
Panax quinquefolius (North American Ginseng) Polysaccharides as Immunomodulators: Current Research Status and Future Directions


Abstract
1. Introduction
2. Current Research Status of NAG Polysaccharides: An Overview
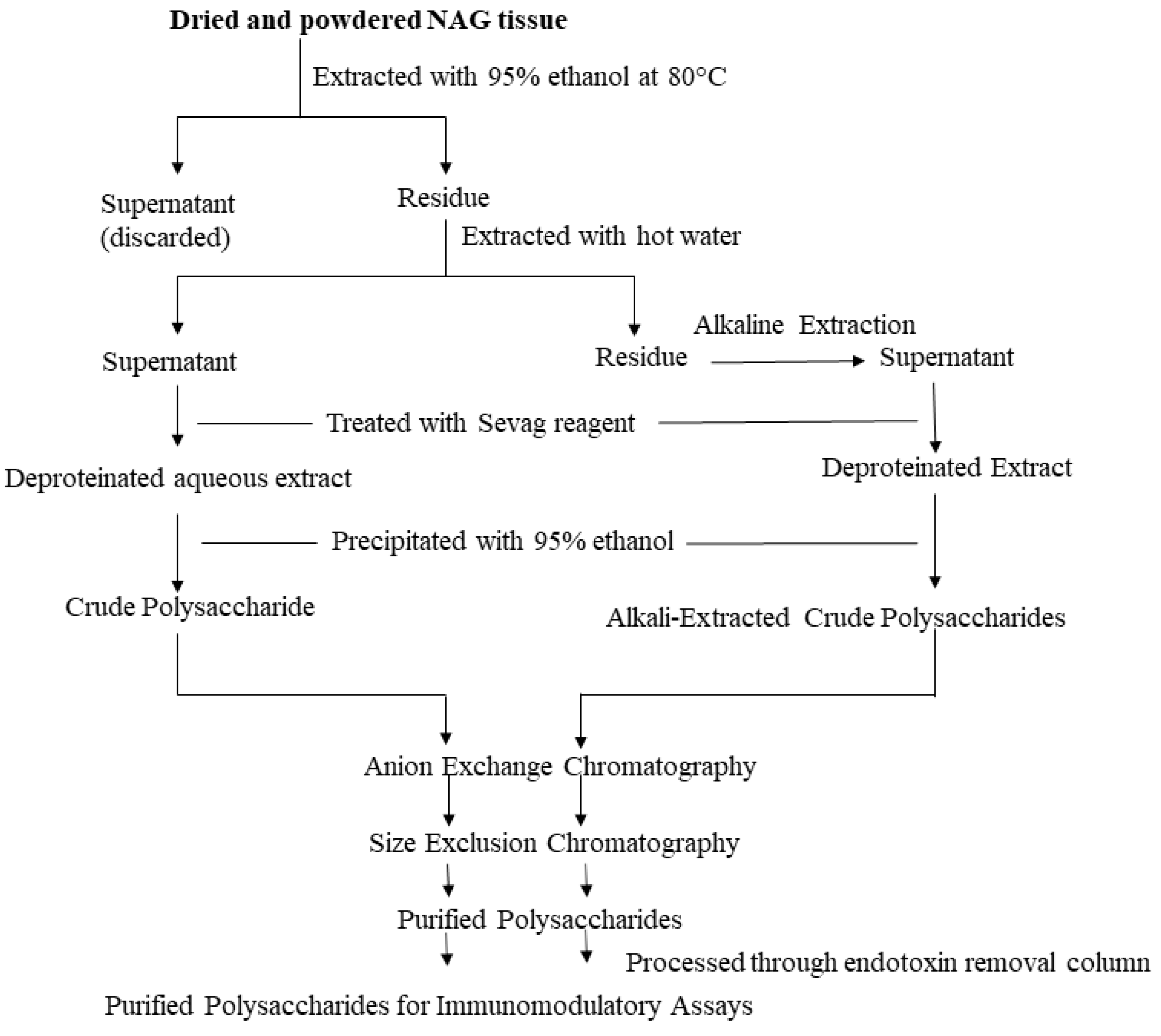
3. Extraction, Purification, and Characterization of NAG Polysaccharides
4. Immunomodulatory Properties of NAG Polysaccharide Extracts
5. Mechanisms of Immunomodulation by NAG Polysaccharides
5.1. Overview
5.2. Receptors of the Innate Immune System that Potentially Bind to NAG Polysaccharides
5.2.1. TLRs
5.2.2. C-type Lectin Receptors
5.2.3. CR3, SRs, and NOD-2
5.3. NAG Polysaccharide Classes and Their Immunomodulatory Mechanism
6. Recent Innovations and Future Directions
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Valentino, G.; Graziani, V.; D’Abrosca, B.; Pacifico, S.; Fiorentino, A.; Scognamiglio, M. NMR-based plant metabolomics in nutraceutical research: An overview. Molecules 2020, 25, 1444. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Kiyohara, H. Immunomodulating activity of plant polysaccharide structures. In Comprehensive Glycoscience; Kamerling, H., Ed.; Elsevier: Oxford, UK, 2007; pp. 663–694. [Google Scholar]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure–function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Date, A.; Chawda, H.; Patel, K. Polysaccharides as potential anticancer agents—A review of their progress. Carbohydr. Polym. 2019, 210, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, X.; He, M.; Zheng, W.; Qi, D.; Zhang, Y.; Han, C.-C. Ginseng polysaccharides: A potential neuroprotective agent. J. Ginseng Res. 2020. [Google Scholar] [CrossRef]
- Chen, F.; Huang, G. Antioxidant activity of polysaccharides from different sources of ginseng. Int. J. Biol. Macromol. 2019, 125, 906–908. [Google Scholar] [CrossRef]
- Yoo, D.-G.; Kim, M.-C.; Park, M.-K.; Park, K.-M.; Quan, F.-S.; Song, J.-M.; Wee, J.J.; Wang, B.-Z.; Cho, Y.-K.; Compans, R.W.; et al. Protective effect of ginseng polysaccharides on influenza viral infection. PLoS ONE 2012, 7, e33678. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Riaz, M.; Rahman, N.U.; Zia-Ul-Haq, M.; Jaffar, H.Z.E.; Manea, R. Ginseng: A dietary supplement as immune-modulator in various diseases. Trends Food Sci. Technol. 2019, 83, 12–30. [Google Scholar] [CrossRef]
- Licciardi, P.V.; Underwood, J.R. Plant-derived medicines: A novel class of immunological adjuvants. Int. Immunopharmacol. 2011, 11, 390–398. [Google Scholar] [CrossRef]
- Azike, C.G.; Charpentier, P.A.; Lui, E.M.K. Stimulation and suppression of innate immune function by American ginseng polysaccharides: Biological relevance and identification of bioactives. Pharm. Res. 2015, 32, 876–897. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, C.; Santangelo, R. Panax ginseng and Panax quinquefolius: From pharmacology to toxicology. Food Chem. Toxicol. 2017, 107, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Smith, S.A.; Nwangwa, E.E.; Arivett, B.A.; Bryant, D.L.; Fuller, M.L.; Hayes, D.; Bowling, J.L.; Nelson, D.E.; DuBois, J.D.; et al. Panax quinquefolius (North American ginseng) cell suspension culture as a source of bioactive polysaccharides: Immunostimulatory activity and characterization of a neutral polysaccharide AGC1. Int. J. Biol. Macromol. 2019, 139, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Choi, H.-K.; Huang, L. State of Panax ginseng research: A global analysis. Molecules 2017, 22, 1518. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ju, Z.; Yang, Y.; Zhang, Y.; Yang, L.; Wang, Z. Phytochemical analysis of Panax species: A review. J. Ginseng Res. 2020. [Google Scholar] [CrossRef]
- Kim, J.H.; Yi, Y.-S.; Kim, M.-Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef]
- Yi, Y.-S. Roles of ginsenosides in inflammasome activation. J. Ginseng Res. 2019, 43, 172–178. [Google Scholar] [CrossRef]
- Szczuka, D.; Nowak, A.; Zakłos-Szyda, M.; Kochan, E.; Szymańska, G.; Motyl, I.; Blasiak, J. American ginseng (Panax quinquefolium L.) as a source of bioactive phytochemicals with pro-health properties. Nutrients 2019, 11, 1041. [Google Scholar] [CrossRef]
- Zhao, B.; Lv, C.; Lu, J. Natural occurring polysaccharides from Panax ginseng C. A. Meyer: A review of isolation, structures, and bioactivities. Int. J. Biol. Macromol. 2019, 133, 324–336. [Google Scholar] [CrossRef]
- Ji, X.; Hou, C.; Shi, M.; Yan, Y.; Liu, Y. An Insight into the research concerning Panax ginseng C. A. Meyer polysaccharides: A review. Food Rev. Int. 2020, 1–17. [Google Scholar] [CrossRef]
- Lim, W.; Mudge, K.W.; Vermeylen, F. Effects of population, age, and cultivation methods on ginsenoside content of wild American ginseng (Panax quinquefolium). J. Agric. Food Chem. 2005, 53, 8498–8505. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, R.C.; Chamberlain, J.L.; Munsell, J.F. Growing American ginseng (Panax quinquefolius) in forestlands. Va. Coop. Ext. 2009. 354–313. Available online: https://www.pubs.ext.vt.edu/354/354-313/354-313.html (accessed on 28 September 2020).
- Qiang, B.; Miao, J.; Phillips, N.; Wei, K.; Gao, Y. Recent advances in the tissue culture of American ginseng (Panax quinquefolius). Chem. Biodivers. 2020, 17, e2000366. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://mtsu.edu/ginseng/ (accessed on 2 October 2020).
- Available online: https://www.uwo.ca/physpharm/ogirc/ (accessed on 2 October 2020).
- Pugh, N.D.; Tamta, H.; Balachandran, P.; Wu, X.; Howell, J.L.; Dayan, F.E.; Pasco, D.S. The majority of in vitro macrophage activation exhibited by extracts of some immune enhancing botanicals is due to bacterial lipoproteins and lipopolysaccharides. Int. Immunopharmacol. 2008, 8, 1023–1032. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gertsch, J.; Viveros-Paredes, J.M.; Taylor, P. Plant immunostimulants—Scientific paradigm or myth? J. Ethnopharmacol. 2011, 136, 385–391. [Google Scholar] [CrossRef]
- Assinewe, V.A.; Arnason, J.T.; Aubry, A.; Mullin, J.; Lemaire, I. Extractable polysaccharides of Panax quinquefolius L. (North American ginseng) root stimulate TNFa production by alveolar macrophages. Phytomedicine 2002, 9, 398–404. [Google Scholar] [CrossRef]
- Lemmon, H.R.; Sham, J.; Chau, L.A.; Madrenas, J. High molecular weight polysaccharides are key immunomodulators in North American ginseng extracts: Characterization of the ginseng genetic signature in primary human immune cells. J. Ethnopharmacol. 2012, 142, 1–13. [Google Scholar] [CrossRef]
- Yu, X.; Yang, X.; Cui, B.; Wang, L.; Ren, G. Antioxidant and immunoregulatory activity of alkali-extractable polysaccharides from North American ginseng. Int. J. Biol. Macromol. 2014, 65, 357–361. [Google Scholar] [CrossRef]
- Sevag, M.G.; Lackman, D.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar]
- Guo, Q.; Cui, S.W.; Kang, J.; Ding, H.; Wang, Q.; Wang, C. Non-starch polysaccharides from American ginseng: Physicochemical investigation and structural characterization. Food Hydrocoll. 2015, 44, 320–327. [Google Scholar] [CrossRef]
- Yang, X.-S.; Wang, L.-J.; Dong, C.; Lui, E.M.K.; Ren, G.-X. Optimization of ultrasonic-assisted extraction process of polysaccharides from American ginseng and evaluation of its immunostimulating activity. J. Integr. Agric. 2014, 13, 2807–2815. [Google Scholar] [CrossRef]
- Yu, X.-H.; Liu, Y.; Wu, X.-L.; Liu, L.-Z.; Fu, W.; Song, D.-D. Isolation, purification, characterization and immunostimulatory activity of polysaccharides derived from American ginseng. Carbohydr. Polym. 2017, 156, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Hosain, N.A.; Ghosh, R.; Bryant, D.L.; Arivett, B.A.; Farone, A.L.; Kline, P.C. Isolation, structure elucidation, and immunostimulatory activity of polysaccharide fractions from Boswellia carterii frankincense resin. Int. J. Biol. Macromol. 2019, 133, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Bryant, D.L.; Arivett, B.A.; Smith, S.A.; Altman, E.; Kline, P.C.; Farone, A.L. An acidic polysaccharide (AGC3) isolated from North American ginseng (Panax quinquefolius) suspension culture as a potential immunomodulatory nutraceutical. Curr. Res. Food Sci. 2020, 3, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yao, Y.; Sang, W.; Yang, X.; Ren, G. Structural features and immunostimulating effects of three acidic polysaccharides isolated from Panax quinquefolius. Int. J. Biol. Macromol. 2015, 80, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Cheong, K.-L.; Wu, D.-T.; Deng, Y.; Leong, F.; Zhao, J.; Zhang, W.-J.; Li, S.-P. Qualitation and quantification of specific polysaccharides from Panax species using GC–MS, saccharide mapping and HPSEC-RID-MALLS. Carbohydr. Polym. 2016, 153, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Khan, N.M.; Nunez, K.M.; Chess, E.K.; Szabo, C.M. Complete monosaccharide analysis by high-performance anion-exchange chromatography with pulsed amperometric detection. Anal. Chem. 2012, 84, 4104–4110. [Google Scholar] [CrossRef]
- Karlsson, G.; Winge, S.; Sandberg, H. Separation of monosaccharides by hydrophilic interaction chromatography with evaporative light scattering detection. J. Chromatogr. A 2005, 1092, 246–249. [Google Scholar] [CrossRef]
- Ghosh, R.; Kline, P. HPLC with charged aerosol detector (CAD) as a quality control platform for analysis of carbohydrate polymers. BMC Res. Notes 2019, 12, 268. [Google Scholar] [CrossRef]
- Yan, J.; Shi, S.; Wang, H.; Liu, R.; Li, N.; Chen, Y.; Wang, S. Neutral monosaccharide composition analysis of plant-derived oligo- and polysaccharides by high performance liquid chromatography. Carbohydr. Polym. 2016, 136, 1273–1280. [Google Scholar] [CrossRef]
- Li, S.-P.; Wu, D.-T.; Lv, G.-P.; Zhao, J. Carbohydrates analysis in herbal glycomics. Trac. Trends Anal. Chem. 2013, 52, 155–169. [Google Scholar] [CrossRef]
- Wang, M.; Guilbert, L.J.; Ling, L.; Li, J.; Wu, Y.; Xu, S.; Pang, P.; Shan, J.J. Immunomodulating activity of CVT-E002, a proprietary extract from North American ginseng (Panax quinquefolium). J. Pharm. Pharmacol. 2001, 53, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Oshima, Y.; Sato, K.; Hikino, H. Isolation and hypoglycemic activity of Quinquefolans A, B, and C, Glycans of Panax quinquefolium roots. J. Nat. Prod. 1987, 50, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Han, B.; Sun, Y.; Wang, Z.; Yang, X. Immunoregulatory effects of a glucogalactan from the root of Panax quinquefolium L. Carbohydr. Polym. 2012, 87, 2725–2729. [Google Scholar] [CrossRef]
- Wang, L.; Yu, X.; Yang, X.; Li, Y.; Yao, Y.; Lui, E.M.K.; Ren, G. Structural and anti-inflammatory characterization of a novel neutral polysaccharide from North American ginseng (Panax quinquefolius). Int. J. Biol. Macromol. 2015, 74, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.A.F.; Wong, M.H.T.; Stryjecki, C.; De Boer, A.; Lui, E.M.K.; Mutch, D.M. Unraveling the adipocyte inflammomodulatory pathways activated by North American ginseng. Int. J. Obes. 2013, 37, 350–356. [Google Scholar] [CrossRef]
- Trammell, R.A.; Cox, L.; Pikora, J.; Murphy, L.L.; Toth, L.A. Evaluation of an extract of North American ginseng (Panax quinquefolius L.) in Candida albicans-infected complement-deficient mice. J. Ethnopharmacol. 2012, 139, 414–421. [Google Scholar] [CrossRef]
- Estaki, M.; Noble, E.G. North American ginseng protects against muscle damage and reduces neutrophil infiltration after an acute bout of downhill running in rats. Appl. Physiol. Nutr. Metab. 2015, 40, 116–121. [Google Scholar] [CrossRef]
- Akhter, K.F.; Mumin, M.A.; Lui, E.K.; Charpentier, P.A. Microfluidic synthesis of ginseng polysaccharide nanoparticles for immunostimulating action on macrophage cell lines. ACS Biomater. Sci. Eng. 2016, 2, 96–103. [Google Scholar] [CrossRef]
- Akhter, K.F.; Mumin, M.A.; Lui, E.M.K.; Charpentier, P.A. Fabrication of fluorescent labeled ginseng polysaccharide nanoparticles for bioimaging and their immunomodulatory activity on macrophage cell lines. Int. J. Biol. Macromol. 2018, 109, 254–262. [Google Scholar] [CrossRef]
- Akhter, K.F.; Mumin, M.A.; Lui, E.M.K.; Charpentier, P.A. Immunoengineering with ginseng polysaccharide nanobiomaterials through oral administration in mice. ACS Biomater. Sci. Eng. 2019, 5, 2916–2925. [Google Scholar] [CrossRef]
- Wang, M.; Guilbert, L.J.; Li, J.; Wu, Y.; Pang, P.; Basu, T.K.; Shan, J.J. A proprietary extract from North American ginseng (Panax quinquefolium) enhances IL-2 and IFN-γ productions in murine spleen cells induced by Con-A. Int. Immunopharmacol. 2004, 4, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Biondo, P.D.; Goruk, S.; Ruth, M.R.; O’Connell, E.; Field, C.J. Effect of CVT-E002™ (COLD-fX®) versus a ginsenoside extract on systemic and gut-associated immune function. Int. Immunopharmacol. 2008, 8, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Mcelhaney, J.E.; Gravenstein, S.; Cole, S.K.; Davidson, E.; O’neill, D.; Petitjean, S.; Rumble, B.; Shan, J.J. A placebo-controlled trial of a proprietary extract of North American ginseng (CVT-E002) to prevent acute respiratory illness in institutionalized older adults. J. Am. Geriatr. Soc. 2004, 52, 13–19. [Google Scholar] [CrossRef]
- McElhaney, J.; Guilbert, L.; Reid, D.; Lowe, K.; Hooten, J.; Shan, J. COLD-FX® stimulates cell mediated immune response of peripheral leukocytes ex-vivo to Influenza Virus in National Hockey League Players. Open Nutraceuticals J. 2010, 3, 25–29. [Google Scholar] [CrossRef]
- Miller, S.C.; Ti, L.; Shan, J. Dietary supplementation with an extract of North American ginseng in adult and juvenile mice increases natural killer cells. Immunol. Investig. 2012, 41, 157–170. [Google Scholar] [CrossRef]
- Predy, G.N.; Goel, V.; Lovlin, R.; Donner, A.; Stitt, L.; Basu, T.K. Efficacy of an extract of North American ginseng containing poly-furanosyl-pyranosyl-saccharides for preventing upper respiratory tract infections: A randomized controlled trial. Can. Med Assoc. J. 2005, 173, 1043–1048. [Google Scholar] [CrossRef]
- Predy, G.N.; Goel, V.; Lovlin, R.E.; Basu, T.K. Immune modulating effects of daily supplementation of COLD-fX (a proprietary extract of North American ginseng) in healthy adults. J. Clin. Biochem. Nutr. 2006, 39, 162–167. [Google Scholar] [CrossRef][Green Version]
- Yan, J.; Ma, Y.; Zhao, F.; Gu, W.; Jiao, Y. Identification of immunomodulatory signatures induced by American ginseng in murine immune cells. Evid. Based Complementary Altern. Med. 2013, 2013, 972814. [Google Scholar] [CrossRef]
- Ebeling, C.; Wu, Y.; Skappak, C.; Gordon, J.R.; Ilarraza, R.; Adamko, D.J. Compound CVT-E002 attenuates allergen-induced airway inflammation and airway hyperresponsiveness, in vivo. Mol. Nutr. Food Res. 2011, 55, 1905–1908. [Google Scholar] [CrossRef]
- Tzianabos, A.O. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef]
- Yin, M.; Zhang, Y.; Li, H. Advances in research on immunoregulation of macrophages by plant polysaccharides. Front. Immunol. 2019, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Barton, G.M.; Kagan, J.C. A cell biological view of Toll-like receptor function: Regulation through compartmentalization. Nat. Rev. Immunol. 2009, 9, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- West, A.P.; Koblansky, A.A.; Ghosh, S. Recognition and Signaling by Toll-Like Receptors. Annu. Rev. Cell Dev. Biol. 2006, 22, 409–437. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef]
- Li, X.; Jiang, S.; Tapping, R.I. Toll-like receptor signaling in cell proliferation and survival. Cytokine 2010, 49, 1–9. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Rowe, D.C.; Golenbock, D.T. Endotoxin recognition and signal transduction by the TLR4/MD2-complex. Microbes Infect. 2004, 6, 1361–1367. [Google Scholar] [CrossRef]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar] [PubMed]
- Lu, Y.-C.; Yeh, W.-C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qi, C.; Guo, Y.; Zhou, W.; Zhang, Y. Toll-like receptor 4-related immunostimulatory polysaccharides: Primary structure, activity relationships, and possible interaction models. Carbohydr. Polym. 2016, 149, 186–206. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB signaling in macrophages: Dynamics, crosstalk, and signal integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Loh, S.H.; Park, J.-Y.; Cho, E.H.; Nah, S.-Y.; Kang, Y.-S. Animal lectins: Potential receptors for ginseng polysaccharides. J. Ginseng Res. 2017, 41, 1–9. [Google Scholar] [CrossRef]
- Osorio, F.; Sousa, C.R. Myeloid C-type lectin receptors in pathogen recognition and host defense. Immunity 2011, 34, 651–664. [Google Scholar] [CrossRef]
- Dennehy, K.M.; Brown, G.D. The role of the β-glucan receptor Dectin-1 in control of fungal infection. J. Leukoc. Biol. 2007, 82, 253–258. [Google Scholar] [CrossRef]
- Tang, J.; Lin, G.; Langdon, W.Y.; Tao, L.; Zhang, J. Regulation of C-type lectin receptor-mediated antifungal immunity. Front. Immunol. 2018, 9, 123. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Simmons, R.M.; Underhill, D.M. Dectin-1 Stimulation by Candida albicans yeast or zymosan triggers NFAT activation in macrophages and dendritic cells. J. Immunol. 2007, 178, 3107–3115. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-J.; Tang, X.-F.; Shuai, X.-X.; Jiang, C.-J.; Liu, X.; Wang, L.-F.; Yao, Y.-F.; Nie, S.-P.; Xie, M.-Y. Mannose receptor mediates the immune response to Ganoderma atrum polysaccharides in macrophages. J. Agric. Food Chem. 2017, 65, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Gazi, U.; Martinez-Pomares, L. Influence of the mannose receptor in host immune responses. Immunobiology 2009, 214, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Zamze, S.; Martinez-Pomares, L.; Jones, H.; Taylor, P.R.; Stillion, R.J.; Gordon, S.; Wong, S.Y.C. Recognition of bacterial capsular polysaccharides and lipopolysaccharides by the macrophage mannose receptor. J. Biol. Chem. 2002, 277, 41613–41623. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Niu, Y.; Xing, P.; Wang, C. Bioactive polysaccharides from natural resources including Chinese medicinal herbs on tissue repair. Chin. Med. 2018, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Prado, S.B.R.; Beukema, M.; Jermendi, E.; Schols, H.A.; de Vos, P.; Fabi, J.P. Pectin interaction with immune receptors is modulated by ripening process in papayas. Sci. Rep. 2020, 10, 1690. [Google Scholar] [CrossRef]
- Ross, G.D.; Cain, J.A.; Myones, B.L.; Newman, S.L.; Lachmann, P.J. Specificity of membrane complement receptor type three (CR3) for ß-Glucans. Complement 1987, 4, 61–74. [Google Scholar] [CrossRef]
- Vorup-Jensen, T.; Jensen, R.K. Structural immunology of complement receptors 3 and 4. Front. Immunol. 2018, 9, 2716. [Google Scholar] [CrossRef]
- Li, B.; Allendorf, D.J.; Hansen, R.; Marroquin, J.; Ding, C.; Cramer, D.E.; Yan, J. Yeast β-Glucan amplifies phagocyte killing of iC3b-opsonized tumor cells via complement receptor 3-Syk-phosphatidylinositol 3-kinase pathway. J. Immunol. 2006, 177, 1661–1669. [Google Scholar] [CrossRef]
- Thieblemont, N.; Haeffner-Cavaillon, N.; Haeffner, A.; Cholley, B.; Weiss, L.; Kazatchkine, M.D. Triggering of complement receptors CR1 (CD35) and CR3 (CD11b/CD18) induces nuclear translocation of NF-kappa B (p50/p65) in human monocytes and enhances viral replication in HIV-infected monocytic cells. J. Immunol. 1995, 155, 4861–4867. [Google Scholar]
- Huang, J.-H.; Lin, C.-Y.; Wu, S.-Y.; Chen, W.-Y.; Chu, C.-L.; Brown, G.D.; Chuu, C.-P.; Wu-Hsieh, B.A. CR3 and dectin-1 collaborate in macrophage cytokine response through association on lipid rafts and activation of Syk-JNK-AP-1 pathway. PLoS Pathog. 2015, 11, e1004985. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, X.; Chen, T.; Li, T.; Cao, K.; Lu, A.; Chen, Y.; Sun, D.; Luo, J.; Fan, J.; et al. Myelin activates FAK/Akt/NF-κB pathways and provokes CR3-dependent inflammatory response in murine system. PLoS ONE 2010, 5, e9380. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.H.; Jain, M.; Lee, K.; Kandimalla, E.R.; Faridi, M.H.; Buyon, J.P.; Gupta, V.; Clancy, R.M. Complement receptor 3 influences Toll-like receptor 7/8-dependent inflammation: Implications for autoimmune diseases characterized by antibody reactivity to ribonucleoproteins. J. Biol. Chem. 2013, 288, 9077–9083. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.L.; Fürnrohr, B.G.; Vyse, T.J.; Rhodes, B. The complement receptor 3 (CD11b/CD18) agonist Leukadherin-1 suppresses human innate inflammatory signalling. Clin. Exp. Immunol. 2016, 185, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Popov, S.V.; Ovodov, Y.S. Polypotency of the immunomodulatory effect of pectins. Biochem. Mosc. 2013, 78, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-Y.; Chiu, S.-L.; Wen, M.-H.; Chen, K.-Y.; Hua, K.-F. Ligands of macrophage scavenger receptor induce cytokine expression via differential modulation of protein kinase signaling pathways. J. Biol. Chem. 2001, 276, 28719–28730. [Google Scholar] [CrossRef]
- Kelley, J.L.; Ozment, T.R.; Li, C.; Schweitzer, J.B.; Williams, D.L. Scavenger receptor-A (CD204): A two-edged sword in health and disease. Crit. Rev. Immunol. 2014, 34, 241–261. [Google Scholar] [CrossRef]
- Ben, J.; Zhang, Y.; Zhou, R.; Zhang, H.; Zhu, X.; Li, X.; Zhang, H.; Li, N.; Zhou, X.; Bai, H.; et al. Major vault protein regulates class A scavenger receptor-mediated tumor necrosis factor-α synthesis and apoptosis in macrophages. J. Biol. Chem. 2013, 288, 20076–20084. [Google Scholar] [CrossRef]
- Means, T.K.; Mylonakis, E.; Tampakakis, E.; Colvin, R.A.; Seung, E.; Puckett, L.; Tai, M.F.; Stewart, C.R.; Pukkila-Worley, R.; Hickman, S.E.; et al. Evolutionarily conserved recognition and innate immunity to fungal pathogens by the scavenger receptors SCARF1 and CD36. J. Exp. Med. 2009, 206, 637–653. [Google Scholar] [CrossRef]
- Patten, D.A. SCARF1: A multifaceted, yet largely understudied, scavenger receptor. Inflamm. Res. 2018, 67, 627–632. [Google Scholar] [CrossRef]
- Abe, T.; Shimamura, M.; Jackman, K.; Kurinami, H.; Anrather, J.; Zhou, P.; Iadecola, C. Key role of CD36 in Toll-like receptor 2 signaling in cerebral ischemia. Stroke 2010, 41, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Watanabe, T. NOD2, an intracellular innate immune sensor involved in host defense and Crohn’s disease. Mucosal Immunol. 2011, 4, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Kim, Y.-G.; McDonald, C.; Kanneganti, T.-D.; Hasegawa, M.; Body-Malapel, M.; Inohara, N.; Núñez, G. RICK/RIP2 mediates innate immune responses induced through Nod1 and Nod2 but not TLRs. J. Immunol. 2007, 178, 2380–2386. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Fujimoto, Y.; Lucas, P.C.; Nakano, H.; Fukase, K.; Núñez, G.; Inohara, N. A critical role of RICK/RIP2 polyubiquitination in Nod-induced NF-kappaB activation. EMBO J. 2008, 27, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Rhee, M.H.; Kim, E.; Cho, J.Y. BAY 11-7082 is a broad-spectrum inhibitor with anti-inflammatory activity against multiple targets. Mediat. Inflamm. 2012, 2012, 416036. [Google Scholar] [CrossRef]
- Luettig, B.; Steinmüller, C.; Gifford, G.E.; Wagner, H.; Lohmann-Matthes, M.-L. Macrophage activation by the polysaccharide arabinogalactan isolated from plant cell cultures of Echinacea purpurea. JNCI J. Natl. Cancer Inst. 1989, 81, 669–675. [Google Scholar] [CrossRef]
- Popov, S.V.; Popova, G.Y.; Ovodova, R.G.; Bushneva, O.A.; Ovodov, Y.S. Effects of polysaccharides from Silene vulgaris on phagocytes. Int. J. Immunopharmacol. 1999, 21, 617–624. [Google Scholar] [CrossRef]
- Holderness, J.; Schepetkin, I.A.; Freedman, B.; Kirpotina, L.N.; Quinn, M.T.; Hedges, J.F.; Jutila, M.A. Polysaccharides isolated from Açaí fruit induce innate immune responses. PLoS ONE 2011, 6, e17301. [Google Scholar] [CrossRef]
- Peng, Q.; Liu, H.; Lei, H.; Wang, X. Relationship between structure and immunological activity of an arabinogalactan from Lycium ruthenicum. Food Chem. 2016, 194, 595–600. [Google Scholar] [CrossRef]
- Lee, J.-B.; Tanikawa, T.; Hayashi, K.; Asagi, M.; Kasahara, Y.; Hayashi, T. Characterization and biological effects of two polysaccharides isolated from Acanthopanax sciadophylloides. Carbohydr. Polym. 2015, 116, 159–166. [Google Scholar] [CrossRef]
- Yang, L.-C.; Lu, T.-J.; Lin, W.-C. A type II arabinogalactan from Anoectochilus formosanus for G-CSF production in macrophages and leukopenia omprovement in CT26-bearing mice treated with 5-Fluorouracil. Evid. Based Complementary Altern. Med. 2013, 2013, 458075. [Google Scholar] [CrossRef] [PubMed]
- Vogt, L.M.; Sahasrabudhe, N.M.; Ramasamy, U.; Meyer, D.; Pullens, G.; Faas, M.M.; Venema, K.; Schols, H.A.; de Vos, P. The impact of lemon pectin characteristics on TLR activation and T84 intestinal epithelial cell barrier function. J. Funct. Foods 2016, 22, 398–407. [Google Scholar] [CrossRef]
- Park, H.-R.; Lee, S.J.; Im, S.-B.; Shin, M.-S.; Choi, H.-J.; Park, H.-Y.; Shin, K.-S. Signaling pathway and structural features of macrophage-activating pectic polysaccharide from Korean citrus, Cheongkyool peels. Int. J. Biol. Macromol. 2019, 137, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Luo, Z.; Liu, D.; Ning, Z.; Yang, J.; Ren, J. Structure characterization of a novel polysaccharide from Dictyophora indusiata and its macrophage immunomodulatory activities. J. Agric. Food Chem. 2015, 63, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Kim, Y.S.; Kwak, Y.S.; Song, Y.B.; Kim, Y.S.; Park, J.D. Enhancement of antitumor effects of paclitaxel (Taxol) in combination with red ginseng acidic polysaccharide (RGAP). Planta Med. 2004, 70, 1033–1038. [Google Scholar] [CrossRef]
- Byeon, S.E.; Lee, J.; Kim, J.H.; Yang, W.S.; Kwak, Y.-S.; Kim, S.Y.; Choung, E.S.; Rhee, M.H.; Cho, J.Y. Molecular mechanism of macrophage activation by red ginseng acidic polysaccharide from Korean red ginseng. Mediat. Inflamm. 2012, 2012, 732860. [Google Scholar] [CrossRef]
- Vanisree, M.; Lee, C.-Y.; Lo, S.-F.; Nalawade, S.; Lin, C.; Tsay, H.-S. Studies on the production of some important secondary metabolites from medicinal plants by plant tissue cultures. Bot. Bull. Acad. Sin. 2004, 45, 1–22. [Google Scholar]
- Murthy, H.N.; Lee, E.-J.; Paek, K.-Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. PCTOC 2014, 118, 1–16. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Name | Structural Features of Isolated Polysaccharides | Molecular Weight (kDa) | Ref |
|---|---|---|---|
| AGC1 | Composed of Gal (60.093%), Ara (19.165%), Xyl (11.363%), Glc (6.298%), Rha (1.548%), and Man (0.79%); contains type II arabinogalactans | 5.2 | [14] |
| AGC3 | Composed of Ara (7.8%), Rha (8.1%), Glc 92%), Gal (74.3%), GalA (6.8%), GluA (1%), and trace amounts of Man and Xyl; contains RG-I polysaccharide | 4.81 and 32.14 | [37] |
| GSP | Composed of Rha, Ara, Gal, Glc, and GalA in a molar ratio of 1:4:8:8:50; pectic polysaccharide with a major homogalacturonan domain and a minor RG-I component | 85.4 | [33] |
| PPQN | Composed of Glc and Gal in a molar ratio of 1:1.15 | 3.1 | [48] |
| PPQA2 | Composed of Ara, Rha, Man, Gal, Glc, GalA, and GluA in a molar ratio of 8:4:2.9:7.2:12.5:26.6:38.8 | 23 | [38] |
| PPQA4 | Composed of Ara, Rha, Man, Gal, Glc, and GluA in a molar ratio of 19.7:5.1:8.1:23.9:41.3:2 | 120 | [38] |
| PPQA5 | Composed of Ara, Rha, Man, Gal, Glc, GalA, and GluA in a molar ratio of 8.5:3.2:5.3:10.8:32.4:15.5:24.4 | 5.3 | [38] |
| AEP-1 | Composed of Glc, Gal, and GalA in a molar ratio of 4.67:0.97:3.92 | N/A | [31] |
| AEP-2 | Composed of Ara, Man, Gal, Glc, and GalA in a relative molar ratio of 1.03:0.76:1.68:3.02:3.65 | N/A | [31] |
| WPS-1 | Composed of Ara, Rha, Man, Gal, and Glc in a ratio of 21.2:2.3:2.6:18.7:55.2 | 1540 | [35] |
| WPS-2 | Composed of Ara, Rha, Man, Gal, and Glc in a ratio of 27.9:1.7:2.9:20.7:46.8 | 14.1 | [35] |
| SPS-1 | Composed of Ara, Xyl, Man, Gal, Glc, Gala, and GlcA in a ratio of 22.3:6.9:9.2:28.6:15.9:13.6:3.5 | 362 | [35] |
| SPS-2 | Composed of Ara, Xyl, Man, Gal, Glc, Gala, and GlcA in a ratio of 14.2:5.3:7.9:22.5:25.3:16.9:7.9 | 9700 | [35] |
| SPS-3 | Composed of Ara, Rha, Xyl, Man, Gal, Glc, GalA, and GlcA in a ratio of 19.2:2.1:9.6:12:15.2:11.5:26.3:4.1 | 512 | [35] |
| PPQ | Composed of Glc and Gal in a molar ratio of 2.1:1 | 54 | [47] |
| Quinquefolan A | Composed of Man and Glc in a ratio of 1.0:2.3 and 10.8% uronic acid | >2000 | [46] |
| Quinquefolan B | Composed of Man and Glc in a ratio of 1.0:5.5 and 11.7% uronic acid | >2000 | [46] |
| Quinquefolan C | Composed of Xyl and 7.1% uronic acid | >2000 | [46] |
| Name of Polysaccharide | Immunomodulatory Findings | Model | Endotoxin Test | Ref |
|---|---|---|---|---|
| N/A (Crude polysaccharide) | Stimulated TNF-α | Alveolar macrophages isolated from male Wistar rats | Yes | [29] |
| N/A (Polysaccharide Nanoparticles) | Stimulated NO, TNF-α, IL-6, and IL-1β | RAW 264.7 murine macrophage cells | Not reported | [52] |
| N/A (Fluorescein-5-thiosemicarbazide labelled polysaccharide nanoparticles | Uptake of nanoparticles by macrophages, stimulated NO, TNF-α, IL-6, and IL-1β | RAW 264.7 murine macrophage cells | Not reported | [53] |
| N/A Gelatin-encapsulated Polysaccharide Nanoparticles | Stimulated NO, TNF-α, IL-6, and IL-1B (in vitro); stimulated NO and TNF-α in blood serum (in vivo) | RAW 264.7 murine macrophage cells (in vitro); swiss albino mice (in vivo) | Not reported | [54] |
| N/A (crude and partially purified polysaccharide fractions of different molecular weights) | Stimulated NO and TNF-α (in vitro, ex vivo, and in vivo); suppressed NO and TNF-α in LPS-stimulated cells (in vitro, ex vivo, and in vivo) | RAW 264.7 murine macrophage cells (in vitro); alveolar macrophages isolated from adult male rats (ex vivo); adult male rats (in vivo) | Not reported | [12] |
| AGC1 (Purified neutral polysaccharide) | Stimulated TNF-α, IL-6, MCP-1, GM-CSF, NOS2 gene expression, iNOS protein expression, NO (in vitro); stimulated splenocyte proliferation (ex vivo) | RAW 264.7 murine macrophage cells (in vitro); splenocytes isolated from CD1 mice (ex vivo) | Yes | [14] |
| AGC3 (Purified acidic polysaccharide fraction) | Stimulated TNF-α, IL-6, and NO (in vitro); upregulated phospho-p65 and phospho-p38 (in vitro); stimulated splenocyte proliferation (ex vivo) | RAW 264.7 murine macrophage cells (in vitro); splenocytes isolated from CD1 mice (ex vivo) | Yes | [37] |
| N/A (High molecular weight crude polysaccharides) | Stimulated IL-6, TNF-α, IFN-y, IL-1β, IL-12, IL-2; induction of Th1 immune response; induced NF-κB, MAPK and PI3K pathways | Human peripheral blood mononuclear cells, CD14+ monocytes, CD14+CD16+ monocytes | Yes | [30] |
| PPQN (Purified neutral polysaccharide) | Suppressed NO, IL-1β, IL-6, and TNF-α in LPS-stimulated macrophages | RAW 264.7 murine macrophage cells | Not reported | [48] |
| PPQA2, PPQA4, PPQA5 (Purified acidic polysaccharides) | Stimulated NO, TNF-α, and IL-6 | RAW 264.7 murine macrophage cells | Not reported | [38] |
| N/A (Crude polysaccharide) | Stimulated IL-6, CCL5, TNF-α, and NF-KB | Mouse 3T3-L1 preadipocyte cells | Not reported | [49] |
| AGP (Crude Polysaccharide) | Stimulated NO, IL-6, and IL-10 | RAW 264.7 murine macrophage cells | Not reported | [34] |
| AEP (alkali-extractable crude polysaccharide), AEP-2 (Purified acidic polysaccharide) | Stimulated NO, IL-6, and TNF-α | RAW 264.7 murine macrophage cells | Not reported | [31] |
| CPS (crude polysaccharide), WPS-1, WPS-2 (purified neutral polysaccharides), SPS-1, SPS-2, and SPS-3 (purified acidic polysaccharides) | Stimulated macrophage phagocytic activity and NO; augmented LPS and ConA induced splenocytic proliferation (SPS-3 > SPS-1 > CPS> WPS-1 > WPS-2 > SPS-2) | Splenocytes and peritoneal macrophages isolated from specific pathogen-free Jilin mice | Not reported | [35] |
| PPQ (purified neutral polysaccharide) | Decreased tumor size; stimulated serum IL-2 and IFN-γ; reduced serum IL-10; increased thymus and spleen index in tumor-bearing mice | C57BL/6 mice (Lewis lung carcinoma model) | Not reported | [47] |
| Immunomodulatory Activities of CVT-E002/COLD-FX | Model | Ref |
|---|---|---|
| Decreased spleen IL-2 and IFN-γ and increased IL-1β following LPS/ConA stimulation of cultured lymphocytes; lowered proportion of CD3+ (T-cells) and activated T-cells | Sprague-Dawley rats (oral administration) followed by isolation and culture of lymphocytes from spleen, mesenteric lymph nodes, and Peyer’s patches | [56] |
| Reduced acute respiratory illness due to influenza or respiratory syncytial virus | Randomized, placebo controlled, double blind human clinical trial (oral administration) | [57] |
| Stimulated TNF-α and IL-2; moderated granzyme-B levels | Human (oral administration) followed by isolation and culture of peripheral blood leukocytes along with influenza viruses | [58] |
| Increased NK cell numbers in spleen, bone marrow, and blood | C3H/OuJ mice (oral administration) | [59] |
| Reduced recurrent and mean number of colds per person and severity of symptoms | Randomized, placebo controlled, double blind human trial (oral administration) | [60] |
| Increased NK cell and T-helper cell numbers and reduced IgA levels in plasma | Randomized, placebo controlled, double blind human trial (oral administration) | [61] |
| Stimulated B lymphocyte proliferation; stimulated IgG in blood serum; stimulated IL-1, IL-6, TNF-α, and NO | Lymphocytes isolated from BALB/c mice; BALB/c mice (oral administration); peritoneal exudate macrophages isolated from C57BL/6 mice | [45] |
| Stimulated ConA-induced IFN-γ and IL-2 | Splenocytes isolated from C57BL/6 mice | [55] |
| Suppressed IFN-γ and related gene expression in dexamethasone cotreated cells | Mouse splenocytes | [62] |
| Inhibited allergic airway inflammation; increased INF-γ, regulatory T-cells, and IL-10 in lungs | BALB/c mice | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, R.; Bryant, D.L.; Farone, A.L. Panax quinquefolius (North American Ginseng) Polysaccharides as Immunomodulators: Current Research Status and Future Directions. Molecules 2020, 25, 5854. https://doi.org/10.3390/molecules25245854
Ghosh R, Bryant DL, Farone AL. Panax quinquefolius (North American Ginseng) Polysaccharides as Immunomodulators: Current Research Status and Future Directions. Molecules. 2020; 25(24):5854. https://doi.org/10.3390/molecules25245854
Chicago/Turabian StyleGhosh, Rajarshi, Daniel L. Bryant, and Anthony L. Farone. 2020. "Panax quinquefolius (North American Ginseng) Polysaccharides as Immunomodulators: Current Research Status and Future Directions" Molecules 25, no. 24: 5854. https://doi.org/10.3390/molecules25245854
APA StyleGhosh, R., Bryant, D. L., & Farone, A. L. (2020). Panax quinquefolius (North American Ginseng) Polysaccharides as Immunomodulators: Current Research Status and Future Directions. Molecules, 25(24), 5854. https://doi.org/10.3390/molecules25245854