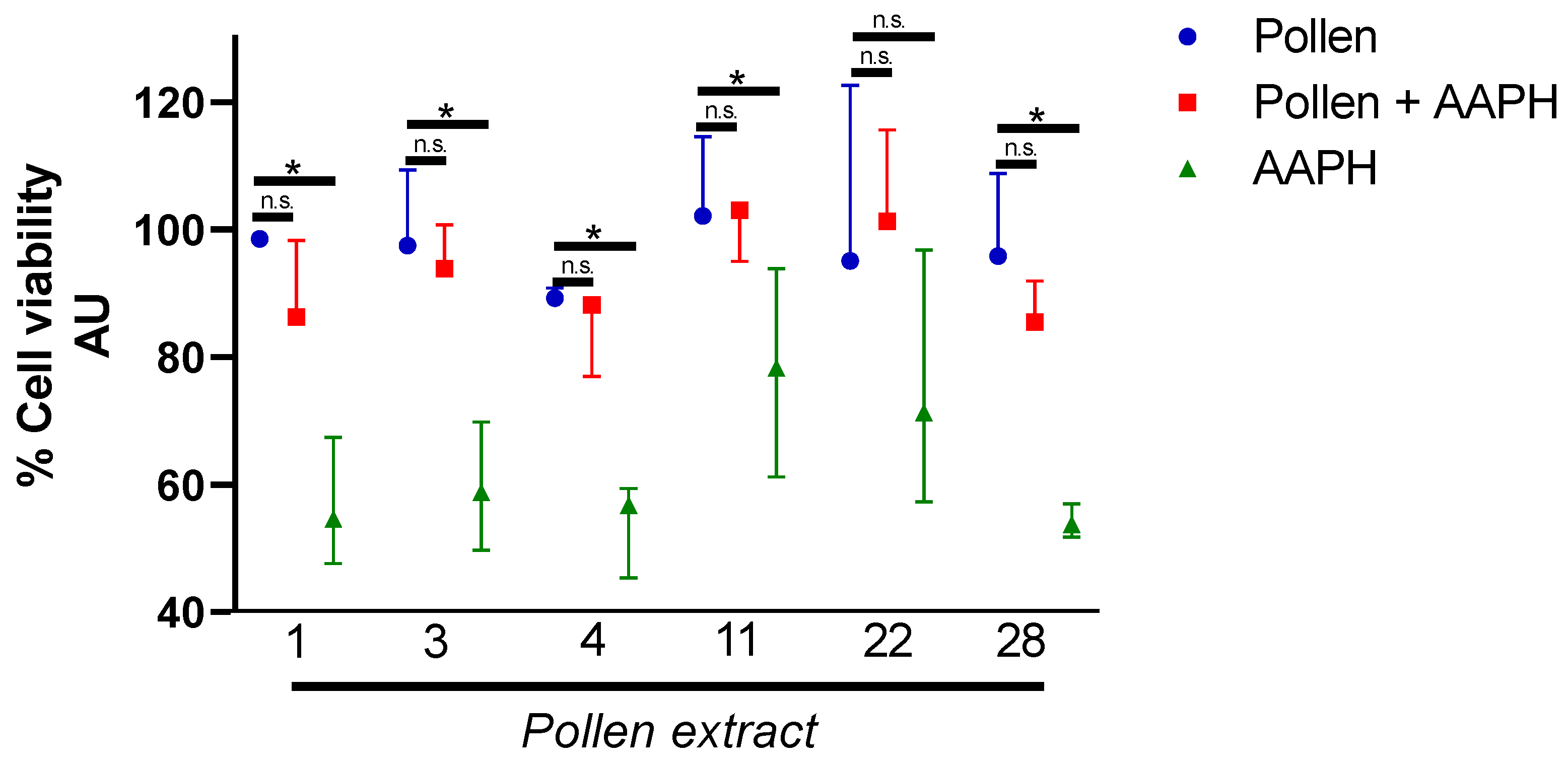
2.3. Cytotoxicity and Hepatoprotective Activity
3. Materials and Methods
3.1. Honeybee Pollen Samples
3.2. Extract Preparation and Phenolic Characterization
3.3. Antioxidant Capacity Assay
3.4. HPLC-DAD Analysis
3.5. Cytotoxicity and Hepatoprotective Activity In Vitro
3.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, L.A.; Harmsen, S.; St Sauver, J.L.; Charatcharoenwitthaya, P.; Enders, F.B.; Therneau, T.; Angulo, P. Nonalcoholic fatty liver disease increases risk of death among patients with diabetes: A community-based cohort study. Am. J. Gastroenterol. 2010, 105, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, J.; Sun, P.; Yi, R.; Han, X.; Zhao, X. Raw Bowl Tea (Tuocha) Polyphenol Prevention of Nonalcoholic Fatty Liver Disease by Regulating Intestinal Function in Mice. Biomolecules 2019, 9, 435. [Google Scholar] [CrossRef] [PubMed]
- Schwenger, K.J.P.; Allard, J.P. Clinical approaches to non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 1712–1723. [Google Scholar] [CrossRef] [PubMed]
- Masarone, M.; Rosato, V.; Dallio, M.; Gravina, A.G.; Aglitti, A.; Loguercio, C.; Federico, A.; Persico, M. Role of Oxidative Stress in Pathophysiology of Nonalcoholic Fatty Liver Disease. Oxid Med. Cell Longev. 2018, 2018, 9547613. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Di Giacomo, M.; Zara, V. Antioxidant dietary approach in treatment of fatty liver: New insights and updates. World J. Gastroenterol. 2017, 23, 4146–4157. [Google Scholar] [CrossRef]
- Sun, X.; Seidman, J.S.; Zhao, P.; Troutman, T.D.; Spann, N.J.; Que, X.; Zhou, F.; Liao, Z.; Pasillas, M.; Yang, X.; et al. Neutralization of Oxidized Phospholipids Ameliorates Non-alcoholic Steatohepatitis. Cell Metab. 2020, 31, 189–206.e8. [Google Scholar] [CrossRef]
- Kitade, H.; Chen, G.; Ni, Y.; Ota, T. Nonalcoholic Fatty Liver Disease and Insulin Resistance: New Insights and Potential New Treatments. Nutrients 2017, 9, 387. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Zhang, P.Y. Polyphenols in Health and Disease. Cell Biochem. Biophys. 2015, 73, 649–664. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef]
- Li, Q.; Wang, K.; Marcucci, M.; Christine, A.; Sawaya, H.; Hu, L.; Xue, X.; Wu, L.-M.; Hu, F.-L. Nutrient-rich bee pollen: A treasure trove of active natural metabolites. J. Funct. Foods 2018, 49, 472. [Google Scholar] [CrossRef]
- Rzepecka-Stojko, A.; Kabała-Dzik, A.; Kubina, R.; Jasik, K.; Kajor, M.; Wrześniok, D.; Stojko, J. Protective Effect of Polyphenol-Rich Extract from Bee Pollen in a High-Fat Diet. Molecules 2018, 23, 805. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Chen, S.; Liu, X.; Zhao, H.; Cao, W. Impact of SchisandraChinensis Bee Pollen on Nonalcoholic Fatty Liver Disease and Gut Microbiota in HighFat Diet Induced Obese Mice. Nutrients 2019, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Kostić, A.Ž.; Milinčić, D.D.; Barać, M.B.; Ali Shariati, M.; Tešić, Ž.L.; Pešić, M.B. The Application of Pollen as a Functional Food and Feed Ingredient-The Present and Perspectives. Biomolecules 2020, 10, 84. [Google Scholar]
- Komosinska-Vassev, K.; Olczyk, P.; Kaźmierczak, J.; Mencner, L.; Olczyk, K. Bee Pollen: Chemical Composition and Therapeutic Application. Evid. Based Complement. Altern. Med. eCAM 2015, 2015, 1–6. [Google Scholar] [CrossRef]
- Bridi, R.; Atala, E.; Pizarro, P.N.; Montenegro, G. Honeybee Pollen Load: Phenolic Composition and Antimicrobial Activity and Antioxidant Capacity. J. Nat. Prod. 2019, 82, 559–565. [Google Scholar] [CrossRef]
- Montenegro, G.; Gómez, M.; Díaz-Forestier, J.; Pizarro, R. Aplicación de la Norma Chilena Oficial de denominación de origen botánico de la miel para la caracterización de la producción apícola. Cienc. Investig. Agrar. 2008, 35, 181–190. [Google Scholar] [CrossRef]
- Levy, G.; Cohen, M.; Nahmias, Y. In Vitro Cell Culture Models of Hepatic Steatosis. Methods Mol. Biol. 2015, 1250, 377–390. [Google Scholar]
- Alkhatatbeh, M.J.; Lincz, L.F.; Thorne, R.F. Low simvastatin concentrations reduce oleic acid-induced steatosis in HepG(2) cells: An in vitro model of non-alcoholic fatty liver disease. Exp. Ther. Med. 2016, 11, 1487–1492. [Google Scholar] [CrossRef]
- Saha, P.; Talukdar, A.D.; Nath, R.; Sarker, S.D.; Nahar, L.; Sahu, J.; Choudhury, M.D. Role of Natural Phenolics in Hepatoprotection: A Mechanistic Review and Analysis of Regulatory Network of Associated Genes. Front. Pharmacol. 2019, 10, 509. [Google Scholar] [CrossRef]
- Sen, S.; Chakraborty, R. Chapter 19—Herbs, Gastrointestinal Protection, and Oxidative Stress. In Gastrointestinal Tissue; Gracia-Sancho, J., Salvadó, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 259–274. [Google Scholar]
- Chen, S.; Zhao, X.; Wan, J.; Ran, L.; Qin, Y.; Wang, X.; Gao, Y.; Shu, F.; Zhang, Y.; Liu, P.; et al. Dihydromyricetin improves glucose and lipid metabolism and exerts anti-inflammatory effects in nonalcoholic fatty liver disease: A randomized controlled trial. Pharmacol Res. 2015, 99, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; An, B.; Yu, Q.; Cao, Y.; Liu, Y.; Li, S. The hepatoprotective effect of myricetin against lipopolysaccharide and D-galactosamine-induced fulminant hepatitis. Int. J. Biol. Macromol. 2020, 155, 1092–1104. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Heusser, C.J.; Moar, N.T. Pollen and spores of Chile: Modern types of the Pteridophyta, Gymnospermae, and Angiospermae. N. Z. J. Bot. 1973, 11, 389–391. [Google Scholar] [CrossRef]
- Marticorena, C. Contribución a la estadística de la flora vascular de Chile. Gayana Bot 1990, 47, 8. [Google Scholar]
- Rodriguez-Gonzalez, I.; Ortega-Toro, R.; Díaz-Moreno, A. Influence of microwave- and ultrasound-assisted extraction on bioactive compounds from pollen. Contemp. Eng. Sci. 2018, 11, 1669–1676. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Madanayake, T.W.; Fidler, T.P.; Fresquez, T.M.; Bajaj, N.; Rowland, A.M. Cytochrome P450 2S1 depletion enhances cell proliferation and migration in bronchial epithelial cells, in part, through modulation of prostaglandin E(2) synthesis. Drug Metab. Dispos. 2012, 40, 2119–2125. [Google Scholar] [CrossRef]
- Rampersad, S.N. Multiple applications of Alamar Blue as an indicator of metabolic function and cellular health in cell viability bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A. Use of multiple assay endpoints to investigate the effects of incubation time, dose of toxin, and plating density in cell-based cytotoxicity assays. Assay Drug Dev. Technol. 2004, 2, 51–62. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Predominant Plant Species | Total Phenols | Flavonoids | FRAP | ORAC-FL | [QE] | ||
|---|---|---|---|---|---|---|---|---|
| First | Second | Third | (mg GAE/100 g Bee Pollen) | (mg QE/100 g Bee Pollen) | (µmol TE/g Bee Pollen) | (µmol TE/g Bee Pollen) | (mg/100 g Bee Pollen) | |
| 1 * | Eschscholzia californica | Adesmia arborea | Taraxacum officinale | 1448 ± 116 | 252 ± 18 | 194 ± 7 | 492 ± 25 | 60.32 ± 0.34 |
| 2 | Eschscholzia californica | Brassica rapa | Eucalyptus sp. | 1301 ± 12 | 504 ± 21 | 111 ± 4 | 312 ± 22 | 99.37 ± 0.18 |
| 3 * | Brassica rapa | Cryptocarya alba | Sophora macrocarpa | 1141 ± 30 | 479 ± 6 | 74 ± 1 | 412 ± 89 | 98.70 ± 18.45 |
| 4 * | Brassica rapa | Cryptocarya alba | Anthemis cotula | 954 ± 83 | 304 ± 8 | 78 ± 15 | 307 ± 61 | 66.99 ± 2.82 |
| 5 | Brassica rapa | Adesmia arborea | Colliguaja odorifera | 920 ± 38 | 261 ± 14 | 101 ± 1 | 224 ± 27 | 16.04 ± 0.08 |
| 6 | Acacia caven | Eschscholzia californica | Acacia caven | 848 ± 2 | 273 ± 12 | 70 ± 4 | 253 ± 8 | 84.22 ± 20.77 |
| 7 | Brassica rapa | Cryptocarya alba | Azara sp. | 8379 ± 55 | 346 ± 8 | 76 ± 15 | 398 ± 34 | 65.13 ± 0.76 |
| 8 | Brassica rapa | Cryptocarya alba | Azara sp. | 780 ± 17 | 326 ± 20 | 91 ± 16 | 397 ± 40 | 39.49 ± 0.75 |
| 9 | Brassica rapa | Colliguaja odorifera | Amomyrtus luma | 767 ± 22 | 168 ± 10 | 86 ± 4 | 371 ± 36 | 133.68 ± 1.10 |
| 10 | Brassica rapa | Eschscholzia californica | Cryptocarya alba | 767 ± 109 | 205 ± 7 | 107 ± 6 | 350 ± 38 | 4.55 ± 0.52 |
| 11 * | Azara celastrina | Mix | Cryptocarya alba | 759 ± 7 | 331 ± 26 | 60 ± 2 | 183 ± 11 | 37.44 ± 0.54 |
| 12 | Cryptocarya alba | Brassica rapa | Eschscholzia californica | 758 ± 72 | 251 ± 12 | 84 ± 8 | 282 ± 68 | 25.05 ± 0.33 |
| 13 | Brassica rapa | Colliguaja odorifera | Eschscholzia californica | 744 ± 15 | 172 ± 9 | 96 ± 9 | 338 ± 27 | 175.22 ± 0.96 |
| 14 | Brassica rapa | Adesmia arborea | Cryptocarya alba | 725 ± 44 | 337 ± 14 | 66 ± 3 | 197 ± 35 | 296.08 ± 3.43 |
| 15 | Brassica rapa | Eucalyptus sp. | Raphanus sativus | 717 ± 15 | 149 ± 16 | 98 ± 1 | 371 ± 45 | 115.74 ± 0.99 |
| 16 | Brassica rapa | Cryptocarya alba | Hypochaeris radicata | 715 ± 52 | 186 ± 14 | 80 ± 17 | 281 ± 87 | 162.38 ± 0.51 |
| 17 | Brassica rapa | Cryptocarya alba | Schinus latifolius | 588 ± 12 | 150 ± 8 | 66 ± 13 | 203 ± 39 | 121.87 ± 0.74 |
| 18 | Eschscholzia californica | Brassica rapa | Schinus latifolius | 580 ± 5 | 165 ± 17 | 56 ± 3 | 231 ± 47 | 145.98 ± 1.18 |
| 19 | Cryptocarya alba | Brassica rapa | Eschscholzia californica | 540 ± 21 | 88 ± 8 | 61 ± 4 | 277 ± 99 | 409.94 ± 1.79 |
| 20 | Eschscholzia californica | Cryptocarya alba | Anthemis cotula | 536 ± 10 | 260 ± 6 | 35 ± 9 | 277 ± 129 | 22.34 ± 0.1 |
| 21 | Brassica rapa | Lythrum hyssopifolia | Robinia pseudoacacia | 518 ± 51 | 102 ± 5 | 54 ± 7 | 185 ± 74 | 63.19 ± 0.36 |
| 22 * | Brassica rapa | Eschscholzia californica | Cactaceae | 368 ± 19 | 75 ± 7 | 51 ± 2 | 109 ± 49 | 6.59 ± 0.19 |
| 23 | Cryptocarya alba | Eschscholzia californica | Taraxacum officinale | 364 ± 14 | 176 ± 18 | 38 ± 6 | 166 ± 45 | 11.2 ± 0.14 |
| 24 | Brassica rapa | Schinus latifolius | Azara celastina | 350 ± 41 | 122 ± 4 | 41 ± 3 | 158 ± 40 | 5.24 ± 0.45 |
| 25 | Brassica rapa | Cryptocarya alba | Azara sp. | 344 ± 27 | 128 ± 6 | 39 ± 10 | 191 ± 28 | 20.61 ± 0.36 |
| 26 | Cryptocarya alba | Eschscholzia californica | Malva sp. | 315 ± 15 | 156 ± 1 | 27 ± 2 | 216 ± 44 | 4.29 ± 0.45 |
| 27 | Cryptocarya alba | Eschscholzia californica | Anthemis cotula | 286 ± 34 | 153 ± 3 | 26 ± 1 | 242 ± 42 | 6.7 ± 0.18 |
| 28 * | Cryptocarya alba | Anthemis cotula | Baccharis linearis | 102 ± 20 | 62 ± 9 | 19 ± 2 | 150 ± 67 | 15.08 ± 0.3 |
| Samples | mg Phenolic Acids/100 g Pollen | ||||||
| Chlorogenic | Caffeic | Syringic | Coumaric | Sinapic | Ferulic | Cinnamic | |
| 1 | 0 | 0 | 6.62 ± 0.05 | 0.8 ± 0.12 | 0 | 6.76 ± 0.57 | 20.18 ± 0.1 |
| 3 | 0 | 0 | 1.77 ± 0.1 | 0.94 ± 0.02 | 0 | 15.34 ± 0.36 | 16.14 ± 0.21 |
| 4 | 0 | 0.93 ± 0.02 | 1.35 ± 0.02 | 0.41 ± 0.23 | 0.98 ± 0.42 | 6.38 ± 0.33 | 21.42 ± 0.31 |
| 11 | 15.92 ± 0.18 | 0 | 13.64 ± 0.13 | 0 | 16.7 ± 0.3 | 20.2 ± 0.29 | 10.15 ± 0.04 |
| 22 | 26.21 ± 0.37 | 5.67 ± 0.08 | 5.81 ± 0.2 | 0.2 ± 0.01 | 5.36 ± 0.22 | 1.03 ± 0.51 | 2.81 ± 0.91 |
| 28 | 16.76 ± 1.16 | 0 | 2.21 ± 0.4 | 0 | 19.05 ± 5.78 | 0 | 5.42 ± 0.23 |
| Samples | mg Flavonoids/ 100 g Pollen | ||||||
| Epicatechin | Rutin | Myricetin | Quercetin | Apigenin | Rhamnetin | Catechin | |
| 1 | 7.95 ± 0.1 | 13.98 ± 0.07 | 775.89 ± 1.87 | 60.32 ± 0.34 | 34.62 ± 2.71 | 0 | 0 |
| 3 | 0 | 7.19 ± 0.18 | 185.65 ± 1.43 | 98.7 ± 18.45 | 767.24 ± 83.92 | 0 | 0 |
| 4 | 38.42 ± 0.14 | 36.2 ± 0.68 | 121.41 ± 3.31 | 66.99 ± 2.82 | 0 | 0 | 0 |
| 11 | 21.4 ± 0.05 | 74.25 ± 2.93 | 96.46 ± 0.41 | 37.44 ± 0.54 | 21.05 ± 2.56 | 13.35 ± 2.88 | 15.8 ± 0.34 |
| 22 | 0 | 2.74 ± 1.43 | 28.51 ± 0.95 | 6.59 ± 0.19 | 13.09 ± 0.06 | 0 | 0 |
| 28 | 0 | 17.57 ± 0.4 | 35.49 ± 3.08 | 15.08 ± 0.3 | 6.98 ± 0.63 | 80.37 ± 3.19 | 17.96 ± 0.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyarzún, J.E.; Andia, M.E.; Uribe, S.; Núñez Pizarro, P.; Núñez, G.; Montenegro, G.; Bridi, R. Honeybee Pollen Extracts Reduce Oxidative Stress and Steatosis in Hepatic Cells. Molecules 2021, 26, 6. https://doi.org/10.3390/molecules26010006
Oyarzún JE, Andia ME, Uribe S, Núñez Pizarro P, Núñez G, Montenegro G, Bridi R. Honeybee Pollen Extracts Reduce Oxidative Stress and Steatosis in Hepatic Cells. Molecules. 2021; 26(1):6. https://doi.org/10.3390/molecules26010006
Chicago/Turabian StyleOyarzún, Juan Esteban, Marcelo E. Andia, Sergio Uribe, Paula Núñez Pizarro, Gabriel Núñez, Gloria Montenegro, and Raquel Bridi. 2021. "Honeybee Pollen Extracts Reduce Oxidative Stress and Steatosis in Hepatic Cells" Molecules 26, no. 1: 6. https://doi.org/10.3390/molecules26010006
APA StyleOyarzún, J. E., Andia, M. E., Uribe, S., Núñez Pizarro, P., Núñez, G., Montenegro, G., & Bridi, R. (2021). Honeybee Pollen Extracts Reduce Oxidative Stress and Steatosis in Hepatic Cells. Molecules, 26(1), 6. https://doi.org/10.3390/molecules26010006