
Spectrofluorometric Analysis of Autofluorescing Components of Crude Serum from a Rat Liver Model of Ischemia and Reperfusion

 ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
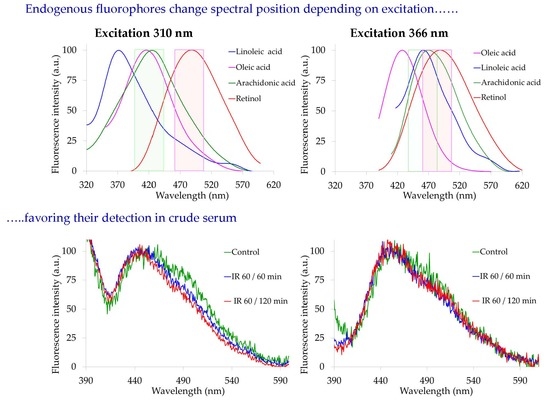
2.1. Spectrofluorometric Analysis of Pure Compounds
2.2. Liver Injury Biochemical Assays
2.3. Spectrofluorometric Analysis of Serum
3. Discussion
4. Materials and Methods
4.1. Pure Compounds and AF Spectrofluorometric and Fitting Analysis
4.2. Animal Models
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Croce, A.C.; Bottiroli, G. Autofluorescence spectroscopy and imaging: A tool for biomedical research and diagnosis. Eur. J. Histochem. 2014, 58, 2461. [Google Scholar] [CrossRef]
- Nazeer, S.S.; Saraswathy, A.; Shenoy, S.J.; Jayasree, R.S. Fluorescence spectroscopy as an efficient tool for staging the degree of liver fibrosis: An in vivo comparison with MRI. Sci. Rep. 2018, 8, 10967. [Google Scholar] [CrossRef]
- Favreau, P.F.; Deal, J.A.; Harris, B.; Weber, D.S.; Rich, T.C.; Leavesley, S.J. Label-free spectroscopic tissue characterization using fluorescence excitation-scanning spectral imaging. J. Biophotonics 2019, 13, e201900183. [Google Scholar] [CrossRef]
- Croce, A.C.; Ferrigno, A.; Bottiroli, G.; Vairetti, M. Autofluorescence-based optical biopsy: An effective diagnostic tool in hepatology. Liver Int. 2018, 38, 1160–1174. [Google Scholar] [CrossRef]
- Pickett-Blakely, O.; Young, K.; Carr, R.M. Micronutrients in Nonalcoholic Fatty Liver Disease Pathogenesis. CMGH 2018, 6, 451–462. [Google Scholar] [CrossRef]
- Saeed, A.; Dullaart, R.P.F.; Schreuder, T.C.M.A.; Blokzijl, H.; Faber, K.N. Disturbed vitamin A metabolism in non-alcoholic fatty liver disease (NAFLD). Nutrients 2018, 10, 29. [Google Scholar] [CrossRef]
- Chance, B. Spectra and reaction kinetics of respiratory pigments of homogenized and intact cells. Nature 1952, 169, 215–221. [Google Scholar] [CrossRef]
- Croce, A.C.; Ferrigno, A.; Di Pasqua, L.G.; Berardo, C.; Bottiroli, G.; Vairetti, M. NAD(P)H and Flavin Autofluorescence Correlation with ATP in Rat Livers with Different Metabolic Steady-State Conditions. Photochem. Photobiol. 2017, 93, 1519–1524. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Heikal, A.A.; Webb, W.W. Two-photon fluorescence spectroscopy and microscopy of NAD(P)H and flavoprotein. Biophys. J. 2002, 82, 2811–2825. [Google Scholar] [CrossRef]
- Popper, H.; Gyorgy, P.; Goldblatt, H. Fluorescent material (ceroid) in experimental nutritional cirrhosis. Arch. Path. 1944, 37, 161–168. [Google Scholar]
- Seehafer, S.S.; Pearce, D.A. You say lipofuscin, we say ceroid: Defining autofluorescent storage material. Neurobiol. Aging 2006, 27, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Liu, Y.; Chen, H.-C.; Chiou, L.-L.; Huang, G.-T.; Lo, W.; Dong, C.-Y. Optical biopsy of liver fibrosis by use of multiphoton microscopy. Opt. Lett. 2004, 29, 2614–2616. [Google Scholar] [CrossRef] [PubMed]
- Croce, A.C.; Ferrigno, A.; Vairetti, M.; Bertone, R.; Freitas, I.; Bottiroli, G. Autofluorescence properties of isolated rat hepatocytes under different metabolic conditions. Photochem. Photobiol. Sci. 2004, 3, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Croce, A.C.; Ferrigno, A.; Santin, G.; Piccolini, V.M.; Bottiroli, G.; Vairetti, M. Autofluorescence of liver tissue and bile: Organ functionality monitoring during ischemia and reoxygenation. Lasers Surg. Med. 2014, 46, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, S.D.; Gilfor, D.; Farber, J.L. Alterations in the metabolism of lipids in ischemia of the liver and kidney. J. Lipid Res. 1985, 26, 726–734. [Google Scholar]
- Aït-Kaddour, A.; Thomas, A.; Mardon, J.; Jacquot, S.; Ferlay, A.; Gruffat, D. Potential of fluorescence spectroscopy to predict fatty acid composition of beef. Meat Sci. 2016, 113, 124–131. [Google Scholar] [CrossRef]
- Croce, A.C.; Ferrigno, A.; Di Pasqua, L.G.; Berardo, C.; Mannucci, B.; Bottiroli, G.; Vairetti, M. Fluorescing fatty acids in rat fatty liver models. J. Biophotonics 2017, 10, 905–910. [Google Scholar] [CrossRef]
- Sanches, S.C.L.; Ramalho, L.N.Z.; Augusto, M.J.; da Silva, D.M.; Ramalho, F.S. Nonalcoholic Steatohepatitis: A Search for Factual Animal Models. Biomed Res. Int. 2015, 574832. [Google Scholar] [CrossRef]
- Quinn, K.P.; Sridharan, G.V.; Hayden, R.S.; Kaplan, D.L.; Lee, K.; Georgakoudi, I. Quantitative metabolic imaging using endogenous fluorescence to detect stem cell differentiation. Sci. Rep. 2013, 3, 3432. [Google Scholar] [CrossRef]
- Croce, A.C.; Di Pasqua, L.G.; Berardo, C.; Ferrigno, A.; Siciliano, V.; Bottiroli, G.; Vairetti, M. Real time optical biopsy: Serum fatty acids profiling in a rat model of nonalcoholic steatohepatitis. Lasers Surg. Med. 2017, 49, E5. [Google Scholar]
- Sahebkar, A.; Sancho, E.; Abelló, D.; Camps, J.; Joven, J. Novel circulating biomarkers for non-alcoholic fatty liver disease: A systematic review. J. Cell. Physiol. 2018, 233, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.; Gilroy, D.W. Lipid Mediators in Inflammation. Microbiol. Spectr. 2016, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Fatty acids and inflammation: The cutting edge between food and pharma. Eur. J. Pharm. 2011, 668, S50–S58. [Google Scholar] [CrossRef] [PubMed]
- Hall, Z.; Bond, N.J.; Ashmore, T.; Sanders, F.; Ament, Z.; Wang, X.; Murray, A.J.; Bellafante, E.; Virtue, S.; Vidal-Puig, A.; et al. Lipid zonation and phospholipid remodeling in nonalcoholic fatty liver disease. Hepatology 2017, 65, 1165–1180. [Google Scholar] [CrossRef]
- Tam, V.C. Lipidomic profiling of bioactive lipids by mass spectrometry during microbial infections. Semin. Immunol. 2013, 25, 240–248. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Nimura, Y.; Nagino, M.; Bland, K.I.; Chaudry, I.H. Role of thromboxane in producing hepatic injury during hepatic stress. Arch. Surg. 2005, 140, 801–807. [Google Scholar] [CrossRef]
- Ross, A.C.; Zolfaghari, R.; Weisz, J. Vitamin A: Recent advances in the biotransformation, transport, and metabolism of retinoids. Curr. Opin. Gastroenterol. 2001, 17, 184–192. [Google Scholar] [CrossRef]
- Shirakami, Y.; Lee, S.A.; Clugston, R.D.; Blaner, W.S. Hepatic metabolism of retinoids and disease associations. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2012, 1821, 124–136. [Google Scholar] [CrossRef]
- Kane, M.A.; Napoli, J.L. Quantification of endogenous retinoids. Methods Mol. Biol. 2010, 652, 1–54. [Google Scholar]
- Craft, N.E. Innovative Approaches to Vitamin A Assessment. J. Nutr. 2001, 131, 1626S–1630S. [Google Scholar] [CrossRef]
- Driskell, W.J.; Hewett, J.S.; Bashor, M.M. Evaluation of a direct fluorometric method for determination of serum retinol. Clin. Chem. 1986, 32, 867–869. [Google Scholar] [CrossRef] [PubMed]
- Futterman, S.; Heller, J. The enhancement of fluorescence and the decreased susceptibility to enzymatic oxidation of retinol complexed with bovine serum albumin, beta-lactoglobulin, and the retinol-binding protein of human plasma. J. Biol. Chem. 1972, 247, 5168–5172. [Google Scholar] [PubMed]
- de Pee, S.; Dary, O. Biochemical Indicators of Vitamin A Deficiency: Serum Retinol and Serum Retinol Binding Protein. J. Nutr. 2002, 132, 2895S–2901S. [Google Scholar] [CrossRef] [PubMed]
- Ferrigno, A.; Rizzo, V.; Bianchi, A.; Di Pasqua, L.G.; Berardo, C.; Richelmi, P.; Vairetti, M. Changes in ADMA/DDAH Pathway after Hepatic Ischemia/Reperfusion Injury in Rats: The Role of Bile. Biomed Res. Int. 2014, 2014, 627434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Q.; Ding, N.; Zeng, Y.F.; Xiang, Y.Y.; Yang, M.W.; Hong, F.F.; Yang, S.L. New progress in roles of nitric oxide during hepatic ischemia reperfusion injury. World J. Gastroenterol. 2017, 23, 2505–2510. [Google Scholar] [CrossRef] [PubMed]
- Pratt, D.S.; Kaplan, M.M. Evaluation of abnormal liver-enzyme results in asymptomatic patients. N. Engl. J. Med. 2000, 342, 1266–1271. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B. An Introduction to Spectroscopy for Biochemists; Brown, S.B., Ed.; Academic Press: London, UK, 1980; pp. 14–65. [Google Scholar]
- Chaves, G.V.; Peres, W.A.F.; Gonçalves, J.C.; Ramalho, A. Vitamin A and retinol-binding protein deficiency among chronic liver disease patients. Nutrition 2015, 31, 664–668. [Google Scholar] [CrossRef]
- Abdelmagid, S.A.; Clarke, S.E.; Nielsen, D.E.; Badawi, A.; El-Sohemy, A.; Mutch, D.M.; Ma, D.W.L. Comprehensive profiling of plasma fatty acid concentrations in young healthy canadian adults. PLoS ONE 2015, 10, e0116195. [Google Scholar] [CrossRef]
- Loomba, R.; Quehenberger, O.; Armando, A.; Dennis, E.A. Polyunsaturated fatty acid metabolites as novel lipidomic biomarkers for noninvasive diagnosis of nonalcoholic steatohepatitis. J. Lipid Res. 2015, 56, 185–192. [Google Scholar] [CrossRef]
- Teoh, N.C.; Ajamieh, H.; Wong, H.J.; Croft, K.; Mori, T.; Allison, A.C.; Farrell, G.C. Microparticles mediate hepatic ischemia-reperfusion injury and are the targets of Diannexin (ASP8597). PLoS ONE 2014, 9, e104376. [Google Scholar] [CrossRef]
- Croce, A.C.; Ferrigno, A.; Bertone, V.; Piccolini, V.M.; Berardo, C.; Di Pasqua, L.G.; Rizzo, V.; Bottiroli, G.; Vairetti, M. Fatty liver oxidative events monitored by autofluorescence optical diagnosis: Comparison between subnormothermic machine perfusion and conventional cold storage preservation. Hepatol. Res. 2017, 47, 668–682. [Google Scholar] [CrossRef] [PubMed]
- Muller, X.; Schlegel, A.; Kron, P.; Eshmuminov, D.; Würdinger, M.; Meierhofer, D.; Clavien, P.A.; Dutkowski, P. Novel Real-time Prediction of Liver Graft Function During Hypothermic Oxygenated Machine Perfusion Before Liver Transplantation. Ann. Surg. 2019, 270, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Chorvat, D.; Kirchnerova, J.; Cagalinec, M.; Smolka, J.; Mateasik, A.; Chorvatova, A. Spectral unmixing of flavin autofluorescence components in cardiac myocytes. Biophys. J. 2005, 89, L55–l57. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, D.W. An Algorithm for Least-Squares Estimation of Nonlinear Parameters. J. Soc. Ind. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- Palladini, G.; Ferrigno, A.; Rizzo, V.; Boncompagni, E.; Richelmi, P.; Freitas, I.; Perlini, S.; Vairetti, M. Lobe-specific heterogeneity and matrix metalloproteinase activation after ischemia/reperfusion injury in rat livers. Toxicol. Pathol. 2012, 40, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Ferrigno, A.; Di Pasqua, L.G.; Berardo, C.; Siciliano, V.; Rizzo, V.; Adorini, L.; Richelmi, P.; Vairetti, M. The farnesoid X receptor agonist obeticholic acid upregulates biliary excretion of asymmetric dimethylarginine via MATE-1 during hepatic ischemia/reperfusion injury. PLoS ONE 2018, 13, e0191430. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | I/R 60 * | I/R 120 * | ||
|---|---|---|---|---|
| Serum | ||||
| AST | 254.63 ± 59.12 | 3503.23 ± 1049.05 | 10405.36 ± 1099.88 | |
| ALT | 69.17 ± 22.91 | 3911.07 ± 893.58 | 9403.55 ± 1055.46 | |
| ALP | 428.33 ± 20.47 | 613.28 ± 58.91 | 772.05 ± 31.27 | |
| Liver Tissue | ||||
| iNOS | 0.89 ± 0.11 | 1.53 ± 0.21 | 2.26 ± 0.26 * | |
| Liver Treatment Time | Endogenous Fluorophores | |||||
|---|---|---|---|---|---|---|
| Retinol (Ret) | Arachidonic Acid (AA) | Linoleic Acid | Oleic Acid | Proteins | (AA)/(Ret) | |
| 310 nm excitation | ||||||
| Sham-operated | 44.26 ± 1.95 | 25.13 ± 1.12 | ---- | 7.93 ± 0.39 | 12.48 ± 0.60 | 0.58 |
| IR 60/60 min | 31.39 ± 0.84 ** | 38.93 ± 1.04 ** | ---- | 8.86 ± 0.23 | 8.20 ± 0.21 | 1.23 |
| IR 60/120 min | 29.09 ± 3.63 ** | 39.16 ± 4.95 * | ---- | 8.56 ± 0.90 | 10.24 ± 1.45 | 1.36 |
| 366 nm excitation | ||||||
| Sham-operated | 33.43 ± 2.68 | 20.22 ± 1.52 | 23.32 ± 1.86 | 12.88 ± 1.03 | ---- | 0.60 |
| IR 60/60 min | 24.03 ± 1.35 * | 26.50 ± 1.49 * | 27.47 ± 1.55 | 16.97 ± 0.67 * | ---- | 1.10 |
| IR 60/120 min | 23.76 ± 3.34 * | 27.17 ± 3.82 | 27.02 ± 3.93 | 16.97 ± 2.38 | ---- | 1.14 |
| Liver Treatment Time | Endogenous Fluorophores | ||||
|---|---|---|---|---|---|
| Retinol (Ret) | Arachidonic Acid (AA) | Linoleic Acid | Oleic Acid | Proteins | |
| 310 nm Excitation | |||||
| Sham-operated | 18786.35 ± 828.23 | 10826.07± 477.01 | ---- | 3791.60 ± 167.59 | 5880.18 ± 251.68 |
| IR 60/60 min | 20694.20 ± 559.98 | 25811.30 ± 688.32** | ---- | 5883.34 ± 157.96 | 5447.30 ± 145.07 |
| IR 60/120 min | 16680.27 ± 2080.75 | 22768.95 ± 2848.29** | ---- | 4350.10 ± 545.00 | 6654.47 ± 833.74 |
| 366 nm Excitation | |||||
| Sham-operated | 6034.63 ± 438.46 | 3648.04 ± 292.31 | 4208.95 ± 337.17 | 2326.57 ± 186.43 | ---- |
| IR 60/60 min | 6180.46 ± 349.19 | 6809.04 ± 384.22 ** | 7077.63 ± 399.72 ** | 4377.84 ± 246.91 * | ---- |
| IR 60/120 min | 5056.64 ± 710.25 | 5780.31 ± 812.19 * | 5953.46 ± 836.49 * | 3611.97 ± 507.93* | ---- |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Croce, A.C.; Ferrigno, A.; Berardo, C.; Bottiroli, G.; Vairetti, M.; Di Pasqua, L.G. Spectrofluorometric Analysis of Autofluorescing Components of Crude Serum from a Rat Liver Model of Ischemia and Reperfusion. Molecules 2020, 25, 1327. https://doi.org/10.3390/molecules25061327
Croce AC, Ferrigno A, Berardo C, Bottiroli G, Vairetti M, Di Pasqua LG. Spectrofluorometric Analysis of Autofluorescing Components of Crude Serum from a Rat Liver Model of Ischemia and Reperfusion. Molecules. 2020; 25(6):1327. https://doi.org/10.3390/molecules25061327
Chicago/Turabian StyleCroce, Anna C., Andrea Ferrigno, Clarissa Berardo, Giovanni Bottiroli, Mariapia Vairetti, and Laura G. Di Pasqua. 2020. "Spectrofluorometric Analysis of Autofluorescing Components of Crude Serum from a Rat Liver Model of Ischemia and Reperfusion" Molecules 25, no. 6: 1327. https://doi.org/10.3390/molecules25061327
APA StyleCroce, A. C., Ferrigno, A., Berardo, C., Bottiroli, G., Vairetti, M., & Di Pasqua, L. G. (2020). Spectrofluorometric Analysis of Autofluorescing Components of Crude Serum from a Rat Liver Model of Ischemia and Reperfusion. Molecules, 25(6), 1327. https://doi.org/10.3390/molecules25061327