Abstract
Salicylic acid (SA) is a very simple phenolic compound (a C7H6O3 compound composed of an aromatic ring, one carboxylic and a hydroxyl group) and this simplicity contrasts with its high versatility and the involvement of SA in several plant processes either in optimal conditions or in plants facing environmental cues, including heavy metal (HM) stress. Nowadays, a huge body of evidence has unveiled that SA plays a pivotal role as plant growth regulator and influences intra- and inter-plant communication attributable to its methyl ester form, methyl salicylate, which is highly volatile. Under stress, including HM stress, SA interacts with other plant hormones (e.g., auxins, abscisic acid, gibberellin) and promotes the stimulation of antioxidant compounds and enzymes thereby alerting HM-treated plants and helping in counteracting HM stress. The present literature survey reviews recent literature concerning the roles of SA in plants suffering from HM stress with the aim of providing a comprehensive picture about SA and HM, in order to orientate the direction of future research on this topic.
1. Introduction
Salicylic acid (SA) (from Latin Salix, willow tree), also known as ortho-hydroxybenzoic acid, is a phenolic derivative widely distributed in the plant kingdom and is known as a regulator of several physiological and biochemical processes such as thermogenesis, plant signaling or plant defense, and response to biotic and abiotic stress [1,2].
From a chemical point of view, SA belongs to a large group of plant phenolics, and SA can be isolated in plants in both free and conjugated form. In particular, the conjugated form proceeds from the methylation, hydroxylation, and/or glucosylation of the aromatic ring [3,4].
Salicin, one of the natural SA derivatives, was first isolated from the bark of the willow tree (Salix sp.) by Johan Büchner in 1828 [5,6]. Successively, it was discovered that almost all the willow trees including Salix alba, S. purpurea, S. fragilis, and S. daphnoides were particularly rich in this natural compound, in which the concentration in plants significantly fluctuates during the different seasons (highest content during spring and summer, lowest content during autumn and winter [7]) reaching values of 3 mg/g of fresh biomass in plants of S. laponum [8]. The first scientist who was able to identify this natural compound in species different from Salix sp. was the Italian chemist Raffaele Piria in the late 1838, who obtained SA in both flower and buds of the European species Spiraea ulmaria successively renamed as Filipendula ulmaria (L.) Maxim. The discovery that this molecule was not exclusive to the Salix genus has opened the door to the study of its biosynthesis, as well as its biochemical and physiological role in plants and in 1899 the Bayer Company formulated a new drug known today as aspirin [9].
Concerning the biosynthesis of SA, it is known to be produced through the shikimate pathway by two metabolic routes (Figure 1). In the first discovered route, also known as phenylalanine route, occuring in the cytoplasm of the cell, the enzyme phenylalanine ammonia lyase (PAL) converts phenylalanine (Phe) to trans-cinnamic acid (t-CA), which gets oxidized to benzoic acid (BA). Subsequently, the enzyme benzoic-acid-2-hydroxylase (BA2H) catalyzes the hydroxylation of BA aromatic ring and leads to SA formation. The enzymatic conversion of BA into SA by BA2H requires the presence of hydrogen peroxide (H2O2) [10,11,12].
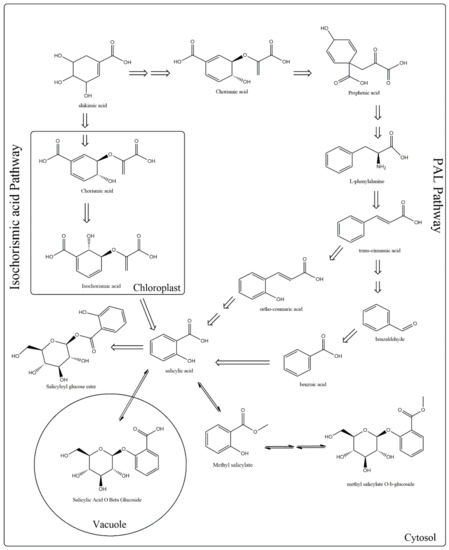
Figure 1.
Metabolic pathways involved in the biosynthesis of salicylic acid (SA). Plants use two pathways for SA production, the phenylalanine ammonia-lyase (PAL) (which is divided into two sub-pathways, benzoic acid, and o-coumaric acid) and the isochorismate. In both routes, shikimate serves as a precursor.
The first evidences for the first route were given by Ellis and Amrchein [13], who observed that feeding Gaultheria procumbens plants with labeled 14C-benzoic acid or 14C-cinnamic acid resulted in the production of labeled SA. Successively, Yalpani et al. [14] and Silverman et al. [15], working on rice and tobacco, proposed that the side chain of trans-cinnamic acid is decarboxylated to generate BA. Then, BA is hydroxylated at the C2 position forming SA. Anyway, recent results indicated that benzoyl glucose, a conjugated form of BA, is more likely to be the direct precursor of SA [12,14].
The second route is called isochorismate (IC) pathway and occurs in the chloroplast [16,17,18]. In plants, chorismate is transformed to isochorismate and then to SA, a reaction which is catalyzed by two enzymes isochorismate synthase (ICS) and isochorismate pyruvate lyase (IPL). Recent studies carried on Arabidopsis thaliana demonstrated that the ~90% of defense-related SA is produced from isochorismate generated by the plastid-localized isochorismate synthase1, whereas ~10% is derived from the cytosolic PAL pathway [1,17].
From the physiological point of view, it is known that SA plays a pivotal role in the regulation of plant growth, development, in defense from biotic and abiotic stress, and in plant immune responses [4,19,20,21,22,23].
For several years, SA was believed to be just one of the several phenolic compounds synthetized by plants with relatively low importance [5,16]. In 1974, after more than a hundred of years from its discovery, it was provided the first evidence that SA could play a role as plant hormone, when Clealand and Ajami [24] observed that SA was a mobile signaling molecule localized in the phloem inducing flowering in different plant species.
However, the final evidence that SA was a plant hormone was only provided several years later by Raskin et al. [25], who described its role during the thermogenesis in Sauromatum guttatum.
From that moment, an exponential increase of manuscripts focused on SA (acting alone or in concert with other plant hormones) as a plant growth regulator, signaling molecule, as well as plant elicitor protecting plants from biotic and abiotic stress, was observed [22,23,26,27,28,29,30,31].
Recently, it has also been demonstrated that SA could play a pivotal role in protecting plants from environmental stress, including heavy metals (HM). In fact, several recent manuscripts reported that SA can alleviate HM toxicity influencing both their uptake and/or accumulation in plant organs [32,33,34,35,36,37,38], as well as scavenging of reactive oxygen species (ROS) and/or decreasing their accumulation and/or enhancing the antioxidant defense system [39,40,41,42], protecting membrane stability and integrity [43], interacting with plant hormones [44], upregulating heme oxygenase [45], and improving the performance of the photosynthetic machinery [42,46,47].
Focusing on these aspects, the present review provides a comprehensive assemblage concerning SA roles in plant defense from HM stress, with the aim to provide a clear view of SA and HM to orientate the direction of future research on this topic.
2. HM Stress and Its Impacts on Plants
Metals and metalloids with atomic density more than 6 g cm−3 are defined as (HM). Both, essential elements, micronutrients that are required in low concentration (e.g., Cu, Cr, Co, and Zn), and nonessential metals such as Pb, Cd, Hg, are incorporated in this group [48,49]. Increased concentration of both essential and nonessential elements is phytotoxic to flora and fauna [50,51]. Heavy metal contamination has become a serious environmental problem worldwide. The increased industrialization, injudicious population growth, and urbanization releases HM that compromise soil and water and pose harms to living biota due to their biomagnification through the food chain [52]. Natural activities such as eruption of volcano and erosion of rocks have contribute in increasing the release of toxic elements to the environment; however, increased human activities such as mining, painting, and refining have enhanced their concentration in the biosphere [53,54,55].
Soil pollution by HM poses serious concerns to the biotic and abiotic components of the ecosystem [56]. The increased amount of HM in soil leads to greater uptake by plants that can reduce plant growth, biomass, photosynthesis, crop yield, and quality in plant [57]. From a biological point of view, the top soil is the most active zone of soil that accumulates a large amount of toxic metals that poses serious concern to the environment [49,58,59].
The increased level of HM accumulation in plant organs negatively affects the cell metabolism in plants [60]. The different physiological activities in plants such as protein metabolism, photosynthesis, respiration, and morphogenesis are naturally affected by a high concentration of toxic compounds, such as HM [53,54,61,62]. For instance, Rascio et al. [63] documented a decreased root growth and altered morphogenesis in rice seedlings upon treatment with Cd. Many plant species such as Brassica napus, Helianthus annuus, Thalaspi caerulescens, Vigna radiata showed inhibition in photosynthesis in response to Cd treatment [64,65,66,67,68]. Recently, Tandon and Srivastava [69] investigated the Pb effect on the morphology and metabolism of Sesamum indicum and found that the increasing concentration of metal affected the growth of the plant. Further, the plant showed severe symptoms of chlorosis, necrosis and reduced chlorophyll, and protein content at higher doses of Pb [69].
The major outcome of metal toxicity is the peaked production of ROS due to impairment of photosynthetic process by HM [70]. ROS such as hydroxyl, superoxide, and hydrogen peroxide are produced as by-product during electron transport in photosynthesis and respiration pathways [71]. Under physiological conditions, ROS play a multitude of signaling roles in plants, as well as in other organisms and they take part in a finely-tuned and well-orchestrated regulatory network [72,73]. ROS are indeed integrated into a complex regulatory system in plants which encompasses ROS, plant hormones (e.g., ethylene (ET) and abscisic acid (ABA)), signaling molecules (e.g., salicylic acid (SA) and jasmonic acid (JA)), and secondary messengers (e.g., Ca2+) [74,75]. However, when ROS production exceeds the physiological levels, their accumulation can lead to oxidative stress in the cells, that cause lipids peroxidation, macromolecular degradation, membrane disruption, DNA breakage, and ion leakage in plants [70,74,75]. For instance, Kaur et al. [76] explored Pb-induced ultrastructural changes in roots of wheat and concluded that Pb inhibited root growth, caused ROS generation, and disrupted mitochondrial and nuclear integrity in the tested plant.
The enhanced generation of ROS in the plant cell is controlled by a complex network of antioxidant machinery that maintains ROS homeostasis in the cell [77]. Plants have a finely-tuned and well-orchestrated defense system that includes enzymatic antioxidants such as catalase (CAT), superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione peroxidase (GPX) and glutathione reductase (GR), and nonenzymatic antioxidants such as ascorbic acid, glutathione, alkaloids, phenol compounds, and α-tocopherol for scavenging excessive ROS [49,61]. Moreover, phytohormones such as auxins, gibberellins, cytokinins, abscisic acid, ethylene, brassinosteroids, jasmonic acid, and SA take part in the defensive mechanism of plants against HM stress.
3. Physiological Roles of SA in Plants Under HM Stress
Concerning the physiological role in plants, SA is known to play a pivotal role in regulating plant morphology, development, flowering, and stomatal closure [78,79]. SA also affects seedling germination, cell growth, and nodulation in legumes [80]. Khan et al. [81] reported increased leaf area and dry weight production in corn and soybean in response to SA. Furthermore, Hussein et al. [82] reported pot studies that documented improved growth, leaf number, dry biomass, and stem diameter in wheat plants when leaves were sprayed with SA. The rate of transpiration and stomatal index of plants increased in response to supplementation of SA [81]. The pigment concentration in wheat seeds significantly enhanced upon exposure to a low concentration (10−5 M) of SA. However, foliar application of SA reduced transpiration rate in test plants, Phaseolus vulgaris and Commelina communis which might be due to the SA-evoked stomatal closure [83,84,85,86,87]. Moreover, SA has been reported to increase the shelf life of cut flowers of rose and defer senescence by controlling water level in rose plants [86].
Plant growth regulators or phytohormones especially, gibberellins, auxin, cytokinins, ethylene, brassinosteroids, and also SA play a key role in providing HM tolerance in plants [83]. SA, a phenolic plant hormone, regulates photosynthesis, respiration, and antioxidant defense mechanism in plants under different abiotic stress such as high temperature, salinity, and HM [78,88,89]. SA pretreatment provides protection from various metals such as Pb, Hg, Cd, in different plants [90,91,92].
Supplementation of SA in combination with plant growth promoting bacteria reduces Cr-induced oxidative damage in maize by enhancing activities of antioxidant and nonantioxidant enzymes [93,94]. Earlier, Song et al. [95] reported SA mediated enhancement in the activities of CAT and SOD enzymes in barley leaves under Zn, Cu, and Mn stress. Further, carbohydrate metabolism in Cr-treated maize plants improved upon exposure to SA [94]. Alleviation of Cd toxicity was reported in mustard plants in response to exogenous treatment of SA [93]. Recently, SA treatment mitigated Cd stress in Brassica juncea plants and enhanced growth and photosynthesis in plants. Moreover, supplementation of SA reduced reactive oxygen species levels by strengthening the antioxidant defense system in plants and provides stability to the plant membrane [96]. The exogenous application of SA upregulates the antioxidant system, improves growth and yield, and results in lowering of oxidative damage under Pb stress in B. campestris [97].
A schematization of the protective role exerted by SA in HM-stressed plants is reported in Figure 2, whereas a literature survey on the effect of different HM on plant metabolism is reported in Table 1.
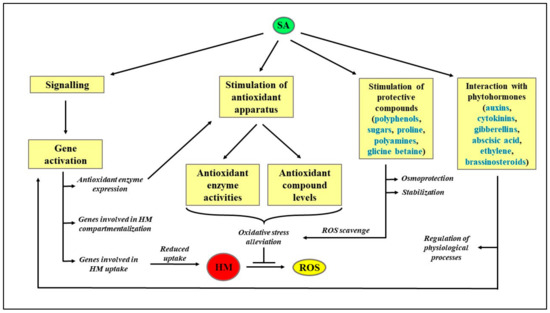
Figure 2.
Schematization of the protective role exerted by SA in HM-stressed plants. HM: Heavy metals; ROS: Reactive oxygen species; SA: Salicylic acid.

Table 1.
Salicylic acid (SA) effect on different heavy metals (HM) stressed plants.
3.1. Effect of SA to Photosynthesis in Plants Subjected to HM Stress
The different stressful conditions encountered by plants affect multiple physiological and biochemical mechanisms in plants. Among these, photosynthesis is usually one of the most affected mechanisms by HM (see a schematization of the effect of HM on chloroplast in Figure 2). HM accumulated in various organs of plants and affect the synthesis of photosynthetic pigments, including carotenoids and chlorophylls [53,54]. HM also alter the chloroplast membrane structure and affect electron transport, thus impairing light-dependent reactions of photosynthesis [120]. Moreover, it was found that the negative effect of HM on PSI and PSII depends on exposure time and concentrations [121,122]. Experiments performed by Khan et al. [123] indicated that PSII is more sensitive to HM stress compared to PSI, however, at high concentrations the activity of PSI resulted inhibited as well. Photosynthesis inhibition caused by HM is also attributable to the impairment of stomatal conductance and transpiration rate [124].
Plants are equipped with multiple mechanisms to preserve the photosynthetic machinery from HM-promoted damages. SA is a major photosynthesis regulator which influences chlorophyll content, stomatal conductivity, and photosynthesis-related enzyme activities in plants [125]. It enhances photosynthetic efficiency and improves photosynthetic apparatus under HM stress [34]. Exogenous application of SA (500 μM) enhanced chlorophyll concentration, CO2 fixation, and activities of phosphoenolpyruvate carboxylase and RuBISCO in Triticum aestivum under Cd toxicity [126]. Further, gas exchange parameters and carbonic anhydrase improved in B. juncea under Ni [120] and Mn [127] stress after the exposure to 10 μM SA. SA treatment enhanced Chla, Chlb, and carotenoid content in barley plants under Pb stress by increasing antioxidant activity in the plants which might be due to blockage of Ca channels that help in translocation of Pb in roots [60]. Recently, Guo et al. [38] studied the role of SA in Cd alleviation and accumulation in tomato plants. The exogenous exposure of SA also increased pigment content and photosynthetic performance in tomato plants [38]. The consistently observed protective role of SA to the photosynthetic apparatus might be due to increased detoxification of ROS species exerted by SA or by the activation of antioxidant apparatus promoted by SA [125].
3.2. Regulation Mechanism of ROS and Enzymatic Antioxidants Promoted by SA Acid under HM Stress
The generation of ROS is one the first response in plants under HM stress. ROS production is either directly due to Haber-Weiss reaction or it is indirectly because of interference in the antioxidant defense system or electron transport chain [128]. ROS (H2O2; hydrogen peroxide, OH·; hydroxyl radical, and O2−·; superoxide radical) are very harmful to plants since they lead to oxidative degeneration of cell membranes and large macromolecules [129]. Plants possess a powerful antioxidant apparatus to counteract oxidative stress, which includes different enzymes (SOD, CAT, APX, GR) and nonenzymatic antioxidants (e.g., glutathione, ascorbic acid, phenolics, carotenoids) that scavenge and detoxify ROS over-production in plants [130].
Lipid peroxidation is the first oxidative injury in plants due to HM stress and SA have been shown to provide stability against HM-induced oxidative damage by increasing antioxidant machinery in plants [125]. Parashar et al. [127] and Zhang et al. [131] observed the reduction in lipid peroxidation, electrolyte leakage, and superoxide ion in Mn- and Cd-treated B. juncea and Cucumis melo upon addition of SA. Few experiments suggest that SA can promote free radical scavenging of HM-promoted ROS by regulating antioxidant enzymes and expression of some proteins and molecules such as OsWRKY45 as reported in rice by Chao et al. [132] that lowers H2O2 accumulation in plants. This helps in maintaining the balance between ROS generation and membrane integrity, thereby preventing membrane disruption [133]. Recently, Lu et al. [98] and Gu et al. [99] documented activation of antioxidant enzymes including SOD, APX, and other peroxidases in Lemna minor and Nymphaea tetragona upon supplementation of SA in plants subjected to Cd stress, which were helpful in conferring Cd tolerance in plants.
3.3. Regulation of Osmolytes and Polyphenols by SA under HM Stress
Plants have evolved various mechanisms to counteract HM-triggered ROS production. Different antioxidant metabolites such as proline, glycine betaine, polyamines, sugars, and polyphenols are all involved in maintaining the ROS balance in plants under stressful conditions, including excess of HM. Below, the intimal connections between SA and other antioxidant compounds are described with the attempt to provide a clear and exhaustive picture about the SA-promoted regulation of antioxidant molecules in plants exposed to HM.
3.3.1. Proline
Proline acts as a free radical scavenger, osmo-protectant, and stabilizer of cellular structures [130,134]. The synthesis of proline occurs from glutamate, which is converted to glutamate-semialdehyde, and then spontaneously to pyrroline-5-carboxylate (P5C) with the help of P5C synthase enzyme. Later, the enzyme P5C reductase aids in the reduction of P5C to proline. The stimulation of proline levels under HM stress was observed, for example, in Olea europaea [135] and Phoenix dactylifera [136]. However, this is not clear whether the accumulation was attributable to enhanced production of enzymes responsible for proline synthesis, the decrease in enzymes related to its oxidation or both. SA is involved in enhancing proline level under HM toxicity [96]. Parashar et al. [127] reported that SA ameliorated the Mn stress through enhanced accumulation of proline in Brassica juncea which might be due to the increased activity of enzymes responsible for proline synthesis [137]. Enhanced proline content also maintains water balance in plants to contrast stressful conditions leading to osmotic stress [138] a condition which can occur when plants reduce the stomatal conductance in order to reduce HM uptake. Further, Chen and Dickman [139] proposed that proline is a powerful ROS scavenger and a pivotal component of protein pathway in plants, besides serving as an osmoprotectant [140]. Zanganeh et al. [141] observed however that SA pre-treatment decreased proline accumulation in Zea mays under Pb stress that was supported by the findings of Mostafa et al. [142] in rice plants. Therefore, the pattern of proline (activation/decrement) can be species- or metal-specific and also dependent on the dose of HM experienced by the plant species.
3.3.2. Glycine Betaine
Glycine betaine (GB) is a quaternary level ammonium compound found in higher plants under stress conditions and it acts as osmoprotectant or compatible solutes in plants [143], in which it accumulates at cytosolic level. GB is involved in providing protection against drought, salinity [93], drought [143], and HM stress, as well [144]. Exogenous application of GB is very effective in providing tolerance from HM stress [94,145]. The role of SA in regulating the accumulation of GB in plants under metal stress is still unknown. However, few studies reported that exogenous treatment of GB together with SA can help in alleviating HM toxicity [145]. Recently, Aldesuquy et al. [146] opined that GB and SA regulates osmotic pressure and concentration of osmolytes in plants that maintain osmotic balance and helps in ameliorating the adverse effect of drought stress in wheat, thereby suggesting a possible cooperation. It was also reported that the SA induced the rise in GB level which helped the growth of Rauwolfia serpentina plants grown under Na excess [147].
3.3.3. Sugars
The term sugars, collectively used for disaccharides (sucrose, trehalose) and fructans, are water-soluble carbohydrates involved in plant stress tolerance. Sucrose, an important product of photosynthesis, is required for growth, development, storage, and signaling in plants [148,149]. Carbohydrates are building blocks of plants that provide energy and act as a signaling molecule during transcriptional, post-transcriptional processes [150]. Accumulation of soluble sugars has been observed in plants under stressful conditions which indicate their role as osmoprotectant and in maintaining cellular balance in plants [151,152]. The exogenous addition of SA enhanced the amount of polysaccharides and sugars in plants and helped in improving their growth [153]. El-tayeb et al. [154] observed that SA provided Cu tolerance in Helianthus annuus. The authors reported an increasing level of soluble sugars in plants treated with SA that protects the photosynthetic pigments from Cu toxicity [154]. Similarly, 0.01 M SA enhanced growth and sugar accumulation in tomato plants and provided stress avoidance and tolerance against Na toxicity [155].
3.3.4. Polyamines
Polyamines (PAs) are water-soluble molecules that play an important role in regulating morphological, developmental, and stress responses in plants [156]. PA have the potential to scavenge HM-triggered ROS [157] and regulate plant defense response to HM toxicity [156,158]. Under stressful conditions, PA operate as signaling compounds and control ion homeostasis and ion transportation in plants, thus actively participating in stress tolerance [159,160]. Many reports suggest that SA treatment influence PA content in plants [131,161]. Recently, Tajti et al. [162] studied the role of putrescine and spermidine on wheat under Cd stress and also reported increased levels of SA in those plants; however, the exact mechanism involved in SA-mediated HM stress tolerance and the relationship between PA and SA in plants are still unknown.
3.3.5. Polyphenols
Phenolics are one of the largest groups of secondary metabolites which include a plethora of compounds with simple aromatic rings to very complex molecules, such as tannins and lignans. They originate from phenylalanine by the activity of PAL. Many reports have demonstrated that enhanced production of phenolic compounds under HM stress can protect from oxidative damage [163,164]. The accumulation of phenolics is principally driven by increased expression of enzymes responsible for phenylpropanoid biosynthesis such as phenylalanine ammonia-lyase, chalcone synthase, shikimate dehydrogenase, cinnamyl alcohol dehydrogenase, and polyphenol oxidase [165,166]. Many studies have documented the role of phytohormones in enhancing the level of some classes of polyphenols, such as anthocyanins [167,168]. Dong et al. [169] reported increased concentrations of phenolics, such as caffeic acid due to exogenous treatment by SA. Similarly, peaked activity of PAL was observed in Matricaria chamomilla plants under Ni and Cd stress with the application of SA [170].
3.4. Regulation of Cell Signaling by SA under HM Stress
The HM stress tolerance induced by SA is supportive for its role in stress signaling. The mechanism of tolerance not only depends on the concentration and mode of application of SA but also on the overall status of plants [171]. Abiotic stress not only affects growth and development of plants, but also regulates DNA replication machinery. SA application upregulates the topoisomerase gene and chloroplast elongation factor that help in plant adaptation under stressful conditions [172,173]. Moreover, SA is known to induce expression of TLC1, a long terminal repeated retrotransposon family in vivo [171]. This family is transcriptionally activated during stressful conditions and its expression by SA suggests their role in SA-mediated signaling pathways [171]. Another mechanism adopted by SA in regulating HM stress plant response is the increased activity of enzymes involved in AsA-GSH pathway [174]. Both AsA and GSH are active redox compounds that maintain cellular redox balance in plants [175]. SA supplementation also increased SOD and POD level in Cannabis sativa and improved Cd-tolerance [34] which might be related to increased concentration of Ca2+ (a second messenger) and H2O2, that eventually promote the activity of antioxidant enzymes which reduce cellular ROS level in plants [176,177].
3.5. Crosstalk of SA with Other Plant Growth Regulators
SA regulates different plant responses both under optimal and stressful conditions through the crosstalk with other plant growth regulators or plant hormones [81,178]. The interaction of SA with other hormones such as auxin [179], cytokinin [180], gibberellins [181], abscisic acid [182], ethylene [178], and brassinosteroids [87] has been studied under optimum and stressful environments. The possible outcome of interaction of SA with hormones can be either synergistic or antagonistic under stressful conditions. Recently, Tamás et al. [44] studied the SA regulated alleviation of Cd-stress by restriction of Cd-induced auxin-mediated ROS production in barley roots. The authors suggest that SA treatment reversed indole-3-acetic acid (IAA)-induced stress responses in plants suggesting a role of SA in IAA signaling pathway. Similarly, Agtuca et al. [183] reported an opposite role of IAA and SA in roots of maize. The exogenous application of IAA enhanced lateral growth by depriving primary root growth, while SA increased total root biomass [183].
Exposure to various environmental stresses, such as HM, can enhance ethylene production and induce oxidative stress in plants [175]. The increased ethylene production is due to peaked expression of ethylene-related biosynthetic genes or expression of ethylene-responsive genes [184]. Exogenous SA was reported to mitigate Cd stress in wheat [174] by increasing GSH content that resulted in metal detoxification and scavenging ROS induced by HM-triggered ethylene production. SA supplementation promoted increased ABA level in wheat seedlings under Cd stress that was attributed to a de novo ABA biosynthesis [185]. Further, endogenous ABA controlled SA-mediated alteration of the concentration of dehydrin proteins under HM stress that demonstrate protective mechanism of SA in wheat plants [185].
Under abiotic stress conditions, crosstalk between SA and jasmonates play a crucial role in regulation of plant growth [186]. Generally, SA and jasmonic acid (JA) signaling pathways work in an antagonistic manner [187]. The Mitogen-activated protein kinase (MAPK) signaling pathway mediates the antagonistic action between SA and JA cell signaling [188]. However, nonantagonistic interaction between SA and JA are also reported, but an exact mechanism is still unclear and it needs further studies [186]. For example, in maize plants Cu stress induced the biosynthesis of SA, which further induced JA priming and JA induced volatile organic compounds [189,190].
4. Conclusions
Heavy metal stress has been accepted as one of the major threats for plants growing in contaminated areas. In order to deal with the harmful effects of heavy metals, plants have developed several molecular, metabolic, and physiological processes which allow them to avoid stressful factors or cope with them.
Several researches highlighted that SA, when used at low doses, plays a pivotal role in both alleviating and reducing heavy metal stress in plants. An increase in the endogenous level, as well as exogenous application of this plant hormone has been demonstrated to be helpful for plants either in optimal or in stress conditions. In fact, this ubiquitous plant hormone is involved in the regulation of several metabolic processes in plants, regulating the ex novo biosynthesis of secondary metabolites and osmoprotectants involved in the protection from oxidative stress, thereby increasing the activity of ROS scavenger enzymes and/or acting as antioxidants. However, at high concentrations SA can also act as a negative plant growth regulator [171,191,192].
The scientific literature cited in the present review highlights the important role played by SA in protecting plants from heavy metal stress. However, most of the researches available on this topic are mainly focused on the role played by this molecule after an exogenous application, while very few researches, because of the complexity of the cascade effects generated, have unveiled the defense mechanisms triggered by its endogenous stimulation in response to heavy metals. Therefore, there are still several questions which need further investigation. For example, it would be extremely interesting to disentangle the complexity of SA signaling in response to heavy metals, as well as to unveil if exogenous application of SA might directly or indirectly enhance endogenous SA levels. In the meantime, more genomic, transcriptomic, proteomic, and metabolomics studies are necessary to detect SA responsive genes, proteins, and metabolites altered by heavy metal stress. In addition, it is necessary that a molecular dissection deeply understands the crosstalk between SA with other phytohormones and/or metabolites and the feedback processes involved in controlling the endogenous levels of SA in response to heavy metal stress.
Author Contributions
All authors contributed in writing part of the original draft and also reviewed and edited the whole manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Z.; Zheng, Z.; Huang, J.; Lai, Z.; Fan, B. Biosynthesis of salicylic acid in plants. Plant Sign. Behav. 2009, 4, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Wani, A.B.; Chadar, H.; Wani, A.H.; Singh, S.; Upadhyay, N. Salicylic acid to decrease plant stress. Environ. Chem. Lett. 2017, 15, 101–123. [Google Scholar] [CrossRef]
- Lovelock, D.A.; Šola, I.; Marschollek, S.; Donald, C.E.; Rusak, G.; van Pée, K.H.; Ludwig-Müller, J.; Cahill, D.M. Analysis of salicylic acid-dependent pathways in Arabidopsis thaliana following infection with Plasmodiophora brassicae and the influence of salicylic acid on disease. Molecul. Plant Pathol. 2016, 17, 1237–1251. [Google Scholar] [CrossRef]
- Maruri-López, I.; Aviles-Baltazar, N.Y.; Buchala, A.; Serrano, M. Intra and extracellular journey of the phytohormone salicylic acid. Front. Plant Sci. 2019, 10, 423. [Google Scholar] [CrossRef] [PubMed]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Muthulakshmi, S.; Lingakumar, K. Role of salicylic acid (SA) in plants—A review. Int. J. Appl. Res. 2017, 3, 33–37. [Google Scholar]
- Foster, S. Tyler’s Honest Herbal: A Sensible Guide to the Use of Herbs and Related Remedies; Routledge: New York, NY, USA, 1999. [Google Scholar]
- Petrek, J.; Havel, L.; Petrlova, J.; Adam, V.; Potesil, D.; Babula, P.; Kizek, R. Analysis of salicylic acid in willow barks and branches by an electrochemical method. Russ. J. Plant Physiol. 2007, 54, 553–558. [Google Scholar] [CrossRef]
- Arif, H.; Aggarwal, S. Salicylic Acid (Aspirin); StatPearls Publishing LLC.: Tampa, FL, USA; St. Petersburg, Russia, 2019. [Google Scholar]
- Shine, M.; Yang, J.W.; El-Habbak, M.; Nagyabhyru, P.; Fu, D.Q.; Navarre, D.; Ghabrial, S.; Kachroo, P.; Kachroo, A. Cooperative functioning between phenylalanine ammonia lyase and isochorismate synthase activities contributes to salicylic acid biosynthesis in soybean. New Phytol. 2016, 212, 627–636. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, X.; Hao, X.; Zhang, L.; Wang, L.; Qian, H.; Zhao, J. Molecular cloning and promoter analysis of the specific salicylic acid biosynthetic pathway gene phenylalanine ammonia-lyase (AaPAL1) from Artemisia annua. Biotech. Appl. Biochem. 2016, 63, 514–524. [Google Scholar] [CrossRef]
- Chong, J.; Pierrel, M.-A.; Atanassova, R.; Werck-Reichhart, D.; Fritig, B.; Saindrenan, P. Free and conjugated benzoic acid in tobacco plants and cell cultures. Induced accumulation upon elicitation of defense responses and role as salicylic acid precursors. Plant Physiol. 2001, 125, 318–328. [Google Scholar] [CrossRef]
- Hayat, S.; Ali, B.; Ahmad, A. Salicylic acid: Biosynthesis, metabolism and physiological role in plants. In Salicylic Acid: A Plant Hormone; Springer: Dordrecht, The Netherlands, 2007; pp. 1–14. [Google Scholar]
- Yalpani, N.; León, J.; Lawton, M.A.; Raskin, I. Pathway of salicylic acid biosynthesis in healthy and virus-inoculated tobacco. Plant Physiol. 1993, 103, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Silverman, P.; Seskar, M.; Kanter, D.; Schweizer, P.; Metraux, J.-P.; Raskin, I. Salicylic acid in rice (biosynthesis, conjugation, and possible role). Plant Physiol. 1995, 108, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Métraux, J.-P. Recent breakthroughs in the study of salicylic acid biosynthesis. Trends Plant Sci. 2002, 7, 332–334. [Google Scholar] [CrossRef]
- Garcion, C.; Lohmann, A.; Lamodière, E.; Catinot, J.; Buchala, A.; Doermann, P.; Métraux, J.-P. Characterization and biological function of the Isochorismate Synthase2 gene of Arabidopsis. Plant Physiol. 2008, 147, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Rekhter, D.; Lüdke, D.; Ding, Y.; Feussner, K.; Zienkiewicz, K.; Lipka, V.; Wiermer, M.; Zhang, Y.; Feussner, I. Isochorismate-derived biosynthesis of the plant stress hormone salicylic acid. Science 2019, 365, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liu, G.; Chang, Y.; He, C.; Shi, H. Heat shock transcription factor 3 regulates plant immune response through modulation of salicylic acid accumulation and signalling in cassava. Mol. Plant Pathol. 2018, 19, 2209–2220. [Google Scholar] [CrossRef]
- Hartmann, M.; Zeier, J. N-Hydroxypipecolic acid and salicylic acid: A metabolic duo for systemic acquired resistance. Curr. Opin. Plant Biol. 2019, 50, 44–57. [Google Scholar] [CrossRef]
- El-Shazoly, R.M.; Metwally, A.A.; Hamada, A.M. Salicylic acid or thiamin increases tolerance to boron toxicity stress in wheat. J. Plant Nutr. 2019, 42, 702–722. [Google Scholar] [CrossRef]
- Luo, J.; Xia, W.; Cao, P.; Xiao, Z.A.; Zhang, Y.; Liu, M.; Zhan, C.; Wang, N. Integrated transcriptome analysis reveals plant hormones jasmonic acid and salicylic acid coordinate growth and defense responses upon fungal infection in poplar. Biomolecules 2019, 9, 12. [Google Scholar] [CrossRef]
- Pasternak, T.; Groot, E.P.; Kazantsev, F.V.; Teale, W.; Omelyanchuk, N.; Kovrizhnykh, V.; Palme, K.; Mironova, V.V. Salicylic acid affects root meristem patterning via auxin distribution in a concentration-dependent manner. Plant Physiol. 2019, 180, 1725–1739. [Google Scholar] [CrossRef]
- Cleland, C.F.; Ajami, A. Identification of the flower-inducing factor isolated from aphid honeydew as being salicylic acid. Plant Physiol. 1974, 54, 904–906. [Google Scholar] [CrossRef] [PubMed]
- Raskin, I.; Skubatz, H.; Tang, W.; Meeuse, B.J. Salicylic acid levels in thermogenic and non-thermogenic plants. Ann. Bot. 1990, 66, 369–373. [Google Scholar] [CrossRef]
- Dempsey, D.M.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 23. [Google Scholar] [CrossRef]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.M.A. Systemic acquired resistance and salicylic acid: Past, present, and future. Mol. Plant-Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef] [PubMed]
- Subban, K.; Subramani, R.; Srinivasan, V.P.M.; Johnpaul, M.; Chelliah, J. Salicylic acid as an effective elicitor for improved taxol production in endophytic fungus Pestalotiopsis microspora. PLoS ONE 2019, 14, e0212736. [Google Scholar] [CrossRef]
- Tripathi, D.; Raikhy, G.; Kumar, D. Chemical elicitors of systemic acquired resistance—Salicylic acid and its functional analogs. Curr. Plant Biol. 2019, 17, 48–59. [Google Scholar] [CrossRef]
- Nadeem, M.; Ahmad, W.; Zahir, A.; Hano, C.; Abbasi, B.H. Salicylic acid-enhanced biosynthesis of pharmacologically important lignans and neo lignans in cell suspension culture of Linum ussitatsimum L. Eng. Life Sci. 2019, 19, 168–174. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.-J. Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef]
- Safari, F.; Akramian, M.; Salehi-Arjmand, H.; Khadivi, A. Physiological and molecular mechanisms underlying salicylic acid-mitigated mercury toxicity in lemon balm (Melissa officinalis L.). Ecotoxic Environ. Safety 2019, 183, 109542. [Google Scholar] [CrossRef]
- Dalvi, A.A.; Bhalerao, S.A. Response of plants towards heavy metal toxicity: An overview of avoidance, tolerance and uptake mechanism. Ann. Plant. Sci. 2013, 2, 362–368. [Google Scholar]
- Shi, G.; Cai, Q.; Liu, Q.; Wu, L. Salicylic acid-mediated alleviation of cadmium toxicity in hemp plants in relation to cadmium uptake, photosynthesis, and antioxidant enzymes. Acta Physiol. Plant 2009, 31, 969–977. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Wang, P.; Hou, J.; Qian, J.; Ao, Y.; Lu, J.; Li, L. Salicylic acid involved in the regulation of nutrient elements uptake and oxidative stress in Vallisneria natans (Lour.) Hara under Pb stress. Chemosphere 2011, 84, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Lv, X.; Jia, H.; Hua, L.; Xu, H.; Zhou, R.; Zhao, J.; Ren, X.; Guo, J. Effects of salicylic acid, Fe (II) and plant growth-promoting bacteria on Cd accumulation and toxicity alleviation of Cd tolerant and sensitive tomato genotypes. J. Environ. Manag. 2018, 214, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Combined effect of 24-epibrassinolide and salicylic acid mitigates lead (Pb) toxicity by modulating various metabolites in Brassica juncea L. seedlings. Protoplasma 2018, 255, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, R.; Ren, X.; Jia, H.; Hua, L.; Xu, H.; Lv, X.; Zhao, J.; Wei, T. Effects of salicylic acid, Epi-brassinolide and calcium on stress alleviation and Cd accumulation in tomato plants. Ecotoxic. Environ. Saf. 2018, 157, 491–496. [Google Scholar] [CrossRef]
- Malik, Z.A.; Lal, E.P.; Mir, Z.A.; Lone, A.H. Effect of salicyclic acid and indole acetic acid on tomato crop under induced salinity and cadmium stressed environment: A Review. Int. J. Plant Soil Sci. 2018, 26, 1–6. [Google Scholar] [CrossRef]
- Mohamed, H.E.; Hassan, A.M. Role of salicylic acid in alleviating cobalt toxicity in wheat (Triticum aestivum L.) seedlings. J. Agric. Sci. 2019, 11. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.; Ansary, M.; Uddin, M.; Fujita, M.; Tran, L.-S.P. Interactive effects of salicylic acid and nitric oxide in enhancing rice tolerance to cadmium stress. Int. J. Mol. Sci. 2019, 20, 5798. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Wang, Y.; Li, G.-Z.; Hao, L. Salicylic acid-altering Arabidopsis plant response to cadmium exposure: Underlying mechanisms affecting antioxidation and photosynthesis-related processes. Ecotoxicol. Environ. Safety 2019, 169, 645–653. [Google Scholar] [CrossRef]
- Belkadhi, A.; De Haro, A.; Obregon, S.; Chaïbi, W.; Djebali, W. Positive effects of salicylic acid pretreatment on the composition of flax plastidial membrane lipids under cadmium stress. Environ. Sci. Poll. Res. 2015, 22, 1457–1467. [Google Scholar] [CrossRef]
- Tamás, L.; Mistrík, I.; Alemayehu, A.; Zelinová, V.; Bočová, B.; Huttová, J. Salicylic acid alleviates cadmium-induced stress responses through the inhibition of Cd-induced auxin-mediated reactive oxygen species production in barley root tips. J. Plant Physiol. 2015, 173, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Li, L.; Gao, Z.; Wu, H.; Xie, Y.; Shen, W. Haem oxygenase-1 is involved in salicylic acid-induced alleviation of oxidative stress due to cadmium stress in Medicago sativa. J. Exp. Bot. 2012, 63, 5521–5534. [Google Scholar] [CrossRef] [PubMed]
- Tahjib-Ul-Arif, M.; Siddiqui, M.N.; Sohag, A.A.M.; Sakil, M.A.; Rahman, M.M.; Polash, M.A.S.; Mostofa, M.G.; Tran, L.-S.P. Salicylic acid-mediated enhancement of photosynthesis attributes and antioxidant capacity contributes to yield improvement of maize plants under salt stress. J. Plant Grow. Regul. 2018, 37, 1318–1330. [Google Scholar] [CrossRef]
- Yin, Q.-S.; Yuan, X.; Jiang, Y.-G.; Huang, L.-L.; Li, G.-Z.; Hao, L. Salicylic acid-mediated alleviation in NO2 phytotoxicity correlated to increased expression levels of the genes related to photosynthesis and carbon metabolism in Arabidopsis. Environ. Exp. Bot. 2018, 156, 141–150. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Zia-ur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci. Poll. Res. 2015, 22, 8148–8162. [Google Scholar] [CrossRef]
- Pinto, A.; De Varennes, A.; Fonseca, R.; Teixeira, D.M. Phytoremediation of soils contaminated with heavy metals: Techniques and strategies. In Phytoremediation; Springer: Dordrecht, The Netherlands, 2015; pp. 133–155. [Google Scholar]
- Kumar, A.; Usmani, Z.; Ahirwal, J.; Rani, P. Phytomanagement of chromium contaminated brown fields. In Phytomanagement of Polluted Sites; Elsevier: Amsterdam, The Neitherland, 2019; pp. 447–469. [Google Scholar]
- Park, J.H.; Lamb, D.; Paneerselvam, P.; Choppala, G.; Bolan, N.; Chung, J.-W. Role of organic amendments on enhanced bioremediation of heavy metal (loid) contaminated soils. J. Haz. Mat. 2011, 185, 549–574. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A.; Kaur, P.; Sidhu, G.P.S.; Bali, A.S.; Bhardwaj, R.; Thukral, A.K.; Cerda, A. Pollution assessment of heavy metals in soils of India and ecological risk assessment: A state-of-the-art. Chemosphere 2019, 216, 449–462. [Google Scholar] [CrossRef]
- Sidhu, G.P.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Effect of lead on oxidative status, antioxidative response and metal accumulation in Coronopus didymus. Plant Physiol. Biochem. 2016, 105, 290–296. [Google Scholar] [CrossRef]
- Sidhu, G.P.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Tolerance and hyperaccumulation of cadmium by a wild, unpalatable herb Coronopus didymus (L.) Sm.(Brassicaceae). Ecotox. Environ. Safety 2017, 135, 209–215. [Google Scholar] [CrossRef]
- Sidhu, G.P.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Appraising the role of environment friendly chelants in alleviating lead by Coronopus didymus from Pb-contaminated soils. Chemosphere 2017, 182, 129–136. [Google Scholar] [CrossRef]
- Keesstra, S.; Mol, G.; de Leeuw, J.; Okx, J.; de Cleen, M.; Visser, S. Soil-related sustainable development goals: Four concepts to make land degradation neutrality and restoration work. Land 2018, 7, 133. [Google Scholar] [CrossRef]
- Ramzani, P.M.A.; Iqbal, M.; Kausar, S.; Ali, S.; Rizwan, M.; Virk, Z.A. Effect of different amendments on rice (Oryza sativa L.) growth, yield, nutrient uptake and grain quality in Ni-contaminated soil. Environ. Sci. Poll. Res. 2016, 23, 18585–18595. [Google Scholar] [CrossRef] [PubMed]
- Ihedioha, J.; Ukoha, P.; Ekere, N. Ecological and human health risk assessment of heavy metal contamination in soil of a municipal solid waste dump in Uyo, Nigeria. Environ. Geochem. Health 2017, 39, 497–515. [Google Scholar] [CrossRef] [PubMed]
- Van Nevel, L.; Mertens, J.; Staelens, J.; de Schrijver, A.; Tack, F.M.; de Neve, S.; Meers, E.; Verheyen, K. Elevated Cd and Zn uptake by aspen limits the phytostabilization potential compared to five other tree species. Ecol. Eng. 2011, 37, 1072–1080. [Google Scholar] [CrossRef]
- Arshad, T.; Maqbool, N.; Javed, F.; Wahid, A.; Arshad, M.U. Enhancing the defensive mechanism of lead affected barley (Hordeum vulgare L.) genotypes by exogenously applied salicylic acid. J. Agric. Sci. 2017, 9, 139–146. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef]
- Guerra, F.; Gainza, F.; Pérez, R.; Zamudio, F. Phytoremediation of heavy metals using poplars (Populus spp.): A glimpse of the plant responses to copper, cadmium and zinc stress. In Handbook of Phytoremediation; Nova Science: New York, NY, USA, 2011; pp. 387–413. [Google Scholar]
- Rascio, N.; Dalla Vecchia, F.; La Rocca, N.; Barbato, R.; Pagliano, C.; Raviolo, M.; Gonnelli, C.; Gabbrielli, R. Metal accumulation and damage in rice (cv. Vialone nano) seedlings exposed to cadmium. Environ. Exp. Bot. 2008, 62, 267–278. [Google Scholar] [CrossRef]
- Baryla, A.; Carrier, P.; Franck, F.; Coulomb, C.; Sahut, C.; Havaux, M. Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cadmium-polluted soil: Causes and consequences for photosynthesis and growth. Planta 2001, 212, 696–709. [Google Scholar] [CrossRef]
- Di Cagno, R.; Guidi, L.; De Gara, L.; Soldatini, G. Combined cadmium and ozone treatments affect photosynthesis and ascorbate-dependent defences in sunflower. New Phytol. 2001, 151, 627–636. [Google Scholar] [CrossRef]
- Küpper, H.; Parameswaran, A.; Leitenmaier, B.; Trtílek, M.; Šetlík, I. Cadmium-induced inhibition of photosynthesis and long-term acclimation to cadmium stress in the hyperaccumulator Thlaspi caerulescens. New Phytol. 2007, 175, 655–674. [Google Scholar] [CrossRef]
- TRAN, T.A.; Popova, L.P. Functions and toxicity of cadmium in plants: Recent advances and future prospects. Turkish J. Bot. 2013, 37, 1–13. [Google Scholar]
- Wahid, A.; Ghani, A.; Javed, F. Effect of cadmium on photosynthesis, nutrition and growth of mungbean. Agr. Sustain. Develop. 2008, 28, 273–280. [Google Scholar] [CrossRef]
- Tandon, P.K.; Srivastava, P. Growth and metabolism of sesame (Sesamum indicum L.) plants in relation to lead toxicity. Agricul. Sci. 2014, 6, 91–92. [Google Scholar]
- Rascio, N.; Navari-Izzo, F. Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 2011, 180, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Kadukova, J.; Kavuličova, J. Phytoremediation of heavy metal contaminated soils—Plant stress assessment. Handbook of Phytoremediation; Nova Science: New York, NY, USA, 2011; pp. 185–222. [Google Scholar]
- Baxter, A.; Mittler, R.; Suzuki, N. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Cotrozzi, L.; Pellegrini, E.; Guidi, L.; Landi, M.; Lorenzini, G.; Massai, R.; Remorini, D.; Tonelli, M.; Trivellini, A.; Vernieri, P.; et al. Losing the warning signal: Drought compromises the cross-talk of signaling molecules in Quercus ilex exposed to ozone. Front. Plant Sci. 2017, 8, 1020. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Cotrozzi, L.; Pellegrini, E.; Remorini, D.; Tonelli, M.; Trivellini, A.; Nali, C.; Guidi, L.; Massai, R.; Vernieri, P.; et al. When “thirsty” means “less able to activate the signalling wave trigged by a pulse of ozone”: A case of study in two Mediterranean deciduous oak species with different drought sensitivity. Sci. Total Environ. 2019, 657, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Lead (Pb)-induced biochemical and ultrastructural changes in wheat (Triticum aestivum) roots. Protoplasma 2013, 250, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Miura, K.; Tada, Y. Regulation of water, salinity, and cold stress responses by salicylic acid. Front. Plant Sci. 2014, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Mohsenzadeh, S.; Shahrtash, M.; Mohabatkar, H. Interactive effects of salicylic acid and silicon on some physiological responses of cadmium-stressed maize seedlings. Iranian J. Sci. Tech. (Sciences) 2011, 35, 57–60. [Google Scholar]
- Vlot, A.C.; Dempsey, D.M.A.; Klessig, D.F. Salicylic acid, a multifaceted hormone to combat disease. Ann. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Per, T.S.; Khan, N.A. Salicylic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Sign. Behav. 2013, 8, e26374. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Balbaa, L.; Gaballah, M. Salicylic acid and salinity effects on growth of maize plants. Res. J. Agricul. Biol. Sci. 2007, 3, 321–328. [Google Scholar]
- Khokon, M.A.R.; Okuma, E.; Hossain, M.A.; Munemasa, S.; Uraji, M.; Nakamura, Y.; Mori, I.C.; Murata, Y. Involvement of extracellular oxidative burst in salicylic acid-induced stomatal closure in Arabidopsis. Plant Cell Environ. 2011, 34, 434–443. [Google Scholar] [CrossRef]
- Larque-Saavedra, A. Stomatal closure in response to acetylsalicylic acid treatment. Zeitschrift für Pflanzenphysiologie 1979, 93, 371–375. [Google Scholar] [CrossRef]
- Larque-Saavedra, A. The antiranspirant effect of acetylsalcylic acid on Phaseolus vulgaris. Physiol. Plant. 1978, 43, 126–128. [Google Scholar] [CrossRef]
- Alaey, M.; Babalar, M.; Naderi, R.; Kafi, M. Effect of pre-and postharvest salicylic acid treatment on physio-chemical attributes in relation to vase-life of rose cut flowers. Postharvest Biol. Technol. 2011, 61, 91–94. [Google Scholar] [CrossRef]
- Divi, U.K.; Rahman, T.; Krishna, P. Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biol. 2010, 10, 151. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, J.; Qin, G.; Cui, H.; Wang, Q. Salicylic acid analogues with biological activity may induce chilling tolerance of maize (Zea mays) seeds. Botany 2012, 90, 845–855. [Google Scholar] [CrossRef]
- Zengin, F. Effects of exogenous salicylic acid on growth characteristics and biochemical content of wheat seeds under arsenic stress. J. Environ. Biol. 2015, 36, 249. [Google Scholar]
- Ghani, A.; Khan, I.; Ahmed, I.; Mustafa, I.; Abd-Ur, R.; Muhammad, N. Amelioration of lead toxicity in Pisum sativum (L.) by foliar application of salicylic acid. J. Environ. Anal. Toxicol 2015, 5, 10–4172. [Google Scholar]
- Gondor, O.K.; Pál, M.; Darkó, É.; Janda, T.; Szalai, G. Salicylic acid and sodium salicylate alleviate cadmium toxicity to different extents in maize (Zea mays L.). PLoS ONE 2016, 11, e0160157. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.S.; Guo, K.; Elbaz, A.A.; Yang, Z.M. Salicylic acid alleviates mercury toxicity by preventing oxidative stress in roots of Medicago sativa. Environ. Exp. Bot. 2009, 65, 27–34. [Google Scholar] [CrossRef]
- Ahmad, P.; Nabi, G.; Ashraf, M. Cadmium-induced oxidative damage in mustard [Brassica juncea (L.) Czern. & Coss.] plants can be alleviated by salicylic acid. South Afr. J. Bot. 2011, 77, 36–44. [Google Scholar]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Riaz, M.; Shahzad, S.M.; Imran, Q.; Ali, I. Combined ability of chromium (Cr) tolerant plant growth promoting bacteria (PGPB) and salicylic acid (SA) in attenuation of chromium stress in maize plants. Plant Physiol. Biochem. 2016, 108, 456–467. [Google Scholar] [CrossRef]
- Song, W.Y.; Yang, H.C.; Shao, H.B.; Zheng, A.Z.; Brestic, M. The alleviative effects of salicylic acid on the activities of catalase and superoxide dismutase in malting barley (Hordeum uhulgare L.) seedling leaves stressed by heavy metals. CLEAN–Soil, Air, Water 2014, 42, 88–97. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Sami, F.; Siddiqui, H.; Hayat, S. Supplementation of salicylic acid and citric acid for alleviation of cadmium toxicity to Brassica juncea. J. Plant Growth Regul. 2019, 1–15. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Matin, M.A.; Fardus, J.; Hasanuzzaman, M.; Hossain, M.S.; Parvin, K. Foliar application of salicylic acid improves growth and yield attributes by upregulating the antioxidant defense system in Brassica campestris plants grown in lead-amended soils. Acta Agrobot. 2019, 72. [Google Scholar] [CrossRef]
- Lu, Q.; Zhang, T.; Zhang, W.; Su, C.; Yang, Y.; Hu, D.; Xu, Q. Alleviation of cadmium toxicity in Lemna minor by exogenous salicylic acid. Ecotox. Environ. Saf. 2018, 147, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.-S.; Yang, Y.-H.; Shao, Y.-F.; Wu, K.-W.; Liu, Z.-L. The effects of exogenous salicylic acid on alleviating cadmium toxicity in Nymphaea tetragona Georgi. S. Afr. J. Bot. 2018, 114, 267–271. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G.; Wang, Y.; Yang, D.; Guan, C.; Ji, J. Foliar application of salicylic acid alleviate the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotox. Environ. Saf. 2019, 172, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Tajti, J.; Németh, E.; Glatz, G.; Janda, T.; Pál, M. Pattern of changes in salicylic acid-induced protein kinase (SIPK) gene expression and salicylic acid accumulation in wheat under cadmium exposure. Plant Biol. 2019, 21, 1176–1180. [Google Scholar] [CrossRef]
- Ahmad, B.; Jaleel, H.; Sadiq, Y.; Khan, M.M.A.; Shabbir, A. Response of exogenous salicylic acid on cadmium induced photosynthetic damage, antioxidant metabolism and essential oil production in peppermint. Plant Growth Regul. 2018, 86, 273–286. [Google Scholar] [CrossRef]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Role of salicylic acid and hydrogen sulfide in promoting lead stress tolerance and regulating free amino acid composition in Zea mays L. Acta Physiol. Plant. 2019, 41, 94. [Google Scholar] [CrossRef]
- Alamri, S.A.D.; Siddiqui, M.H.; Al-Khaishany, M.Y.; Ali, H.M.; Al-Amri, A.; AlRabiah, H.K. Exogenous application of salicylic acid improves tolerance of wheat plants to lead stress. Adv. Agric. Sci. 2018, 6, 25–35. [Google Scholar]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Impacts of seed priming with salicylic acid and sodium hydrosulfide on possible metabolic pathway of two amino acids in maize plant under lead stress. Mol. Biol. Res. Commun. 2018, 7, 83. [Google Scholar]
- Mabrouk, B.; Kâab, S.; Rezgui, M.; Majdoub, N.; da Silva, J.T.; Kâab, L. Salicylic acid alleviates arsenic and zinc toxicity in the process of reserve mobilization in germinating fenugreek (Trigonella foenum-graecum L.) seeds. S. Afr. J. Bot. 2019, 124, 235–243. [Google Scholar] [CrossRef]
- Kumari, A.; Pandey, N.; Pandey-Rai, S. Exogenous salicylic acid-mediated modulation of arsenic stress tolerance with enhanced accumulation of secondary metabolites and improved size of glandular trichomes in Artemisia annua L. Protoplasma 2018, 255, 139–152. [Google Scholar] [CrossRef]
- Kumari, A.; Pandey-Rai, S. Enhanced arsenic tolerance and secondary metabolism by modulation of gene expression and proteome profile in Artemisia annua L. after application of exogenous salicylic acid. Plant Physiol. Biochem. 2018, 132, 590–602. [Google Scholar] [CrossRef]
- Saidi, I.; Yousfi, N.; Borgi, M.A. Salicylic acid improves the antioxidant ability against arsenic-induced oxidative stress in sunflower (Helianthus annuus) seedling. J. Plant Nutr. 2017, 40, 2326–2335. [Google Scholar] [CrossRef]
- Singh, A.P.; Dixit, G.; Kumar, A.; Mishra, S.; Kumar, N.; Dixit, S.; Singh, P.K.; Dwivedi, S.; Trivedi, P.K.; Pandey, V. A protective role for nitric oxide and salicylic acid for arsenite phytotoxicity in rice (Oryza sativa L.). Plant Physiol. Biochem. 2017, 115, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Sihag, S.; Brar, B.; Joshi, U. Salicylic acid induces amelioration of chromium toxicity and affects antioxidant enzyme activity in Sorghum bicolor L. Int. J. Phytorem. 2019, 21, 293–304. [Google Scholar] [CrossRef]
- Gill, R.A.; Zhang, N.; Ali, B.; Farooq, M.A.; Xu, J.; Gill, M.B.; Mao, B.; Zhou, W. Role of exogenous salicylic acid in regulating physio-morphic and molecular changes under chromium toxicity in black-and yellow-seeded Brassica napus L. Environ. Sci. Poll. Res. 2016, 23, 20483–20496. [Google Scholar] [CrossRef]
- Huda, A.N.; Swaraz, A.; Reza, M.A.; Haque, M.A.; Kabir, A.H. Remediation of chromium toxicity through exogenous salicylic acid in rice (Oryza sativa L.). Water Air Soil Poll. 2016, 227, 278. [Google Scholar] [CrossRef]
- Zaid, A.; Mohammad, F.; Wani, S.H.; Siddique, K.M. Salicylic acid enhances nickel stress tolerance by up-regulating antioxidant defense and glyoxalase systems in mustard plants. Ecotoxicol. Environ. Saf. 2019, 180, 575–587. [Google Scholar] [CrossRef]
- Kotapati, K.V.; Palaka, B.K.; Ampasala, D.R. Alleviation of nickel toxicity in finger millet (Eleusine coracana L.) germinating seedlings by exogenous application of salicylic acid and nitric oxide. Crop J. 2017, 5, 240–250. [Google Scholar] [CrossRef]
- Soltani Maivan, E.; Radjabian, T.; Abrishamchi, P.; Talei, D. Physiological and biochemical responses of Melissa officinalis L. to nickel stress and the protective role of salicylic acid. Arch. Agron. Soil Sci. 2017, 63, 330–343. [Google Scholar] [CrossRef]
- Karimi, N.; Ghasempour, H.-R. Salicylic acid and jasmonic acid restrains nickel toxicity by ameliorating antioxidant defense system in shoots of metallicolous and non-metallicolous Alyssum inflatum Náyr. Populations. Plant Physiol. Biochem. 2019, 135, 450–459. [Google Scholar]
- Mei, L.; Daud, M.; Ullah, N.; Ali, S.; Khan, M.; Malik, Z.; Zhu, S. Pretreatment with salicylic acid and ascorbic acid significantly mitigate oxidative stress induced by copper in cotton genotypes. Environ. Sci. Poll. Res. 2015, 22, 9922–9931. [Google Scholar] [CrossRef] [PubMed]
- Moravcová, Š.; Tůma, J.; Dučaiová, Z.K.; Waligórski, P.; Kula, M.; Saja, D.; Słomka, A.; Bąba, W.; Libik-Konieczny, M. Influence of salicylic acid pretreatment on seeds germination and some defence mechanisms of Zea mays plants under copper stress. Plant Physiol. Biochem. 2018, 122, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Ventrella, A.; Catucci, L.; Piletska, E.; Piletsky, S.; Agostiano, A. Interactions between heavy metals and photosynthetic materials studied by optical techniques. Bioelectrochemistry 2009, 77, 19–25. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Babu, N.G.; Sarma, P.A.; Attitalla, I.H.; Murthy, S. Effect of selected heavy metal ions on the photosynthetic electron transport and energy transfer in the thylakoid membrane of the cyanobacterium, Spirulina platensis. Acad. J. Plant Sci. 2010, 3, 46–49. [Google Scholar]
- Chugh, L.K.; Sawhney, S.K. Photosynthetic activities of Pisum sativum seedlings grown in presence of cadmium. Plant Physiol. Biochem. 1999, 37, 297–303. [Google Scholar] [CrossRef]
- Khan, N.; Samiullah Singh, S.; Nazar, R. Activities of antioxidative enzymes, sulphur assimilation, photosynthetic activity and growth of wheat (Triticum aestivum) cultivars differing in yield potential under cadmium stress. J. Agron. Crop Sci. 2007, 193, 435–444. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Varshney, P.; Ahmad, A. Salicylic acid minimizes nickel and/or salinity-induced toxicity in Indian mustard (Brassica juncea) through an improved antioxidant system. Environ. Sci. Poll. Res. 2012, 19, 8–18. [Google Scholar] [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef]
- Moussa, H.; El-Gamal, S.M. Effect of salicylic acid pretreatment on cadmium toxicity in wheat. Biol. Plant. 2010, 54, 315–320. [Google Scholar] [CrossRef]
- Parashar, A.; Yusuf, M.; Fariduddin, Q.; Ahmad, A. Salicylic acid enhances antioxidant system in Brassica juncea grown under different levels of manganese. Int. J. Biol. Macromol. 2014, 70, 551–558. [Google Scholar] [CrossRef]
- Yadav, S. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Yang, S.; Chen, Y. Salicylic acid alleviates cadmium-induced inhibition of growth and photosynthesis through upregulating antioxidant defense system in two melon cultivars (Cucumis melo L.). Protoplasma 2015, 252, 911–924. [Google Scholar] [CrossRef]
- Chao, Y.-Y.; Chen, C.-Y.; Huang, W.-D.; Kao, C.H. Salicylic acid-mediated hydrogen peroxide accumulation and protection against Cd toxicity in rice leaves. Plant Soil 2010, 329, 327–337. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Zouari, M.; Ahmed, C.B.; Elloumi, N.; Bellassoued, K.; Delmail, D.; Labrousse, P.; Abdallah, F.B.; Rouina, B.B. Impact of proline application on cadmium accumulation, mineral nutrition and enzymatic antioxidant defense system of Olea europaea L. cv Chemlali exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2016, 128, 195–205. [Google Scholar] [CrossRef]
- Zouari, M.; Ahmed, C.B.; Zorrig, W.; Elloumi, N.; Rabhi, M.; Delmail, D.; Rouina, B.B.; Labrousse, P.; Abdallah, F.B. Exogenous proline mediates alleviation of cadmium stress by promoting photosynthetic activity, water status and antioxidative enzymes activities of young date palm (Phoenix dactylifera L.). Ecotoxicol. Environ. Saf. 2016, 128, 100–108. [Google Scholar] [CrossRef]
- Misra, N.; Saxena, P. Effect of salicylic acid on proline metabolism in lentil grown under salinity stress. Plant Sci. 2009, 177, 181–189. [Google Scholar] [CrossRef]
- Zengin, F.K.; Munzuroglu, O. Effects of some heavy metals on content of chlorophyll, proline and some antioxidant chemicals in bean (Phaseolus vulgaris L.) seedlings. Acta Biol. Cracov. Bot. 2005, 47, 157–164. [Google Scholar]
- Chen, C.; Dickman, M.B. Proline suppresses apoptosis in the fungal pathogen Colletotrichum trifolii. Proc. Natl. Acad. Sci. USA 2005, 102, 3459–3464. [Google Scholar] [CrossRef] [PubMed]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Modulation of growth and oxidative stress by seed priming with salicylic acid in Zea mays L. under lead stress. J. Plant Inter. 2019, 14, 369–375. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M.; Tran, L.-S.P. Nitric oxide mediates hydrogen peroxide-and salicylic acid-induced salt tolerance in rice (Oryza sativa L.) seedlings. Plant Growth Regul. 2015, 77, 265–277. [Google Scholar] [CrossRef]
- Raza, S.; Aown, M.; Saleem, M.F.; Jamil, M.; Khan, I.H. Impact of foliar applied glycinebetaine on growth and physiology of wheat (Triticum aestivum L.) under drought conditions. Pak. J. Agric. Sci. 2014, 51, 327–334. [Google Scholar]
- Jabeen, N.; Abbas, Z.; Iqbal, M.; Rizwan, M.; Jabbar, A.; Farid, M.; Ali, S.; Ibrahim, M.; Abbas, F. Glycinebetaine mediates chromium tolerance in mung bean through lowering of Cr uptake and improved antioxidant system. Arch. Agron. Soil Sci. 2016, 62, 648–662. [Google Scholar] [CrossRef]
- Cao, F.; Liu, L.; Ibrahim, W.; Cai, Y.; Wu, F. Alleviating effects of exogenous glutathione, glycinebetaine, brassinosteroids and salicylic acid on cadmium toxicity in rice seedlings (Oryza sativa). Agrotechnology 2013, 2, 107–112. [Google Scholar] [CrossRef]
- Aldesuquy, H.S.; Abbas, M.A.; Abo-Hamed, S.A.; Elhakem, A.H. Does glycine betaine and salicylic acid ameliorate the negative effect of drought on wheat by regulating osmotic adjustment through solutes accumulation? J. Stress Physol. Biochem. 2013, 9, 5–22. [Google Scholar]
- Misra, N.; Misra, R. Salicylic acid changes plant growth parameters and proline metabolism in Rauwolfia serpentina leaves grown under salinity stress. Am-Eurasian J. Agric. Environ. Sci. 2012, 12, 1601–1609. [Google Scholar]
- Keunen, E.; Peshev, D.; Vangronsveld, J.; Van Den Ende, W.; Cuypers, A. Plant sugars are crucial players in the oxidative challenge during abiotic stress: Extending the traditional concept. Plant Cell Environ. 2013, 36, 1242–1255. [Google Scholar] [CrossRef] [PubMed]
- Salerno, G.L.; Curatti, L. Origin of sucrose metabolism in higher plants: When, how and why? Trends Plant Sci. 2003, 8, 63–69. [Google Scholar] [CrossRef]
- Muller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-J.; Yang, S.-H.; Hsu, C.-S. Synthesis of conjugated polymers for organic solar cell applications. Chem. Rev. 2009, 109, 5868–5923. [Google Scholar] [CrossRef] [PubMed]
- Van den Ende, W.; Peshev, D. Sugars as antioxidants in plants. In Crop Improvement under Adverse Conditions; Springer: Dordrecht, The Netherlands, 2013; pp. 285–307. [Google Scholar]
- Luo, Y.; Su, Z.; Bi, T.; Cui, X.; Lan, Q. Salicylic acid improves chilling tolerance by affecting antioxidant enzymes and osmoregulators in sacha inchi (Plukenetia volubilis). Braz. J. Bot. 2014, 37, 357–363. [Google Scholar] [CrossRef]
- El-Tayeb, M.; El-Enany, A.; Ahmed, N. Salicylic acid-induced adaptive response to copper stress in sunflower (Helianthus annuus L.). Plant Growth Regul. 2006, 50, 191–199. [Google Scholar] [CrossRef]
- Wasti, S.; Mimouni, H.; Smiti, S.; Zid, E.; Ben Ahmed, H. Enhanced salt tolerance of tomatoes by exogenous salicylic acid applied through rooting medium. OMICS 2012, 16, 200–207. [Google Scholar] [CrossRef]
- Nahar, K.; Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Suzuki, T.; Fujita, M. Polyamine and nitric oxide crosstalk: Antagonistic effects on cadmium toxicity in mung bean plants through upregulating the metal detoxification, antioxidant defense and methylglyoxal detoxification systems. Ecotoxicol. Environ. Saf. 2016, 126, 245–255. [Google Scholar] [CrossRef]
- Benavides, M.P.; Groppa, M.D.; Recalde, L.; Verstraeten, S.V. Effects of polyamines on cadmium-and copper-mediated alterations in wheat (Triticum aestivum L.) and sunflower (Helianthus annuus L.) seedling membrane fluidity. Arch. Biochem. Biophys. 2018, 654, 27–39. [Google Scholar] [CrossRef]
- Groppa, M.D.; Benavides, M.P.; Tomaro, M.L. Polyamine metabolism in sunflower and wheat leaf discs under cadmium or copper stress. Plant Sci. 2003, 164, 293–299. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M. Regulatory role of polyamines in growth, development and abiotic stress tolerance in plants. In Plant Adaptation to Environmental Change: Significance of Amino Acids and Their Derivatives; MD University: Haryana, India, 2014; pp. 157–193. [Google Scholar]
- Liu, J.-H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [PubMed]
- Szepesi, Á. Interaction between salicylic acid and polyamines and their possible roles in tomato hardening processes. Acta Biol. Szeged. 2011, 55, 165–166. [Google Scholar]
- Tajti, J.; Janda, T.; Majláth, I.; Szalai, G.; Pál, M. Comparative study on the effects of putrescine and spermidine pre-treatment on cadmium stress in wheat. Ecotoxicol. Environ. Saf. 2018, 148, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Yadav, P.; Sharma, A.; Thukral, A.K.; Kumar, V.; Kohli, S.K.; Bhardwaj, R. Castasterone and citric acid treatment restores photosynthetic attributes in Brassica juncea L. under Cd (II) toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Interaction of 24-epibrassinolide and salicylic acid regulates pigment contents, antioxidative defense responses, and gene expression in Brassica juncea L. seedlings under Pb stress. Environ. Sci. Poll. Res. 2018, 25, 15159–15173. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia obovata under Cd and Zn stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef]
- Zafari, S.; Sharifi, M.; Chashmi, N.A.; Mur, L.A. Modulation of Pb-induced stress in Prosopis shoots through an interconnected network of signaling molecules, phenolic compounds and amino acids. Plant Physiol. Biochem. 2016, 99, 11–20. [Google Scholar] [CrossRef]
- Nakamura, M.; Takeuchi, Y.; Miyanaga, K.; Seki, M.; Furusaki, S. High anthocyanin accumulation in the dark by strawberry (Fragaria ananassa) callus. Biotechnol. Lett. 1999, 21, 695–699. [Google Scholar] [CrossRef]
- Narayan, M.; Thimmaraju, R.; Bhagyalakshmi, N. Interplay of growth regulators during solid-state and liquid-state batch cultivation of anthocyanin producing cell line of Daucus carota. Process Biochem. 2005, 40, 351–358. [Google Scholar] [CrossRef]
- Dong, J.; Wan, G.; Liang, Z. Accumulation of salicylic acid-induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture. J. Biotechnol. 2010, 148, 99–104. [Google Scholar] [CrossRef]
- Kováčik, J.; Gruz, J.; Hedbavny, J.; Klejdus, B.I.; Strnad, M. Cadmium and nickel uptake are differentially modulated by salicylic acid in Matricaria chamomilla plants. J. Agric. Food Chem. 2009, 57, 9848–9855. [Google Scholar] [CrossRef] [PubMed]
- Horváth, E.; Szalai, G.; Janda, T. Induction of abiotic stress tolerance by salicylic acid signaling. J. Plant Growth Regul. 2007, 26, 290–300. [Google Scholar] [CrossRef]
- Hettiarachchi, G.H.; Reddy, M.K.; Sopory, S.K.; Chattopadhyay, S. Regulation of TOP2 by various abiotic stresses including cold and salinity in pea and transgenic tobacco plants. Plant Cell Physiol. 2005, 46, 1154–1160. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Singh, B.; Mishra, R.; Agarwal, P.K.; Goswami, M.; Nair, S.; Sopory, S.; Reddy, M. A pea chloroplast translation elongation factor that is regulated by abiotic factors. Biochem. Byophys. Res. Commun. 2004, 320, 523–530. [Google Scholar] [CrossRef]
- Kovács, V.; Gondor, O.K.; Szalai, G.; Darkó, É.; Majláth, I.; Janda, T.; Pál, M. Synthesis and role of salicylic acid in wheat varieties with different levels of cadmium tolerance. J. Hazard. Mater. 2014, 280, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant. Sci. 2015, 6, 462. [Google Scholar] [CrossRef]
- Arfan, M. Exogenous application of salicylic acid through rooting medium modulates ionaccumulation and antioxidant activity in spring wheat under salt stress. Int. J. Agric. Biol. 2009, 11, 437–442. [Google Scholar]
- Song, W.; Zheng, A.; Shao, H.B.; Chu, L.; Brestic, M.; Zhang, Z. The alleviative effect of salicylic acid on the physiological indices of the seedling leaves in six different wheat genotypes under lead stress. Plant Omics J. 2012, 5, 486–493. [Google Scholar]
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of salt-induced photosynthesis and growth inhibition by salicylic acid involves glycinebetaine and ethylene in mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Terrile, M.C.; Casalongué, C.A. Auxin and salicylic acid signalings counteract the regulation of adaptive responses to stress. Plant Signal. Behav. 2011, 6, 452–454. [Google Scholar] [CrossRef]
- Jiang, C.-J.; Shimono, M.; Sugano, S.; Kojima, M.; Liu, X.; Inoue, H.; Sakakibara, H.; Takatsuji, H. Cytokinins act synergistically with salicylic acid to activate defense gene expression in rice. Mol. Plant. Microbe. Interact. 2013, 26, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Roghayyeh, S.; Saeede, R.; Omid, A.; Mohammad, S. The effect of salicylic acid and gibberellin on seed reserve utilization, germination and enzyme activity of sorghum (Sorghum bicolor L.) seeds under drought stress. J. Stress Physiol. Biochem. 2014, 10, 5–13. [Google Scholar]
- Jiang, C.-J.; Shimono, M.; Sugano, S.; Kojima, M.; Yazawa, K.; Yoshida, R.; Inoue, H.; Hayashi, N.; Sakakibara, H.; Takatsuji, H. Abscisic acid interacts antagonistically with salicylic acid signaling pathway in rice–Magnaporthe grisea interaction. Mol. Plant. Microbe. Interact. 2010, 23, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Agtuca, B.; Rieger, E.; Hilger, K.; Song, L.; Robert, C.A.; Erb, M.; Karve, A.; Ferrieri, R.A. Carbon-11 reveals opposing roles of auxin and salicylic acid in regulating leaf physiology, leaf metabolism, and resource allocation patterns that impact root growth in Zea mays. J. Plant Growth Regul. 2014, 33, 328–339. [Google Scholar] [CrossRef]
- Thao, N.P.; Khan, M.I.R.; Thu, N.B.A.; Hoang, X.L.T.; Asgher, M.; Khan, N.A.; Tran, L.-S.P. Role of ethylene and its cross talk with other signaling molecules in plant responses to heavy metal stress. Plant Physiol. 2015, 169, 73–84. [Google Scholar] [CrossRef]
- Shakirova, F.; Allagulova, C.R.; Maslennikova, D.; Klyuchnikova, E.; Avalbaev, A.; Bezrukova, M. Salicylic acid-induced protection against cadmium toxicity in wheat plants. Environ. Exp. Bot. 2016, 122, 19–28. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, M.I.R.; Anjum, N.A.; Masood, A.; Hussain, S.J.; Khan, N.A. Jasmonates in plants under abiotic stresses: Crosstalk with other phytohormones matters. Environ. Exp. Bot. 2018, 145, 104–120. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Khan, N.A. Salicylic acid and jasmonates: Approaches in abiotic stress tolerance. J. Plant Biochem. Physiol. 2013, 1, e113. [Google Scholar] [CrossRef]
- Petersen, M.; Brodersen, P.; Naested, H.; Andreasson, E.; Lindhart, U.; Johansen, B.; Nielsen, H.B.; Lacy, M.; Austin, M.J.; Parker, J.E.; et al. Arabidopsis MAP kinase 4 negatively regulates systemic acquired resistance. Cell 2000, 103, 1111–1120. [Google Scholar] [CrossRef]
- Engelberth, J.; Viswanathan, S.; Engelberth, M.J. Low concentrations of salicylic acid stimulate insect elicitor responses in Zea mays seedlings. J. Chem. Ecol. 2011, 37, 263–266. [Google Scholar] [CrossRef]
- Rostás, M.; Winter, T.R.; Borkowski, L.; Zeier, J. Copper and herbivory lead to priming and synergism in phytohormones and plant volatiles in the absence of salicylate-jasmonate antagonism. Plant Signal. Behav. 2013, 8, e24264. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Ram, H.; Sarlach, R.S. Enhancing seed yield and quality of Egyptian clover (Trifolium alexandrinum L.) with foliar application of bio-regulators. Field Crops Res. 2013, 146, 25–30. [Google Scholar] [CrossRef]
- Canakci, S. Effects of salicylic acid on growth, biochemical constituents in pepper (Capsicum annuum L.) seedlings. Pak. J. Biol. Sci. 2011, 14, 300. [Google Scholar] [CrossRef] [PubMed][Green Version]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).