
Bacterial Biotransformation of Oleic Acid: New Findings on the Formation of γ-Dodecalactone and 10-Ketostearic Acid in the Culture of Micrococcus luteus †

 , ,
, ,  and
and
Abstract
1. Introduction
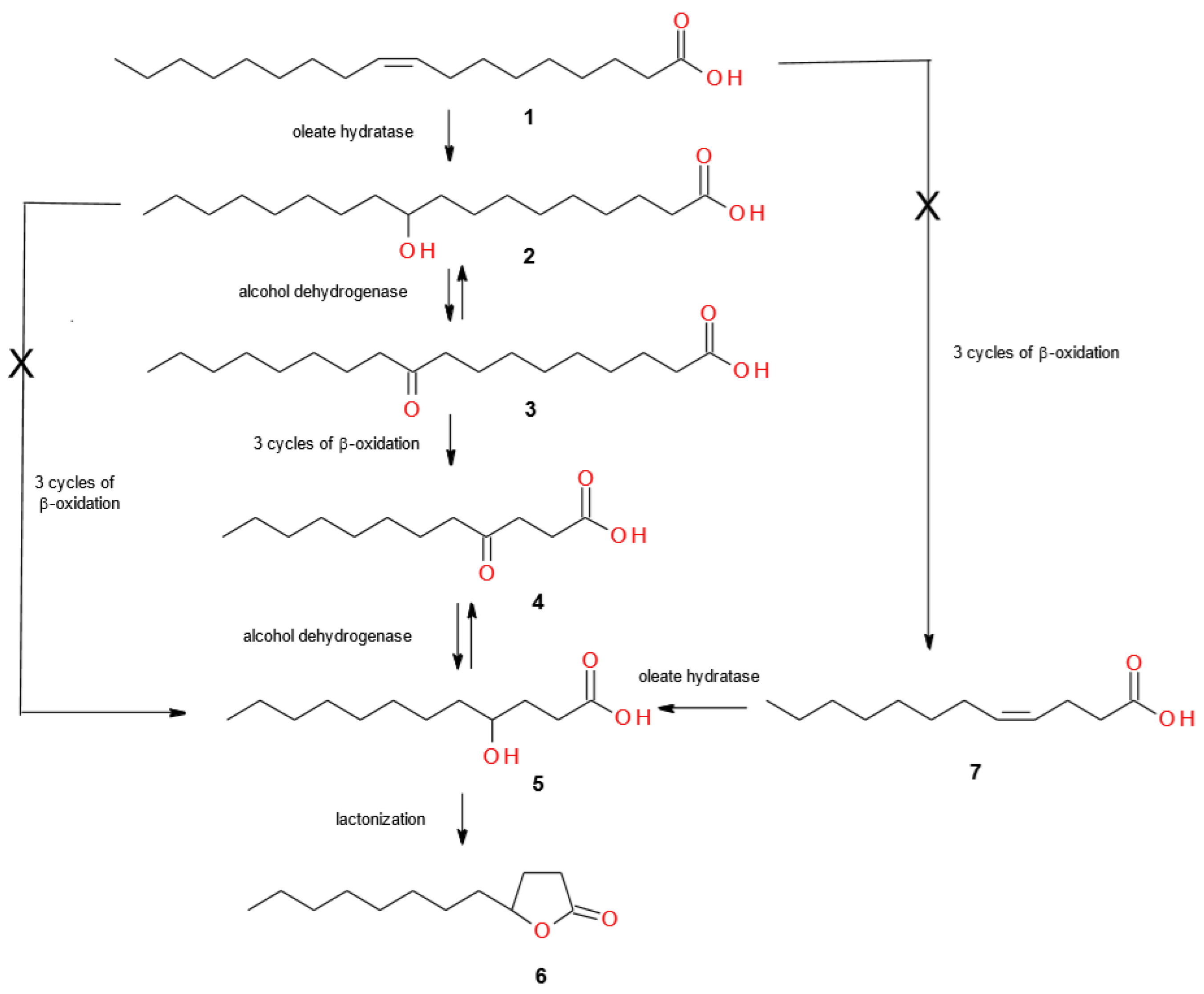
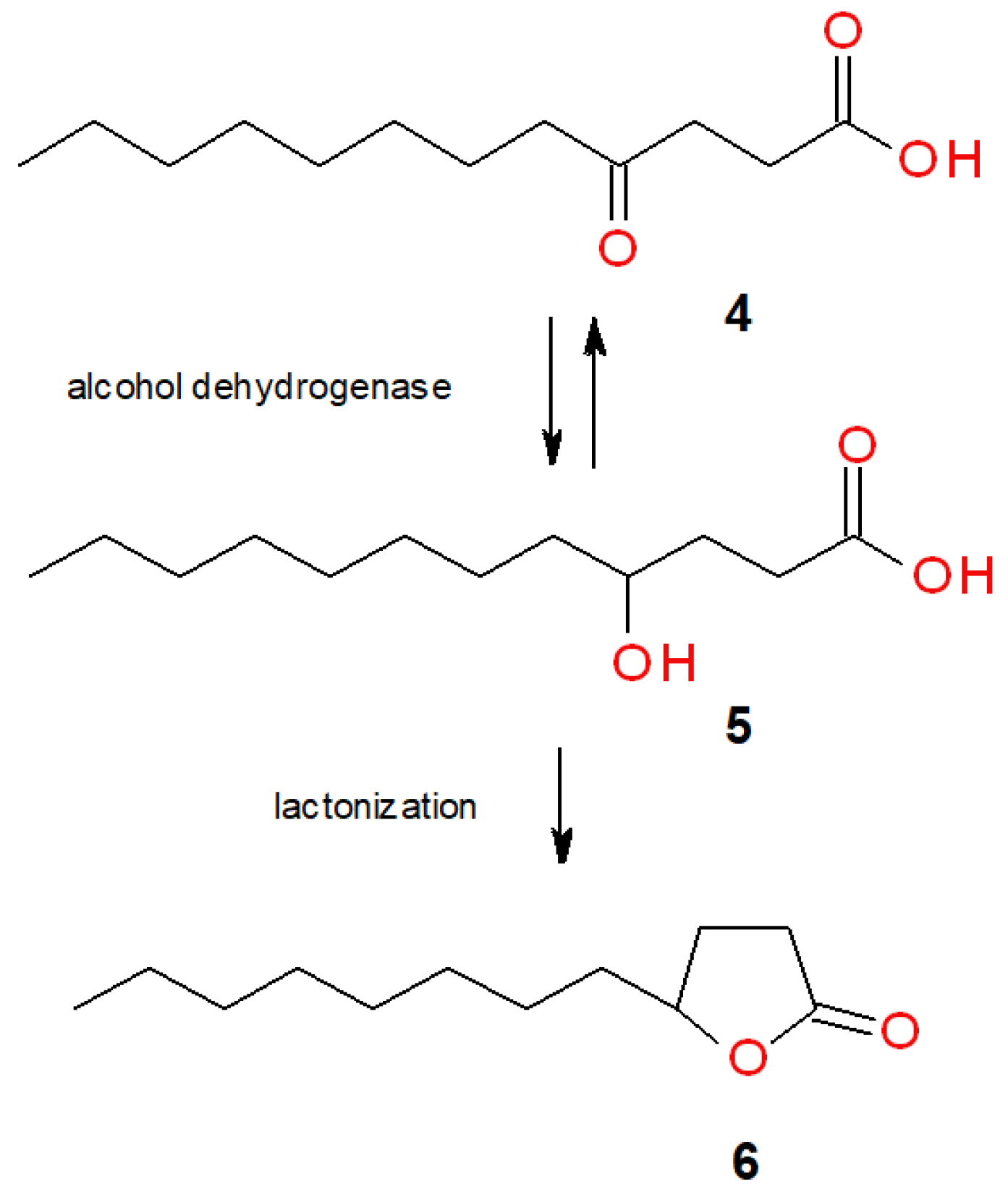
2. Results and Discussion
2.1. Screening Biotransformations of Oleic Acid (1)
2.2. Micrococcus Luteus PCM525 Transformations of Linoleic and α-Linolenic Acids
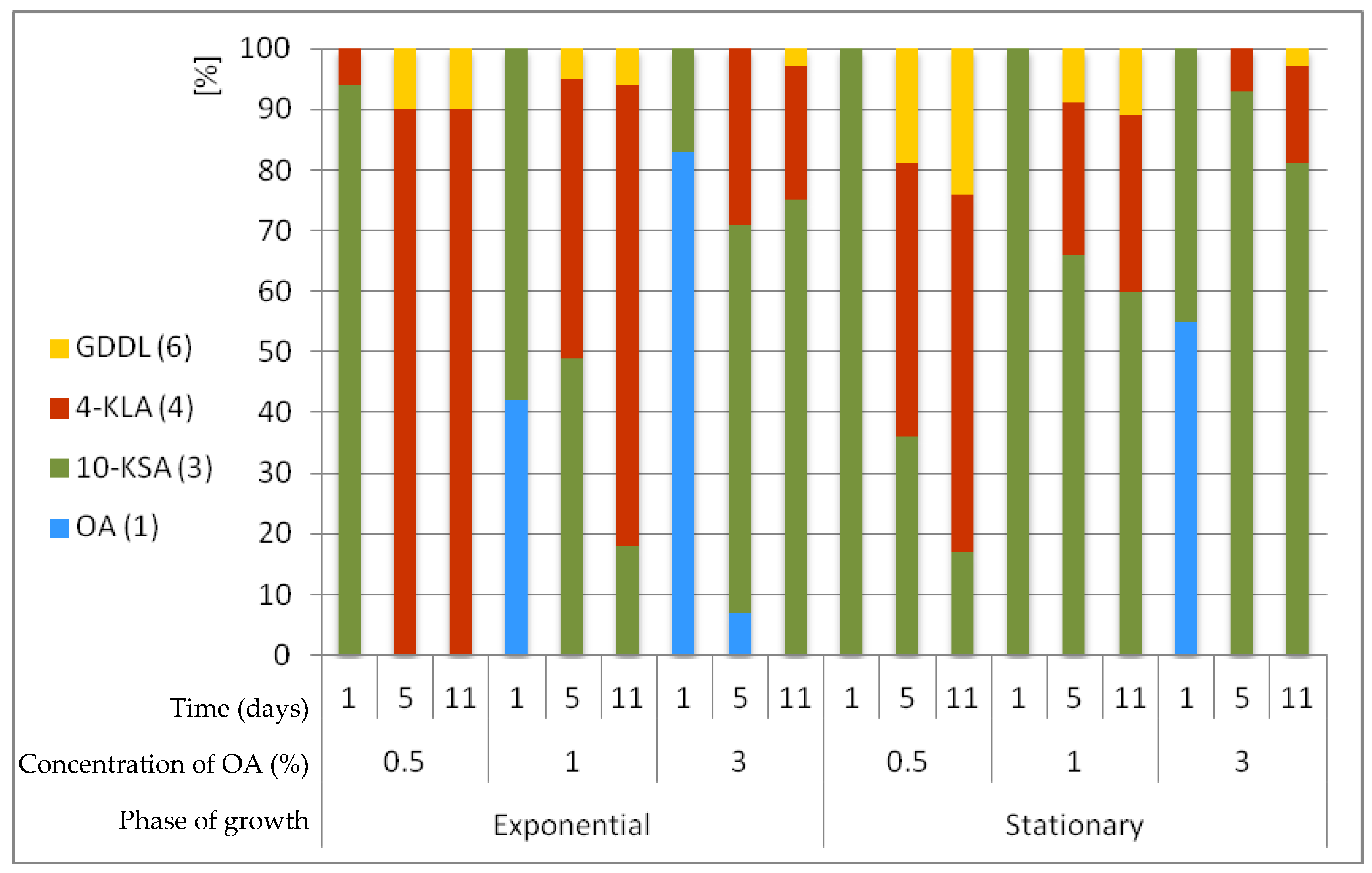
2.3. Micrococcus Luteus PCM525 Transformations Depending on the Growth Phase and Concentration of Oleic Acid (1)
2.4. Selection of Optimal Conditions for Biotransformations of Oleic Acid (1)
2.4.1. The Effect of Surfactants
2.4.2. The Effect of Media Aeration and Agitation
2.4.3. The Effect of Redox Potential
2.4.4. The Effect of Carbon Source and Technique of Biotransformation
3. Materials and Methods
3.1. Materials
3.2. Microorganisms
3.3. Media Composition
3.4. Biotransformation Process
3.4.1. Screening Procedure in Microtiter Plate
3.4.2. Screening Procedure in Erlenmeyer Flask
3.5. Process Optimization for Micrococcus luteus PCM525 Transformation
3.6. Preparation of Resting Cells of Micrococcus luteus PCM525
3.7. Pre-Growing of Micrococcus Luteus PCM525 in the Presence of a High Concentration of Oleic Acid (1)
3.8. Analysis
3.9. Characterization of Substrates and Products from Biotransformation Experiments
GC–MS Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maga, J.A. Lactones in foods. CRC Crit. Rev. Food Sci. Nutr. 1976, 8, 1–56. [Google Scholar] [CrossRef]
- Mosandl, A.; Guenther, C. Stereoisomeric flavor compounds. 20. Structure and properties of gamma-lactone enantiomers. J. Agric. Food Chem. 1989, 37, 413–418. [Google Scholar] [CrossRef]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Thonart, P.; Teixeira, J.A.; Destain, J.; Waché, Y. Biochemistry of lactone formation in yeast and fungi and its utilisation for the production of flavour and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Brenna, E.; Fuganti, C.; Gatti, F.G.; Serra, S. Biocatalytic methods for the synthesis of enantioenriched odor active compounds. Chem. Rev. 2011, 111, 4036–4072. [Google Scholar] [CrossRef] [PubMed]
- Boratynski, F.; Smuga, M.; Wawrzenczyk, C. Lactones 42. Stereoselective enzymatic/microbial synthesis of optically active isomers of whisky lactone. Food Chem. 2013, 141, 419–427. [Google Scholar] [CrossRef]
- Boratynski, F.; Dancewicz, K.; Paprocka, M.; Gabrys, B.; Wawrzenczyk, C. Chemo-Enzymatic Synthesis of Optically Active gamma- and delta-Decalactones and Their Effect on Aphid Probing, Feeding and Settling Behavior. PLoS ONE 2016, 11, e0146160. [Google Scholar] [CrossRef]
- Wache, Y.; Aguedo, M.; Nicaud, J.-M.; Belin, J.-M. Catabolism of hydroxyacids and biotechnological production of lactones by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2003, 61, 393–404. [Google Scholar] [CrossRef]
- Schöttler, M.; Boland, W. Über die Biosynthese von γ-Dodecanolacton in reifenden Früchten: Aroma-Komponenten der Erdbeere (Fragaria ananassa) und des Pfirsichs (Prunus persica). Helv. Chim. Acta 1995, 78, 847–856. [Google Scholar] [CrossRef]
- Elss, S.; Preston, C.; Hertzig, C.; Heckel, F.; Richling, E.; Schreier, P. Aroma profiles of pineapple fruit (Ananas comosus [L.] Merr.) and pineapple products. LWT-Food Sci. Technol. 2005, 38, 263–274. [Google Scholar] [CrossRef]
- Lalel, H.J.D.; Singh, Z.; Tan, S.C. Aroma volatiles production during fruit ripening of ‘Kensington Pride’ mango. Postharvest Biol. Technol. 2003, 27, 323–336. [Google Scholar] [CrossRef]
- Nunes, C.; Coimbra, M.A.; Saraiva, J.; Rocha, S.M. Study of the volatile components of a candied plum and estimation of their contribution to the aroma. Food Chem. 2008, 111, 897–905. [Google Scholar] [CrossRef]
- Pino, J.A.; Marbot, R. Volatile flavor constituents of acerola (Malpighia emarginata DC.) fruit. J. Agric. Food Chem. 2001, 49, 5880–5882. [Google Scholar] [CrossRef]
- Farbood, M.I.; Morris, J.A.; McLean, L.B. Fermentation process for preparing 10-hydroxy-C18-carboxylic acid and gamma-dodecalactone derivatives. European Patent 578,388, 12 January 1994. [Google Scholar]
- Serra, S.; Fuganti, C.; Brenna, E. Biocatalytic preparation of natural flavours and fragrances. Trends Biotechnol. 2005, 23, 193–198. [Google Scholar] [CrossRef]
- The European Parliament; The Council of the European Union. Flavourings and certain food ingredients with flavouring properties for use in and on foods. L 354/34. Off. J. Eur. Union 2008, 1334, 34–50. [Google Scholar]
- Gocho, S.; Tabogami, N.; Inagaki, M.; Kawabata, C.; Komai, T. Biotransformation of Oleic Acid to Optically Active γ-Dodecalactone. Biosci. Biotechnol. Biochem. 1995, 59, 1571–1572. [Google Scholar] [CrossRef]
- Yu, A.-Q.; Pratomo Juwono, N.K.; Leong, S.S.J.; Chang, M.W. Production of fatty acid-derived valuable chemicals in synthetic microbes. Front. Bioeng. Biotechnol. 2014, 2, 78. [Google Scholar] [CrossRef]
- Gomes, N.; Teixeira, J.A.; Belo, I. Fed-batch versus batch cultures of Yarrowia lipolytica for gamma-decalactone production from methyl ricinoleate. Biotechnol. Lett. 2012, 34, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.; Belo, I. Biotechnological production of gamma-decalactone, a peach like aroma, by Yarrowia lipolytica. World J. Microbiol. Biotechnol. 2016, 32, 169. [Google Scholar] [CrossRef]
- Scheller, U.; Zimmer, T.; Becher, D.; Schauer, F.; Schunck, W.H. Oxygenation cascade in conversion of n-alkanes to alpha,omega-dioic acids catalyzed by cytochrome P450 52A3. J. Biol. Chem. 1998, 273, 32528–32534. [Google Scholar] [CrossRef] [PubMed]
- Burg, N. ter.; Schaft, P.H. van der. Some aspects of the biocatalytic production of natural delta-decalactone from Massoia lactone by baker’s yeast. Dev. food Sci. 1994, 35, 481–486. [Google Scholar]
- Heshof, R.; de Graaff, L.H.; Villaverde, J.J.; Silvestre, A.J.D.; Haarmann, T.; Dalsgaard, T.K.; Buchert, J. Industrial potential of lipoxygenases. Crit. Rev. Biotechnol. 2016, 36, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, J.; Driller, R.; Waldow, A.; Qoura, F.; Loll, B.; Brück, T. Cover Feature: Rhodococcus erythropolis Oleate Hydratase: A New Member in the Oleate Hydratase Family Tree—Biochemical and Structural Studies (ChemCatChem 2/2018). ChemCatChem 2018, 10, 336. [Google Scholar] [CrossRef]
- Wallen, L.L.; Benedict, R.G.; Jackson, R.W. The microbiological production of 10-hydroxystearic acid from oleic acid. Arch. Biochem. Biophys. 1962, 99, 249–253. [Google Scholar] [CrossRef]
- Hou, C.T. Production of 10-Ketostearic Acid from Oleic Acid by Flavobacterium sp. Strain DS5 (NRRL B-14859). Appl. Environ. Microbiol. 1994, 60, 3760–3763. [Google Scholar] [CrossRef] [PubMed]
- Lanser, A.C.; Plattner, R.D.; Bagby, M.O. Production of 15-, 16- and 17-hydroxy-9-octadecenoic acids by bioconversion of oleic acid with Bacillus pumilus. J. Am. Oil Chem. Soc. 1992, 69, 363–366. [Google Scholar] [CrossRef]
- El-Sharkawy, S.H.; Yang, W.; Dostal, L.; Rosazza, J.P. Microbial oxidation of oleic acid. Appl. Environ. Microbiol. 1992, 58, 2116–2122. [Google Scholar] [CrossRef]
- Lanser, A.C. Conversion of oleic acid to 10-ketostearic acid by a Staphylococcus species. J. Am. Oil Chem. Soc. 1993, 70, 543–545. [Google Scholar] [CrossRef]
- Wanikawa, A.; Hosoi, K.; Kato, T. Conversion of unsaturated fatty acids to precursors of γ-lactones by lactic acid bacteria during the production of malt whisky. J. Am. Soc. Brew. Chem. 2000, 58, 51–56. [Google Scholar] [CrossRef]
- Serra, S.; De Simeis, D. Use of Lactobacillus rhamnosus (ATCC 53103) as Whole-Cell Biocatalyst for the Regio- and Stereoselective Hydration of Oleic, Linoleic, and Linolenic Acid. Catalysts 2018, 8, 109. [Google Scholar] [CrossRef]
- Haffner, T.; Tressl, R. Biosynthesis of (R)-γ-Decanolactone in the Yeast Sporobolomyces odorus. J. Agric. Food Chem. 1996, 44, 1218–1223. [Google Scholar] [CrossRef]
- Adam, P.; Hannemann, K.; Reiner, J.; Spiteller, G. 10-Hydroxystearic acid—Identified after homogenization of tissue—Is derived from bacteria. Z. Naturforsch.—Sect. C J. Biosci. 2000, 55, 965–970. [Google Scholar] [CrossRef]
- Serra, S.; De Simeis, D. New insights on the baker’s yeast-mediated hydration of oleic acid: The bacterial contaminants of yeast are responsible for the stereoselective formation of (R)-10-hydroxystearic acid. J. Appl. Microbiol. 2018, 124, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Serra, S.; De Simeis, D.; Castagna, A.; Valentino, M. The fatty-acid hydratase activity of the most common probiotic microorganisms. Catalysts 2020, 10, 154. [Google Scholar] [CrossRef]
- Jo, Y.-S.; An, J.-U.; Oh, D.-K. γ-Dodecelactone Production from Safflower Oil via 10-Hydroxy-12(Z)-octadecenoic Acid Intermediate by Whole Cells of Candida boidinii and Stenotrophomonas nitritireducens. J. Agric. Food Chem. 2014, 62, 6736–6745. [Google Scholar] [CrossRef] [PubMed]
- Tzirita, M.; Papanikolaou, S.; Quilty, B. Degradation of Fat by a Bioaugmentation Product Comprising of Bacillus spp. Before and After the Addition of a Pseudomonas sp. Eur. J. Lipid Sci. Technol. 2018, 120, 1–9. [Google Scholar] [CrossRef]
- Matsuoka, H.; Hirooka, K.; Fujita, Y. Organization and function of the YsiA regulon of Bacillus subtilis involved in fatty acid degradation. J. Biol. Chem. 2007, 282, 5180–5194. [Google Scholar] [CrossRef] [PubMed]
- Iram, S.H.; Cronan, J.E. The β-Oxidation Systems of Escherichia coli and Salmonella enterica Are Not Functionally Equivalent. Society 2006, 188, 599–608. [Google Scholar] [CrossRef]
- Zarzycki-Siek, J.; Norris, M.H.; Kang, Y.; Sun, Z.; Bluhm, A.P.; McMillan, I.A.; Hoang, T.T. Elucidating the Pseudomonas aeruginosa Fatty Acid Degradation Pathway: Identification of Additional Fatty Acyl-CoA Synthetase Homologues. PLoS ONE 2013, 8, e64554. [Google Scholar] [CrossRef]
- An, J.-U.; Joo, Y.-C.; Oh, D.-K. New Biotransformation Process for Production of the Fragrant Compound γ-Dodecalactone from 10-Hydroxystearate by Permeabilized Waltomyces lipofer Cells. Appl. Environ. Microbiol. 2013, 79, 2636–2641. [Google Scholar] [CrossRef]
- Marella, E.R.; Dahlin, J.; Dam, M.I.; ter Horst, J.; Christensen, H.B.; Sudarsan, S.; Wang, G.; Holkenbrink, C.; Borodina, I. A single-host fermentation process for the production of flavor lactones from non-hydroxylated fatty acids. Metab. Eng. 2019, 5, 1–10. [Google Scholar] [CrossRef]
- Ratledge, C. Yeasts, Moulds, Algae and Bacteria as Sources of Lipids BT—Technological Advances in Improved and Alternative Sources of Lipids; Kamel, B.S., Kakuda, Y., Eds.; Springer: Boston, MA, USA, 1994; pp. 235–291. ISBN 978-1-4615-2109-9. [Google Scholar]
- Zabeti, N.; Bonin, P.; Volkman, J.K.; Guasco, S.; Rontani, J.-F. Fatty acid composition of bacterial strains associated with living cells of the haptophyte Emiliania huxleyi. Org. Geochem. 2010, 41, 627–636. [Google Scholar] [CrossRef]
- Milkessa, T.J.; Abate, D. Oleaginous microorganisms, diversity, lipid biosynthesis pathway and strain improvement. WebPub J. Sci. Res. 2014, 2, 55–65. [Google Scholar]
- Kocur, M.; Pacova, Z.; Martinec, T. Taxonomic Status of Micrococcus luteus (Schroeter 1872) Cohn 1872, and Designation ofthe Neotype Strain. Int. J. Syst. Bacteriol. 1972, 22, 218–223. [Google Scholar] [CrossRef][Green Version]
- Kaprelyants, A.S.; Kell, D.B. Dormancy in Stationary-Phase Cultures of Micrococcus luteus: Flow Cytometric Analysis of Starvation and Resuscitation. Appl. Environ. Microbiol. 1993, 59, 3187–3196. [Google Scholar] [CrossRef]
- Akbar, A.; Sitara, U.; Ali, I.; Muhammad, N.; Khan, S.A. Isolation and characterization of biotechnologically potent Micrococcus luteus strain from environment. Pak. J. Zool. 2014, 46, 967–973. [Google Scholar]
- Abd El-Rhman, A.M.; Khattab, Y.A.E.; Shalaby, A.M.E. Micrococcus luteus and Pseudomonas species as probiotics for promoting the growth performance and health of Nile tilapia, Oreochromis niloticus. Fish. Shellfish Immunol. 2009, 27, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Alsulaiman, A.; Nizam, A.A. Evaluation of the ability of Micrococcus sp. isolated from Barada river to biodegradation of vegetable oil wastes. Int. Food Res. J. 2018, 25, 345–350. [Google Scholar]
- Sandrin, T.R.; Maier, R.M. Impact of metals on the biodegradation of organic pollutants. Environ. Health Perspect. 2003, 111, 1093–1101. [Google Scholar] [CrossRef]
- Dib, J.R.; Liebl, W.; Wagenknecht, M.; Farías, M.E.; Meinhardt, F. Extrachromosomal genetic elements in Micrococcus. Appl. Microbiol. Biotechnol. 2013, 97, 63–75. [Google Scholar] [CrossRef]
- Biskupiak, J.E.; Meyers, E.; Gillum, A.M.; Dean, L.; Trejo, W.H.; Kirsch, D.R. Neoberninamycin, a new antibiotic produced by Micrococcus luteus. J. Antibiot. (Tokyo) 1988, 41, 684–687. [Google Scholar] [CrossRef][Green Version]
- Akita, K.; Naitou, C.; Maruyama, K. Purification and characterization of an esterase from Micrococcus sp. YGJ1 hydrolyzing phthalate esters. Biosci. Biotechnol. Biochem. 2001, 65, 1680–1683. [Google Scholar] [CrossRef]
- Manikandan, M.; Kannan, V.; Pasic, L. Extraction, purification and characterization of a protease from Micrococcus sp. VKMM 037. Environ. Technol. 2011, 32, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-K.; Park, J.K.; Dhungana, B.R.; Youngblut, N.D.; Lin, C.-T.; Wen, L. A novel secondary alcohol dehydrogenase from Micrococcus luteus WIUJH20: Purification, cloning, and properties. FASEB J. 2010, 24, 835.5. [Google Scholar]
- Youngblut, N.D.; Huang, J.-K.; Wen, L. Secondary Alcohol Dehydrogenase in Micrococcus luteus (WIUJH-20). FASEB J. 2008, 22, 625-9. [Google Scholar]
- Seo, E.J.; Kim, H.J.; Kim, M.J.; Kim, J.S.; Park, J.B. Cofactor specificity engineering of a long-chain secondary alcohol dehydrogenase from: Micrococcus luteus for redox-neutral biotransformation of fatty acids. Chem. Commun. 2019, 55, 14462–14465. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.X.; Pan, J.; Yu, H.L.; Xu, J.H. Enzymatic synthesis of 10-oxostearic acid in high space-time yield via cascade reaction of a new oleate hydratase and an alcohol dehydrogenase. J. Biotechnol. X 2019, 2, 100008. [Google Scholar] [CrossRef]
- Kuo, T.M.; Lanser, A.C.; Kaneshiro, T.; Hou, C.T. Conversion of oleic acid to 10-ketostearic acid by Sphingobacterium sp. strain O22. JAOCS J. Am. Oil Chem. Soc. 1999, 76, 709–712. [Google Scholar] [CrossRef]
- Wanikawa, A.; Shoji, H.; Hosoi, K.; Nakagawa, K.I. Stereospecificity of 10-hydroxystearic acid and formation of 10-ketostearic acid by lactic acid bacteria. J. Am. Soc. Brew. Chem. 2002, 60, 14–20. [Google Scholar] [CrossRef]
- Guerrero, A.; Casals, I.; Busquets, M.; Leon, Y.; Manresa, A. Oxydation of oleic acid to (E)-10-hydroperoxy-8-octadecenoic and (E)-10-hydroxy-8-octadecenoic acids by Pseudomonas sp. 42A2. Biochim. Biophys. Acta 1997, 1347, 75–81. [Google Scholar] [CrossRef]
- Kuo, T.M.; Levinson, W.E. Biocatalytic production of 10-hydroxystearic acid, 10-ketostearic acid, and their primary fatty amides. J. Am. Oil Chem. Soc. 2006, 83, 671–675. [Google Scholar] [CrossRef]
- Heilig, M.L. United States Patent Office. ACM SIGGRAPH Comput. Graph. 1994, 28, 131–134. [Google Scholar] [CrossRef]
- Lu, W.; Ness, J.E.; Xie, W.; Zhang, X.; Minshull, J.; Gross, R.A. Biosynthesis of Monomers for Plastics from Renewable Oils. J. Am. Chem. Soc. 2010, 132, 15451–15455. [Google Scholar] [CrossRef] [PubMed]
- Boratyński, F.; Pannek, J.; Walczak, P.; Janik-Polanowicz, A.; Huszcza, E.; Szczepańska, E.; Martinez-Rojas, E.; Olejniczak, T. Microbial alcohol dehydrogenase screening for enantiopure lactone synthesis: Down-stream process from microtiter plate to bench bioreactor. Process. Biochem. 2014, 49, 1637–1646. [Google Scholar] [CrossRef]
- Duetz, W.A. Microtiter plates as mini-bioreactors: Miniaturization of fermentation methods. Trends Microbiol. 2007, 15, 469–475. [Google Scholar] [CrossRef]
- Márquez-Ruiz, G.; Rodríguez-Pino, V.; de la Fuente, M.A. Determination of 10-hydroxystearic, 10-ketostearic, 8-hydroxypalmitic, and 8-ketopalmitic acids in milk fat by solid-phase extraction plus gas chromatography-mass spectrometry. J. Dairy Sci. 2011, 94, 4810–4819. [Google Scholar] [CrossRef]
- Weihrauch, J.L.; Brewington, C.R.; Schwartz, D.P. Trace constituents in milk fat: Isolation and identification of oxofatty acids. Lipids 1974, 9, 883–890. [Google Scholar] [CrossRef]
- Esaki, N.; Ito, S.; Blank, W.; Soda, K. Biotransformation of Oleic Acid by Micrococcus luteus Cells. Biosci. Biotechnol. Biochem. 1994, 58, 319–321. [Google Scholar] [CrossRef]
- Blank, W.; Takayanagi, H.; Kido, T.; Esaki, N.; Soda, K.; Meussdoerffer, F. Transformation of Oleic Acid and Its Esters by Sarcina lutea. Agric. Biol. Chem. 1991, 55, 2651–2652. [Google Scholar] [CrossRef]
- Litchfield, J.H.; Pierce, G.E. United States Patent (19) 4,582,804, 15 April 1986.
- Horikawa, M.; Tateda, K.; Tuzuki, E.; Ishii, Y.; Ueda, C.; Takabatake, T.; Miyairi, S.; Yamaguchi, K.; Ishiguro, M. Synthesis of Pseudomonas quorum-sensing autoinducer analogs and structural entities required for induction of apoptosis in macrophages. Bioorg. Med. Chem. Lett. 2006, 16, 2130–2133. [Google Scholar] [CrossRef]
Sample Availability: Samples of the tested compounds (1,2,6) are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Strain | Time (Days) | Conversion (%) | Products | |||
|---|---|---|---|---|---|---|
| 10-HSA (2) (%) | 10-KSA (3) (%) | 4-KLA (4) (%) | GDDL (6) (%) | |||
| B.benzoevorans DSM5391 | 3 | 13 | 13 | 0 | 0 | 0 |
| Dietzia sp. DSM44016 | 1 | 100 | 0 | 100 | 0 | 0 |
| 2 | 100 | 0 | 0 * | 0 | 0 | |
| M. luteus PCM525 | 1 | 100 | 0 | 83 | 12 | 5 |
| 5 | 100 | 0 | 0 | 50 | 50 | |
| R. erythropolis DSM44534 | 1 | 100 | 0 | 100 | 0 | 0 |
| 6 | 100 | 0 | 0 * | 0 | 0 | |
| R. ruber PCM2166 | 1 | 100 | 5 | 95 | 0 | 0 |
| 6 | 100 | 4 | 96 | 0 | 0 | |
| Surfactant | Time (Days) | Conversion (%) | Substrate | Products | ||
|---|---|---|---|---|---|---|
| OA (1) (%) | 10-KSA (3) (%) | 4-KLA (4) (%) | GDDL (6) (%) | |||
| Glycerol | 5 | 100 | 0 | 65 (±3) * | 25 (±1) | 10 (±2) |
| Triton X-100 | 5 | 100 | 0 | 62 (±4) | 30 (±2) | 8 (±2) |
| Tween-80 | 5 | 98 (±2) | 2 | 93 | 3 | 2 (±2) |
| No. | Additive | Time (Days) | Conversion (%) | Substrate | Products | |||
|---|---|---|---|---|---|---|---|---|
| OA (1) (%) | 10-KSA (3) (%) | 4-KLA (4) (%) | GDDL (6) (%) | |||||
| 1 | With addition of OA (1) | HCl * | 5 | 100 | 0 | 59 (±2) ** | 25 (±1) | 16 (±1) |
| 11 | 100 | 0 | 44 (±4) | 36 (±2) | 20 (±2) | |||
| 2 | Glucose | 5 | 100 | 0 | 100 | 0 | 0 | |
| 11 | 100 | 0 | 100 | 0 | 0 | |||
| 3 | HCl + Glucose | 5 | 100 | 0 | 100 | 0 | 0 | |
| 11 | 100 | 0 | 100 | 0 | 0 | |||
| 4 | When 4-KDDA (4) was formed | HCl | 5 | 100 | 0 | 48 (±3) | 33 (±2) | 19 (±1) |
| 11 | 100 | 0 | 19 (±2) | 54 (±1) | 27 (±3) | |||
| 5 | HCl + Glucose | 5 | 100 | 0 | 54 | 28 (±4) | 18 (±4) | |
| 11 | 100 | 0 | 25 (±1) | 47 (±2) | 28 (±3) | |||
| No. | Substrate | Media | Time (Days) | Conversion (%) | Products | |||
|---|---|---|---|---|---|---|---|---|
| 10-HSA (2) (%) | 10-KSA (3) (%) | 4-KLA (4) (%) | GDDL (6) (%) | |||||
| 1 | OA (1) | PCM | 2 | 100 | 0 | 0 | 69 (±4) * | 31 (±4) |
| 5 | 100 | 0 | 0 | 63 (±1) | 37 (±1) | |||
| 2 | PCM (no glucose) | 2 | 0 | 0 | 0 | 0 | 0 | |
| 6 | 0 | 0 | 0 | 0 | 0 | |||
| 3 | buffer | 2 | 100 | 15 (±5) | 85 (±5) | 0 | 0 | |
| 5 | 100 | 12 (±1) | 88 (±1) | 0 | 0 | |||
| 4 | 10-HSA (2) | PCM | 2 | 93 (±4) | 7 (±4) | 90 (±2) | 3 (±2) | 0 |
| 5 | 100 | 0 | 96 (±4) | 4 (±4) | 0 | |||
| 5 | PCM (no glucose) | 2 | 100 | 0 | 100 | 0 | 0 | |
| 6 | 100 | 0 | 100 | 0 | 0 | |||
| 6 | buffer | 2 | 25 (±2) | 75 (±2) | 25 (±2) | 0 | 0 | |
| 5 | 28 (±1) | 72 (±1) | 28 (±1) | 0 | 0 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boratyński, F.; Szczepańska, E.; De Simeis, D.; Serra, S.; Brenna, E. Bacterial Biotransformation of Oleic Acid: New Findings on the Formation of γ-Dodecalactone and 10-Ketostearic Acid in the Culture of Micrococcus luteus. Molecules 2020, 25, 3024. https://doi.org/10.3390/molecules25133024
Boratyński F, Szczepańska E, De Simeis D, Serra S, Brenna E. Bacterial Biotransformation of Oleic Acid: New Findings on the Formation of γ-Dodecalactone and 10-Ketostearic Acid in the Culture of Micrococcus luteus. Molecules. 2020; 25(13):3024. https://doi.org/10.3390/molecules25133024
Chicago/Turabian StyleBoratyński, Filip, Ewa Szczepańska, Davide De Simeis, Stefano Serra, and Elisabetta Brenna. 2020. "Bacterial Biotransformation of Oleic Acid: New Findings on the Formation of γ-Dodecalactone and 10-Ketostearic Acid in the Culture of Micrococcus luteus" Molecules 25, no. 13: 3024. https://doi.org/10.3390/molecules25133024
APA StyleBoratyński, F., Szczepańska, E., De Simeis, D., Serra, S., & Brenna, E. (2020). Bacterial Biotransformation of Oleic Acid: New Findings on the Formation of γ-Dodecalactone and 10-Ketostearic Acid in the Culture of Micrococcus luteus. Molecules, 25(13), 3024. https://doi.org/10.3390/molecules25133024