
Hydromethanolic Extracts from Adansonia digitata L. Edible Parts Positively Modulate Pathophysiological Mechanisms Related to the Metabolic Syndrome

 , , ,
, , ,  ,
, 
 and
and 
Abstract
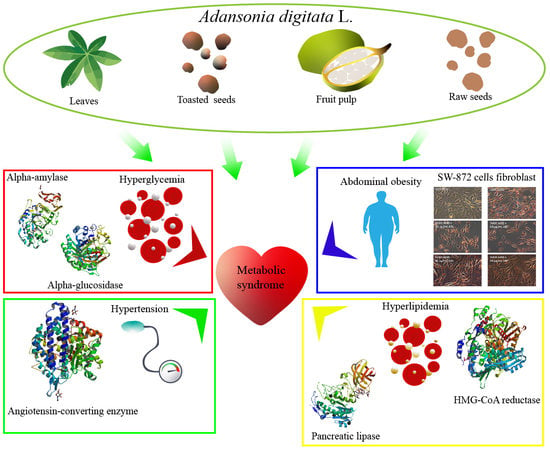
:
1. Introduction
2. Results
2.1. Baobab Edible Part Extracts
2.2. In Vitro Enzymatic Inhibition Assays
2.2.1. Alpha-Amylase Inhibition Assay
2.2.2. Alpha-Glucosidase Inhibition Assay
2.2.3. ACE Inhibition Assay
2.2.4. HMG-CoAR Inhibition Assay
2.2.5. Pancreatic Lipase Inhibition Assay
2.3. Effects of Baobab Fruit Pulp and Leaf Extracts on the Differentiation of SW-872 Human Liposarcoma Cells to Mature Adipocytes
2.3.1. Differentiation of SW-872 Preadipocytes to Adipocytes
2.3.2. Effect of Baobab Fruit Pulp and Leaf Extracts on SW-872 Cell Viability
2.3.3. Effect of Baobab Fruit Pulp and Leaf Extracts on SW-872 Differentiation
2.4. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of Food Materials and Food Extracts
4.3. In Vitro Enzymatic Assays
4.4. Evaluation of the Effects of Baobab Pulp and Leaf Plant Extracts on SW872 Human Liposarcoma Cells Differentiated to Mature Adipocytes
4.4.1. SW-872 Cell Culture
4.4.2. Differentiation of SW-872 Preadipocytes to Mature Adipocytes
4.4.3. Oil red O (ORO) Staining/Hematoxylin Counterstaining of Nuclei
4.4.4. Triglyceride Content Measurement
4.5. Ligand-Based Pharmacophore Modeling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C. Harmonizing the metabolic syndrome: A joint interim statement of the international diabetes federation task force on epidemiology and prevention; national heart, lung, and blood institute; american heart association; world heart federation; international atherosclerosis society; and international association for the study of obesity. Circ. J. 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desroches, S.; Lamarche, B. The evolving definitions and increasing prevalence of the metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Kolovou, G.D.; Anagnostopoulou, K.K.; Salpea, K.D.; Mikhailidis, D.P. The prevalence of metabolic syndrome in various populations. Am. J. Med. Sci. 2007, 333, 362–371. [Google Scholar] [CrossRef]
- Rask Larsen, J.; Dima, L.; Correll, C.U.; Manu, P. The pharmacological management of metabolic syndrome. Expert. Rev. Clin. Pharmacol. 2018, 11, 397–410. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic syndrome: Pathophysiology, management, and modulation by natural compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Ramadan, A.; Harraz, F.M.H.; Elmougy, S.A. Anti-inflammatory, analgesic and antipyretic effects of the fruit pulp of adansonia digitata. Fitoterapia 1994, 65, 418–422. [Google Scholar]
- Rahul, J.; Jain, M.K.; Singh, S.P.; Kamal, R.K.; Naz, A.; Gupta, A.K.; Mrityunjay, S.K. Adansonia digitata L. (baobab): A review of traditional information and taxonomic description. Asian Pac. J. Trop. Biomed. 2015, 5, 79–84. [Google Scholar] [CrossRef] [Green Version]
- Sugandha, S.; Shashi, R. Spectroscopic determination of total phenol, flavonoid content and anti-oxidant activity in different parts of Adansonia digitata L.: An important medicinal tree. Eur. J. Pharm. Med. Res. 2017, 4, 549–552. [Google Scholar]
- Zahra’u, B.; Mohammed, A.S.; Ghazali, H.M.; Karim, R. Baobab Tree (Adansonia digitata L.) Parts: Nutrition, Applications in Food and Uses in Ethno-medicine—A Review. Ann Nutr. Disord. Ther. 2014, 1, 1011. [Google Scholar] [CrossRef]
- Tanko, Y.; Yerima, M.; Mahdi, M.; Yaro, A. Hypoglycemic activity of methanolic stem bark of adansonnia digitata extract on blood glucose levels of streptozocin-induced diabetic wistar rats. Int. J. Appl. 2008, 1, 32–36. [Google Scholar]
- Geidam, M.; Chabiri, S.; Shettima, Y. Effects of methanolic leaf extract of adansonia digitata on serum lipid levels in normal and ethanol fed rats. J. Biol. Sci. 2004, 7, 1094–1095. [Google Scholar] [CrossRef]
- Bako, H.Y.; Mohammad, J.S.; Wazir, P.M.; Bulus, T.; Gwarzo, M.Y.; Zubairu, M.M. Lipid profile of alloxan-induced diabetic wistar rats treated with methanolic extract of adansonia digitata fruit pulp. Sci. World J. 2014, 9, 19–24. [Google Scholar]
- Ebaid, H.; Bashandy, S.A.E.; Alhazza, I.M.; Hassan, I.; Al-Tamimi, J. Efficacy of a methanolic extract of adansonia digitata leaf in alleviating hyperglycemia, hyperlipidemia, and oxidative stress of diabetic rats. Biomed. Res. Int. 2019, 2019, 2835152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassef, H.; Bernier, L.; Davignon, J.; Cohn, J.S. Synthesis and secretion of apoc-i and apoe during maturation of human sw872 liposarcoma cells. J. Nutr. 2004, 134, 2935–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmel, J.F.; Tarnus, E.; Cohn, J.S.; Bourdon, E.; Davignon, J.; Bernier, L. High expression of apolipoprotein e impairs lipid storage and promotes cell proliferation in human adipocytes. J. Cell. Biochem. 2009, 106, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Campioli, E.; Batarseh, A.; Li, J.; Papadopoulos, V. The endocrine disruptor mono-(2-ethylhexyl) phthalate affects the differentiation of human liposarcoma cells (sw 872). PLoS ONE 2011, 6, e28750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsetegho Sokeng, A.J.; Sobolev, A.P.; Di Lorenzo, A.; Xiao, J.; Mannina, L.; Capitani, D.; Daglia, M. Metabolite characterization of powdered fruits and leaves from Adansonia digitata L. (baobab): A multi-methodological approach. Food Chem. 2019, 272, 93–108. [Google Scholar] [CrossRef]
- Jhong, C.H.; Riyaphan, J.; Lin, S.H.; Chia, Y.C.; Weng, C.F. Screening alpha-glucosidase and alpha-amylase inhibitors from natural compounds by molecular docking in silico. Biofactors 2015, 41, 242–251. [Google Scholar] [CrossRef]
- Islam, B.; Sharma, C.; Adem, A.; Aburawi, E.; Ojha, S. Insight into the mechanism of polyphenols on the activity of hmgr by molecular docking. Drug Des. Dev. Ther. 2015, 9, 4943–4951. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Gonzalez, A.I.; Alvarez-Parrilla, E.; Diaz-Sanchez, A.G.; de la Rosa, L.A.; Nunez-Gastelum, J.A.; Vazquez-Flores, A.A.; Gonzalez-Aguilar, G.A. In vitro inhibition of pancreatic lipase by polyphenols:A kinetic, fluorescence spectroscopy and molecular docking study. Food Technol. Biotechnol. 2017, 55, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, L.; Castillo, J.; Quinones, M.; Garcia-Vallve, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of angiotensin-converting enzyme activity by flavonoids: Structure-activity relationship studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [Green Version]
- Irondi, E.A.; Akintunde, J.K.; Agboola, S.O.; Boligon, A.A.; Athayde, M.L. Blanching influences the phenolics composition, antioxidant activity, and inhibitory effect of adansonia digitata leaves extract on alpha-amylase, alpha-glucosidase, and aldose reductase. Food Sci. Nutr. 2017, 5, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Coe, S.A.; Clegg, M.; Armengol, M.; Ryan, L. The polyphenol-rich baobab fruit (Adansonia digitata L.) reduces starch digestion and glycemic response in humans. Nutr. Res. 2013, 33, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Gadour, M.O.; Khidir, H.B.; Adam, I.; Gasim, G.I. Effects of a powder of the fruit of adansonia digitata (tabaldia, gongolase, or baobab tree) on serum lipids. J. Herb. Med. 2017, 8, 14–16. [Google Scholar] [CrossRef]
- Saravanaraj, M.G. Anti-diabetic effect of the ethanolic extract of dried fruits of adansonia digitata linn. World J. Pharm. Pharm. Sci. 2017, 6, 1597–1605. [Google Scholar] [CrossRef] [Green Version]
- Parellada, J.; Suarez, G.; Guinea, M. Inhibition of zinc metallopeptidases by flavonoids and related phenolic compounds: Structure-activity relationships. J. Enzyme Inhib. 1998, 13, 347–359. [Google Scholar] [CrossRef]
- Ende, C.; Gebhardt, R. Inhibition of matrix metalloproteinase-2 and -9 activities by selected flavonoids. Planta Med. 2004, 70, 1006–1008. [Google Scholar] [CrossRef]
- Atchan Nwakiban, A.P.; Sokeng, A.J.; Dell’Agli, M.; Bossi, L.; Beretta, G.; Gelmini, F.; Deutou Tchamgoue, A.; Agbor Agbor, G.; Kuiaté, J.R.; Daglia, M.; et al. Hydroethanolic plant extracts from cameroon positively modulate enzymes relevant to carbohydrate/lipid digestion and cardio-metabolic diseases. Food Funct. 2019, 10, 6533–6542. [Google Scholar] [CrossRef]
- Wolber, G.; Dornhofer, A.A.; Langer, T. Efficient overlay of small organic molecules using 3d pharmacophores. J. Comput. Aided Mol. Des. 2006, 20, 773–788. [Google Scholar] [CrossRef]
- Hawkins, P.C.; Skillman, A.G.; Warren, G.L.; Ellingson, B.A.; Stahl, M.T. Conformer generation with omega: Algorithm and validation using high quality structures from the protein databank and cambridge structural database. J. Chem. Inf. Model. 2010, 50, 572–584. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the baobab extracts are available from the authors. |






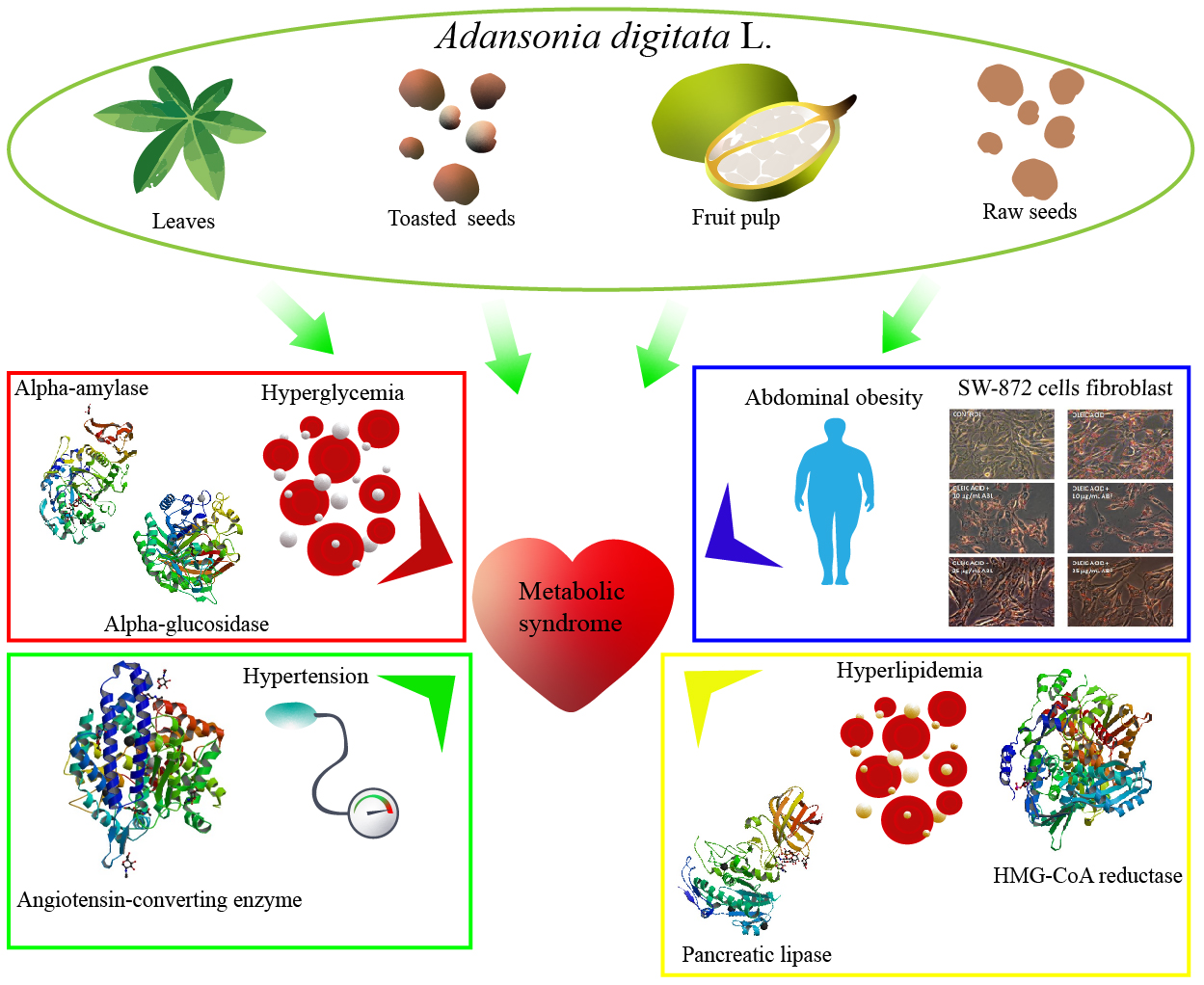{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Treatment | mg/mL (Mean (SD)) | % |
|---|---|---|
| Control | 1.318 (0.131) | 100 |
| Oleic acid | 2.821 (0.660) | 214 |
| Oleic acid + baobab leaf extract | 3.158 (0.281) | 240 |
| Oleic acid + baobab fruit pulp extract | 3.005 (0.213) | 228 |
| Molecular Target | Catalytic Site | Allosteric Site | Residues Involved | Ref. |
|---|---|---|---|---|
| Alpha-glucosidase | √ | R407, R197, R407, F282 | Jhong et al., 2015 [19] | |
| Alpha-Amylase | √ | K200, E233, D300, W151, A198, L162 | Jhong et al., 2015 [19] | |
| 3-Hydroxy-3-methylglutaryl-coenzyme A reductase (HMGCoAR) | √ (NADP binding site) | K691, E559, D767 | Islam et al., 2015 [20] | |
| Pancreatic Lipase | √ | F78, Y115, H152, F216 | Martinez Gonzales et al., 2017 [21] | |
| Angiotensin-converting enzyme (ACE) | √ | S1′, S2′/S1′, S2′ regions | Guerrero et al., 2012 [22] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cicolari, S.; Dacrema, M.; Tsetegho Sokeng, A.J.; Xiao, J.; Atchan Nwakiban, A.P.; Di Giovanni, C.; Santarcangelo, C.; Magni, P.; Daglia, M. Hydromethanolic Extracts from Adansonia digitata L. Edible Parts Positively Modulate Pathophysiological Mechanisms Related to the Metabolic Syndrome. Molecules 2020, 25, 2858. https://doi.org/10.3390/molecules25122858
Cicolari S, Dacrema M, Tsetegho Sokeng AJ, Xiao J, Atchan Nwakiban AP, Di Giovanni C, Santarcangelo C, Magni P, Daglia M. Hydromethanolic Extracts from Adansonia digitata L. Edible Parts Positively Modulate Pathophysiological Mechanisms Related to the Metabolic Syndrome. Molecules. 2020; 25(12):2858. https://doi.org/10.3390/molecules25122858
Chicago/Turabian StyleCicolari, Stefania, Marco Dacrema, Arold Jorel Tsetegho Sokeng, Jianbo Xiao, Achille Parfait Atchan Nwakiban, Carmen Di Giovanni, Cristina Santarcangelo, Paolo Magni, and Maria Daglia. 2020. "Hydromethanolic Extracts from Adansonia digitata L. Edible Parts Positively Modulate Pathophysiological Mechanisms Related to the Metabolic Syndrome" Molecules 25, no. 12: 2858. https://doi.org/10.3390/molecules25122858
APA StyleCicolari, S., Dacrema, M., Tsetegho Sokeng, A. J., Xiao, J., Atchan Nwakiban, A. P., Di Giovanni, C., Santarcangelo, C., Magni, P., & Daglia, M. (2020). Hydromethanolic Extracts from Adansonia digitata L. Edible Parts Positively Modulate Pathophysiological Mechanisms Related to the Metabolic Syndrome. Molecules, 25(12), 2858. https://doi.org/10.3390/molecules25122858