
Chinese Yellow Rice Wine Processing with Reduced Ethyl Carbamate Formation by Deleting Transcriptional Regulator Dal80p in Saccharomyces cerevisiae



Abstract
1. Introduction
2. Results and Discussion
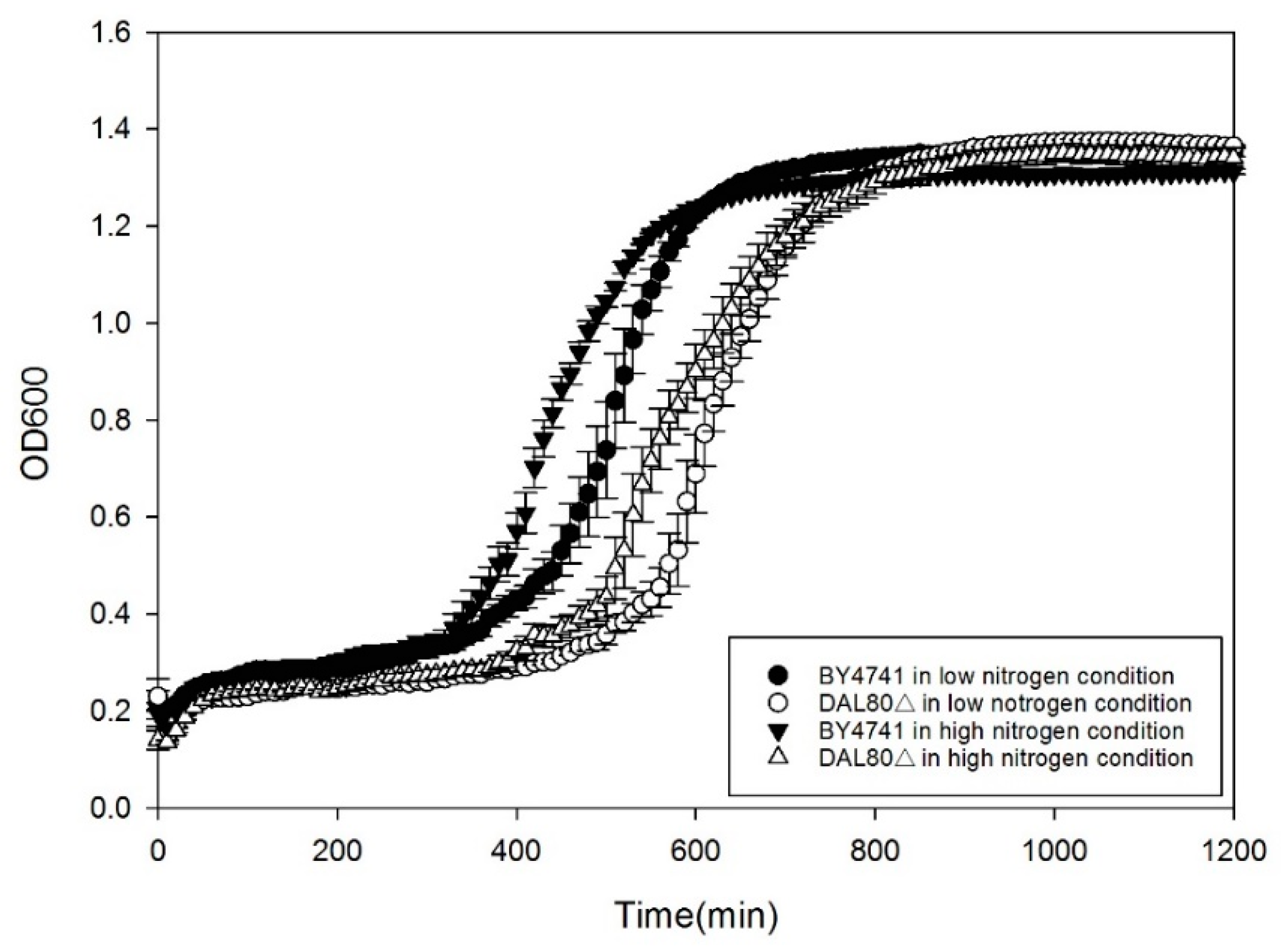
2.1. Effect of DAL80 Deletion on Cell Growth, Intracellular Arginase Activity and Urease Activity
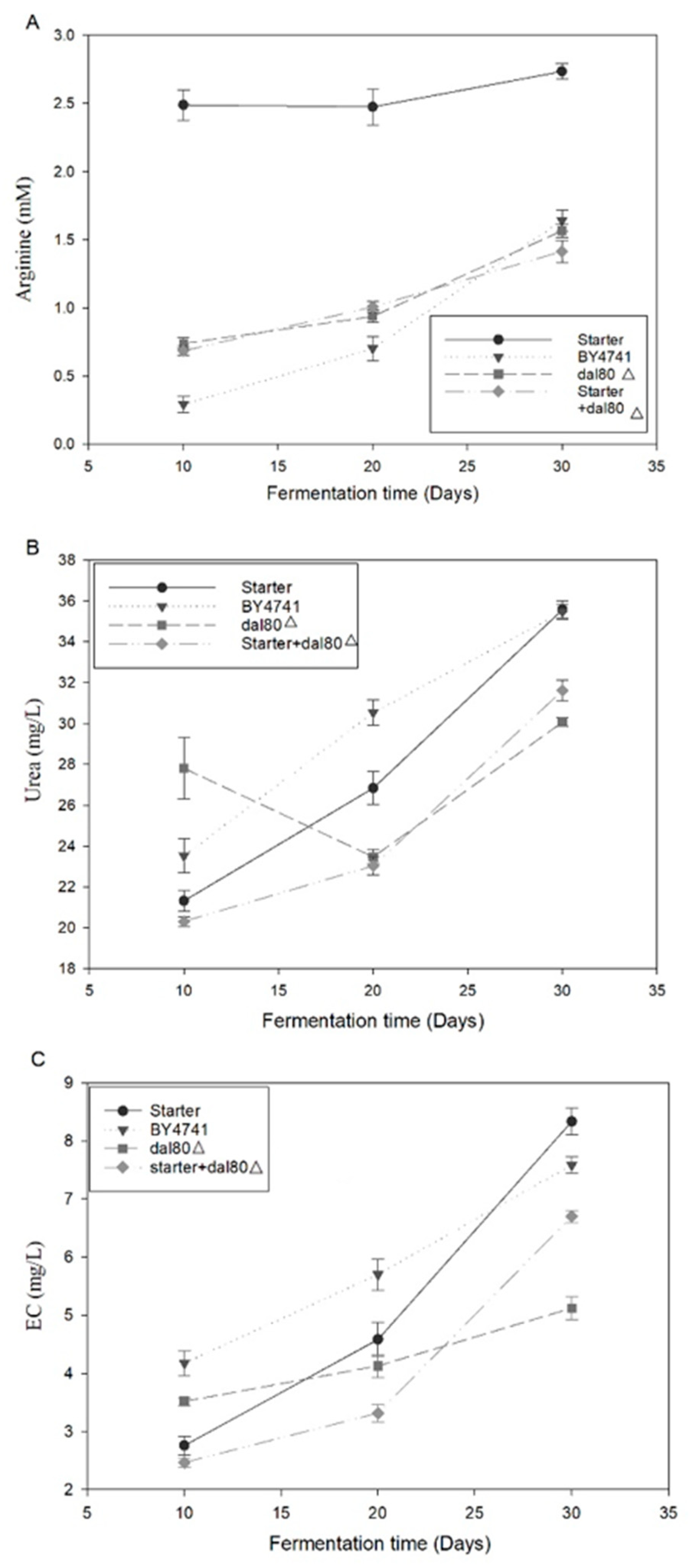
2.2. Comparative Analysis of Urea and Arginine Metabolism
2.3. Transcriptional Analysis of Genes Related to Arginine and Urea Metabolism
2.4. The Genetic Strain with the Modification of DAL80 Can Reduce Urea Accumulation during Chinese Rice Wine Fermentation
2.5. DAL80 Deletion Can Decrease EC Formation during Chinese Rice Wine Fermentation
3. Materials and Methods
3.1. Strains, Plasmids, and Culture Conditions
3.2. Yeast Cell Growth Curve Assay
3.3. Reverse-Transcribed Quantitative PCR (RT-qPCR) Analysis
3.4. Shaking-Flask Cultivation
3.5. Chinese Yellow Rice Wine Fermentation with Genetically Modified Strain
3.6. Assay of Arginase Activity, Urease Activity, Urea, Arginine, EC, Ethanol, and Amino Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baan, R.; Straif, K.; Grosse, Y.; Secretan, B.; El Ghissassi, F.; Bouvard, V.; Altieri, A.; Cogliano, V. Carcinogenicity of alcoholic beverages. Lancet Oncol. 2007, 8, 292–293. [Google Scholar] [CrossRef]
- Dahabieh, M.S.; Husnik, J.I.; van Vuuren, H.J.J. Functional enhancement of Sake yeast strains to minimize the production of ethyl carbamate in Sake wine. J. Appl. Microbiol. 2010, 109, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Alberts, P.; Stander, M.A.; De Villiers, A. Development of a novel solid-phase extraction, LC-MS/MS method for the analysis of ethyl carbamate in alcoholic beverages: Application to South African wine and spirits. Food Addit. Contam. Part A 2011, 28, 826–839. [Google Scholar] [CrossRef] [PubMed]
- Alcarde, A.R.; Souza, L.M.; Bortoletto, A.M. Ethyl carbamate kinetics in double distillation of sugar cane spirit. Part 2: Influence of type of pot still. J. Inst. Brew. 2012, 118, 352–355. [Google Scholar]
- Wu, P.G.; Pan, X.D.; Wang, L.Y.; Shen, X.H.; Yang, D.J. A survey of ethyl carbamate in fermented foods and beverages from Zhejiang, China. Food Control. 2012, 23, 286–288. [Google Scholar] [CrossRef]
- Stevens, D.F.; Ough, C.S. Ethyl carbamate formation-reaction of urea and citrulline with ethanol in wine under low to normal temperature conditions. Am. J. Enol. Vitic. 1993, 44, 309–312. [Google Scholar]
- Zhao, X.R.; Zou, H.J.; Fu, J.W.; Zhou, J.W.; Du, G.C.; Chen, J. Metabolic engineering of the regulators in nitrogen catabolite repression to reduce the production of ethyl carbamate in a model rice wine system. Appl. Environ. Microbiol. 2014, 80, 392–398. [Google Scholar] [CrossRef]
- Cooper, T.G.; Sumrada, R.A. What is the function of nitrogen catabolite repression in saccharomyces-cerevisiae. J. Bacteriol. 1983, 155, 623–627. [Google Scholar] [CrossRef]
- Wiame, J.M.; Grenson, M.; Arst, H.N. Nitrogen catabolite repression in yeasts and filamentous fungi. Adv. Microb. Physiol. 1985, 26, 1–88. [Google Scholar]
- Beltran, G.; Novo, M.; Rozes, N.; Mas, A.; Guillamon, J.M. Nitrogen catabolite repression in Saccharomyces cerevisiae during wine fermentations. FEMS Yeast Res. 2004, 4, 625–632. [Google Scholar] [CrossRef]
- Crépin, L.; Nidelet, T.; Sanchez, I.; Dequin, S.; Camarasa, C. Sequential use of nitrogen compounds by yeast during wine fermentation: A model based on kinetic and regulation characteristics of nitrogen permeases. Appl. Environ. Microbiol. 2012, 78, 8102–8111. [Google Scholar] [CrossRef] [PubMed]
- Coffman, J.A.; Cooper, T.G. Nitrogen GATA factors participate in transcriptional regulation of vacuolar protease genes in Saccharomyces cerevisiae. J. Bacteriol. 1997, 179, 5609–5613. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ljungdahl, P.O.; Daignan-Fornier, B. Regulation of amino acid, nucleotide, and phosphate metabolism in saccharomyces cerevisiae. Genetics 2012, 190, 885–929. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.R.; Zou, H.J.; Chen, J.; Du, G.C.; Zhou, J.W. The modification of Gat1p in nitrogen catabolite repression to enhance non-preferred nitrogen utilization in Saccharomyces cerevisiae. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Z.H.; Dong, Y.C.; Chen, Q.H. Comparative transcriptomic analysis of Saccharomyces cerevisiae under limited nitrogen conditions during fermentation of Chinese rice wine. Metabolomics 2016, 6, 1–6. [Google Scholar]
- Cunningham, T.S.; Cooper, T.G. Expression of the dal80 gene, whose product is homologous to the gata factors and is a negative regulator of multiple nitrogen catabolic genes in saccharomyces-cerevisiae, is sensitive to nitrogen catabolite repression. Cell Mol. Biol. 1991, 11, 6205–6215. [Google Scholar] [CrossRef]
- Jiao, Z.H.; Dong, Y.C.; Chen, Q.H. Ethyl carbamate in fermented beverages: Presence, analytical chemistry, formation mechanism, and mitigation proposals. Compr. Rev. Food Sci. Food Saf. 2014, 13, 611–626. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, W.; Zheng, D.; Zhou, Z.; Yu, W.; Zhang, L.; Feng, L.; Liang, X.; Guan, W.; Zhou, J.; et al. Genomic evolution of Saccharomyces cerevisiae under Chinese rice wine fermentation. Genome Biol. Evol. 2014, 6, 2516–2526. [Google Scholar] [CrossRef]
- Monteiro, F.F.; Trousdale, E.K.; Bisson, L.F. Ethyl carbamate formation in wine—Use of radioactively labeled precursors to demonstrate the involvement of urea. Am. J. Enol. Vitic. 1989, 40, 1–8. [Google Scholar]
- Fang, R.S.; Dong, Y.C.; Li, H.J.; Chen, Q.H. Ethyl carbamate formation regulated by Saccharomyces cerevisiae ZJU in the processing of Chinese yellow rice wine. Int. J. Food Sci. Technol. 2015, 50, 626–632. [Google Scholar] [CrossRef]
- Arena, M.E.; Saguir, F.M.; Manca de Nadra, M.C. Arginine, citrulline and ornithine metabolism by lactic acid bacteria from wine. Int. J. Food Microbiol. 1999, 52, 155–161. [Google Scholar] [CrossRef]
- Wang, P.; Sun, J.; Li, X.; Wu, D.; Li, T.; Lu, J.; Chen, J.; Xie, G. Contribution of citrulline to the formation of ethyl carbamate during Chinese rice wine production. Food Addit. Contam. Part A Chem. Anal. Control. Exp. Risk Assess. 2014, 31, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Livak KJ, S.T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Clark, S.; Francis, P.S.; Conlan, X.A.; Barnett, N.W. Determination of urea using high-performance liquid chromatography with fluorescence detection after automated derivatisation with xanthydrol. J. Chromatogr. A 2007, 1161, 207–213. [Google Scholar] [CrossRef]
- Cigic, I.K.; Vodosek, T.V.; Kosmerl, T.; Strlic, M. Amino acid quantification in the presence of sugars using hplc and pre-column derivatization with 3-MPA/OPA and FMOC-Cl. Acta Chim. Slov. 2008, 55, 660–664. [Google Scholar]
- Fu, M.L.; Liu, J.; Chen, Q.H.; Liu, X.J.; He, G.Q.; Chen, J.C. Determination of ethyl carbamate in Chinese yellow rice wine using high-performance liquid chromatography with fluorescence detection. Int. J. Food Sci. Technol. 2010, 45, 1297–1302. [Google Scholar] [CrossRef]
- Fang, R.; Dong, Y.; Xu, T.; Fan, L.; Chen, Q. Regulatory effect of arginine metabolism enzymes on ethyl carbamate formation during the fermentation of Chinese yellow rice wine. J. Zhejiang Univ. Sci. 2013, 39, 203–208. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Starter | BY4741 | dal80Δ | Starter + dal80Δ |
|---|---|---|---|---|
| Ethanol (% vol) | 11.90 ± 0.20 | 11.40 ± 0.30 | 10.20 ± 0.30 | 13.40 ± 0.50 |
| Amino acids (mg/liter) | ||||
| Asparagine | 125.32 ± 0.56 | 124.65 ± 0.56 | 134.32 ± 0.46 | 114.32 ± 0.53 |
| Threonine | 60.23 ± 0.32 | 54.65 ± 0.16 | 56.98 ± 0.52 | 58.65 ± 0.14 |
| Serine | 112.32 ± 0.71 | 116.65 ± 0.76 | 115.65 ± 0.33 | 130.32 ± 0.58 |
| Glutamate | 169.12 ± 0.78 | 145.65 ± 0.58 | 169.32 ± 0.54 | 164.32 ± 0.62 |
| Glycine | 150.32 ± 0.64 | 154.32 ± 0.46 | 171.32 ± 0.5 | 162.32 ± 0.27 |
| Alanine | 333.32 ± 0.93 | 352.32 ± 1.46 | 361.06 ± 0.41 | 345.32 ± 0.74 |
| Cystine | 100.32 ± 0.55 | 114.65 ± 0.66 | 133.32 ± 0.52 | 129.32 ± 0.53 |
| Valine | 127.65 ± 0.56 | 134.65 ± 0.44 | 140.65 ± 0.26 | 139.32 ± 0.25 |
| Methionine | 9.32 ± 0.06 | 10.65 ± 0.02 | 9.65 ± 0.02 | 11.65 ± 0.62 |
| Isoleucine | 90.32 ± 0.26 | 100.32 ± 0.40 | 95.65 ± 0.63 | 97.65 ± 0.27 |
| Leucine | 216.32 ± 0.66 | 241.65 ± 0.52 | 236.3 ± 0.62 | 244.32 ± 0.36 |
| Tyrosine | 215.65 ± 0.98 | 245.23 ± 0.83 | 236.32 ± 0.73 | 242.32 ± 0.17 |
| Phenylalanine | 216.32 ± 0.86 | 231.65 ± 0.47 | 198.96 ± 0.71 | 200.65 ± 0.62 |
| Lysine | 154.65 ± 0.56 | 188.32 ± 0.82 | 159.65 ± 0.26 | 184.32 ± 0.25 |
| Histidine | 90.65 ± 0.12 | 103.65 ± 0.43 | 99.46 ± 0.29 | 100.65 ± 0.74 |
| Arginine | 399.98 ± 0.76 | 395.32 ± 0.23 | 397.32 ± 0.66 | 421.32 ± 0.76 |
| Proline | 245.32 ± 0.53 | 288.32 ± 0.52 | 245.65 ± 0.73 | 253.32 ± 0.45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, T.; Jiao, Z.; Hu, J.; Lou, H.; Chen, Q. Chinese Yellow Rice Wine Processing with Reduced Ethyl Carbamate Formation by Deleting Transcriptional Regulator Dal80p in Saccharomyces cerevisiae. Molecules 2020, 25, 3580. https://doi.org/10.3390/molecules25163580
Wei T, Jiao Z, Hu J, Lou H, Chen Q. Chinese Yellow Rice Wine Processing with Reduced Ethyl Carbamate Formation by Deleting Transcriptional Regulator Dal80p in Saccharomyces cerevisiae. Molecules. 2020; 25(16):3580. https://doi.org/10.3390/molecules25163580
Chicago/Turabian StyleWei, Tianyu, Zhihua Jiao, Jingjin Hu, Hanghang Lou, and Qihe Chen. 2020. "Chinese Yellow Rice Wine Processing with Reduced Ethyl Carbamate Formation by Deleting Transcriptional Regulator Dal80p in Saccharomyces cerevisiae" Molecules 25, no. 16: 3580. https://doi.org/10.3390/molecules25163580
APA StyleWei, T., Jiao, Z., Hu, J., Lou, H., & Chen, Q. (2020). Chinese Yellow Rice Wine Processing with Reduced Ethyl Carbamate Formation by Deleting Transcriptional Regulator Dal80p in Saccharomyces cerevisiae. Molecules, 25(16), 3580. https://doi.org/10.3390/molecules25163580