
The Inhibitory Activity of Anthraquinones against Pathogenic Protozoa, Bacteria, and Fungi and the Relationship to Structure

 , ,
, ,  ,
, 

Abstract
1. Introduction
2. Results and Discussion
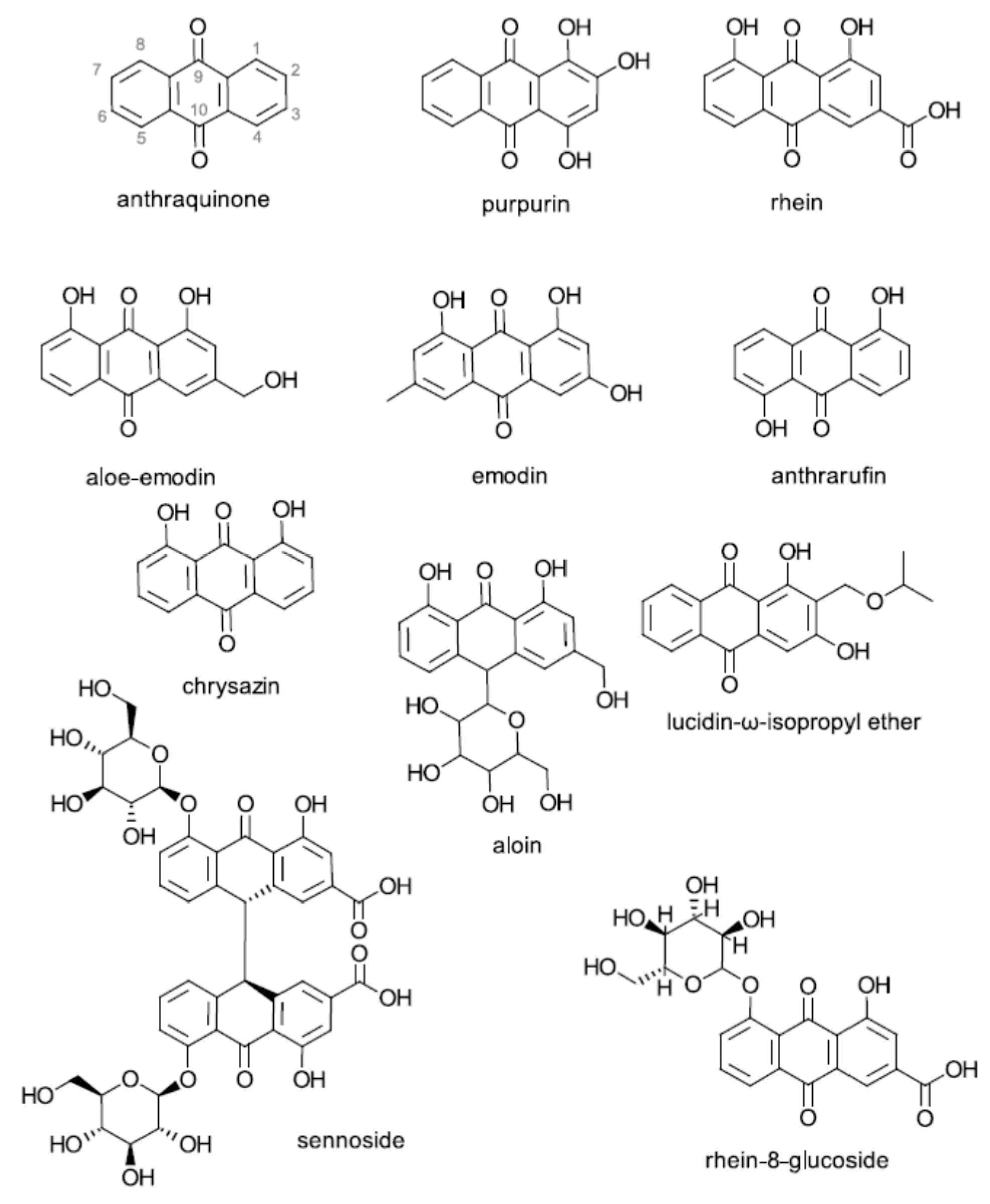
2.1. Structure–Activity Relationships
2.2. Antifungal Activity of Emodin
2.3. Biological Activities of Purpurin that Impact Human Health
2.4. Anti-Oxidative Properties of Anthraquinones
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Stock Solutions of the Anthraquinone Powders
3.2.2. Trichomonad Growth Inhibition Assays
3.2.3. Bacterial and Fungal Screens with Anthraquinone Compounds
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xu, J.S.; Cui, Y.; Liao, X.M.; Tan, X.B.; Cao, X. Effect of emodin on the cariogenic properties of Streptococcus mutans and the development of caries in rats. Exp. Ther. Med. 2014, 8, 1308–1312. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Niu, M.; Zhang, W.; Yan, J.; Li, J.; Tan, X.; Li, B.; Su, M.; Di, B.; Yan, F. Emodin reverses leukemia multidrug resistance by competitive inhibition and downregulation of P-glycoprotein. PLoS ONE 2017, 12, e0187971. [Google Scholar] [CrossRef]
- Dai, H.; Chen, Z.; Shang, B.; Chen, Q. Identification and quantification of four anthraquinones in rhubarb and its preparations by gas chromatography-mass spectrometry. J. Chromatogr. Sci. 2018, 56, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Chien, S.C.; Wu, Y.C.; Chen, Z.W.; Yang, W.C. Naturally occurring anthraquinones: Chemistry and therapeutic potential in autoimmune diabetes. Evid.-Based Complement. Altern. Med.: Ecam 2015, 2015, 357357. [Google Scholar] [CrossRef] [PubMed]
- Cáceres-Castillo, D.; Pérez-Navarro, Y.; Torres-Romero, J.C.; Mirón-López, G.; Ceballos-Cruz, J.; Arana-Argáez, V.; Vázquez-Carrillo, L.; Fernández-Sánchez, J.M.; Alvarez-Sánchez, M.E. Trichomonicidal activity of a new anthraquinone isolated from the roots of Morinda panamensis Seem. Drug Dev. Res. 2019, 80, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Meier, N.; Meier, B.; Peter, S.; Wolfram, E. In-silico UHPLC method optimization for aglycones in the herbal laxatives Aloe barbadensis Mill., Cassia angustifolia vahl pods, Rhamnus frangula L. Bark, Rhamnus purshianus DC. bark, and Rheum palmatum L. roots. Molecules 2017, 22, 1838. [Google Scholar] [CrossRef]
- Jafer, M.; Patil, S.; Hosmani, J.; Bhandi, S.H.; Chalisserry, E.P.; Anil, S. Chemical plaque control strategies in the prevention of biofilm-associated oral diseases. J. Contemp. Dent. Pract. 2016, 17, 337–343. [Google Scholar]
- Ji, X.; Liu, X.; Peng, Y.; Zhan, R.; Xu, H.; Ge, X. Comparative analysis of methicillin-sensitive and resistant Staphylococcus aureus exposed to emodin based on proteomic profiling. Biochem. Biophys. Res. Commun. 2017, 494, 318–324. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, J.G. Health functions and structure-activity relationships of natural anthraquinones from plants. Food Funct. 2018, 9, 6063–6080. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, C.; He, R.; Zhou, M.; Liu, Y.; Guo, X.; Wang, M.; Zhu, F.; Qin, R.; Li, X. Danthron suppresses autophagy and sensitizes pancreatic cancer cells to doxorubicin. Toxicol. Vitr.: Int. J. Publ. Assoc. Bibra 2019, 54, 345–353. [Google Scholar] [CrossRef]
- Yang, L.; Lin, S.; Kang, Y.; Xiang, Y.; Xu, L.; Li, J.; Dai, X.; Liang, G.; Huang, X.; Zhao, C. Rhein sensitizes human pancreatic cancer cells to EGFR inhibitors by inhibiting STAT3 pathway. J. Exp. Clin. Cancer Res.: Cr 2019, 38, 31. [Google Scholar] [CrossRef]
- Janeczko, M.; Maslyk, M.; Kubinski, K.; Golczyk, H. Emodin, a natural inhibitor of protein kinase CK2, suppresses growth, hyphal development, and biofilm formation of Candida albicans. Yeast (Chichesterengland) 2017, 34, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Nam, W.; Kim, S.P.; Nam, S.-H.; Friedman, M. Structure-antioxidative and anti-inflammatory activity relationships of purpurin and related anthraquinones in chemical and cell assays. Molecules 2017, 22, 265. [Google Scholar] [CrossRef] [PubMed]
- Nam, W.; Nam, S.H.; Kim, S.P.; Levin, C.; Friedman, M. Anti-adipogenic and anti-obesity activities of purpurin in 3T3-L1 preadipocyte cells and in mice on a high-fat diet. BMC Complement. Altern. Med. 2019, 19, 364. [Google Scholar] [CrossRef] [PubMed]
- Marković, Z.; Jeremić, S.; Dimitrić Marković, J.; Stanojević Pirković, M.; Amić, D. Influence of structural characteristics of substituents on the antioxidant activity of some anthraquinone derivatives. Comput. Theor. Chem. 2016, 1077, 25–31. [Google Scholar] [CrossRef]
- Wang, H.-H. Antitrichomonal action of emodin in mice. J. Ethnopharmacol. 1993, 40, 111–116. [Google Scholar]
- Dalimi, A.; Delavari, M.; Ghaffarifar, F.; Sadraei, J. In vitro and in vivo antileishmanial effects of aloe-emodin on Leishmania major. J. Tradit. Complement. Med. 2015, 5, 96–99. [Google Scholar] [CrossRef]
- Kumar, S.; Yadav, M.; Yadav, A.; Rohilla, P.; Yadav, J.P. Antiplasmodial potential and quantification of aloin and aloe-emodin in Aloe vera collected from different climatic regions of India. BMC Complement. Altern. Med. 2017, 17, 369. [Google Scholar] [CrossRef]
- Zhong, T.; Zhang, L.Y.; Wang, Z.Y.; Wang, Y.; Song, F.M.; Zhang, Y.H.; Yu, J.H. Rheum emodin inhibits enterovirus 71 viral replication and affects the host cell cycle environment. Acta Pharmacol. Sin. 2017, 38, 392–401. [Google Scholar] [CrossRef]
- Bai, Y.; Su, Z.; Sun, H.; Zhao, W.; Chen, X.; Hang, P.; Zhu, W.; Du, Z. Aloe-emodin relieves high-fat diet induced QT prolongation via MiR-1 inhibition and IK1 up-regulation in rats. Cell. Physiol. Biochem.: Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 1961–1973. [Google Scholar] [CrossRef]
- Bu, T.; Wang, C.; Meng, Q.; Huo, X.; Sun, H.; Sun, P.; Zheng, S.; Ma, X.; Liu, Z.; Liu, K. Hepatoprotective effect of rhein against methotrexate-induced liver toxicity. Eur. J. Pharmacol. 2018, 834, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chu, S.; Liu, Y.; Chen, N. Neuroprotective effects of anthraquinones from rhubarb in central nervous system diseases. Evid.-Based Complement. Altern. Med.: Ecam 2019, 2019, 3790728. [Google Scholar] [CrossRef] [PubMed]
- Prasad, C.; Nurko, S.; Borovoy, J.; Korson, M.S. The importance of gut motility in the metabolic control of propionic acidemia. J. Pediatrics 2004, 144, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.F.; Liu, C.F.; Lai, W.F.; Xiang, Q.; Li, Z.F.; Wang, H.; Lin, N. The laxative effect of emodin is attributable to increased aquaporin 3 expression in the colon of mice and HT-29 cells. Fitoterapia 2014, 96, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Stemmer, S.M.; Mordechai, E.; Adelson, M.E.; Gygax, S.E.; Hilbert, D.W. Trichomonas vaginalis is most frequently detected in women at the age of peri-/premenopause: An unusual pattern for a sexually transmitted pathogen. Am. J. Obstet. Gynecol. 2018, 218, 328.e1–328.e13. [Google Scholar] [CrossRef]
- Okafor, C.C.; Strickland, L.G.; Jones, B.M.; Kania, S.; Anderson, D.E.; Whitlock, B.K. Prevalence of Tritrichomonas foetus in Tennessee bulls. Vet. Parasitol. 2017, 243, 169–175. [Google Scholar] [CrossRef]
- Michi, A.N.; Favetto, P.H.; Kastelic, J.; Cobo, E.R. A review of sexually transmitted bovine trichomoniasis and campylobacteriosis affecting cattle reproductive health. Theriogenology 2016, 85, 781–791. [Google Scholar] [CrossRef]
- Gookin, J.L.; Hanrahan, K.; Levy, M.G. The conundrum of feline Trichomonosis. J. Feline Med. Surg. 2017, 19, 261–274. [Google Scholar] [CrossRef]
- Paul, A.; Stayt, J. The intestinal microbiome in dogs and cats with diarrhoea as detected by a faecal polymerase chain reaction-based panel in Perth, Western Australia. Aust. Vet. J. 2019, 97, 418–421. [Google Scholar] [CrossRef]
- Arranz-Solis, D.; Pedraza-Diaz, S.; Miro, G.; Rojo-Montejo, S.; Hernandez, L.; Ortega-Mora, L.M.; Collantes-Fernandez, E. Tritrichomonas foetus infection in cats with diarrhea from densely housed origins. Vet. Parasitol. 2016, 221, 118–122. [Google Scholar] [CrossRef]
- Bradic, M.; Warring, S.D.; Tooley, G.E.; Scheid, P.; Secor, W.E.; Land, K.M.; Huang, P.-J.; Chen, T.-W.; Lee, C.-C.; Tang, P.; et al. Genetic indicators of drug resistance in the highly repetitive genome of Trichomonas vaginalis. Genome Biol. Evol. 2017, 9, 1658–1672. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, R.W.; Erbelding, E.; Fauci, A.S. Refocusing research on sexually transmitted infections. J. Infect. Dis. 2019, jiz442. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kanetake, S.; Wu, Y.-H.; Tam, C.; Cheng, L.W.; Land, K.M.; Friedman, M. Anti-protozoal effects of the tomato tetrasaccharide glycoalkaloid tomatine and the aglycone tomatidine on mucosal trichomonads. J. Agric. Food Chem. 2016, 64, 8806–8810. [Google Scholar] [CrossRef] [PubMed]
- Noritake, S.M.; Liu, J.; Kanetake, S.; Levin, C.E.; Tam, C.; Cheng, L.W.; Land, K.M.; Friedman, M. Phytochemical-rich foods inhibit the growth of pathogenic trichomonads. BMC Complementary Altern. Med. 2017, 17, 461. [Google Scholar] [CrossRef]
- Friedman, M.; Huang, V.; Quiambao, Q.; Noritake, S.; Liu, J.; Kwon, O.; Chintalapati, S.; Levin, C.E.; Tam, C.; Cheng, L.W.; et al. Potato peels and their bioactive glycoalkaloids and phenolic compounds inhibit the growth of pathogenic trichomonads. J. Agric. Food Chem. 2018, 66, 7942–7947. [Google Scholar] [CrossRef]
- Elkahoui, S.; Bartley, G.E.; Yokoyama, W.H.; Friedman, M. Dietary supplementation of potato peel powders prepared from conventional and organic russet and nonorganic gold and red potatoes reduces weight gain in mice on a high-fat diet. J. Agric. Food Chem. 2018, 66, 6064–6072. [Google Scholar] [CrossRef]
- Sobel, J.D.; Wiesenfeld, H.C.; Martens, M.; Danna, P.; Hooton, T.M.; Rompalo, A.; Sperling, M.; Livengood, C.; Horowitz, B.; Von Thron, J.; et al. Maintenance fluconazole therapy for recurrent vulvovaginal candidiasis. N. Engl. J. Med. 2004, 351, 876–883. [Google Scholar] [CrossRef]
- Beardsley, J.; Halliday, C.L.; Chen, S.C.-A.; Sorrell, T.C. Responding to the emergence of antifungal drug resistance: Perspectives from the bench and the bedside. Future Microbiol. 2018, 13, 1175–1191. [Google Scholar] [CrossRef]
- Leonardelli, F.; Macedo, D.; Dudiuk, C.; Cabeza, M.S.; Gamarra, S.; Garcia-Effron, G. Aspergillus fumigatus intrinsic fluconazole resistance is due to the naturally occurring T301I substitution in Cyp51Ap. Antimicrob Agents Chemother. 2016, 60, 5420–5426. [Google Scholar] [CrossRef]
- Tsang, P.W.; Wong, A.P.; Yang, H.P.; Li, N.F. Purpurin triggers caspase-independent apoptosis in Candida dubliniensis biofilms. PLoS ONE 2013, 8, e86032. [Google Scholar] [CrossRef]
- Viswanathan, G.K.; Shwartz, D.; Losev, Y.; Arad, E.; Shemesh, C.; Pichinuk, E.; Engel, H.; Raveh, A.; Jelinek, R.; Cooper, I.; et al. Purpurin modulates Tau-derived VQIVYK fibrillization and ameliorates Alzheimer’s disease-like symptoms in animal model. Cell. Mol. Life Sci. 2019, 77, 2795–2813. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Hu, P.; Zhang, J.; Cui, W.; Zhao, X. Purpurin exerted antidepressant-like effects on behavior and stress axis reactivity: Evidence of serotonergic engagement. Psychopharmacology 2020, 237, 887–899. [Google Scholar] [CrossRef]
- Park, H.; Shim, J.S.; Kim, B.S.; Jung, H.J.; Huh, T.-L.; Kwon, H.J. Purpurin inhibits adipocyte-derived leucine aminopeptidase and angiogenesis in a zebrafish model. Biochem. Biophys. Res. Commun. 2014, 450, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, E.; Fujita, K.; Kamataki, T.; Arimoto-Kobayashi, S.; Okamoto, K.; Negishi, T. Inhibition of human cytochrome P450 1B1, 1A1 and 1A2 by antigenotoxic compounds, purpurin and alizarin. Mutat. Res. 2002, 508, 147–156. [Google Scholar] [CrossRef]
- Takahashi, E.; Arimoto, S.; Okamoto, K.; Negishi, T. Enhancement of phase II enzyme activity by purpurin resulting in the suppression of MeIQx-DNA-adduct formation in mice. Mutat. Res. 2007, 626, 128–134. [Google Scholar] [CrossRef]
- Friedman, M.; Zhu, L.; Feinstein, Y.; Ravishankar, S. Carvacrol facilitates heat-induced inactivation of Escherichia coli O157:H7 and inhibits formation of heterocyclic amines in grilled ground beef patties. J. Agric. Food Chem. 2009, 57, 1848–1853. [Google Scholar] [CrossRef]
- Rounds, L.; Havens, C.M.; Feinstein, Y.; Friedman, M.; Ravishankar, S. Plant extracts, spices, and essential oils inactivate Escherichia coli O157:H7 and reduce formation of potentially carcinogenic heterocyclic amines in cooked beef patties. J. Agric. Food Chem. 2012, 60, 3792–3799. [Google Scholar] [CrossRef]
- Rounds, L.; Havens, C.M.; Feinstein, Y.; Friedman, M.; Ravishankar, S. Concentration-dependent inhibition of Escherichia coli O157:H7 and heterocyclic amines in heated ground beef patties by apple and olive extracts, onion powder and clove bud oil. Meat Sci. 2013, 94, 461–467. [Google Scholar] [CrossRef]
- Zengin, G.; Degirmenci, N.S.; Alpsoy, L.; Aktumsek, A. Evaluation of antioxidant, enzyme inhibition, and cytotoxic activity of three anthraquinones (alizarin, purpurin, and quinizarin). Hum. Exp. Toxicol. 2016, 35, 544–553. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, biochemistry, and dietary role of potato polyphenols. A review. J. Agric. Food Chem. 1997, 45, 1523–1540. [Google Scholar] [CrossRef]
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular dynamics study on the biophysical interactions of seven green tea catechins with lipid bilayers of cell membranes. J. Agric. Food Chem. 2008, 56, 7750–7758. [Google Scholar] [CrossRef]
- Sirk, T.W.; Friedman, M.; Brown, E.F. Molecular binding of black tea theaflavins to biological membranes: Relationship to bioactivities. J. Agric. Food Chem. 2011, 59, 3780–3787. [Google Scholar] [CrossRef] [PubMed]
- Le, J.; Zhang, X.; Jia, W.; Zhang, Y.; Luo, J.; Sun, Y.; Ye, J. Regulation of microbiota–GLP1 axis by sennoside A in diet-induced obese mice. Acta Pharm. Sin. B 2019, 9, 758–768. [Google Scholar] [CrossRef]
- Sgibnev, A.; Kremleva, E. Probiotics in addition to metronidazole for treatment Trichomonas vaginalis in the presence of BV: A randomized, placebo-controlled, double-blind study. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Abdali, K.; Jahed, L.; Amooee, S.; Zarshenas, M.; Tabatabaee, H.; Bekhradi, R. Comparison of the effect of vaginal Zataria multiflora cream and oral metronidazole pill on results of treatments for vaginal infections including trichomoniasis and bacterial vaginosis in women of reproductive age. Biomed. Res. Int. 2015, 2015, 683640. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. They can be purchased from chemical companies directly. |


{kind=link}
| T. vaginalis G3 (human) | T. foetus C1 (Feline) | T. foetus D1 (Bovine) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Compound | IC50 (µM) | Percent Inhibition (%) | Percent Error (%) | IC50 (µM) | Percent Inhibition (%) | Percent Error (%) | IC50 (µM) | Percent Inhibition (%) | Percent Error (%) |
| Purpurin | 101.60(1.0) b | 51.2 | 2.5 | 133.10(1.0) | 50.29 | 0.59 | 78.62(1.0) | 49.64 | 0.72 |
| Anthraquinone | 10.43 (9.8) | 50.7 | 1.4 | 25.55(5.2) | 45.6 | 8.8 | 9.19(8.6) | 51.4 | 2.9 |
| Anthrarufin | ND | ND | ND | 8.10(16.4) | 46.1 | 7.8 | 7.10(7.9) | 45.4 | 9.1 |
| Chrysazin | 8.90(11.4) | 52.9 | 5.7 | 0.979(137.8) | 47.4 | 5.3 | 0.98(80.2) | 53.1 | 6.3 |
| Aloe-Emodin | 0.6109(166.2) | 47.9 | 4.1 | 1.41(94.4) | 52.2 | 4.4 | 2.47(37.8) | 50.36 | 0.72 |
| Emodin | ND | ND | ND | ND | ND | ND | 8.00(9.8) | 48.0 | 4.0 |
| Rhein w/HPLC | 5.06(20.1) | 53.3 | 6.5 | 21.00(33.8) | 52.2 | 4.4 | 14.04(5.6) | 51.9 | 3.9 |
| Compound | Salmonella Enterica | Listeria Monocytogenes | Staphylococcus Aureus | Bacillus Cereus |
|---|---|---|---|---|
| DMSO | 0 | 0 | 0 | 0 |
| Levofloxacin 5 µg | 30 | 17 | 36 | 26 |
| Gentamicin 10 µg | 16 | 22 | 25 | 15 |
| Gentamicin 120 µg | 22 | 30 | 30 | 20 |
| Purpurin a | 0 | 0 | 0 | 0 |
| Anthraquinone a | 0 | 0 | 0 | 0 |
| Anthrarufin b | 0 | 0 | 0 | 0 |
| Chrysazin b | 0 | 0 | 0 | 0 |
| Aloe-Emodin b | 0 | 0 | 0 | 0 |
| Emodin b | 0 | 0 | 0 | 0 |
| Rhein w/HPLC b | 0 | 0 | 0 | 0 |
| Compound | Escherichia coli K12 | Lactobacillus Acidophilus | Lactobacillus Rhamnosus GG | Lactobacillus Reuteri |
|---|---|---|---|---|
| DMSO | 0 | 0 | 0 | 0 |
| Levofloxacin 5 µg | 30 | 0 | 15 | 5 |
| Gentamicin 10 µg | 18 | 8 | 8 | 11 |
| Gentamicin 120 µg | 20 | 15 | 15 | 20 |
| Purpurin a | 0 | 0 | 0 | 0 |
| Anthraquinone a | 0 | 0 | 0 | 0 |
| Anthrarufin b | 0 | 0 | 0 | 0 |
| Chrysazin b | 0 | 0 | 0 | 0 |
| Aloe-Emodin b | 0 | 0 | 0 | 0 |
| Emodin b | 0 | 0 | 0 | 0 |
| Rhein w/HPLC b | 0 | 0 | 0 | 0 |
| Compound | Concentration (mM) | A. fumigatus AF293 | C. albicans ATCC10231 |
|---|---|---|---|
| Aloin | 20 | 0.00 | 0.00 |
| Purpurin | 20 | 0.00 | 0.00 |
| Anthraquinone | 5 | 0.00 | 0.00 |
| Anthrarufin | 5 | 0.00 | 0.00 |
| Chrysazin | 5 | 0.00 | 0.00 |
| Aloe-Emodin | 5 | 0.00 | 0.00 |
| Emodin | 5 | 0.00 | 1.10 (76%) c |
| Octyl gallate | 5 | 1.15 (100%) c | 1.45 (100%) c |
| DMSO | 50 b | 0.00 (0%) c | 0.00 (0%) c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedman, M.; Xu, A.; Lee, R.; Nguyen, D.N.; Phan, T.A.; Hamada, S.M.; Panchel, R.; Tam, C.C.; Kim, J.H.; Cheng, L.W.; et al. The Inhibitory Activity of Anthraquinones against Pathogenic Protozoa, Bacteria, and Fungi and the Relationship to Structure. Molecules 2020, 25, 3101. https://doi.org/10.3390/molecules25133101
Friedman M, Xu A, Lee R, Nguyen DN, Phan TA, Hamada SM, Panchel R, Tam CC, Kim JH, Cheng LW, et al. The Inhibitory Activity of Anthraquinones against Pathogenic Protozoa, Bacteria, and Fungi and the Relationship to Structure. Molecules. 2020; 25(13):3101. https://doi.org/10.3390/molecules25133101
Chicago/Turabian StyleFriedman, Mendel, Alexander Xu, Rani Lee, Daniel N. Nguyen, Tina A. Phan, Sabrina M. Hamada, Rima Panchel, Christina C. Tam, Jong H. Kim, Luisa W. Cheng, and et al. 2020. "The Inhibitory Activity of Anthraquinones against Pathogenic Protozoa, Bacteria, and Fungi and the Relationship to Structure" Molecules 25, no. 13: 3101. https://doi.org/10.3390/molecules25133101
APA StyleFriedman, M., Xu, A., Lee, R., Nguyen, D. N., Phan, T. A., Hamada, S. M., Panchel, R., Tam, C. C., Kim, J. H., Cheng, L. W., & Land, K. M. (2020). The Inhibitory Activity of Anthraquinones against Pathogenic Protozoa, Bacteria, and Fungi and the Relationship to Structure. Molecules, 25(13), 3101. https://doi.org/10.3390/molecules25133101