
Mosquitocidal and Anti-Inflammatory Properties of The Essential Oils Obtained from Monoecious, Male, and Female Inflorescences of Hemp (Cannabis sativa L.) and Their Encapsulation in Nanoemulsions


 ,
,  ,
, 
 ,
, 
 and
and 
Abstract
1. Introduction
2. Results
2.1. Chemical Composition of C. sativa Essential Oils
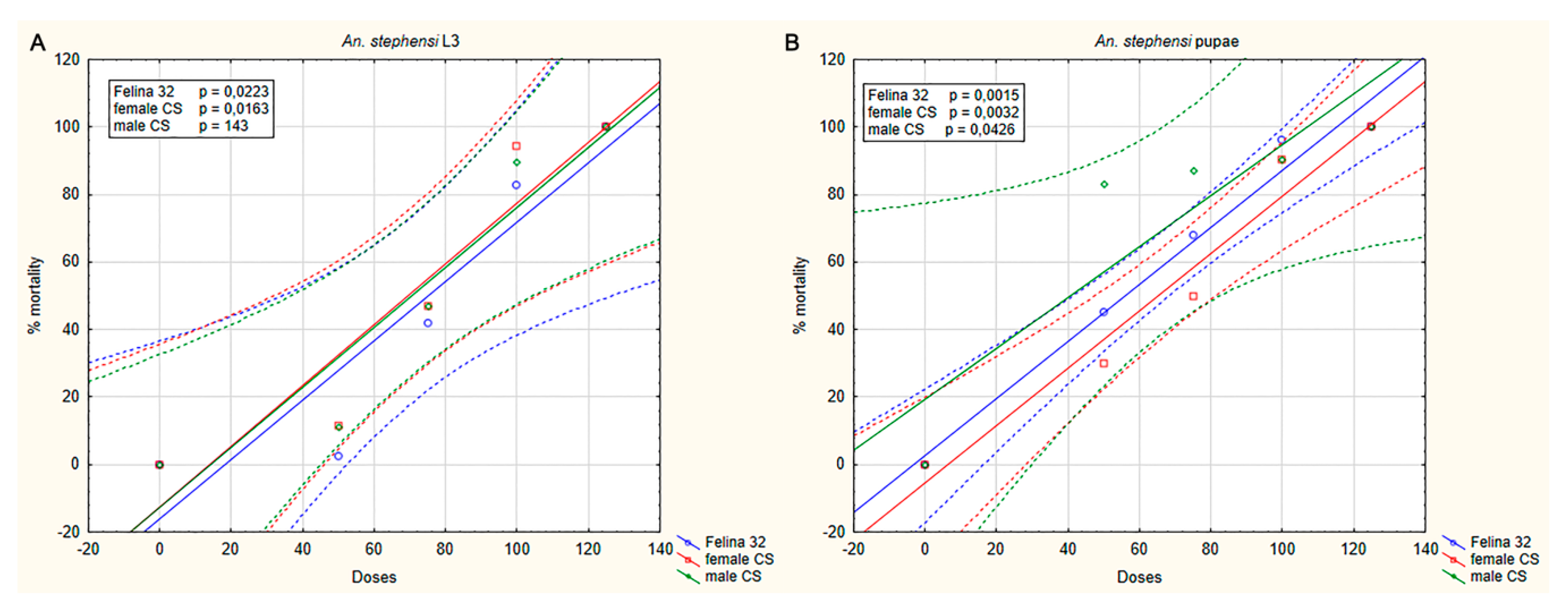
2.2. Toxicity of C. sativa Essential Oils on Mosquito Larvae and Pupae
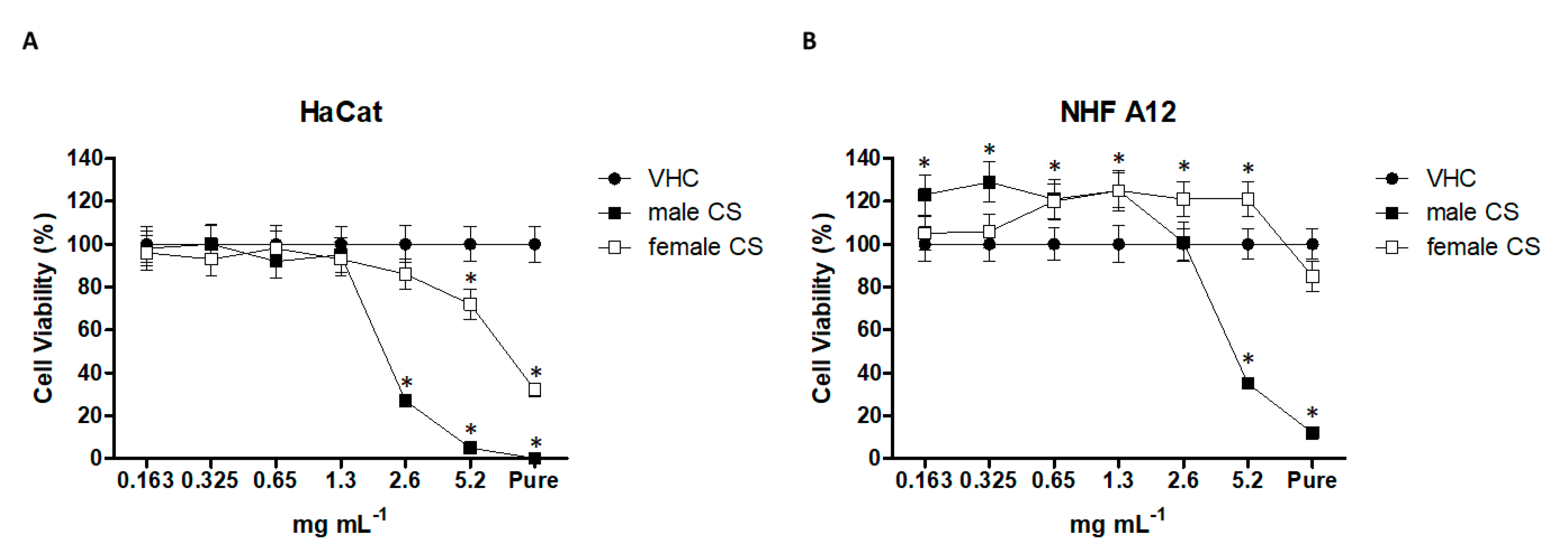
2.3. Effect of Hemp Essential Oils in HaCaT and NHF A12 Cell Lines
2.3.1. Essential Oil Effects on Cell Damage.
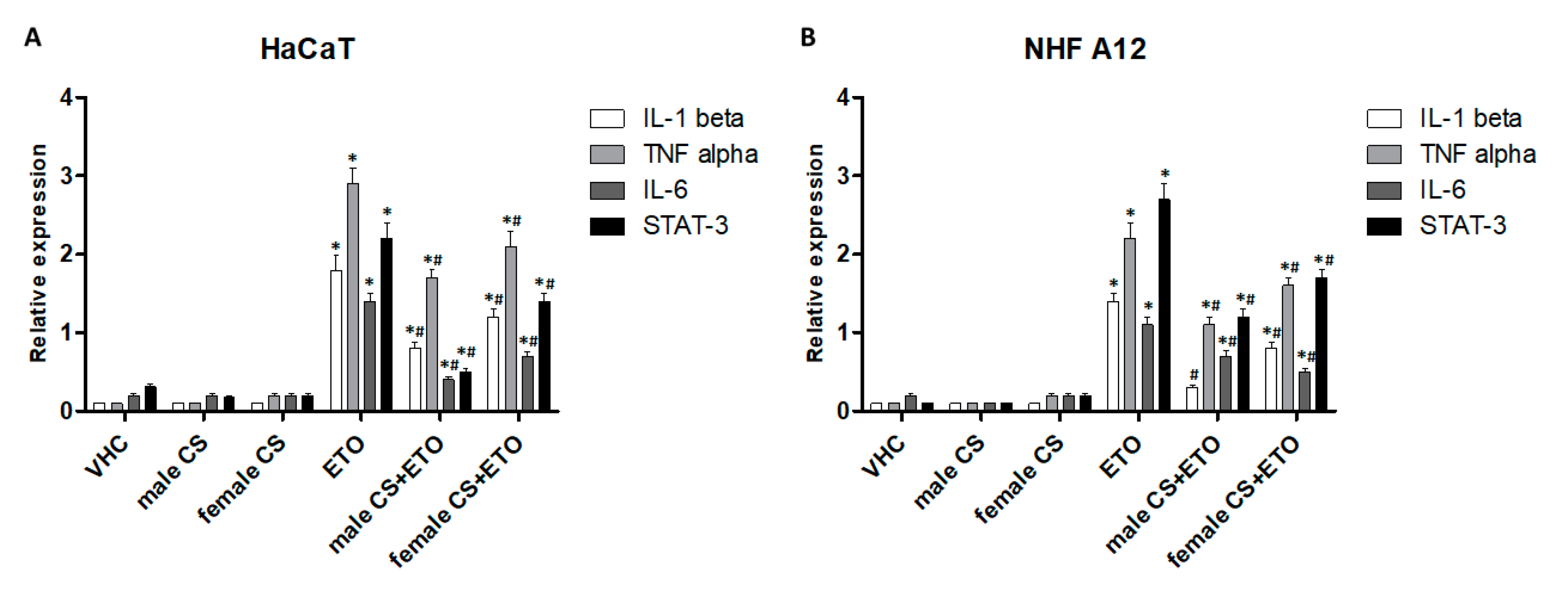
2.3.2. Hemp Essential Oils Reduced Proinflammatory Gene Expression in ETO-treated HaCaT and NHF A12 Cell Lines.
2.3.3. Hemp Essential Oils Reduced Proinflammatory Protein Release in ETO-treated HACAT and NHF A12 Cell Lines
2.4. Preparation and Characterization of C. sativa Essential Oil Nanoemulsions
2.5. Stability of C. sativa Essential Oil Nanoemulsions
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Reagents
4.3. Mosquitoes
4.4. Cell Lines
4.5. Steam Distillation
4.6. Hydrodistillation
4.7. GC-MS Analysis
4.8. Insecticidal Assays
4.9. MTT Assay
4.10. Gene Expression Analysis by TaqMan Array
4.11. ELISA Assay
4.12. Alkaline Comet Assay
4.13. Statistical Analysis
4.14. Preparation of EO-Based Nanoemulsions
4.15. Nanoemulsions Characterization
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fiorini, D.; Molle, A.; Nabissi, M.; Santini, G.; Benelli, G.; Maggi, F. Valorizing industrial hemp (Cannabis sativa L.) by-products: Cannabidiol enrichment in the inflorescence essential oil optimizing sample pre-treatment prior to distillation. Ind. Crops Prod. 2019, 128, 581–589. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R.; Lupidi, G.; Nabissi, M.; Petrelli, R.; Kamte, S.L.N.; Cappellacci, L.; Fiorini, D.; Sut, S.; Dall’Acqua, S.; et al. The crop-residue of fiber hemp cv. Futura 75: From a waste product to a source of botanical insecticides. Env. Sci. Pollut. Res. 2018, 25, 10515–10525. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R.; Petrelli, R.; Cappellacci, L.; Santini, G.; Fiorini, D.; Sut, S.; Dall’Acqua, S.; Canale, A.; Maggi, F. The essential oil from industrial hemp (Cannabis sativa L.) by-products as an effective tool for insect pest management in organic crops. Ind. Crops Prod. 2018, 122, 308–315. [Google Scholar] [CrossRef]
- Petrelli, R.; Ranjbarian, F.; Dall’Acqua, S.; Papa, F.; Iannarelli, R.; Kamte, S.L.N.; Vittori, S.; Benelli, G.; Maggi, F.; Hofer, A.; et al. An overlooked horticultural crop, Smyrnium olusatrum, as a potential source of compounds effective against African trypanosomiasis. Parasitol. Int. 2017, 66, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Kamte, S.L.N.; Ranjbarian, F.; Cianfaglione, K.; Sut, S.; Dall’Acqua, S.; Bruno, M.; Afshar, F.H.; Iannarelli, R.; Benelli, G.; Cappellacci, L.; et al. Identification of highly effective antitrypanosomal compounds in essential oils from the Apiaceae family. Ecotoxicol. Environ. Saf. 2018, 156, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Pavela, R.; Iannarelli, R.; Petrelli, R.; Cappellacci, L.; Cianfaglione, K.; Afshar, F.H.; Nicoletti, M.; Canale, A.; Maggi, F. Synergized mixtures of Apiaceae essential oils and related plant-borne compounds: Larvicidal effectiveness on the filariasis vector Culex quinquefasciatus Say. Ind. Crops Prod. 2017, 96, 186–195. [Google Scholar] [CrossRef]
- Vitali, L.A.; Beghelli, D.; Nya, P.C.B.; Bistoni, O.; Cappellacci, L.; Damiano, S.; Lupidi, G.; Maggi, F.; Orsomando, G.; Papa, F.; et al. Diverse biological effects of the essential oil from Iranian Trachyspermum ammi. Arab. J. Chem. 2016, 9, 775–786. [Google Scholar] [CrossRef]
- Woguem, V.; Fogang, H.P.; Maggi, F.; Tapondjou, L.A.; Womeni, H.M.; Quassinti, L.; Bramucci, M.; Vitali, L.A.; Petrelli, D.; Lupidi, G.; et al. Volatile oil from striped African pepper (Xylopia parviflora, Annonaceae) possesses notable chemopreventive, anti-inflammatory and antimicrobial potential. Food Chem. 2014, 149, 183–189. [Google Scholar] [CrossRef]
- World Health Organization. World Malaria Report 2018; World Health Organization: New York, NY, USA, 2018; pp. 1–210. [Google Scholar]
- Miller, L.H.; Baruch, D.I.; Marsh, K.; Doumbo, O.K. The pathogenic basis of malaria. Nature 2002, 415, 673–679. [Google Scholar] [CrossRef]
- Hanafi-Bojd, A.A.; Vatandoost, H.; Oshaghi, M.A.; Haghdoost, A.A.; Shahi, M.; Sedaghat, M.M.; Abedi, F.; Yeryan, M.; Pakari, A. Entomological and epidemiological attributes for malaria transmission and implementation of vector control in southern Iran. Acta Trop. 2012, 121, 85–92. [Google Scholar] [CrossRef]
- Manimaran, A.; Cruz, M.M.J.J.; Muthu, C.; Vincent, S.; Ignacimuthu, S. Larvicidal and knockdown effects of some essential oils against Culex quinquefasciatus Say, Aedes aegypti (L.) and Anopheles stephensi (Liston). Adv. Biosci. Biotechnol. 2012, 3, 855–862. [Google Scholar] [CrossRef][Green Version]
- Chandre, F.; Darriet, F.; Manguin, S.; Brengues, C.; Carnevale, P.; Guillet, P. Pyrethroid cross resistance spectrum among populations of Anopheles gambiae s.s. from Cote d’Ivoire. J. Am. Mosq. Contr. 1999, 15, 53–59. [Google Scholar]
- Sedaghat, M.M.; Dehkordi, A.S.; Khanavi, M.; Abai, M.R.; Mohtarami, F.; Vatandoost, H. Chemical composition and larvicidal activity of essential oil of Cupressus arizonica E.L. Greene against malaria vector Anopheles stephensi Liston (Diptera: Culicidae). Pharmacogn. Res. 2011, 3, 135–139. [Google Scholar]
- Asase, A.; Oteng-Yeboah, A.A.; Odamtten, G.T.; Simmonds, M.S.J. Ethnobotanical Study of Some Ghanaian Anti-Malarial Plants. J. Ethnopharmacol. 2005, 99, 273–279. [Google Scholar] [CrossRef]
- Pavela, R.; Maggi, F.; Iannarelli, R.; Benelli, G. Plant extracts for developing mosquito larvicides: From laboratory to the field, with insights on the modes of action. Acta Trop. 2019, 193, 236–271. [Google Scholar] [CrossRef]
- Wongsrichanalai, C.; Pickard, A.L.; Wernsdorfer, W.H.; Meshnick, S.R. Epidemiology of drug-resistant malaria. Lancet. Infect. Dis. 2002, 2, 209–218. [Google Scholar] [CrossRef]
- Potter, D.J. The Propagation, Characterisation and Optimisation of Cannabis sativa L. as a Phytopharmaceutical. Ph.D. Thesis, King’s College, London, UK, March 2009. [Google Scholar]
- Fiorini, D.; Scortichini, S.; Bonacucina, G.; Greco, N.G.; Mazzara, E.; Petrelli, R.; Torresi, J.; Maggi, F.; Cespi, M. Cannabidiol-enriched hemp essential oil obtained by an optimized microwave-assisted extraction using a central composite design. Ind. Crops Prod. 2020, 154, 112688. [Google Scholar] [CrossRef]
- Tabari, M.A.; Khodashenas, A.; Jafari, M.; Petrelli, R.; Cappellacci, L.; Nabissi, M.; Maggi, F.; Pavela, R.; Youssefi, M.R. Acaricidal properties of hemp (Cannabis sativa L.) essential oil against Dermanyssus gallinae and Hyalomma dromedarii. Ind. Crops Prod. 2020, 147, 112238. [Google Scholar] [CrossRef]
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef]
- Hänel, K.H.; Cornelissen, C.; Lüscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care (New Rochelle) 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, C.; Kurosaka, K.; Watanabe, N.; Kobayashi, Y. Cytokine production by macrophages in association with phagocytosis of etoposide-treated P388 cells in vitro and in vivo. Biophys. Acta 2001, 1541, 221–230. [Google Scholar] [CrossRef]
- Darst, M.; Al-Hassani, M.; Li, T.; Yi, Q.; Travers, J.M.; Lewis, D.A.; Travers, J.B. Augmentation of chemotherapy-induced cytokine production by expression of the platelet-activating factor receptor in a human epithelial carcinoma cell line. J. Immunol. 2004, 172, 6330–6335. [Google Scholar] [CrossRef] [PubMed]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Bertoli, A.; Ruffoni, B.; Pistelli, L.; Pistelli, L. Analytical methods for the extraction and identification of secondary metabolite production in ‘in vitro’ plant cell cultures. Adv. Exp. Med. Biol. 2010, 698, 250–266. [Google Scholar] [PubMed]
- Nagy, D.U.; Cianfaglione, K.; Maggi, F.; Sut, S.; Dall’Acqua, S. Chemical characterization of leaves, male and female flowers from spontaneous Cannabis (Cannabis sativa L.) growing in Hungary. Chem. Biodivers. 2019, 16, e1800562. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Franz, C. Composition of the essential oils and extracts of two populations of Cannabis sativa L. ssp. spontanea from Austria. J. Essent. Oil Res. 2003, 15, 158–160. [Google Scholar] [CrossRef]
- Kendall, D.A.; Yudowski, G.A. Cannabinoid receptors in the central nervous system: Their signaling and roles in disease. Front. Cell. Neurosci. 2017, 10, 294. [Google Scholar] [CrossRef]
- Subramaniam, J.; Kovendan, K.; Mahesh Kumar, P.; Murugan, K.; Walton, W. Mosquito larvicidal activity of Aloe vera (Family: Liliaceae) leaf extract and Bacillus sphaericus, against Chikungunya vector, Aedes aegypti. Saudi J. Biol. Sci. 2012, 19, 503–509. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R. Beyond mosquitoes—Essential oil toxicity and repellency against bloodsucking insects. Ind. Crops Prod. 2018, 117, 382–392. [Google Scholar] [CrossRef]
- Alkofahi, A.; Rupprecht, J.K.; Anderson, J.E.; McLaughlin, J.L.; Mikolajczak, K.L.; Scott, B.A. Search for new pesticides from higher plants. In Insecticides of Plant Origin; Arnason, J.T., Philogene, B.J.R., Morand, P., Eds.; American Chemical Society: Washington, DC, USA, 1989; pp. 25–43. [Google Scholar]
- Benelli, G.; Maggi, F.; Pavela, R.; Murugan, K.; Govindarajan, M.; Vaseeharan, B.; Petrelli, R.; Cappellacci, L.; Kumar, S.; Hofer, A.; et al. Mosquito control with green nanopesticides: Towards the One Health approach? A review of non-target effects. Environ. Sci. Pollut. Res. Int. 2018, 25, 10184–10206. [Google Scholar] [CrossRef] [PubMed]
- Kovendan, K.; Murugan, K. Effect of medicinal plants on the mosquito vectors from the different agroclimatic regions of Tamil Nadu, India. Adv. Environ. Biol. 2011, 5, 335–344. [Google Scholar]
- Benelli, G.; Pavel, R.; Zorzetto, C.; Sánchez-Mateo, C.C.; Santini, G.; Canale, A.; Maggi, F. Insecticidal activity of the essential oil from Schizogyne sericea (Asteraceae) on four insect pests and two non-target species. Entomol. Gen. 2019, 39, 9–18. [Google Scholar] [CrossRef]
- Cheng, S.S.; Huang, C.G.; Chen, Y.J.; Yu, J.J.; Chen, W.J.; Chang, S.T. Chemical compositions and larvicidal activities of leaf essential oils from two eucalyptus species. Bioresour. Technol. 2009, 100, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhu, F.; Zhou, X.M.; Niu, C.Y.; Lei, C.I. Repellent and fumigant activity of essential oil from Artemisia vulgaris to Tribolium eastaneum (Herbst). J. Stored Prod. Res. 2006, 42, 339–347. [Google Scholar] [CrossRef]
- Pavela, R.; Murugan, K.; Canale, A.; Benelli, G. Saponaria officinalis-synthesized silver nanocrystals as effective biopesticides and oviposition inhibitors against Tetranychus urticae Koch. Ind. Crops Prod. 2017, 97, 338–344. [Google Scholar] [CrossRef]
- Thomas, T.G.; Sharma, S.K.; Prakash, A.; Sharma, B.R. Insecticidal properties of essential oil of Cannabis sativa Linn. against mosquito larvae. Entomon 2000, 25, 21–24. [Google Scholar]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R. Cannabis sativa and Humulus lupulus essential oils as novel control tools against the invasive mosquito Aedes albopictus and fresh water snail Physella acuta. Ind. Crops Prod. 2016, 85, 318–323. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crops Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Khanavi, M.; Fallah, A.; Vatandoost, H.; Sedaghat, M.; Abai, M.R.; Hadjiakhoondi, A. Larvicidal activity of essential oil and methanol extract of Nepeta menthoides against malaria vector Anopheles stephensi. Asian Pac. J. Trop. Med. 2012, 5, 962–965. [Google Scholar]
- Da Silva, R.C.S.; Milet-Pinheiro, P.; Bezerra da Silva, P.C.; da Silva, A.G.; da Silva, M.V.; Navarro, D.M.D.A.F.; da Silva, N.H. (E)-caryophyllene and α-humulene: Aedes aegypti oviposition deterrents elucidated by gas chromatography-electrophysiological assay of Commiphora leptophloeos leaf oil. PLoS ONE 2015, 10, e0144586. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Fumagalli, M.; Pacchetti, B.; Piazza, S.; Magnavacca, A.; Khalilpour, S.; Melzi, G.; Martinelli, G.; Dell’Agli, M. Cannabis sativa L. extract and cannabidiol inhibit in vitro mediators of skin inflammation and wound injury. Phytother. Res. 2019, 33, 2083–2093. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowski, L.F.; Calixto, J.B. Anti-inflammatory effects of compounds alpha-humulene and (−)-trans-caryophyllene isolated from the essential oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef]
- Pavoni, L.; Benelli, G.; Maggi, F.; Bonacucina, G. Green nanoemulsion interventions for biopesticide formulations. In Nano-Biopesticides Today and Future Perspectives; Academic Press: Cambridge, MA, USA, 2019; pp. 133–160. [Google Scholar]
- Pavoni, L.; Pavela, R.; Cespi, M.; Bonacucina, G.; Maggi, F.; Zeni, V.; Canale, A.; Lucchi, A.; Bruschi, F.; Benelli, G. Green Micro-and Nanoemulsions for Managing Parasites, Vectors and Pests. Nanomaterials 2019, 9, 1285. [Google Scholar] [CrossRef]
- Quassinti, L.; Bramucci, M.; Lupidi, G.; Barboni, L.; Ricciutelli, M.; Sagratini, G.; Papa, F.; Caprioli, G.; Petrelli, D.; Vitali, L.A.; et al. In vitro biological activity of essential oils and isolated furanosesquiterpenes from the neglected vegetable Smyrnium olusatrum L. (Apiaceae). Food Chem. 2013, 138, 808–813. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organization: New York, NY, USA, 2005; pp. 5–39. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Component a | RI Calc. b | RI Lit. c | % Felina 32 | Carmagnola CS | |
|---|---|---|---|---|---|---|
| % Female | % Male | |||||
| 1 | Tricyclene | 914 | 921 | 0.2 | ||
| 2 | α-Thujene | 921 | 924 | 0.1 | 0.1 | |
| 3 | α-Pinene | 926 | 932 | 15.1 | 11.4 | 8.0 |
| 4 | Camphene | 939 | 946 | 0.2 | 0.2 | |
| 5 | Sabinene | 966 | 967 | 0.2 | ||
| 6 | β-Pinene | 968 | 974 | 5.0 | 5.7 | 3.1 |
| 7 | Myrcene | 989 | 988 | 11.8 | 24.3 | 10.6 |
| 8 | α-Phellandrene | 1003 | 1002 | 0.4 | ||
| 9 | δ-3-Carene | 1008 | 1008 | 0.4 | ||
| 10 | α-Terpinene | 1014 | 1014 | 0.1 | 0.3 | |
| 11 | Limonene | 1025 | 1024 | 6.0 | 6.7 | 4.1 |
| 12 | (1,8)-Cineole | 1027 | 1026 | 3.5 | 1.4 | 1.6 |
| 13 | (Z)-β-Ocimene | 1037 | 1032 | 0.1 | 0.2 | |
| 14 | (E)-β-Ocimene | 1047 | 1044 | 0.8 | 2.3 | 0.3 |
| 15 | γ-Terpinene | 1056 | 1054 | 0.3 | 0.4 | 0.2 |
| 16 | cis-Sabinene Hydrate | 1064 | 1065 | 0.2 | ||
| 17 | Terpinolene | 1085 | 1086 | 0.9 | 13.5 | 0.3 |
| 18 | Linalool | 1101 | 1095 | 0.8 | ||
| 19 | (Z)-Caryophyllene | 1407 | 1408 | 0.3 | 0.6 | |
| 20 | (E)-Caryophyllene | 1409 | 1417 | 34.8 | 19.3 | 47.2 |
| 21 | α-trans-Bergamotene | 1430 | 1432 | 1.0 | 0.5 | |
| 22 | α-Humulene | 1443 | 1452 | 11.4 | 6.4 | 15.1 |
| 23 | allo-Aromadendrene | 1450 | 1458 | 0.6 | 0.8 | |
| 24 | (E)-β-Farnesene | 1456 | 1454 | 0.9 | ||
| 25 | β-Selinene | 1476 | 1519 | 1.5 | 0.4 | 1.7 |
| 26 | α-Selinene | 1485 | 1498 | 1.0 | 0.3 | 1.2 |
| 27 | δ-Cadinene | 1519 | 1522 | 0.1 | ||
| 28 | Selina-3,7(11)-diene | 1531 | 1545 | 0.5 | 0.6 | |
| 29 | Caryophyllene oxide | 1571 | 1582 | 2.1 | 2.2 | 3.6 |
| 30 | Humulene epoxide II | 1593 | 1608 | 0.4 | 0.5 | 0.5 |
| 31 | Cannabidiol | 2419 | 2430 | 0.1 | 0.2 | |
| Total identified (%) | 99.0 | 97.6 | 100 | |||
| monoterpene hydrocarbons | 40.6 | 66.1 | 26.6 | |||
| oxygenated monoterpenes | 3.7 | 2.2 | 1.6 | |||
| sesquiterpene hydrocarbons | 52.1 | 26.4 | 67.7 | |||
| oxygenated sesquiterpenes | 2.5 | 2.7 | 4.1 | |||
| cannabinoids | 0.1 | 0.2 | ||||
| Mosquito Species | LC50 a | |||||
|---|---|---|---|---|---|---|
| Larvae | Pupae | |||||
| Felina 32 b | Female CS b | Male CS b | Felina 32 b | Female CS b | Male CS b | |
| An. stephensi | 78.8 | 75.12 | 75.23 | 54.41 | 67.19 | 20.13 |
| An. gambiae | 73.5 | 75.54 | 75.04 | 50.06 | 41.51 | 50.27 |
| Sample | Essential Oil | Surfactant | Other Components | Description |
|---|---|---|---|---|
| A8 | C. sativa cv CS 6% (male inflorescences) | Tween 80 4% | -Polydispersed system at t0 -Instability phenomena after 1 month | |
| A9 | C. sativa cv CS 6% (female inflorescences) | Tween 80 4% | -Polydispersed system at t0 -Instability phenomena after 1 month | |
| A8B | C. sativa cv CS 4% (male inflorescences) | Tween 80 4% | -Polydispersed system at t0 -Instability phenomena between 1 and 2 months | |
| A9B | C. sativa cv CS 4% (female inflorescences) | Tween 80 4% | -Polydispersed system at t0 -Instability phenomena between 1 and 2 months | |
| A10 | C. sativa cv CS 3% (female inflorescences) | Tween 80 4% | Ethyl oleate 3% | -Mean diameter ≈ 200 nm PDI < 0.2 -Stable system after 6 months |
| A21 | C. sativa cv CS 3% (male inflorescences) | Tween 80 4% | Ethyl oleate 3% | -Mean diameter ≈ 200 nm PDI < 0.2 -Stable system after 6 months |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, P.; Cappelli, A.; Marinelli, O.; Valzano, M.; Pavoni, L.; Bonacucina, G.; Petrelli, R.; Pompei, P.; Mazzara, E.; Ricci, I.; et al. Mosquitocidal and Anti-Inflammatory Properties of The Essential Oils Obtained from Monoecious, Male, and Female Inflorescences of Hemp (Cannabis sativa L.) and Their Encapsulation in Nanoemulsions. Molecules 2020, 25, 3451. https://doi.org/10.3390/molecules25153451
Rossi P, Cappelli A, Marinelli O, Valzano M, Pavoni L, Bonacucina G, Petrelli R, Pompei P, Mazzara E, Ricci I, et al. Mosquitocidal and Anti-Inflammatory Properties of The Essential Oils Obtained from Monoecious, Male, and Female Inflorescences of Hemp (Cannabis sativa L.) and Their Encapsulation in Nanoemulsions. Molecules. 2020; 25(15):3451. https://doi.org/10.3390/molecules25153451
Chicago/Turabian StyleRossi, Paolo, Alessia Cappelli, Oliviero Marinelli, Matteo Valzano, Lucia Pavoni, Giulia Bonacucina, Riccardo Petrelli, Pierluigi Pompei, Eugenia Mazzara, Irene Ricci, and et al. 2020. "Mosquitocidal and Anti-Inflammatory Properties of The Essential Oils Obtained from Monoecious, Male, and Female Inflorescences of Hemp (Cannabis sativa L.) and Their Encapsulation in Nanoemulsions" Molecules 25, no. 15: 3451. https://doi.org/10.3390/molecules25153451
APA StyleRossi, P., Cappelli, A., Marinelli, O., Valzano, M., Pavoni, L., Bonacucina, G., Petrelli, R., Pompei, P., Mazzara, E., Ricci, I., Maggi, F., & Nabissi, M. (2020). Mosquitocidal and Anti-Inflammatory Properties of The Essential Oils Obtained from Monoecious, Male, and Female Inflorescences of Hemp (Cannabis sativa L.) and Their Encapsulation in Nanoemulsions. Molecules, 25(15), 3451. https://doi.org/10.3390/molecules25153451