
Anti-Inflammatory Effect of Homo- and Heterodimers of Natural Enkephalinase Inhibitors in Experimental Colitis in Mice



Abstract
1. Introduction
2. Results and Discussion
2.1. Peptide Synthesis and Purification
2.2. Effect of Sialorphin, Opiorphin and Homo- and Heterodimers on Degradation of Met-Enkephalin by NEP
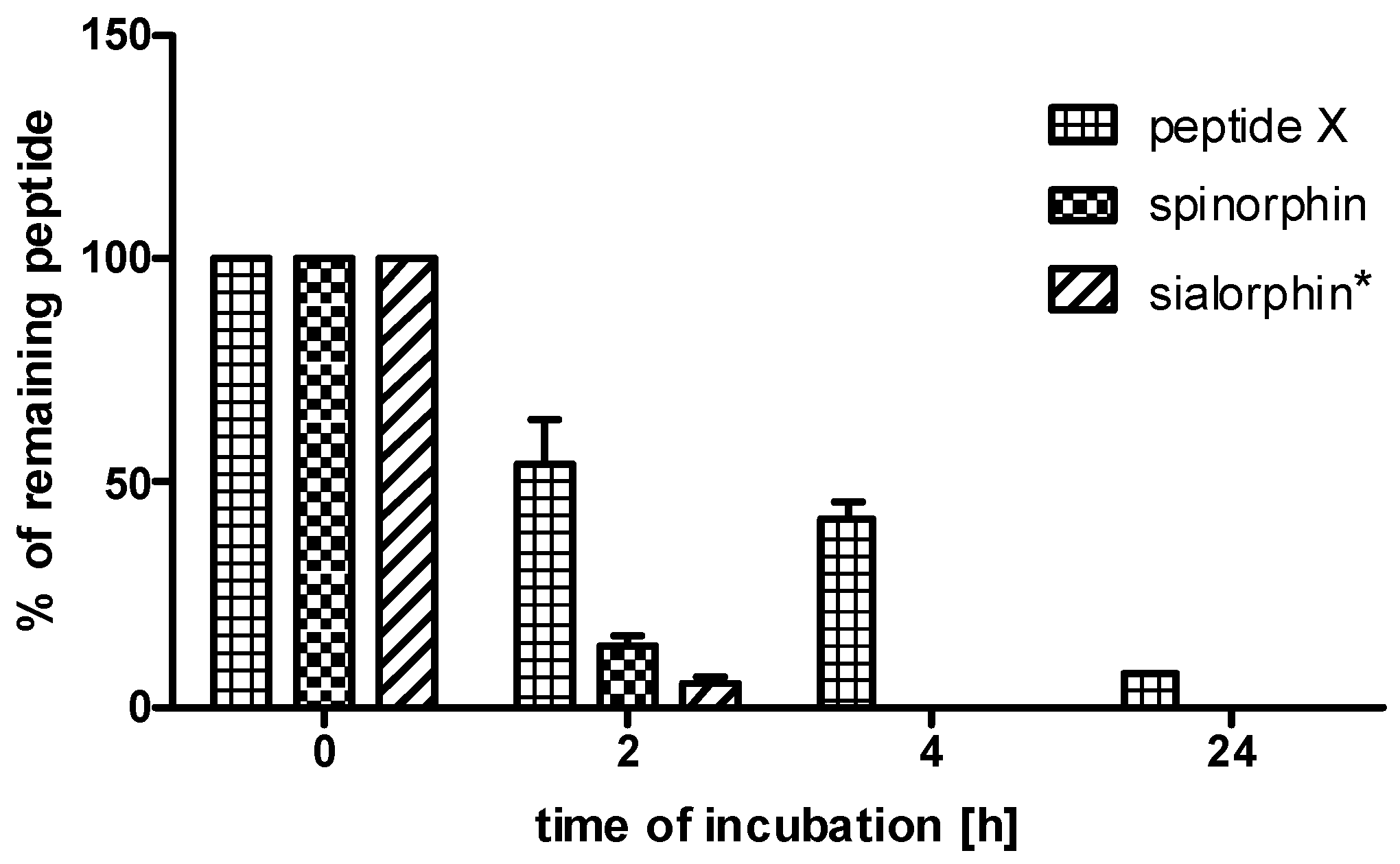
2.3. Stability in Human Plasma
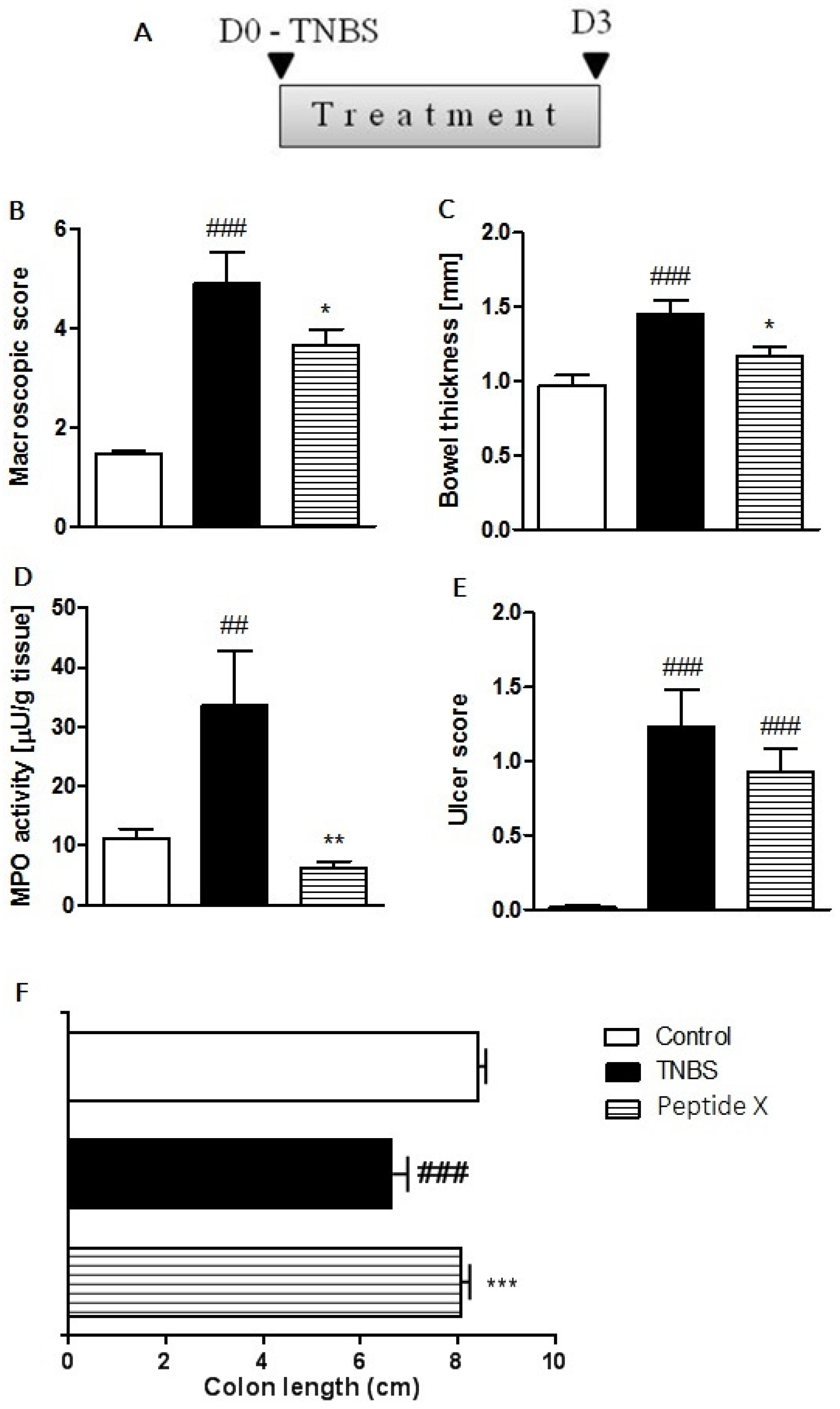
2.4. Anti-Inflammatory Effect in TNBS-Induced Colitis
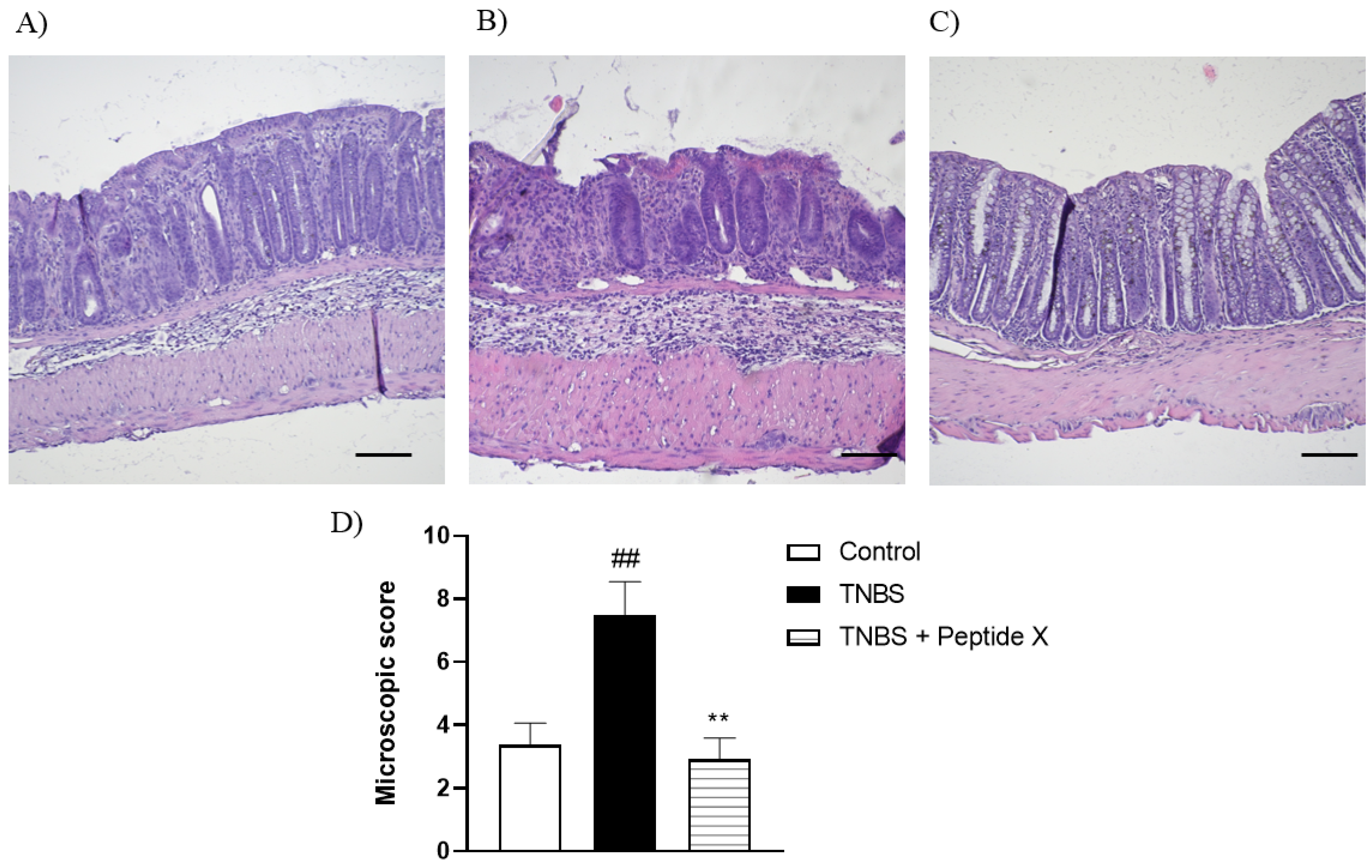
2.5. Histological
3. Materials and Methods
3.1. Peptide Synthesis
3.2. Peptide Purification and Analysis
3.3. Homo- and Heterodimers Synthesis
3.4. Determination of Enkephalins Degradation Rates
3.5. Stability of Sialorphin, Spinorphin and Peptide X in the Human Plasma
3.6. Animals
3.7. TNBS Model
3.8. Evaluation of Colonic Damage
3.9. Determination of Tissue Myeloperoxidase Activity
3.10. Histology
3.11. Statistics
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Baumgart, D.C.; Carding, S.R. Inflammatory bowel disease: Cause and immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef]
- Sobczak, M.; Fabisiak, A.; Murawska, N.; Wesołowska, E.; Wierzbicka, P.; Wlazłowski, M.; Wójcikowska, M.; Zatorski, H.; Zwolińska, M.; Fichna, J. Current overview of extrinsic and intrinsic factors in etiology and progression of inflammatory bowel diseases. Pharmacol. Rep. 2014, 66, 766–775. [Google Scholar] [CrossRef]
- Jump, R.L.; Levine, A.D. Mechanisms of natural tolerance in the intestine: Implications for inflammatory bowel disease. Inflamm. Bowel Dis. 2004, 10, 462–478. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Biancheri, P.; Rovedatti, L.; MacDonald, T.T.; Corazza, G.R. New pathogenic paradigms in inflammatory bowel disease. Inflamm. Bowel Dis. 2012, 18, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Mosińska, P.; Zielińska, M.; Fichna, J. Expression and physiology of opioid receptors in the gastrointestinal tract. Curr. Opin. Endocrinol. Diabetes Obes. 2016, 23, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, D.B.; Maric, D. In vivo modulation of the immune system by enkephalins. Int. J. Neurosci. 1990, 51, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Salzet, M.; Tasiemski, A. Involvement of pro-enkephalin-derived peptides in immunity. Dev. Comp. Immunol. 2001, 25, 177–185. [Google Scholar] [CrossRef]
- Bilkei-Gorzo, A.; Racz, I.; Michel, K.; Mauer, D.; Zimmer, A.; Klingmüller, D.; Zimmer, A. Control of hormonal stress reactivity by the endogenous opioid system. Psychoneuroendocrinology 2008, 33, 425–436. [Google Scholar] [CrossRef]
- Marini, M.; Roscetti, G.; Bongiorno, L.; Urbani, A.; Roda, L.G. Hydrolysis and protection from hydrolysis of enkephalins in human plasma. Neurochem. Res. 1990, 15, 61–67. [Google Scholar] [CrossRef]
- Roques, B.P.; Noble, F.; Dauge, V.; Fournié-Zaluski, M.C.; Beaumont, A. Neutral endopeptidase 24.11: Structure, inhibition, and experimental and clinical pharmacology. Pharmacol. Rev. 1993, 45, 87–146. [Google Scholar]
- Gabrilovac, J.; Cupic, B.; Breljak, D.; Zekusić, M.; Boranić, M. Expression of CD13/aminopeptidase N and CD10/neutral endopeptidase on cultured human keratinocytes. Immunol. Lett. 2004, 91, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Rougeot, C.; Messaoudi, M.; Hermitte, V.; Rigault, A.G.; Blisnick, T.; Dugave, C.; Desor, D.; Rougeon, F. Sialorphin, a natural inhibitor of rat membrane-bound neutral endopeptidase that displays analgesic activity. Proc. Natl. Acad. Sci. USA 2003, 100, 8549–8554. [Google Scholar] [CrossRef]
- Wisner, A.; Dufour, E.; Messaoudi, M.; Nejdi, A.; Marcel, A.; Ungeheuer, M.N.; Rougeot, C. Human opiorphin, a natural antinociceptive modulator of opioid-dependent pathways. Proc. Natl. Acad. Sci. USA 2006, 103, 17979–17984. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Hazato, T. Spinorphin, a new inhibitor of enkephalin-degrading enzymes derived from the bovine spinal cord. Masui 1993, 42, 1497–1503. [Google Scholar] [PubMed]
- Kamysz, E.; Sałaga, M.; Sobocińska, M.; Giełdoń, A.; Fichna, J. Antiinflammatory effect of novel analogs of natural enkephalinase inhibitors in a mouse model of experimental colitis. Future Med. Chem. 2016, 8, 2231–2243. [Google Scholar] [CrossRef]
- Sałaga, M.; Mokrowiecka, A.; Jacenik, D.; Cygankiewicz, A.I.; Malecka-Panas, E.; Kordek, R.; Krajewska, W.M.; Sobocinska, M.K.; Kamysz, E.; Fichna, J. Systemic administration of sialorphin attenuates experimental colitis in mice via interaction with mu and kappa opioid receptors. J. Crohns. Colitis 2017, 11, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Popik, P.; Kamysz, E.; Kreczko, J.; Wróbel, M. Human opiorphin: The lack of physiological dependence, tolerance to antinociceptive effects and abuse liability in laboratory mice. Behav. Brain Res. 2010, 213, 88–93. [Google Scholar] [CrossRef]
- Kamysz, E.; Sałaga, M.; Sobczak, M.; Kamysz, W.; Fichna, J. Characterization of the effects of opiorphin and sialorphin and their analogs substituted in position 1 with pyroglutamic acid on motility in the mouse ileum. J. Pept. Sci. 2013, 19, 166–172. [Google Scholar] [CrossRef]
- Schilling, S.; Hoffmann, T.; Manhart, S.; Hoffmann, M.; Demuth, H. Glutaminyl cyclases unfold glutamyl cyclase activity under mild acid conditions. FEBS Lett. 2004, 563, 191–196. [Google Scholar] [CrossRef]
- Bogeas, A.; DuFour, E.; Bisson, J.F.; Messaoudi, M.; Rougeot, C. Structure–activity relationship study and function-based petidomimetic design of human opiorphin with improved bioavailability property and unaltered analgesic activity. Biochem. Pharmacol. 2013, 2, 1000122. [Google Scholar]
- Sobocińska, M.; Giełdoń, A.; Fichna, J.; Kamysz, E. 1‑Substituted sialorphin analogues -synthesis, molecular modelling and in vitro effect on enkephalins degradation by NEP. Amino Acids 2019, 51, 1201. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.C.; White, P.D. Fmoc Solid Phase Peptide Synthesis. In A Practical Approach; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Pinto, M.; Rougeot, C.; Gracia, L.; Rosa, M.; García, A.; Arsequell, G.; Valencia, G.; Centeno, N.B. Proposed bioactive conformations of opiorphin, an endogenous dual APN/NEP inhibitor. ACS Med. Chem. Lett. 2012, 3, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Sobocińska, M.; Giełdoń, A.; Fichna, J.; Kamysz, E. Alanine scan of sialorphin and its hybrids with opiorphin: Synthesis, molecular modelling and effect on enkephalins degradation. Amino Acids 2018, 50, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Barlos, K.; Chatzi, O.; Gatos DStavropoulos, G. 2-Chlorotrityl chloride resin. Studies on anchoring of Fmoc-amino acids and peptide cleavage. Int. J. Pept. Protein Res. 1991, 37, 513–520. [Google Scholar]
- Vojkovsky, T. Detection of secondary amines on solid phase. Pept. Res. 1995, 8, 236–237. [Google Scholar]
- Kamysz, E.; Sikorska, E.; Dawgul, M.; Tyszkowski, R.; Kamysz, W. Influence of Dimerization of Lipopeptide Laur-Orn-Orn-Cys–NH2 and an N-terminal Peptide of Human Lactoferricin on Biological Activity. Int. J. Pept. Res. Ther. 2015, 21, 39–46. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peptide | Name | Molecular Formula | HPLC tR(min) | [M]+ Calc. | [M + H]+ Found |
|---|---|---|---|---|---|
| I | Sialorphin (Gln-His-Asn-Pro-Arg) | C26H42N12O8 | 5.5 * | 650.3 | 651.0 |
| II | Opiorphin (Gln-Arg-Phe-Ser-Arg) | C29H48N12O8 | 6.7 * | 692.3 | 693.2 |
| III | Spinorphin (Leu-Val-Val-Tyr-Pro-Trp-Thr) | C45H64N8O10 | 8.3 ** | 876.5 | 877.4 |
| IV | Cys-Gln-His-Asn-Pro-Arg | C29H47N13O9S | 6.2 * | 753.3 | 754.2 |
| V | Cys-Gln-Arg-Phe-Ser-Arg | C32H53N13O9S | 8.8 * | 795.3 | 796.3 |
| VI | Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr | C48H69N9O11S | 7.3 ** | 979.2 | 1002.4 [M + Na]+ |
| VII | (Cys-Gln-His-Asn-Pro-Arg)2 | C58H92N26O18S2 | 7.6 * | 1504.6 | 1505.9 |
| VIII | (Cys-Gln-Arg-Phe-Ser-Arg)2 | C64H104N26O18S2 | 9.4 * | 1588.7 | 1589.2 |
| IX | (Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr)2 | C96H136N18O22S2 | 9.9 ** | 1956.9 | 1957.5 |
| X | Cys-Gln-His-Asn-Pro-Arg │ Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr | C77H114N22O20S2 | 18.6 *** | 1731.0 | 1732.5 |
| XI | Cys-Gln-Arg-Phe-Ser-Arg │ Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr | C80H120N22O20S2 | 19.4 *** | 1772.8 | 1774.2 |
| Peptide | Inhibitor | 1000 × k [1/min] | t1/2 [min] |
|---|---|---|---|
| Without inhibitor * | 25.35 ± 1.05 | 27 ± 1 | |
| I | Sialorphin * (Gln-His-Asn-Pro-Arg) | 8.84 ± 0.27 ** | 78 ± 2 ** |
| II | Opiorphin * (Gln-Arg-Phe-Ser-Arg) | 7.38 ± 0.20 ** | 93 ± 2 ** |
| V | Cys-Gln-Arg-Phe-Ser-Arg | 3.47 ± 0.12 ** | 199 ± 7 ** |
| VI | Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr | 6.43 ± 0.17 ** | 107 ± 3 ** |
| VII | (Cys-Gln-His-Asn-Pro-Arg)2 | 4.75 ± 0.08 ** | 145 ± 2 ** |
| VIII | (Cys-Gln-Arg-Phe-Ser-Arg)2 | 1.53 ± 0.01 ** | 451 ± 2 ** |
| X | Cys-Gln-His-Asn-Pro-Arg │ Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr | 3.27 ± 0.10 ** | 211 ± 6 ** |
| XI | Cys-Gln-Arg-Phe-Ser-Arg │ Cys-Leu-Val-Val-Tyr-Pro-Trp-Thr | 1.13 ± 0.04 ** | 612 ± 22 ** |
Sample Availability: Samples of the compounds are not available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobocińska, M.; Salaga, M.; Fichna, J.; Kamysz, E. Anti-Inflammatory Effect of Homo- and Heterodimers of Natural Enkephalinase Inhibitors in Experimental Colitis in Mice. Molecules 2020, 25, 5820. https://doi.org/10.3390/molecules25245820
Sobocińska M, Salaga M, Fichna J, Kamysz E. Anti-Inflammatory Effect of Homo- and Heterodimers of Natural Enkephalinase Inhibitors in Experimental Colitis in Mice. Molecules. 2020; 25(24):5820. https://doi.org/10.3390/molecules25245820
Chicago/Turabian StyleSobocińska, Małgorzata, Maciej Salaga, Jakub Fichna, and Elżbieta Kamysz. 2020. "Anti-Inflammatory Effect of Homo- and Heterodimers of Natural Enkephalinase Inhibitors in Experimental Colitis in Mice" Molecules 25, no. 24: 5820. https://doi.org/10.3390/molecules25245820
APA StyleSobocińska, M., Salaga, M., Fichna, J., & Kamysz, E. (2020). Anti-Inflammatory Effect of Homo- and Heterodimers of Natural Enkephalinase Inhibitors in Experimental Colitis in Mice. Molecules, 25(24), 5820. https://doi.org/10.3390/molecules25245820