Enzymatic Modification of Porphyra dioica-Derived Proteins to Improve their Antioxidant Potential

 ,
,  ,
,  ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Protein Content
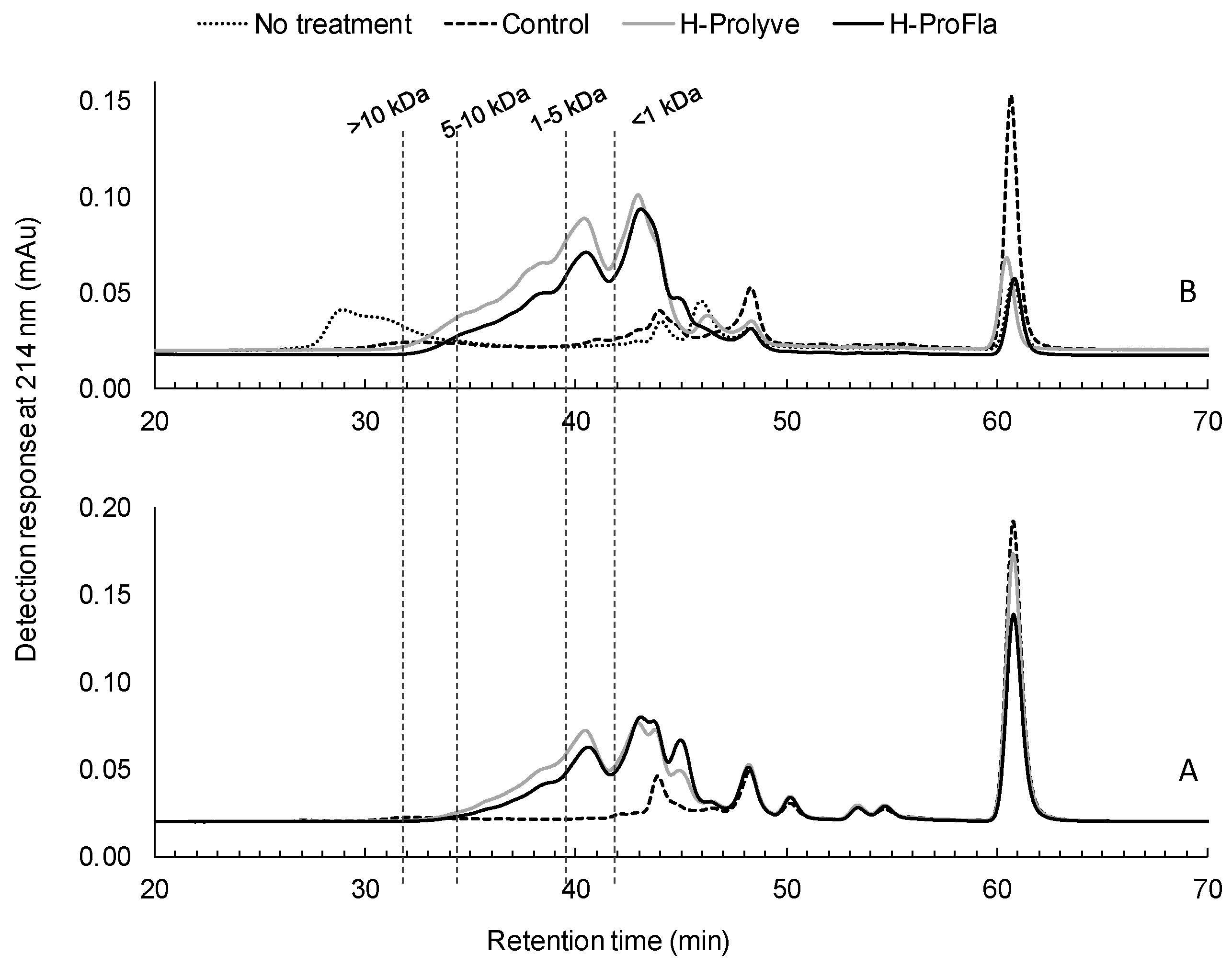
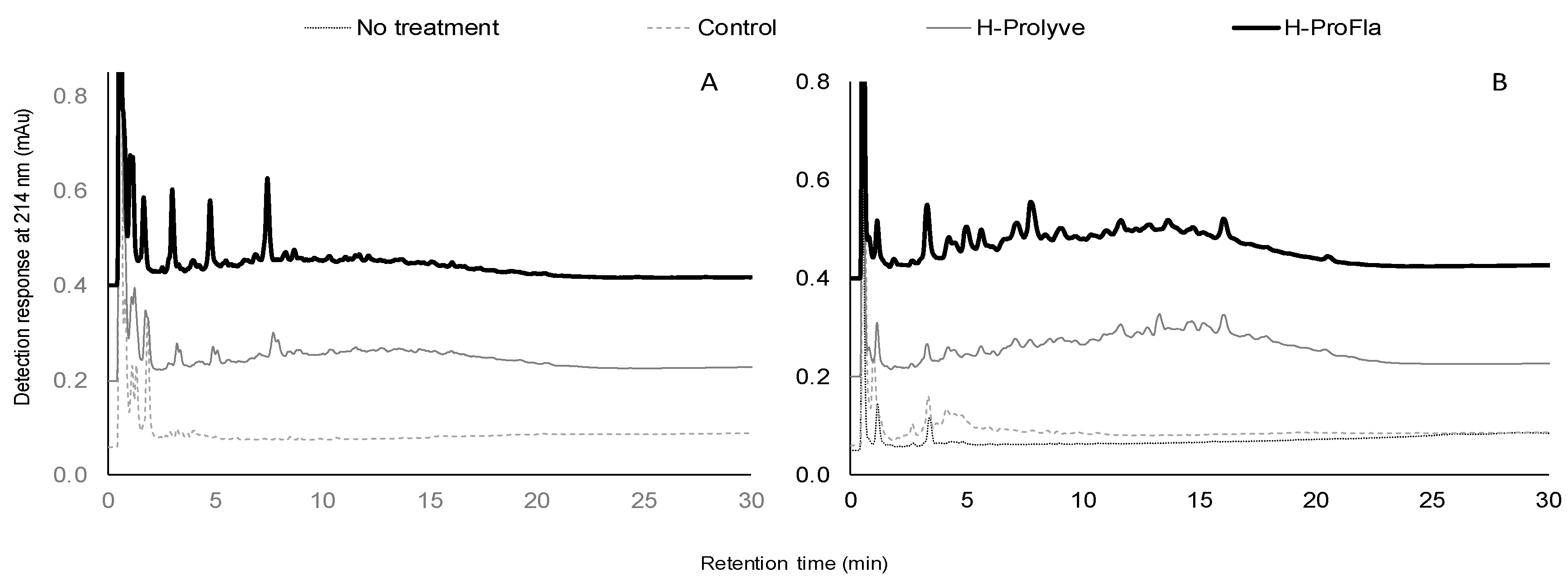
2.2. Physicochemical Characterization of Protein Hydrolysates
2.3. Antioxidant Activity
3. Materials and Methods
3.1. Reagents and Standards
3.2. Samples
3.2.1. Algal Biomass
3.2.2. Crude Protein Extracts
3.2.3. Determination of Nitrogenous Composition
3.2.4. Hydrolysate Generation
3.2.5. Characterization of the Hydrolysates
3.2.6. Preparation of Samples and Antioxidant Bioassays
3.2.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Barizão, E.O.; Visentainer, J.V.; de Cinque Almeida, V.; Ribeiro, D.; Chiste, R.C.; Fernandes, E. Citharexylum solanaceum fruit extracts: Profiles of phenolic compounds and carotenoids and their relation with ROS and RNS scavenging capacities. Food Res. Int. 2016, 86, 24–33. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Choe, E.; Min, D.B. Mechanisms of antioxidants in the oxidation of foods. Compr. Rev. Food Sci. 2009, 8, 345–358. [Google Scholar] [CrossRef]
- Tkaczewska, J.; Bukowski, M.; Mak, P. Identification of Antioxidant Peptides in Enzymatic Hydrolysates of Carp (Cyprinus Carpio) Skin Gelatin. Molecules 2019, 24, 97. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Kristinsson, H.G.; Hreggvidsson, G.O.; Jónsson, J.Ó.; Thorkelsson, G.; Ólafsdóttir, G. Enzyme-enhanced extraction of antioxidant ingredients from red algae Palmaria palmata. LWT Food Sci. Technol. 2010, 43, 1387–1393. [Google Scholar] [CrossRef]
- Wu, R.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H.; Li, H. Overview of antioxidant peptides derived from marine resources: The sources, characteristic, purification, and evaluation methods. Appl. Biochem. Biotechnol. 2015, 176, 1815–1833. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; FitzGerald, R. Antioxidative peptides: Enzymatic production, In vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Nasri, M. Protein hydrolysates and biopeptides: Production, biological activities, and applications in foods and health benefits. A review. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 81, pp. 109–159. [Google Scholar]
- Freitas, A.; Andrade, J.; Silva, F.; Rocha-Santos, T.; Duarte, A.; Gomes, A. Antioxidative peptides: Trends and perspectives for future research. Curr. Med. Chem. 2013, 20, 4575–4594. [Google Scholar] [CrossRef]
- Merz, M.; Eisele, T.; Berends, P.; Appel, D.; Rabe, S.; Blank, I.; Stressler, T.; Fischer, L. Flavourzyme, an enzyme preparation with industrial relevance: Automated nine-step purification and partial characterization of eight enzymes. J. Agric. Food Chem. 2015, 63, 5682–5693. [Google Scholar] [CrossRef]
- Lafarga, T.; Acién-Fernández, F.G.; Garcia-Vaquero, M. Bioactive peptides and carbohydrates from seaweed for food applications: Natural occurrence, isolation, purification, and identification. Algal Res. 2020, 48, 101909. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. The Global Status of Seaweed Production, Trade and Utilization, Globefish Research Programme; FAO: Rome, Italy, 2018. [Google Scholar]
- Fleurence, J.; Morançais, M.; Dumay, J. Seaweed Proteins. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 245–262. [Google Scholar]
- Yao, X. Preparation of the bioactive peptides from Porphyra yezoensis and their hydroxyl radical scavenging activity. Agro. Food Ind. Hi-Tech. 2013, 24, 41–43. [Google Scholar]
- Cian, R.E.; Martínez-Augustin, O.; Drago, S.R. Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byproducts of Porphyra columbina. Food Res. Int. 2012, 49, 364–372. [Google Scholar] [CrossRef]
- Cian, R.E.; Alaiz, M.; Vioque, J.; Drago, S.R. Enzyme proteolysis enhanced extraction of ACE inhibitory and antioxidant compounds (peptides and polyphenols) from Porphyra columbina residual cake. J. Appl. Phycol. 2013, 25, 1197–1206. [Google Scholar] [CrossRef]
- Bhatia, S.; Sharma, K.; Sharma, A.; Namdeo, A.G.; Chaugule, B.B. Anti-oxidant potential of Indian Porphyra. Pharmacologyonline 2011, 1, 248–257. [Google Scholar]
- Cermeño, M.; Stack, J.; Tobin, P.R.; O’Keeffe, M.B.; Harnedy, P.A.; Stengel, D.B.; FitzGerald, R.J. Peptide identification from a Porphyra dioica protein hydrolysate with antioxidant, angiotensin converting enzyme and dipeptidyl peptidase IV inhibitory activities. Food Funct. 2019, 10, 3421–3429. [Google Scholar] [CrossRef]
- Senevirathne, M.; Ahn, C.B.; Je, J.Y. Enzymatic extracts from edible red algae, Porphyra tenera, and their antioxidant, anti-acetylcholinesterase, and anti-inflammatory activities. Food Sci. Biotechnol. 2010, 19, 1551–1557. [Google Scholar] [CrossRef]
- Hwang, E.S.; Thi, N.D. Effects of extraction and processing methods on antioxidant compound contents and radical scavenging activities of Laver (Porphyra tenera). Prev. Nutr. Food Sci. 2014, 19, 40. [Google Scholar] [CrossRef]
- Hwang, E.; Choi, M. Antioxidant activity of commercially processed laver (Porphyra tenera). Proceed. Nutr. Soc. 2013, 72, OCE4. [Google Scholar] [CrossRef]
- Pimentel, F.; Alves, R.; Harnedy, P.; FitzGerald, R.; Oliveira, M. Macroalgal-derived protein hydrolysates and bioactive peptides: Enzymatic release and potential health enhancing properties. Trends Food. Sci. Tech. 2019, 93, 106–124. [Google Scholar] [CrossRef]
- Stack, J.; Tobin, P.R.; Gietl, A.; Harnedy, P.A.; Stengel, D.B.; FitzGerald, R.J. Seasonal variation in nitrogenous components and bioactivity of protein hydrolysates from Porphyra dioica. J. Appl. Phycol. 2017, 29, 2439–2450. [Google Scholar] [CrossRef]
- Harnedy, P.; FitzGerald, R. In vitro assessment of the cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Beaulieu, L.; Sirois, M.; Tamigneaux, É. Evaluation of the In vitro biological activity of protein hydrolysates of the edible red alga, Palmaria palmata (dulse) harvested from the Gaspe coast and cultivated in tanks. J. Appl. Phycol. 2016, 28, 3101–3115. [Google Scholar] [CrossRef]
- Pimentel, F.B.; Cermeño, M.; Kleekayai, T.; Harnedy, P.A.; FitzGerald, R.J.; Alves, R.C.; Oliveira, M.B.P.P. Effect of In vitro simulated gastrointestinal digestion on the antioxidant activity of the red seaweed Porphyra dioica. Food Res. Int. 2020, 136, 109309. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Extraction of protein from the macroalga Palmaria palmata. LWT- Food Sci. Technol. 2013, 51, 375–382. [Google Scholar] [CrossRef]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2016, 28, 511–524. [Google Scholar] [CrossRef]
- Spellman, D.; O’cuinn, G.; FitzGerald, R. Bitterness in Bacillus proteinase hydrolysates of whey proteins. Food Chem. 2009, 114, 440–446. [Google Scholar] [CrossRef]
- Sinthusamran, S.; Idowu, A.T.; Benjakul, S.; Prodpran, T.; Yesilsu, A.F.; Kishimura, H. Effect of proteases and alcohols used for debittering on characteristics and antioxidative activity of protein hydrolysate from salmon frames. J. Food Sci. Tech. 2020, 57, 473–483. [Google Scholar] [CrossRef]
- Aryee, A.N.A.; Boye, J.I. Improving the Digestibility of Lentil Flours and Protein Isolate and Characterization of Their Enzymatically Prepared Hydrolysates. Int. J. Food Prop. 2016, 19, 2649–2665. [Google Scholar] [CrossRef]
- Meisel, H.; Walsh, D.; Murray, B.; FitzGerald, R. ACE inhibitory peptides. In Nutraceutical Proteins and Peptides in Health and Disease, Mine, Y., Shahidi, F., Eds.; CRC Press: Boca Raton, FL, USA, 2005; Volume 4, pp. 269–315. [Google Scholar]
- Opitz, S.E.W.; Smrke, S.; Goodman, B.A.; Yeretzian, C. Methodology for the Measurement of Antioxidant Capacity of Coffee: A Validated Platform Composed of Three Complementary Antioxidant Assays. In Processing and Impact on Antioxidants in Beverages; Preedy, V., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 253–264. [Google Scholar]
- Spellman, D.; Kenny, P.; O’Cuinn, G.; FitzGerald, R.J. Aggregation properties of whey protein hydrolysates generated with Bacillus licheniformis proteinase activities. J. Agric. Food Chem. 2005, 53, 1258–1265. [Google Scholar] [CrossRef]
- Kalyankar, P.; Zhu, Y.; O’ Keeffe, M.; O’ Cuinn, G.; FitzGerald, R.J. Substrate specificity of glutamyl endopeptidase (GE): Hydrolysis studies with a bovine α-casein preparation. Food Chem. 2013, 136, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Matsui, R.; Honda, R.; Kanome, M.; Hagiwara, A.; Matsuda, Y.; Togitani, T.; Ikemoto, N.; Terashima, M. Designing antioxidant peptides based on the antioxidant properties of the amino acid side-chains. Food Chem. 2018, 245, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xiong, H.; Selomulya, C.; Chen, X.D.; Zhong, H.; Wang, S.; Sun, W.; Zhou, Q. Enzymatic hydrolysis of rice dreg protein: Effects of enzyme type on the functional properties and antioxidant activities of recovered proteins. Food Chem. 2012, 134, 1360–1367. [Google Scholar] [CrossRef]
- Song, L.; Li, T.; Yu, R.; Yan, C.; Ren, S.; Zhao, Y. Antioxidant activities of hydrolysates of Arca subcrenata prepared with three proteases. Marine Drugs 2008, 6, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Everette, J.D.; Bryant, Q.M.; Green, A.M.; Abbey, Y.A.; Wangila, G.W.; Walker, R.B. Thorough study of reactivity of various compound classes toward the Folin−Ciocalteu reagent. J. Agric. Food. Chem. 2010, 58, 8139–8144. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Park, P.-J.; Je, J.-Y. Preparation and biological evaluation of enzyme-assisted extracts from edible seaweed (Enteromorpha prolifera) as antioxidant, anti-acetylcholinesterase and inhibition of lipopolysaccharide-induced nitric oxide production in murine macrophages. Int. J. Food Sci. Nutr. 2012, 63, 187–193. [Google Scholar] [CrossRef]
- Ahn, C.-B.; Jeon, Y.-J.; Kang, D.-S.; Shin, T.-S.; Jung, B.-M. Free radical scavenging activity of enzymatic extracts from a brown seaweed Scytosiphon lomentaria by electron spin resonance spectrometry. Food Res. Int. 2004, 37, 253–258. [Google Scholar] [CrossRef]
- Connolly, A.; Piggott, C.O.; FitzGerald, R.J. Characterisation of protein-rich isolates and antioxidative phenolic extracts from pale and black brewers’ spent grain. Int. J. Food Sci. Tech. 2013, 48, 1670–1681. [Google Scholar] [CrossRef]
- Le Maux, S.; Nongonierma, A.B.; Barre, C.; FitzGerald, R.J. Enzymatic generation of whey protein hydrolysates under pH-controlled and non pH-controlled conditions: Impact on physicochemical and bioactive properties. Food Chem. 2016, 199, 246–251. [Google Scholar] [CrossRef]
- Nunes, M.A.; Costa, A.S.; Bessada, S.; Santos, J.; Puga, H.; Alves, R.C.; Freitas, V.; Oliveira, M.B.P. Olive pomace as a valuable source of bioactive compounds: A study regarding its lipid-and water-soluble components. Sci. Total Environ. 2018, 644, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Wang, K.; Lei, Y.; Yao, S.; Lu, B.; Huang, W. Antioxidant synergistic effects of Osmanthus fragrans flowers with green tea and their major contributed antioxidant compounds. Sci. Rep. 2017, 7, 46501. [Google Scholar] [CrossRef] [PubMed]
- Kleekayai, T.; Harnedy, P.A.; O’Keeffe, M.B.; Poyarkov, A.A.; CunhaNeves, A.; Suntornsuk, W.; FitzGerald, R.J. Extraction of antioxidant and ACE inhibitory peptides from Thai traditional fermented shrimp pastes. Food Chem. 2015, 176, 441–447. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | NPN | PN | TN | Protein* |
|---|---|---|---|---|
| (% dw) | (% dw) | (% dw) | (% dw) | |
| Blades | 0.99 ± 0.07 a | 3.69 ± 0.20 b | 4.60 ± 0.18 b | 23.09 ± 0.90 b |
| PI | 0.06 ± 0.001 b | 8.26 ± 0.27 a | 10.10 ± 0.30 a | 50.30 ± 1.52 a |
| Treatment | Blades (mg GAE/g FDS) | Protein isolate (mg GAE/g FDS) |
|---|---|---|
| No treatment (4 °C, 16 h) | 2.55 ± 0.47 a | na |
| Control | 2.25 ± 0.10 a | 1.89 ± 0.11 a |
| H-Prolyve | 3.19 ± 0.11 b | 3.96 ± 0.09 b |
| H-ProFla | 3.42 ± 0.08 b | 4.34 ± 0.12 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pimentel, F.B.; Cermeño, M.; Kleekayai, T.; Harnedy-Rothwell, P.A.; Fernandes, E.; Alves, R.C.; Oliveira, M.B.P.P.; FitzGerald, R.J. Enzymatic Modification of Porphyra dioica-Derived Proteins to Improve their Antioxidant Potential. Molecules 2020, 25, 2838. https://doi.org/10.3390/molecules25122838
Pimentel FB, Cermeño M, Kleekayai T, Harnedy-Rothwell PA, Fernandes E, Alves RC, Oliveira MBPP, FitzGerald RJ. Enzymatic Modification of Porphyra dioica-Derived Proteins to Improve their Antioxidant Potential. Molecules. 2020; 25(12):2838. https://doi.org/10.3390/molecules25122838
Chicago/Turabian StylePimentel, Filipa B., Maria Cermeño, Thanyaporn Kleekayai, Pádraigín A. Harnedy-Rothwell, Eduarda Fernandes, Rita C. Alves, M. Beatriz P.P. Oliveira, and Richard J. FitzGerald. 2020. "Enzymatic Modification of Porphyra dioica-Derived Proteins to Improve their Antioxidant Potential" Molecules 25, no. 12: 2838. https://doi.org/10.3390/molecules25122838
APA StylePimentel, F. B., Cermeño, M., Kleekayai, T., Harnedy-Rothwell, P. A., Fernandes, E., Alves, R. C., Oliveira, M. B. P. P., & FitzGerald, R. J. (2020). Enzymatic Modification of Porphyra dioica-Derived Proteins to Improve their Antioxidant Potential. Molecules, 25(12), 2838. https://doi.org/10.3390/molecules25122838