
Fragaria Genus: Chemical Composition and Biological Activities

 ,
, 
 and
and
Abstract
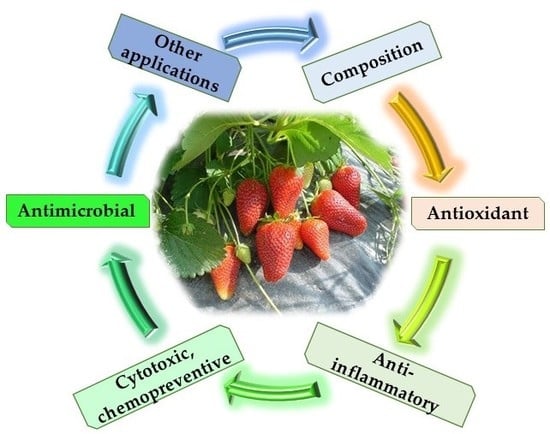
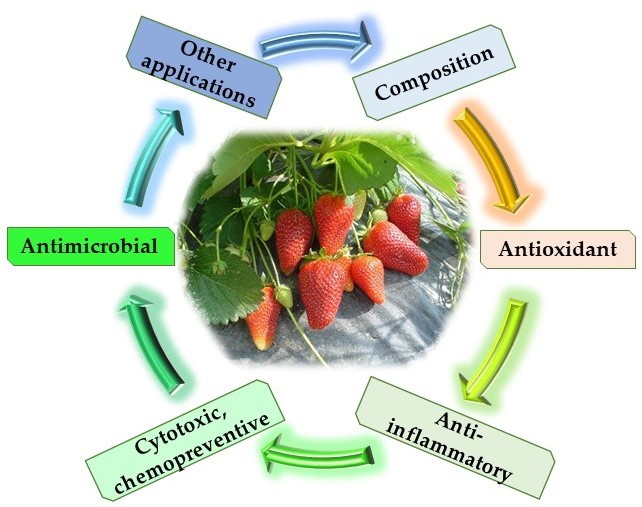
1. Introduction
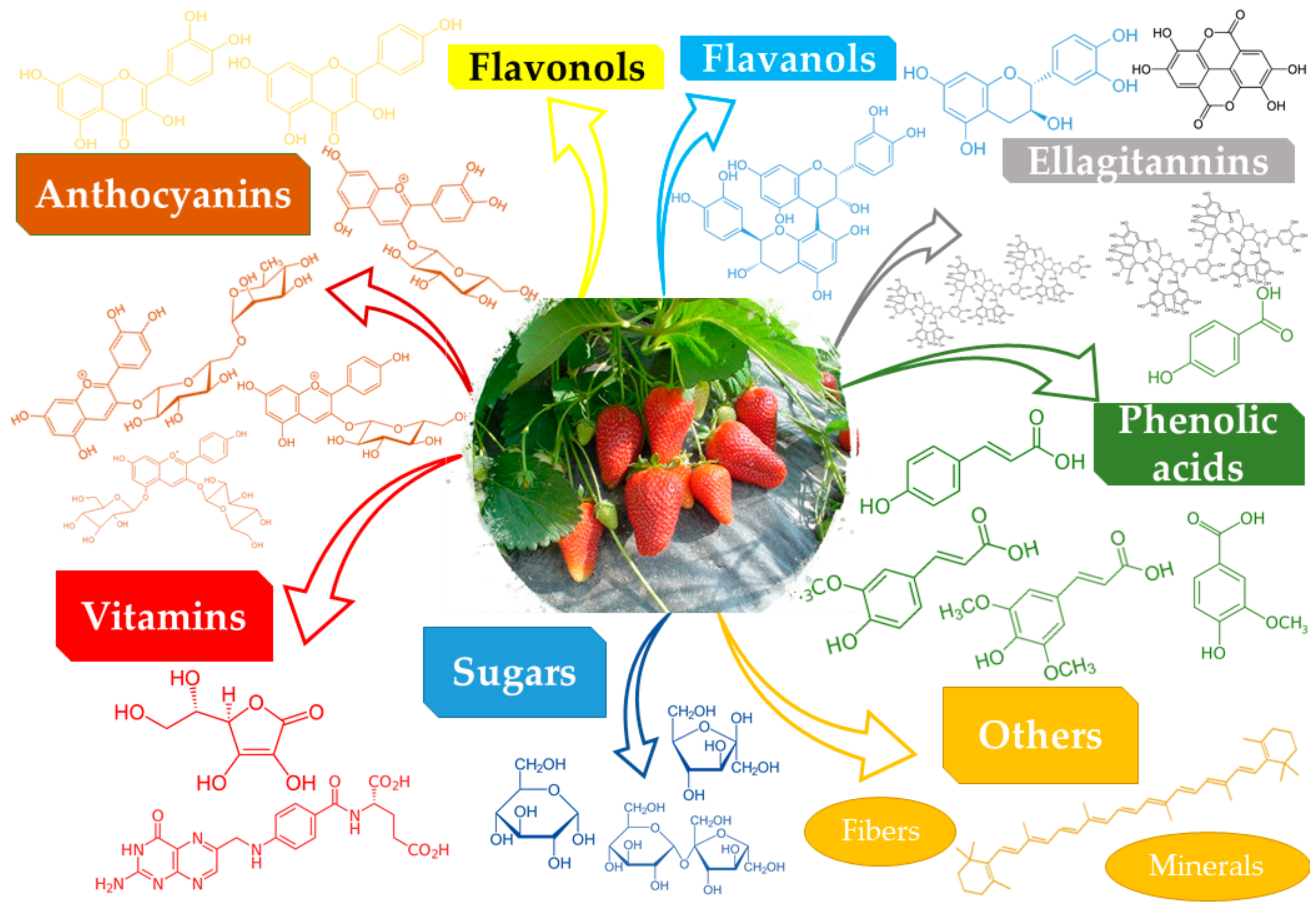
2. Composition of Fragaria L. Genus
3. Biological Activities of Fragaria Genus
3.1. Antioxidant Properties
3.2. Anti-Inflammatory Properties
3.3. Other Potential Applications
4. Current Limitations and Future Perspectives
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liston, A.; Cronn, R.; Ashman, T.L. Fragaria: A genus with deep historical roots and ripe for evolutionary and ecological insights. Am. J. Bot. 2014, 101, 1686–1699. [Google Scholar] [CrossRef]
- Awad, M.A.; De Jager, A. Influences of air and controlled atmosphere storage on the concentration of potentially healthful phenolics in apples and other fruits. Postharv. Biol. Technol. 2003, 27, 53–58. [Google Scholar] [CrossRef]
- Pauketat, T.R.; Kelly, L.S.; Fritz, G.J.; Lopinot, N.H.; Elias, S.; Hargrave, E. The residues of feasting and public ritual at Early Cahokia. Amer. Antiq. 2002, 67, 257–279. [Google Scholar] [CrossRef]
- Finn, C.E.; Retamales, J.B.; Lobos, G.A.; Hancock, J.F. The Chilean strawberry (Fragaria chiloensis): Over 1000 years of domestication. HortScience 2013, 48, 418–421. [Google Scholar] [CrossRef]
- Duchesne, A.N. Histoire Naturelle des Fraisiers Contenant les Vues d’économie Réunies à la Botanique et Suivie de Remarques Particulières sur Plusieurs Points qui ont Rapport à l’Histoire Naturelle Générale; Didot le Jeune: Paris, France, 1766. [Google Scholar]
- Plants of the World Online: Fragaria L. Available online: http://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:30014957-2 (accessed on 3 January 2020).
- The Plant List: Fragaria. Available online: http://www.theplantlist.org/1.1/browse/A/Rosaceae/Fragaria/ (accessed on 3 January 2020).
- Petran, A.J. Performance and impact of strawberry (Fragaria x ananassa) season extension in the United States Upper Midwest using organic practices. Ph.D. Thesis, University of Minnesota, Minneapolis, MN, USA, April 2016. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). 2017. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 3 January 2020).
- Sato, T.; Ikeya, Y.; Adachi, S.I.; Yagasaki, K.; Nihei, K.I.; Itoh, N. Extraction of strawberry leaves with supercritical carbon dioxide and entrainers: Antioxidant capacity, total phenolic content, and inhibitory effect on uric acid production of the extract. Food Bioprod. Process 2019, 117, 160–169. [Google Scholar] [CrossRef]
- Giampieri, F.; Tulipani, S.; Alvarez-Suarez, J.M.; Quiles, J.L.; Mezzetti, B.; Battino, M. The strawberry: Composition, nutritional quality, and impact on human health. Nutrition 2012, 28, 9–19. [Google Scholar] [CrossRef]
- Morales-Quintana, L.; Ramos, P. Chilean strawberry (Fragaria chiloensis): An integrative and comprehensive review. Food Res. Int. 2019, 119, 769–776. [Google Scholar] [CrossRef]
- Jimenez-Garcia, S.N.; Guevara-Gonzalez, R.G.; Miranda-Lopez, R.; Feregrino-Perez, A.A.; Torres-Pacheco, I.; Vazquez-Cruz, M.A. Functional properties and quality characteristics of bioactive compounds in berries: Biochemistry, biotechnology, and genomics. Food Res. Int. 2013, 54, 1195–1207. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Vuong, Q.V.; Hirun, S.; Phillips, P.A.; Chuen, T.L.; Bowyer, M.C.; Goldsmith, C.D.; Scarlett, C.J. Fruit-derived phenolic compounds and pancreatic cancer: Perspectives from Australian native fruits. J. Ethnopharmacol. 2014, 152, 227–242. [Google Scholar] [CrossRef]
- Khan, N.; Syed, D.N.; Ahmad, N.; Mukhtar, H. Fisetin: A dietary antioxidant for health promotion. Antioxid. Redox Signal. 2013, 2, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Cerezo, A.B.; Cuevas, E.; Winterhalter, P.; Garcia-Parrilla, M.C.; Troncoso, A.M. Isolation, identification, and antioxidant activity of anthocyanin compounds in Camarosa strawberry. Food Chem. 2010, 123, 574–582. [Google Scholar] [CrossRef]
- Crecente-Campo, J.; Nunes-Damaceno, M.; Romero-Rodrıguez, M.A.; Vazquez-Oderiz, M.L. Color, anthocyanin pigment, ascorbic acid and total phenolic compound determination in organic versus conventional strawberries (Fragaria x ananassa Duch, cv Selva). J. Food Compos. Anal. 2012, 28, 23–30. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, Y.; Tang, X.; Jin, W.; Han, Z. Differences in volatile ester composition between Fragaria×ananassa and F. vesca and implications for strawberry aroma patterns. Sci. Horticult. 2013, 150, 47–53. [Google Scholar] [CrossRef]
- Yang, D.; Xie, H.; Jiang, Y.; Wei, X. Phenolics from strawberry cv. Falandi and their antioxidant and α-glucosidase inhibitory activities. Food Chem. 2016, 194, 857–863. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Wu, B.; Liu, W.; Archbold, D.D. Comparative analyses of polyphenolic composition of Fragaria spp. Color mutants. Plant Physiol. Biochem. 2018, 125, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Alici, E.H.; Arabaci, G. A novel serine protease from strawberry (Fragaria ananassa): Purification and biochemical characterization. Int. J. Biol. Macromol. 2018, 114, 1295–1304. [Google Scholar] [CrossRef]
- Aceituno-Valenzuela, U.; Covarrubias, M.P.; Aguayo, M.F.; Valenzuela-Riffo, F.; Espinoza, A.; Gaete-Eastman, C.; Herrera, R.; Handford, M.; Norambuena, L. Identification of a type II cystatin in Fragaria chiloensis: A proteinase inhibitor differentially regulated during achene development and in response to biotic stress-related stimuli. Plant Physiol. Biochem. 2018, 129, 158–167. [Google Scholar] [CrossRef]
- Karlińska, E.; Pecio, L.; Macierzyński, J.; Stochmal, A.; Kosmala, M. Structural elucidation of the ellagitannin with a molecular weight of 2038 isolated from strawberry fruit (Fragaria ananassa Duch.) and named fragariin A. Food Chem. 2019, 296, 109–115. [Google Scholar] [CrossRef]
- Nizioł, J.; Misiorek, M.; Ruman, T. Mass spectrometry imaging of low molecular weight metabolites in strawberry fruit (Fragaria x ananassa Duch.) cv. Primoris with 109Ag nanoparticle enhanced target. Phytochemistry 2019, 159, 11–19. [Google Scholar]
- Simirgiotis, M.J.; Schmeda-Hirschmann, G. Determination of phenolic composition and antioxidant activity in fruits, rhizomes and leaves of the white strawberry (Fragaria chiloensis spp. Chiloensis form chiloensis) using HPLC-DAD–ESI-MS and free radical quenching techniques. J. Food Compos. Anal. 2010, 23, 545–553. [Google Scholar] [CrossRef]
- Singh, A.; Singh, B.K.; Deka, B.C.; Sanwal, S.K.; Patel, R.K.; Verma, M.R. The genetic variability, inheritance and inter-relationships of ascorbic acid, β-carotene, phenol and anthocyanin content in strawberry (Fragaria×ananassa Duch.). Sci. Horticult. 2011, 129, 86–90. [Google Scholar] [CrossRef]
- Pineli, L.L.O.; Moretti, C.L.; dos Santos, M.S.; Campos, A.B.; Brasileiro, A.V.; Cordova, A.C.; Chiarello, M.D. Antioxidants and other chemical and physical characteristics of two strawberry cultivars at different ripeness stages. J. Food Compos. Anal. 2011, 24, 11–16. [Google Scholar] [CrossRef]
- Aaby, K.; Mazur, S.; Nes, A.; Skrede, G. Phenolic compounds in strawberry (Fragaria x ananassa Duch.) fruits: Composition in 27 cultivars and changes during ripening. Food Chem. 2012, 132, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Mikulic-Petkovsek, M.; Slatnar, A.; Stampar, F.; Veberic, R. HPLC-MSn identification and quantification of flavonol glycosides in 28 wild and cultivated berry species. Food Chem. 2012, 135, 2138–2146. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, X.; Yang, T.; Slovin, J.; Chen, P. Profiling polyphenols of two diploid strawberry (Fragaria vesca) inbred lines using UHPLC-HRMSn. Food Chem. 2014, 146, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Veberic, R.; Slatnar, A.; Bizjak, J.; Stampar, F.; Mikulic-Petkovsek, M. Anthocyanin composition of different wild and cultivated berry species. LWT Food Sci. Technol. 2015, 60, 509–517. [Google Scholar] [CrossRef]
- Liberal, J.; Costa, G.; Carmo, A.; Vitorino, R.; Marques, C.; Domingues, M.R.; Domingues, P.; Goncalves, A.C.; Alves, R.; Sarmento-Ribeiro, A.B.; et al. Chemical characterization and cytotoxic potential of an ellagitannin-enriched fraction from Fragaria vesca leaves. Arab. J. Chem. 2015. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, D.S.; Kim, D.Y.; Chun, C. Variation of bioactive compounds content of 14 oriental strawberry cultivars. Food Chem. 2015, 184, 196–202. [Google Scholar] [CrossRef]
- Zhu, Q.; Nakagawa, T.; Kishikawa, A.; Ohnuki, K.; Shimizu, K. In vitro bioactivities and phytochemical profile of various parts of the strawberry (Fragaria × ananassa var. Amaou). J. Funct. Food 2015, 13, 38–49. [Google Scholar] [CrossRef]
- Fernández-Lara, R.; Gordillo, B.; Rodríguez-Pulido, F.J.; González-Miret, M.L.; del Villar-Martínez, A.A.; Dávila-Ortiz, G.; Heredia, F.J. Assessment of the differences in the phenolic composition and color characteristics of new strawberry (Fragaria x ananassa Duch.) cultivars by HPLC-MS and Imaging Tristimulus Colorimetry. Food Res. Int. 2015, 76, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Chavez, G.; Scampicchio, M.; Andreotti, C. Influence of the site altitude on strawberry phenolic composition and quality. Sci. Horticult. 2015, 192, 21–28. [Google Scholar] [CrossRef]
- D’Urso, G.; Maldini, M.; Pintore, G.; d’Aquino, L.; Montoro, P.; Pizza, C. Characterisation of Fragaria vesca fruit from Italy following a metabolomics approach through integrated mass spectrometry techniques. LWT Food Sci. Technol. 2016, 74, 387–395. [Google Scholar] [CrossRef]
- Chaves, V.C.; Calvete, E.; Reginatto, F.H. Quality properties and antioxidant activity of seven strawberry (Fragaria x ananassa Duch) cultivars. Sci. Horticult. 2017, 225, 293–298. [Google Scholar] [CrossRef]
- Urrutia, M.; Rambla, J.L.; Alexiou, K.G.; Granell, A.; Monfort, A. Genetic analysis of the wild strawberry (Fragaria vesca) volatile composition. Plant Physiol. Biochem. 2017, 121, 99–117. [Google Scholar] [CrossRef]
- D’Urso, G.; Pizza, C.; Piacente, S.; Montoro, P. Combination of LC–MS based metabolomics and antioxidant activity for evaluation of bioactive compounds in Fragaria vesca leaves from Italy. J. Pharmaceut. Biomed. Anal. 2018, 150, 233–240. [Google Scholar] [CrossRef]
- Chamorro, M.F.; Reiner, G.; Theoduloz, C.; Ladio, A.; Schmeda-Hirschmann, G.; Gómez-Alonso, S.; Jiménez-Aspee, F. Polyphenol composition and (bio)activity of berberis species and wild strawberry from the Argentinean Patagonia. Molecules 2019, 24, 3331. [Google Scholar] [CrossRef]
- Nowicka, A.; Kucharska, A.Z.; Sokół-Łętowska, A.; Fecka, I. Comparison of polyphenol content and antioxidant capacity of strawberry fruit from 90 cultivars of Fragaria × ananassa Duch. Food Chem. 2019, 270, 32–46. [Google Scholar] [CrossRef]
- Vandendriessche, T.; Vermeir, S.; Mayayo Martinez, C.; Hendrickx, Y.; Lammertyn, J.; Nicolaï, B.M.; Hertog, M.L.A.T.M. Effect of ripening and inter-cultivar differences on strawberry quality. LWT Food Sci. Technol. 2013, 52, 62–70. [Google Scholar] [CrossRef]
- Mazur, S.P.; Nes, A.; Wold, A.B.; Remberg, S.F.; Martinsen, B.K.; Aaby, K. Effects of ripeness and cultivar on chemical composition of strawberry (Fragaria x ananassa Duch.) fruits and their suitability for jam production as a stable product at different storage temperatures. Food Chem. 2014, 146, 412–422. [Google Scholar] [CrossRef]
- Akšic, M.F.; Zagorac, D.D.; Sredojevic, M.; Milivojevic, J.; Gašic, U.; Meland, M.; Nati, M. Chemometric characterization of strawberries and blueberries according to their phenolic profile: Combined effect of cultivar and cultivation system. Molecules 2019, 24, 4310. [Google Scholar] [CrossRef] [PubMed]
- Moshiur Rahman, M.; Rahman, M.M.; Hossain, M.M.; Khaliq, Q.A.; Moniruzzaman, M. Effect of planting time and genotypes growth, yield and quality of strawberry (Fragaria x ananassa Duch.). Sci. Horticult. 2014, 167, 56–62. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, L.; Li, Y.; Chen, Q.; Ye, Y.; Zhang, Y.; Luo, Y.; Sun, B.; Wang, X.; Tang, H. Effect of red and blue light on anthocyanin accumulation and differential gene expression in strawberry (Fragaria x ananassa). Molecules 2018, 23, 820. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.F.S.; Liu, X.; Suarez, D.L. Fruit yield and survival of five commercial strawberry cultivars under field cultivation and salinity stress. Sci. Horticult. 2019, 243, 401–410. [Google Scholar] [CrossRef]
- Mozafari, A.A.; Ghaderi, N.; Havas, F.; Dedejani, S. Comparative investigation of structural relationships among morphophysiological and biochemical properties of strawberry (Fragaria × ananassa Duch.) under drought and salinity stresses: A study based on in vitro culture. Sci. Horticult. 2019, 256, 108601. [Google Scholar] [CrossRef]
- Voća, S.; Žlabur, J.S.; Dobričević, N.; Jakobek, L.; Šeruga, M.; Galić, A.; Pliestić, S. Variation in the bioactive compound content at three ripening stages of strawberry fruit. Molecules 2014, 19, 10370–10385. [Google Scholar] [CrossRef]
- Salvatierra, A.; Pimentel, P.; Moya-León, M.A.; Herrera, R. Increased accumulation of anthocyanins in Fragaria chiloensis fruits by transient suppression of FcMYB1 gene. Phytochemistry 2013, 290, 25–36. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Sousa, M.J.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Enhancement of nutritional and bioactive compounds by in vitro culture of wild Fragaria vesca L. vegetative parts. Food Chem. 2017, 235, 212–219. [Google Scholar] [CrossRef]
- Delgado, L.D.; Zúñiga, P.E.; Figueroa, N.E.; Pastene, E.; Escobar-Sepúlveda, H.F.; Figueroa, P.M.; Garrido-Bigotes, A.; Figueroa, C.R. Application of a JA-Ile biosynthesis inhibitor to methyl jasmonate-treated strawberry fruit induces upregulation of specific MBW complex-related genes and accumulation of proanthocyanidins. Molecules 2018, 23, 1433. [Google Scholar] [CrossRef]
- Pawlaczyk-Graja, I.; Balicki, S.; Wilk, K.A. Effect of various extraction methods on the structure of polyphenolic-polysaccharide conjugates from Fragaria vesca L. leaf. Int. J. Biol. Macromol. 2019, 130, 664–674. [Google Scholar] [CrossRef]
- Méndez-Lagunas, L.; Rodríguez-Ramírez, J.; Cruz-Gracida, M.; Sandoval-Torres, S.; Barriada-Bernal, G. Convective drying kinetics of strawberry (Fragaria ananassa): Effects on antioxidant activity, anthocyanins and total phenolic content. Food Chem. 2017, 230, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Fernández, M.A.; Hornedo-Ortega, R.; Cerezo, A.B.; Troncoso, A.M.; García-Parrilla, M.C. Effects of the strawberry (Fragaria ananassa) purée elaboration process on non-anthocyanin phenolic composition and antioxidant activity. Food Chem. 2014, 164, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Pieszka, M.; Tombarkiewicz, B.; Roman, A.; Migdał, W.; Niedziółka, J. Effect of bioactive substances found in rapeseed, raspberry and strawberry seed oils on blood lipid profile and selected parameters of oxidative status in rats. Environ. Toxicol. Pharmacol. 2013, 36, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Sikora, E.; Michorczyk, P.; Olszanska, M.; Ogonowsk, J. Supercritical CO2 extract from strawberry seeds as a valuable component of mild cleansing compositions. Int. J. Cosmet. Sci. 2015, 37, 574–578. [Google Scholar] [CrossRef]
- Grzelak-Błaszczyk, K.; Karlińska, E.; Grzęda, K.; Rój, E.; Kołodziejczyk, K. Defatted strawberry seeds as a source of phenolics, dietary fiber and minerals. LWT Food Sci. Technol. 2017, 84, 18–22. [Google Scholar] [CrossRef]
- Wichtl, M. Herbal drugs and phytopharmaceuticals. In A Handbook of Practice on a Scientific Basis; Brinckmann, J.A., Lindenmaier, M.P., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 220–221. [Google Scholar]
- Saric-Kundalic, B.; Dobes, C.; Klatte-Asselmeyer, V.; Saukel, J. Ethnobotanical study on medicinal use of wild and cultivated plants in middle, south and west Bosnia and Herzegovina. J. Ethnopharmacol. 2010, 131, 33–55. [Google Scholar] [CrossRef]
- Zhu, F. Anthocyanins in cereals: Composition and health effects. Food Res. Int. 2018, 109, 232–249. [Google Scholar] [CrossRef]
- Sinopoli, A.; Calogero, G.; Bartolotta, A. Computational aspects of anthocyanidins and anthocyanins: A review. Food Chem. 2019, 297, 124898. [Google Scholar] [CrossRef]
- Braga, A.R.C.; Murador, D.C.; de Souza Mesquita, L.M.; Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Ortan, A.; Fierascu, I.C.; Fierascu, I. In vitro and in vivo evaluation of antioxidant properties of wild-growing plants. A short review. Curr. Opin. Food Sci. 2018, 24, 1–8. [Google Scholar] [CrossRef]
- Heinrich, M.; Appendino, G.; Efferth, T.; Fürst, R.; Izzo, A.A.; Kayser, O.; Pezzuto, J.M.; Viljoen, A. Best practice in research–Overcoming common challenges in phytopharmacological research. J. Ethnopharmacol. 2020, 246, 112230. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Fernández, M.A.; Hornedo-Ortega, R.; Cerezo, A.B.; Troncoso, A.M.; García-Parrilla, M.C. Non-anthocyanin phenolic compounds and antioxidant activity of beverages obtained by gluconic fermentation of strawberry. Innov. Food Emerg. Technol. 2014, 26, 469–481. [Google Scholar] [CrossRef]
- Kebede, M.; Admassu, S. Application of antioxidants in food processing industry: Options to improve the extraction yields and market value of natural products. Adv. Food. Technol. Nutr. Sci. Open J. 2019, 5, 38–49. [Google Scholar] [CrossRef]
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Pateiro, M.; Domínguez, R.; Barba, F.J.; Putnik, P.; Bursać Kovačević, D.; Shpigelman, A.; Granato, D.; Franco, D. Berries extracts as natural antioxidants in meat products: A review. Food Res. Int. 2018, 106, 1095–1104. [Google Scholar] [CrossRef]
- Zugic, A.; Ðordevic, S.; Arsic, I.; Markovic, G.; Zivkovic, J.; Jovanovic, S.; Tadi, V. Antioxidant activity and phenolic compounds in 10 selected herbs from Vrujci Spa, Serbia. Ind. Crop Prod. 2014, 52, 519–527. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Phenolic profile and antioxidant properties of commercial and wild Fragaria vesca L. roots: A comparison between hydromethanolic and aqueous extracts. Ind. Crop Prod. 2015, 63, 125–132. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Fernandes, I.P.; Ruphuy, G.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Barreiro, M.F.; Ferreira, I.C.F.R. A bioactive formulation based on Fragaria vesca L. vegetative parts: Chemical characterization and application in κ-carrageenan gelatin. J. Funct. Food. 2015, 16, 243–255. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and human health: Effects beyond antioxidant activity. J. Agricult. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef]
- Vendrame, S.; Klimis-Zacas, D.J. Anti-inflammatory effect of anthocyanins via modulation of nuclear factor-κB and mitogen-activated protein kinase signaling cascades. Nutr. Rev. 2015, 73, 348–358. [Google Scholar] [CrossRef]
- Li, S.; Wu, B.; Fu, W.; Reddivari, L. The anti-inflammatory effects of dietary anthocyanins against ulcerative colitis. Int. J. Mol. Sci. 2019, 20, 2588. [Google Scholar] [CrossRef] [PubMed]
- Szymanowska, U.; Złotek, U.; Karaś, M.; Baraniak, B. Anti-inflammatory and antioxidative activity of anthocyanins from purple basil leaves induced by selected abiotic elicitors. Food Chem. 2015, 172, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.; Li, X.; et al. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Liberal, J.; Francisco, V.; Costa, G.; Figueirinha, A.; Amaral, M.T.; Marques, C.; Girão, H.; Lopes, M.C.; Cruz, M.T.; Batista, M.T. Bioactivity of Fragaria vesca leaves through inflammation, proteasome and autophagy modulation. J. Ethnopharmacol. 2014, 158, 113–122. [Google Scholar] [CrossRef]
- Gasparrini, M.; Giampieri, F.; Forbes-Hernandez, T.Y.; Afrin, S.; Cianciosi, D.; Reboredo-Rodriguez, P.; Varela-Lopez, A.; Zhang, J.; Quiles, J.L.; Mezzetti, B.; et al. Strawberry extracts efficiently counteract inflammatory stress induced by the endotoxin lipopolysaccharide in Human Dermal Fibroblast. Food Chem. Toxicol. 2018, 114, 128–140. [Google Scholar] [CrossRef]
- Molinett, S.; Nuñez, F.; Moya-León, M.A.; Zúñiga-Hernández, J. Chilean strawberry consumption protects against LPS-induced liver injury by anti-inflammatory and antioxidant capability in Sprague-Dawley rats. Evid.-Based Compl. Alt. Med. 2015, 2015, 320136. [Google Scholar] [CrossRef]
- Duarte, L.J.; Chaves, V.C.; dos Santos Nascimento, M.V.P.; Calvete, E.; Li, M.; Ciraolo, E.; Ghigo, A.; Hirsch, E.; Simões, C.M.O.; Reginatto, F.H.; et al. Molecular mechanism of action of Pelargonidin-3-O-glucoside, the main anthocyanin responsible for the anti-inflammatory effect of strawberry fruits. Food Chem. 2018, 247, 56–65. [Google Scholar] [CrossRef]
- Van de Velde, F.; Esposito, D.; Grace, M.H.; Pirovani, M.E.; Lila, M.A. Anti-inflammatory and wound healing properties of polyphenolic extracts from strawberry and blackberry fruits. Food Res. Int. 2019, 121, 453–462. [Google Scholar] [CrossRef]
- Gomes, F.; Martins, N.; Barros, L.; Rodrigues, M.E.; Oliveira, M.B.P.P.; Henriques, M.; Ferreira, I.C.F.R. Plant phenolic extracts as an effective strategy to control Staphylococcus aureus, the dairy industry pathogen. Ind. Crop. Prod. 2018, 112, 515–520. [Google Scholar] [CrossRef]
- Cardoso, O.; Donato, M.M.; Luxo, C.; Almeida, N.; Liberal, J.; Figueirinha, A.; Batista, M.T. Anti-Helicobacter pylori potential of Agrimonia eupatoria L. and Fragaria vesca. J. Funct. Food. 2018, 44, 299–303. [Google Scholar] [CrossRef]
- Ninomiya, M.; Itoh, T.; Ishikawa, S.; Saiki, M.; Narumiya, K.; Yasuda, M.; Koshikawa, K.; Nozawa, Y.; Koketsu, M. Phenolic constituents isolated from Fragaria ananassa Duch. Inhibit antigen-stimulated degranulation through direct inhibition of spleen tyrosine kinase activation. Bioorg. Med. Chem. 2010, 18, 5932–5937. [Google Scholar] [CrossRef] [PubMed]
- Abdulazeez, S.S. Effects of freeze-dried Fragaria x ananassa powder on alloxan-induced diabetic complications in Wistar rats. J. Taibah Univ. Med. Sci. 2014, 9, 268–273. [Google Scholar] [CrossRef]
- Somasagara, R.R.; Hegde, M.; Chiruvella, K.K.; Musini, A.; Choudhary, B.; Raghavan, S.C. Extracts of strawberry fruits induce intrinsic pathway of apoptosis in breast cancer cells and inhibits tumor progression in mice. PLoS ONE 2012, 7, 47021. [Google Scholar] [CrossRef] [PubMed]
- Forni, C.; Braglia, R.; Mulinacci, N.; Urbani, A.; Ronci, M.; Gismondi, A.; Tabolacci, C.; Provenzano, B.; Lentini, A.; Beninati, S. Antineoplastic activity of strawberry (Fragaria x ananassa Duch.) crude extracts on B16-F10 melanoma cells. Mol. Biosyst. 2014, 10, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Lucioli, S.; Pastorino, F.; Nota, P.; Ballan, G.; Frattarelli, A.; Fabbri, A.; Forni, C.; Caboni, E. Extracts from cell suspension cultures of strawberry (Fragaria x ananassa Duch): Cytotoxic effects on human cancer cells. Molecules 2019, 24, 1738. [Google Scholar] [CrossRef] [PubMed]
- Casto, B.C.; Knobloch, T.J.; Galioto, R.L.; Yu, Z.; Accurso, B.T.; Warner, B.M. Chemoprevention of oral cancer by lyophilized strawberries. Anticancer Res. 2013, 33, 4757–4766. [Google Scholar]
- Chen, T.; Shi, N.; Afzali, A. Chemopreventive effects of strawberry and black raspberry on colorectal cancer in inflammatory bowel disease. Nutrients 2019, 11, 1261. [Google Scholar] [CrossRef]
- Kanodia, L.; Borgohain, M.; Das, S. Effect of fruit extract of Fragaria vesca L. on experimentally induced inflammatory bowel disease in albino rats. Indian. J. Pharmacol. 2011, 43, 18–21. [Google Scholar] [CrossRef]
- Shi, N.; Clinton, S.K.; Liu, Z.; Wang, Y.; Riedl, K.M.; Schwartz, S.J.; Zhang, X.; Pan, Z.; Chen, T. Strawberry phytochemicals inhibit azoxymethane/dextran sodium sulfate-induced colorectal carcinogenesis in Crj: CD-1 mice. Nutrients 2015, 7, 1696–1715. [Google Scholar] [CrossRef]
- Gasperotti, M.; Masuero, D.; Mattivi, F.; Vrhovsek, U. Overall dietary polyphenol intake in a bowl of strawberries: The influence of Fragaria spp. in nutritional studies. J. Funct. Foods 2015, 18, 1057–1070. [Google Scholar] [CrossRef]
- Valenzuela, C.; Tapia, C.; López, L.; Bunger, A.; Escalona, V.; Abugoch, L. Effect of edible quinoa protein-chitosan based films on refrigerated strawberry (Fragaria × ananassa) quality. Electron. J. Biotechnol. 2015, 18, 406–411. [Google Scholar] [CrossRef]
- Shin, Y.J.; Song, H.Y.; Song, K.B. Effect of a combined treatment of rice bran protein film packaging with aqueous chlorine dioxide washing and ultraviolet-C irradiation on the postharvest quality of ‘Goha’ strawberries. J. Food. Eng. 2012, 113, 374–379. [Google Scholar] [CrossRef]
- Weisany, W.; Amini, J.; Samadi, S.; Hossaini, S.; Yousefi, S.; Struik, P.C. Nano silver-encapsulation of Thymus daenensis and Anethum graveolens essential oils enhances antifungal potential against strawberry anthracnose. Ind. Crop. Prod. 2019, 141, 111808. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Fierascu, I.; Avramescu, S.M.; Sieniawska, E. Recovery of natural antioxidants from agro-industrial side streams through advanced extraction techniques. Molecules 2019, 24, 4212. [Google Scholar] [CrossRef]
- Demirbas, A.; Yilmaz, V.; Ildiz, N.; Baldemir, A.; Ocsoy, I. Anthocyanins-rich berry extracts directed formation of Ag NPs with the investigation of their antioxidant and antimicrobial activities. J. Molec. Liq. 2017, 248, 1044–1049. [Google Scholar] [CrossRef]
- Fierascu, I.; Georgiev, M.I.; Ortan, A.; Fierascu, R.C.; Avramescu, S.M.; Ionescu, D.; Sutan, A.; Brinzan, A.; Ditu, L.M. Phyto-mediated metallic nanoarchitectures via Melissa officinalis L.: Synthesis, characterization and biological properties. Sci. Rep. 2017, 7, 12428. [Google Scholar] [CrossRef]
- Sutan, N.A.; Manolescu, D.S.; Fierascu, I.; Neblea, A.M.; Sutan, C.; Ducu, C.; Soare, L.C.; Negrea, D.; Avramescu, S.M.; Fierascu, R.C. Phytosynthesis of gold and silver nanoparticles enhance in vitro antioxidant and mitostimulatory activity of Aconitum toxicum Reichenb. rhizomes alcoholic extracts. Mat. Sci. Eng. C 2018, 93, 746–758. [Google Scholar] [CrossRef]
- Fierascu, I.; Fierascu, I.C.; Dinu-Pirvu, C.E.; Fierascu, R.C.; Anuta, V.; Velescu, B.S.; Jinga, M.; Jinga, V. A short overview of recent developments on antimicrobial coatings based on phytosynthesized metal nanoparticles. Coatings 2019, 9, 787. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Ortan, A.; Avramescu, S.M.; Fierascu, I. Phyto-nanocatalysts: Green synthesis, characterization, and applications. Molecules 2019, 24, 3418. [Google Scholar] [CrossRef]
Sample Availability: Not available.. |


{kind=link}
{kind=link}
| Class | Compound | Ref. |
|---|---|---|
| Anthocyanins | Pelargonidin 3-glucoside, cyanidin 3-glucoside, cyanidin 3-rutinoside, pelargonidin 3-galactoside, pelargonidin 3-rutinoside, pelargonidin 3-arabinoside, pelargonidin 3-malylglucoside | [11,12,13] |
| Flavonols | Quercetin, kaempferol, fisetin, their glucuronides, and glycosides | [11,12,13,16] |
| Flavanols | Catechin, proanthocyanidin B1, proanthocyanidin trimer, proanthocyanidin B3 | [11] |
| Ellagitannins | Sanguiin H-6, ellagitannin, ellagic acid, lambertianin C, galloylbis-hexahydroxydiphenoyl-glucose | [11] |
| Phenolic acids | 4-coumaric acid, p-hydroxybenzoic acid, ferulic acid, vanillic acid, sinapic acid | [15] |
| Vitamins | Vitamin C, vitamin B9 | [14] |
| Minerals | Mn, K, Mg, P, Ca | [11] |
| Others | Sugars (glucose, fructose, and sucrose), fibers | [11] |
| Species | Plant Part, Other Variables | Identified Compounds and Main Findings | Identification Method | Ref. |
|---|---|---|---|---|
| F. chiloensis | Ripe fruits | Anthocyanins (cyanidin 3-O-glucoside, pelargonidin 3-O-glucoside cyanidin-malonyl-glucoside and pelargonidin-malonyl-glucoside); procyanidins, ellagitannins, ellagic acid and flavonol derivatives | HPLC-DAD, LC-ESI-MS | [26] |
| F. chiloensis | Leaves | Procyanidins, ellagitannins, ellagic acid and flavonol derivatives | HPLC-DAD, LC-ESI-MS | [26] |
| F. chiloensis | Rhizomes | Procyanidins, ellagitannins, ellagic acid and flavonol derivatives | HPLC-DAD, LC-ESI-MS | [26] |
| Fragaria × ananassa | Fruits | Anthocyanins (pelargonidin-3-glucoside, pelargonidin-3-rutinoside, cyanidin-3-rutinoside, pelargonidin-3,5-diglucoside, pelargonidin-3-(6-acetyl)-glucoside, 5-carboxypyranopelargonidin-3-glucoside, delphinidin-3-glucoside, peonidin-3-glucoside, cyanidin-3-galactoside), p-hydroxybenzoic acid, (+)-catechin, ellagic acid, p-coumaric acid, quercetin glucoside | LC-MS/MS, HPLC-UV/Vis | [17] |
| Fragaria × ananassa | Fruits, cultivar and seasonal variations | Vitamin C, β-carotene, total phenolics, total anthocyanins; genotype influence is stronger than the environmental influence | Colorimetric | [27] |
| Fragaria × ananassa | Fruits, different cultivars on different ripeness stage | Total vitamin C, total phenolics, total anthocyanins, total ellagic acid/pelargonidin-3-glucoside and cyanidin-3-glucoside; higher amounts in pink fruits compared with fully ripped fruits | Colorimetric/HPLC-DAD | [28] |
| Fragaria × ananassa | Fruits, different farming methods | Total phenolics/pelargonidin-3-glucoside and cyanidin-3-glucoside, vitamin C, higher in organic farming fruits | Colorimetric/HPLC-DAD | [18] |
| Fragaria × ananassa | Fruits, different cultivars (27) and ripening stages | Phenolic compounds (multiple classes, including anthocyanins, flavanols and ellagitannins); composition dependent on cultivar, cinnamic acid conjugates and anthocyanins levels increased with the ripening stage | HPLC-DAD-MS | [29] |
| Fragaria × ananassa, F. vesca | Fruits | Quercetin and isorhamnetin glycosides (higher levels in wild strawberry) | HPLC-DAD, LC-ESI-MS | [30] |
| Fragaria × ananassa, F. vesca | Fruits, different cultivars | Volatile esters (including ethyl acetate, hexyl acetate, methyl butanoate, ethyl butanoate, hexyl butanoate, methyl hexanoate, ethyl hexanoate, hexyl hexanoate); higher levels in cultivated strawberries. | GC-MS | [19] |
| F. vesca | Fruits, two different cultivars | Anthocyanins (cyanidin 3-O-glucoside, pelargonidin 3-O-glucoside, peonidin 3-O-glucoside, cyanidin 3-O-malonylglucoside, pelargonidin 3-O-malonylglucoside, peonidin 3-O-malonylglucoside), dihydroflavonol and flavonols (taxifolin 3-O-arabinoside, kaempferol 3-O-glucoside, quercetin 3-O-glucoside, quercetin-acetylhexoside, kaempferol 3-O-acetylhexosides), flavan-3-ols and proanthocyanidins (catechin, B type proanthocyanidin dimers, trimers, and tetramers), ellagic acid and derivatives (glycosylated, methyl pentoside, methylellagic acid methyl pentoside, ellagitannins), other compounds (benzoic acid, ferulic acid hexose derivative, citric acid, furaneol glucoside) | HPLC-DAD | [31] |
| Fragaria × ananassa, F. vesca | Fruits | Anthocyanins (cyanidin, pelargonidin), cyanidin glycosides (cyanidin 3-glucoside, cyanidin 3-arabinoside, cyanidin 3-sambubioside, delphinidin 3-galactoside, delphinidin 3-glucoside, delphinidin 3-malonylglucoside); higher levels of cyanidin glycosides in wild species | HPLC-DAD | [32] |
| F. vesca | Leaves | Ellagitannins (sanguiin H-2 isomer, sanguiin H-10 isomer, sanguiin H-6/agrimoniin/lambertianin A isomer, castalagin/vescalagin isomer, sanguiin H-10 isomer, sanguiin H-2 isomer, casuarictin/potentillin isomer) | LC-PDA-ESI-MS | [33] |
| Fragaria × ananassa | Fruits, different cultivars and production years | Vitamin C, anthocyanins (pelargonidin 3-glucoside, cyanidin 3-glucoside, pelargonidin 3-rutinoside), ellagic acid; strongly dependent on the cultivar and production year | HPLC-UV/Vis | [34] |
| Fragaria × ananassa | Fruits, at different ripening stage | Vitamin C, pelargonidin-3-rutinoside, ellagic acid, cyanidin-3-glucoside, quercetin (red fruits), neochlorogenic, pelargonidin-3-glucoside, pelargonidin-3-rutinoside, epicatechin, quercetin-3-β-d-glucoside, ellagic acid (green fruits) | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Calyx (red and green) | Quercetin-3-β-d-glucoside, ellagic acid, kaempferol-3-O-glucoside, vitamin C (red), catechin, quercetin-3-β-d-glucoside, ellagic acid (green) | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Flower | Catechin, quercetin-3-β-d-glucoside, ellagic acid, kaempferol-3-O-glucoside, vitamin C | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Leaf | Procyanidin dimer and trimer, catechin, quercetin-3-β-d-glucoside, vitamin C, ellagic acid | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Stolon | Neochlorogenic, procyanidin dimer, catechin, quercetin-3-β-d-glucoside, ellagic acid, vitamin C, kaempferol-3-O-glucoside | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Stem | Procyanidin dimer, catechin, ferulic acid, quercetin-3-β-d-glucoside, ellagic acid | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Crown | Procyanidin dimer and trimer, catechin, propelargonidin dimer, ellagic acid | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Root | Procyanidin dimer and trimer, catechin, neochlorogenic, propelargonidin dimer | LC-ESI-TOF | [35] |
| Fragaria × ananassa | Fruits, different novel cultivars | Phenolic acids (p-coumaric acid, ellagic acid, ferulic acid derivative, p-coumaric acid derivatives), monomeric flavanols ((+)-catechin), flavonols (quercetin 3-O-glucoside, fisetin, quercetin 3-O-glucoside derivative), anthocyanins (cyanidin 3-glucoside, cyanidin 3-rutinoside, cyanidin pentoside, pelargonidin 3-galactoside, pelargonidin 3,5-diglucoside, pelargonidin 3-glucoside, pelargonidin 3-rutinoside, cyanidin 3-Oacetylglucoside, cyanidin hexoside, pelargonidin 3-O-monoglucuronide, pelargonidin derivatives) | HPLC-DAD, LC-ESI-QTOF | [36] |
| Fragaria × ananassa | Fruits, grown on different altitudes, on consecutive years | Hydroxybenzoic acid, p-coumaric acid, other hydroxycinnamic acids, (+)-catechin, (−)-epicatechin, procyanidins, flavonols, anthocyanins (cyanidin 3-glucoside, pelargonidin 3-glucoside, pelargonidin derivative); higher levels recorded at lower altitudes. | HPLC-DAD | [37] |
| Fragaria × ananassa | Fruits | Kaempferol 3-(6-methylglucuronide), quercetin 3-(6-methylglucuronide), isorhamnetin 3-(6-methylglucuronide), trichocarpin, 2-p-hydroxybenzoyl-2,4,6-tri hydroxyphenylacetate, 2-p-hydroxyphene thyl-6-caffeoylglucoside, zingerone 4-glucoside, b-hydroxypropiovanillone 3-glucoside, (+)-isolariciresinol 90-glucoside, (−)-isolariciresinol 90-glucoside, aviculin, (−)-secoisolariciresinol 4-glucoside, cupressoside A, cedrusin, icariside E4, dihydrodehydrodiconiferyl alcohol 90-glucoside, massonianoside A, urolignoside, (−)-pinoresinol 4-glucoside, 2,3”-epoxy-4-(butan-2-one-3-yl)-5,7,40-trihydroxy flavane 3-glucoside, kaempferol 3-(6-butylglucuronide), benzyl 2-glucosyl-6-rhamnosylbenzoate | 1H NMR, 13C NMR, HMBC, HPLC-UV/Vis, LC-MS/MS, HR-ESI-MS, | [20] |
| F. vesca | Fruits, wild and cultivated, from different geographical areas | 39 phenolic compounds (including cyanidin 3-O-glucoside, delphinidin-3-O-glucoside, pelargonidin-3-O-glucoside, pelargonidin-3-O-rutinoside, (+) catechin, (−) epicatechin, procyanidin B1 and B2, isoquercetin, gallic acid, p-coumaric acid, phloridzin); composition dependent on the geographical area | LC-ESI-Orbitrap-MS, LC-ESI-QTrap-MS, LC-ESI-QTrap-MS/MS | [38] |
| Fragaria × ananassa | Fruits, different cultivars | Cyanidin 3-O-glucoside, pelargonidin-3-O-glucoside, pelargonidin-O-rutinoside, total anthocyanins content, dependent on the cultivar | UPLC-PDA-ESI-MS, HPLC-DAD | [39] |
| F. vesca | Fruits | Volatile composition—one hundred compounds (including esters, aldehydes, ketones, alcohols, terpenoids, furans and lactones). | GS-MS | [40] |
| F. vesca | Leaves | 27 metabolites (organic acids, flavonoids, catechin and its oligomers, ellagitannins), including quinic acid, chelidonic acid, quercetin derivatives, catechin and procyanidins, phloridzin, pedunculagin, methyl ellagic acid glucuronide. | LC-ESI-Orbitrap-MS | [41] |
| Fragaria × ananassa, F. vesca | White-fruited mutants, different genotypes | Anthocyanins, flavonols, flavan-3-ols, hydroxycinnamic acids, and ellagic acid—derived compounds, dependent on genotype | LC-ESI-MS/MS | [21] |
| F. chiloensis | Fruits | Anthocyanins (cyanidin-3-O-glucoside, pelargonidin hexoside, cyanidin manlonyl hexoside, pelargonidin-malonyl hexoside), ellagitannins (ellagic acid hexoside, pentoside, rhamnoside), proanthocyanidin dimers, epicatechin, flavonols (quercetin pentoside, glucuronide) | HPLC-DAD, LC-ESI-MS | [42] |
| Fragaria × ananassa | Fruits, different cultivars | Anthocyanins, flavonoids, cinnamic acid derivatives, tannins and related compounds, triterpenoids; concentration dependent on the cultivar | UPLC-ESI-QTOF-MS/MS, HPLC-DAD | [43] |
| Species | Extraction Method | Antioxidant Assay | Antioxidant Potential | Responsible Compounds | Ref. |
|---|---|---|---|---|---|
| Fragaria × ananassa, Camarosa var. fruits | Anthocyanins isolated using CCC | ORAC, FRAP | ORAC: 2.7–24.46 mmol Trolox/g; FRAP: 2.75–12.5 mmol Fe2+/g (depending on the fraction) | Anthocyanins | [17] |
| Fragaria chiloensis spp. chiloensis form chiloensis fruits | Methanol: formic acid (99:1 v/v) extraction | DPPH, SAS | DPPH assay: IC50 = 38.7 mg/L; SAS: 79.3%) | Aglycone and glycosylated ellagic acid and flavonoids | [26] |
| Fragaria chiloensis spp. chiloensis form chiloensis leaves | Methanol: formic acid (99:1 v/v) extraction | DPPH, SAS | DPPH assay: IC50 = 49.4 mg/L; SAS: 67.60% | Aglycone and glycosylated ellagic acid and flavonoids | [26] |
| Fragaria chiloensis spp. chiloensis form chiloensis rhizomes | Methanol: formic acid (99:1 v/v) extraction | DPPH, SAS | DPPH assay: IC50 = 64.8 mg/L; SAS: 55% | Aglycone and glycosylated ellagic acid and flavonoids | [26] |
| Fragaria x ananassa Osogrande var. frozen fruits | Acetone (80%) extraction | DPPH, FRAP | DPPH: 11.91–12.83 μmol BHT eq./g FW; best results for ripe fruits FRAP: 27.37–36.75 μmol FS eq./g FW; best results for green fruits | Total phenolic content, vitamin C | [28] |
| Fragaria x ananassa Camino Real var. frozen fruits | Acetone (80%) extraction | DPPH, FRAP | DPPH: 9.75–12.01 μmol BHT eq./g FW, FRAP: 24.13–28.49 μmol FS eq./g FW (best results for pink fruits) | Total phenolic content, vitamin C | [28] |
| F. vesca leaves | Methanol, ultrasounds extraction | DPPH, FRAP | DPPH: IC50 = 13.46 mg/L; FRAP: 0.878 mmol Fe2+/g DW | Total phenols, total tannins | [72] |
| F. vesca roots, wild-growing | Hydromethanolic extraction, infusion, decoction | DPPH, FRAP, β-Carotene bleaching inhibition, TBARS | IC50, mg/L: DPPH—50.03/50.56/50.62; FRAP—40.98/44.78/49.23; β-C bleaching—116.26/44.88/66.10; TBARS—35.76/4.76/6.14 | Total phenolics, total flavan-3-ols, total dihydroflavonols, | [73] |
| F. vesca roots, commercial | Hydromethanolic extraction, infusion, decoction | DPPH, FRAP, β-Carotene bleaching inhibition, TBARS | IC50, mg/L: DPPH—68.89/255.81/51.32; FRAP—327.75/78.99/67.92; β-C bleaching—68.34/23.44/114.67; TBARS—6.69/24.25/10.62 | Total phenolics, total flavan-3-ols, total dihydroflavonols, | [73] |
| Fragaria × ananassa var. Amaou, fruits, at different ripening stage | Ethanol or water room temperature extraction | Modified ABTS assay | Ethanol: 150.5/151.9; water: 227.2/189.4 (red/green fruits) μmol TE/100 g FW | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou calyx (red and green) | Ethanol or water room temperature extraction | Modified ABTS assay | Ethanol: 241.1/1239.9; water: 1716.6/577.7 μmol TE/100 g FW (red/green calyx) | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou flower | Ethanol or water room temperature extraction | Modified ABTS assay | 4234.4/387.5 μmol TE/100 g FW (ethanol/water) | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou leaves | Ethanol or water room temperature extraction | Modified ABTS assay | 2401.7/241.1 μmol TE/100 g FW (ethanol/water) | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou stolon | Ethanol or water room temperature extraction | Modified ABTS assay | 1089.4/1856.7 μmol TE/100 g FW (ethanol/water) | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou stem | Ethanol or water room temperature extraction | Modified ABTS assay | 1338.6/1123.1 μmol TE/100 g FW (ethanol/water) | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou crown | Ethanol or water room temperature extraction | Modified ABTS assay | 6213.3/128.7 μmol TE/100 g FW (ethanol/water) | Total phenolic content | [35] |
| Fragaria × ananassa var. Amaou root | Ethanol or water room temperature extraction | Modified ABTS assay | 253.1/69.2 μmol TE/100 g FW (ethanol/water) | Total phenolic content | [35] |
| F. vesca vegetative parts (leaves and stems), wild-growing | Hydromethanolic and aqueous extracts; wild-growing infusion microencapsulated in alginate and incorporated in k-carrageenan gelatine | DPPH, FRAP, β-Carotene bleaching inhibition, TBARS | IC50, mg/L: DPPH—123.67/86.17/109.10; FRAP—81.40/62.36/77.28; β-C bleaching—56.71/12.34/13.40; TBARS—12.63/3.12/5.03 (hydromethanolic/infusion/decoction); Final formulation (mg/mL)—DPPH—2.74; FRAP = 1.23 | Total phenolics, total flavan-3-ols, total dihydroflavonols, | [74] |
| F. vesca vegetative parts (leaves and stems), commercial | Hydromethanolic and aqueous extracts | DPPH, FRAP, β-Carotene bleaching inhibition, TBARS | IC50, mg/L: DPPH—139.33/121.94/118.89; FRAP—324.49/91.88/88.20; β-C bleaching—388.90/76.41/69.98; TBARS—24.36/23.07/17.52 (hydromethanolic/infusion/decoction). | Total phenolics, total flavan-3-ols, total dihydroflavonols, | [74] |
| Fragaria x ananassa cv. Falandi fruit | 22 compounds isolated from ethanolic extracts | ABTS, DPPH, FRAP | Best results (IC50): ABTS—4.42 μM kaempferol 3-(6-methylglucuronide); DPPH—32.12 μM quercetin 3-(6-methylglucuronide); FRAP—0.05 mmol/g—urolignoside. | Individual compounds | [20] |
| Fragaria x ananassa cv. Albion, Aromas, Camarosa, Camino Real, Monte Rey, Portola, and San Andreas fruits | Ultrasonic extraction with acidified methanol | DPPH | IC50 (mg/mL) ranging from 76.73 (Camarosa)—100 (Camino Real) | Total anthocyanin content | [39] |
| F. vesca leaves native to Italy | Ultrasonic extraction with ethanol: water solvent (70:30, v/v) | TEAC | 0.34–0.35 mg/mL Trolox eq., compared with quercetin (0.40) | Condensed tannins and flavonoid derivatives | [41] |
| Fragaria x ananassa cv. Tochiotome leaves | Supercritical CO2 extraction with different entrainers | DPPH | 0.07 (simple supercritical extraction)—5.82 μmol BHT/g sample (with ethanol, dried at 40 °C) | Phenolic compounds | [10] |
| Fragaria × ananassa fruits (90 cultivars) | Ultrasonic aqueous methanol (70%) acidified with 1.5% formic acid, at room temperature | DPPH, ABTS | Average values (μmol Trolox/100 g):765.06 (DPPH), 1637.96 (ABTS) | Tannin-based compounds. | [43] |
| Action | Plant | Extraction Method | Assay | Results | Responsible Compounds | Ref. |
|---|---|---|---|---|---|---|
| Anti-inflammatory on inflammatory bowel disease | Fragaria vesca leaves | Eth. extraction | MPO activity; GSH, SOD and CAT levels | Prevention of increase in colon weight and disease activity index, decrease in macroscopic and microscopic lesion score; significant improvement of MPO, CAT and SOD levels at 500 mg/kg 5 days oral treatment | Phenolic acids, flavonoids | [94] |
| Anti-inflammatory | Fragaria vesca leaves | Eth. extraction at room temperature, infusion | Nitric oxide production, western blot analysis (expression of pro-inflammatory proteins in lipopolysaccharide-triggered macrophages); nitric oxide scavenger activity | Inhibition of nitrite production on pre-treated cells (at 80 and 160 mg/L—31%/40%); 23% inhibition in culture media, at 160 mg/L | Phenolic content | [80] |
| Anti-inflammatory | Fragaria x ananassa, var. Alba fruits | Meth. extraction at room temperature, infusion | Determination of ROS intracellular levels, apoptosis detection, antioxidant enzyme activities, immunoblotting analysis, determination of mitochondrial respiration and extracellular acidification rate in cells | Reduction of intracellular ROS levels (significant at 100 mg/L), decreased apoptotic rate (significant at 50 and 100 mg/L); Increased ARE-antioxidant enzymes expression, reduced NO and inflammatory cytokines production (at 50 and 100 mg/L) to control levels | Vitamin C, anthocyanins, flavonoids | [81] |
| Anti-inflammatory, hepatoprotective | Fragaria chiloensisssp. Chiloensis fruits | Aq. extracts | Histological analyses, determination of transaminases, cytokines, F2-isoprostanes, and glutathione assays | maintained hepatocellular membrane, structural integrity, attenuated hepatic oxidative stress, and inhibited inflammatory response in LPS-induced liver injury; downregulation of cytokines (TNFa, IL-1β, and IL-6) | Phenolic content | [82] |
| Anti-inflammatory | Fragaria x ananassa var. Camarosa fruits | Ultrasonic-assisted, acidified meth. extraction, separation | In vivo: quantification of the leukocyte content, exudate concentration, MPO and ADA activities, nitric oxide products, TNF-α and IL-6 levels; in vitro: MTT assay, measurement of nitric oxide products, TNF-α and IL-6 levels, western blot analysis | Inhibition of the carrageenan-induced leukocyte influx to the pleural cavity; reduction of myeloperoxidase activity, exudate concentration, NO levels. | Phenolic compounds, anthocyanins (particularly pelargonidin-3-O-glucoside) | [83] |
| Anti-inflammatory, wound healing | Fragaria x ananassa var. San Andreas fruits | Ultrasound-assisted extraction, acidified meth.: aq. (80:20); separation of different fractions | MTT assay, ROS, NO levels, effects on inflammatory markers and on skin fibroblast migration | ROS reduction, suppression of IL-1β, IL-6 and iNOS gene expressions; enhanced skin fibroblast migration | Polyphenolic compounds, especially anthocyanins | [84] |
| Anti-microbial | Fragaria vesca leaves and roots | Centrifugation extraction with meth.: aq. (80:20) | Disc diffusion assay | 6–9 mm inhibition zones for leaves, 5–9 mm for roots (depending on S. aureus strain) | Phenolic compounds | [85] |
| Anti-microbial | Fragaria vesca leaves | Hydroalcoholic extraction, separation | Disc diffusion assay | Good inhibition potential at 25 mg/mL, better effect for the ellagitannin-enriched fraction | Ellagitannins | [86] |
| Anti-allergenic | Fragaria x ananassa var. Minomusume fruits | Methanol fraction of fruits juice (obtained by squeezing) | Antigen-stimulated degranulation in RBL-2H3 cells | degranulation suppression (95–60% inhibition for linocinnamarin, cinnamic acid, chrysin, kaempferol, trans-tiliroside) | Best results - phenylpropanoid glycoside | [87] |
| Anti-diabetic | Fragaria x ananassa var. Falandi fruits | Compounds isolated from eth. extracts | α-glucosidase inhibitory activity | IC50 values better than the positive control (acarbose) for nine compounds (537.43 to 25.39 μM) | Individual compounds | [20] |
| Anti-obesity, anti-allergy, skin-lightening | Fragaria ×ananassa var. Amaou, entire plant (red fruit, green fruit, red calyx, green calyx, flower, leaf, stolon, stolon leaf, stem, crown and root) | Eth. or aq. room temperature extraction | Anti-lipase assay, adipocyte differentiation inhibition assay, melanogenesis inhibition assay, β-hexosaminidase inhibition assay, tyrosinase inhibition assay | Crown, stolon leaf and flowers extracts exhibited the highest effects | Total phenolic content | [35] |
| Antihyperuricemic | Fragaria x ananassa cv. Tochiotome leaves | Supercritical CO2 extraction with different entrainers | Uric acid production in AML12 hepatocytes | Reduction of uric acid at 100 mg/mL (96 mmol/2 h/mg protein), compared with the control (16,096 mmol/2 h/mg protein) | Kaempferol, quercetin | [10] |
| Cytotoxic, anti-proliferative | Fragaria x ananassa fruits | Meth. extraction | Ex vivo: cell viability assay; in vivo: developing tumor size determination | Cytotoxic on cancer cells, blocked the proliferation of tumor cells | Phenolic compounds | [89] |
| Antineoplastic | Fragaria x ananassa var. Pajaro fruits | Acidified hydro-eth. extraction | Transglutaminase assay and polyamine detection, immunoblot analysis | reduction of cell proliferation, lowering of the intracellular levels of polyamine, enhancement of tissue transglutaminase activity | Anthocyanins | [90] |
| Cytotoxic | Fragaria vesca L. leaves | Hydroalcoholic extract at room temperature, ellagitannins-enriched fraction | Effects on HepG2 cells—cell viability assessment, cell proliferation, cell cycle and cell death analysis, Western blot analysis, proteasome chymotrypsin-like activity | Inhibition of HepG2 cell viability IC50 = 690 mg/L (extract)/113 mg/L (fraction); fraction induced necrosis and apoptosis, influenced the cellular proteolytic mechanisms | Ellagitannins | [33] |
| Chemopreventive | Lyophilized Fragaria x ananassa fruits | Ultrasound-assisted extraction with acidified acetone | Histological studies, Western blot analysis, PGE2 measurement, and nitrate/nitrite colorimetric assay | Decreased tumor incidence, decreased levels of TNF-α, IL-1β, IL-6, COX-2 and iNOS, inhibition of the phosphorylation of PI3K, Akt, ERK, and NFκB | anthocyanins, ellagitannin/ellagic acid/ellagic acid derivatives flavonols | [95] |
| Cytotoxic | Fragaria x ananassa leaves | Hydroalcoholic extracts (meth., eth., isopropanol) from in vitro cell suspension | Cell proliferation, cell viability | Under 50% viable cells for colorectal adenocarcinoma and colon adenocarcinoma upon treatment with extracts containing 0.29 mM ethoxy-dihydrofuro-furan | Polyphenols | [91] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fierascu, R.C.; Temocico, G.; Fierascu, I.; Ortan, A.; Babeanu, N.E. Fragaria Genus: Chemical Composition and Biological Activities. Molecules 2020, 25, 498. https://doi.org/10.3390/molecules25030498
Fierascu RC, Temocico G, Fierascu I, Ortan A, Babeanu NE. Fragaria Genus: Chemical Composition and Biological Activities. Molecules. 2020; 25(3):498. https://doi.org/10.3390/molecules25030498
Chicago/Turabian StyleFierascu, Radu Claudiu, Georgeta Temocico, Irina Fierascu, Alina Ortan, and Narcisa Elena Babeanu. 2020. "Fragaria Genus: Chemical Composition and Biological Activities" Molecules 25, no. 3: 498. https://doi.org/10.3390/molecules25030498
APA StyleFierascu, R. C., Temocico, G., Fierascu, I., Ortan, A., & Babeanu, N. E. (2020). Fragaria Genus: Chemical Composition and Biological Activities. Molecules, 25(3), 498. https://doi.org/10.3390/molecules25030498