Effect of Stereochemical Configuration on the Transport and Metabolism of Catechins from Green Tea across Caco-2 Monolayers

Abstract
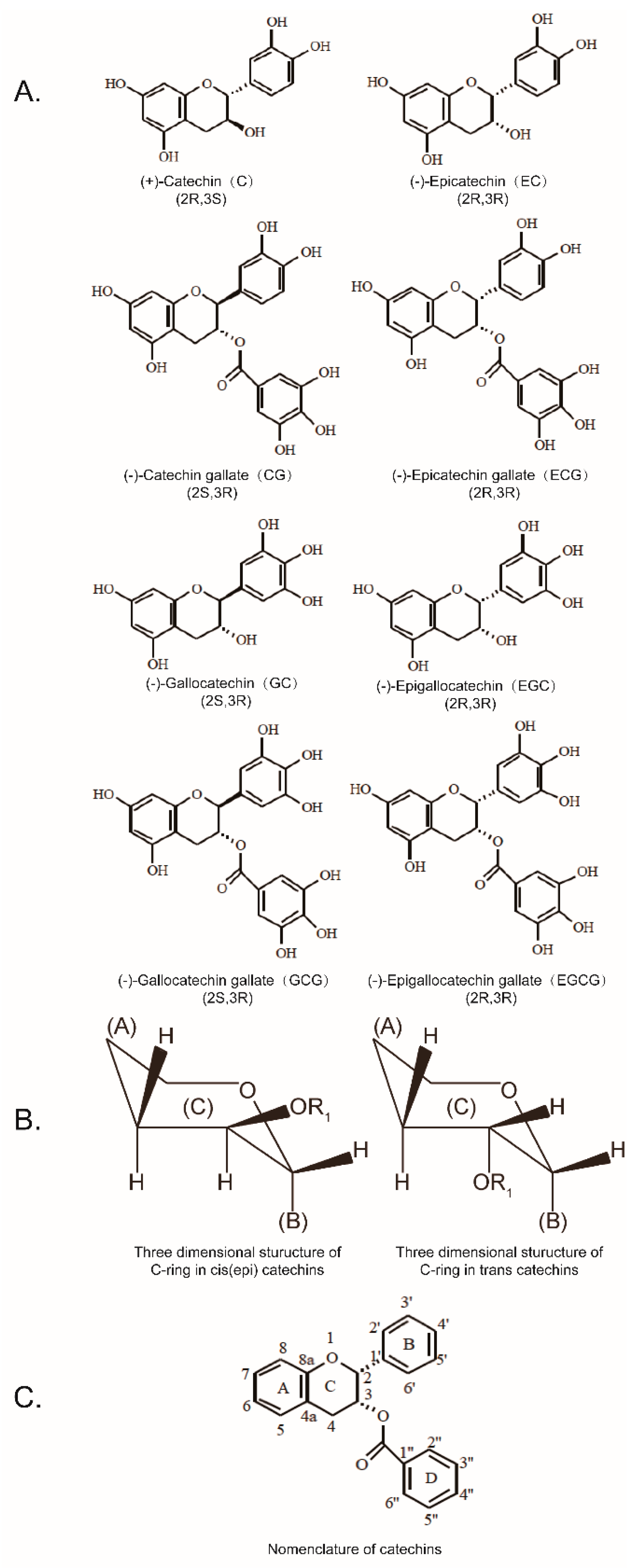
1. Introduction
2. Results and Discussion
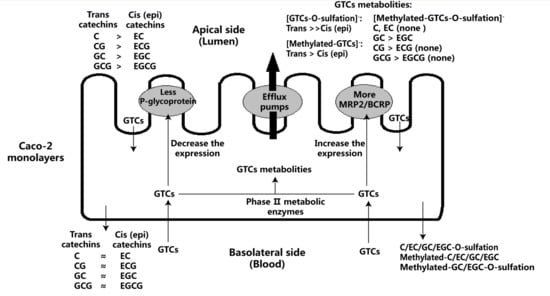
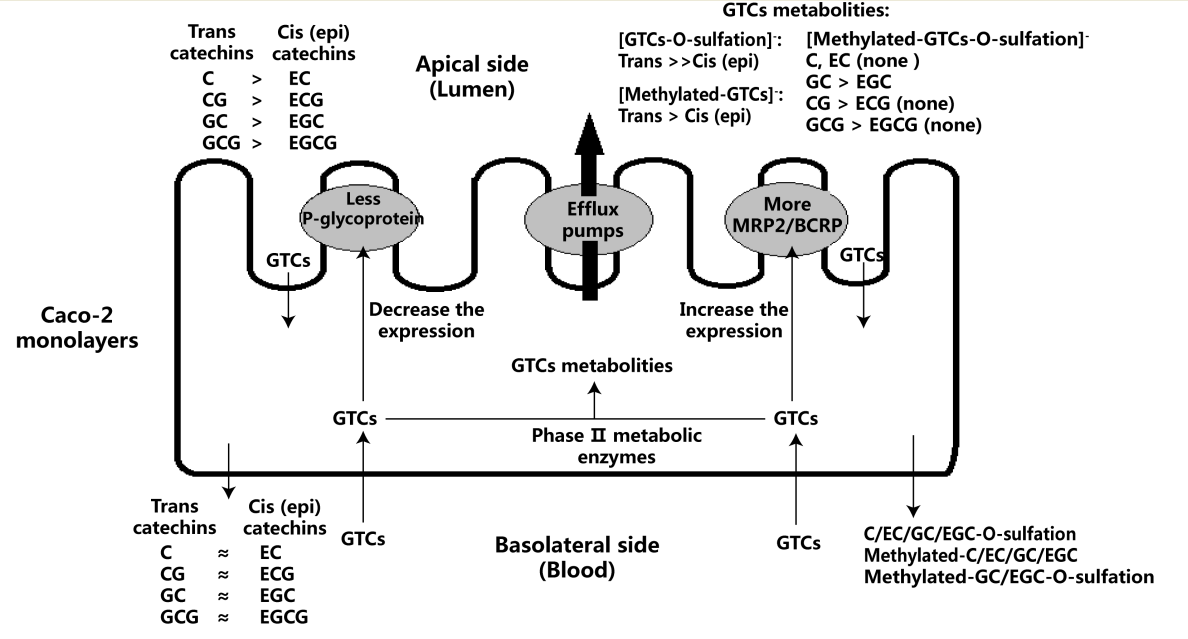
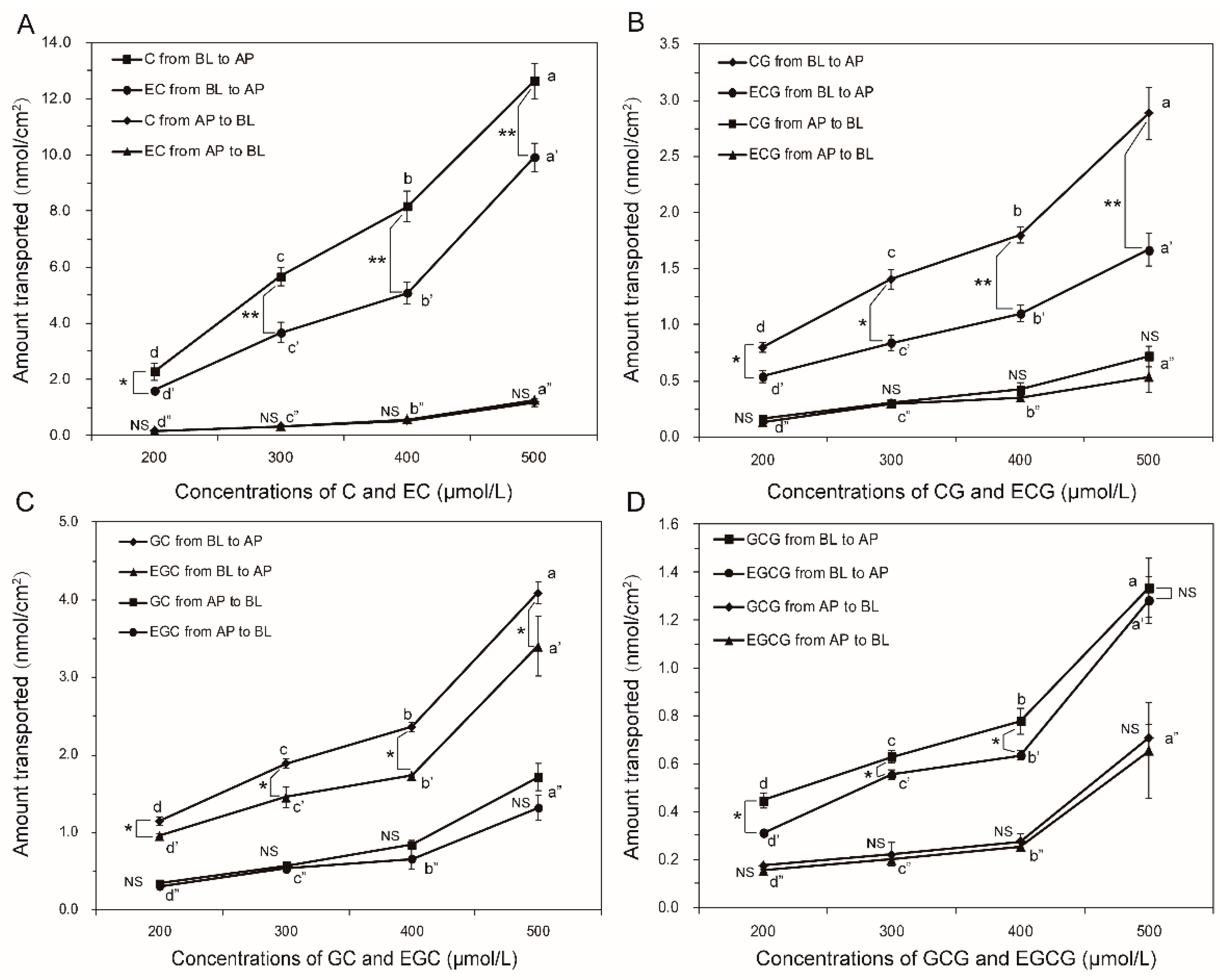
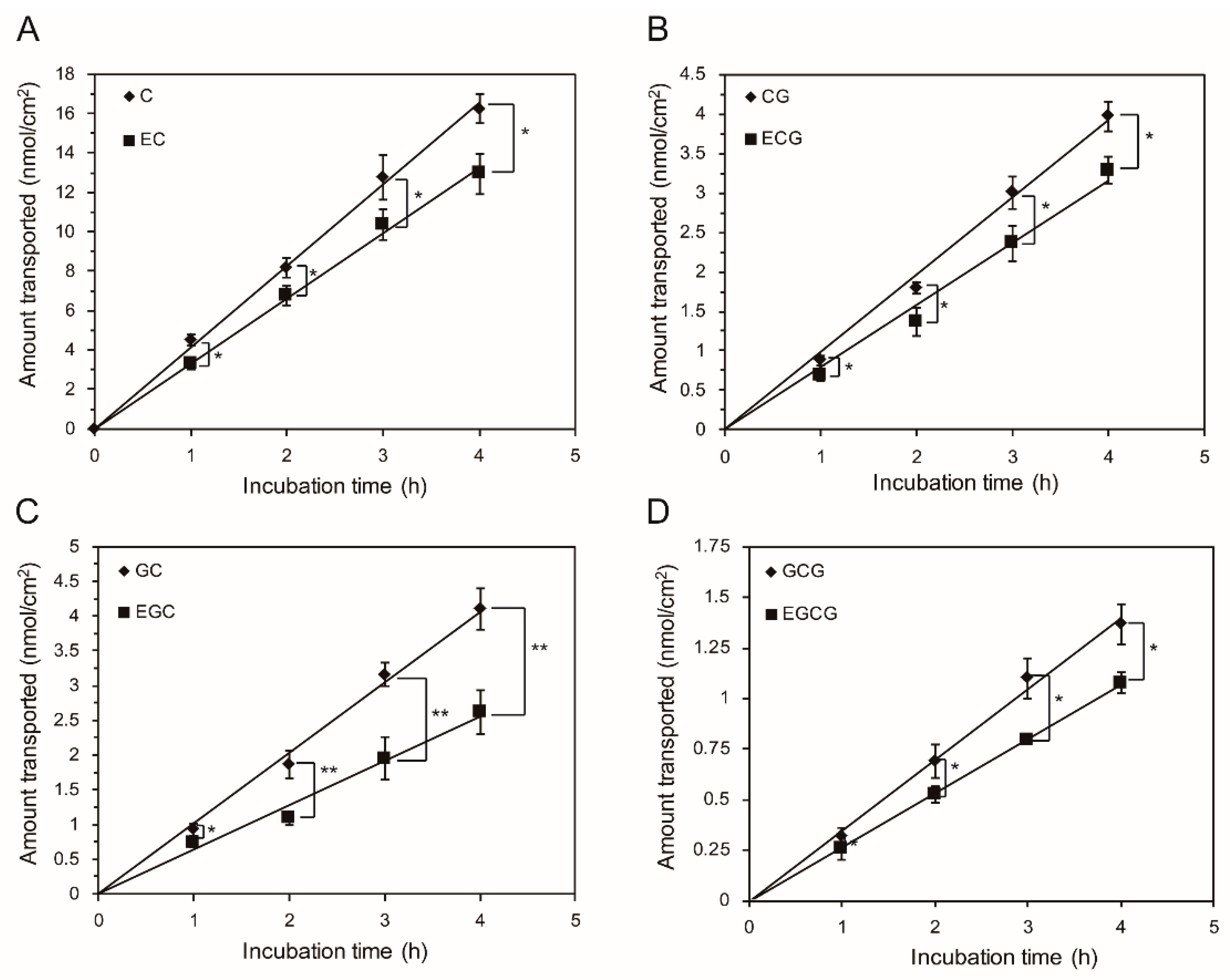
2.1. Bidirectional Transport of Eight GTCs
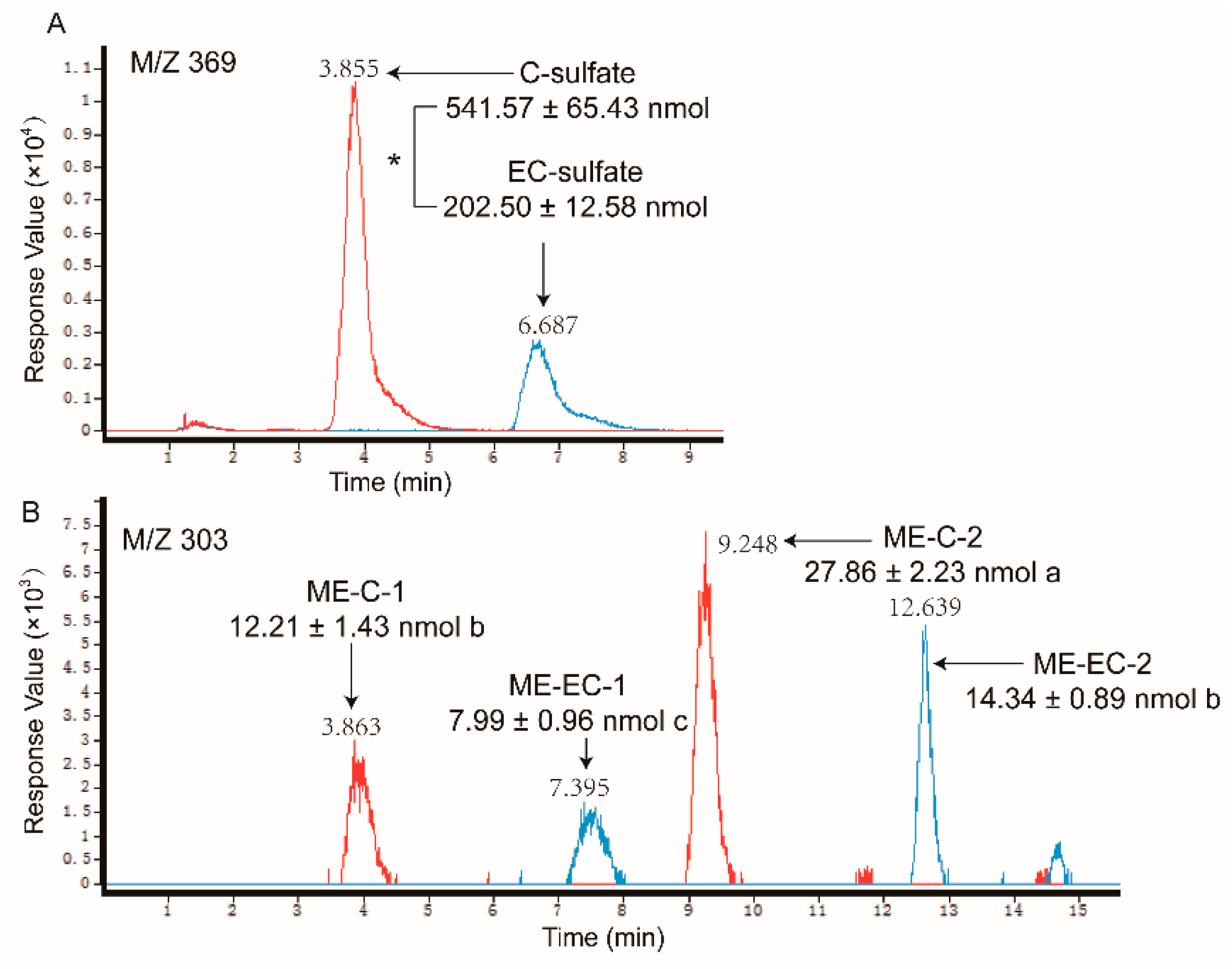
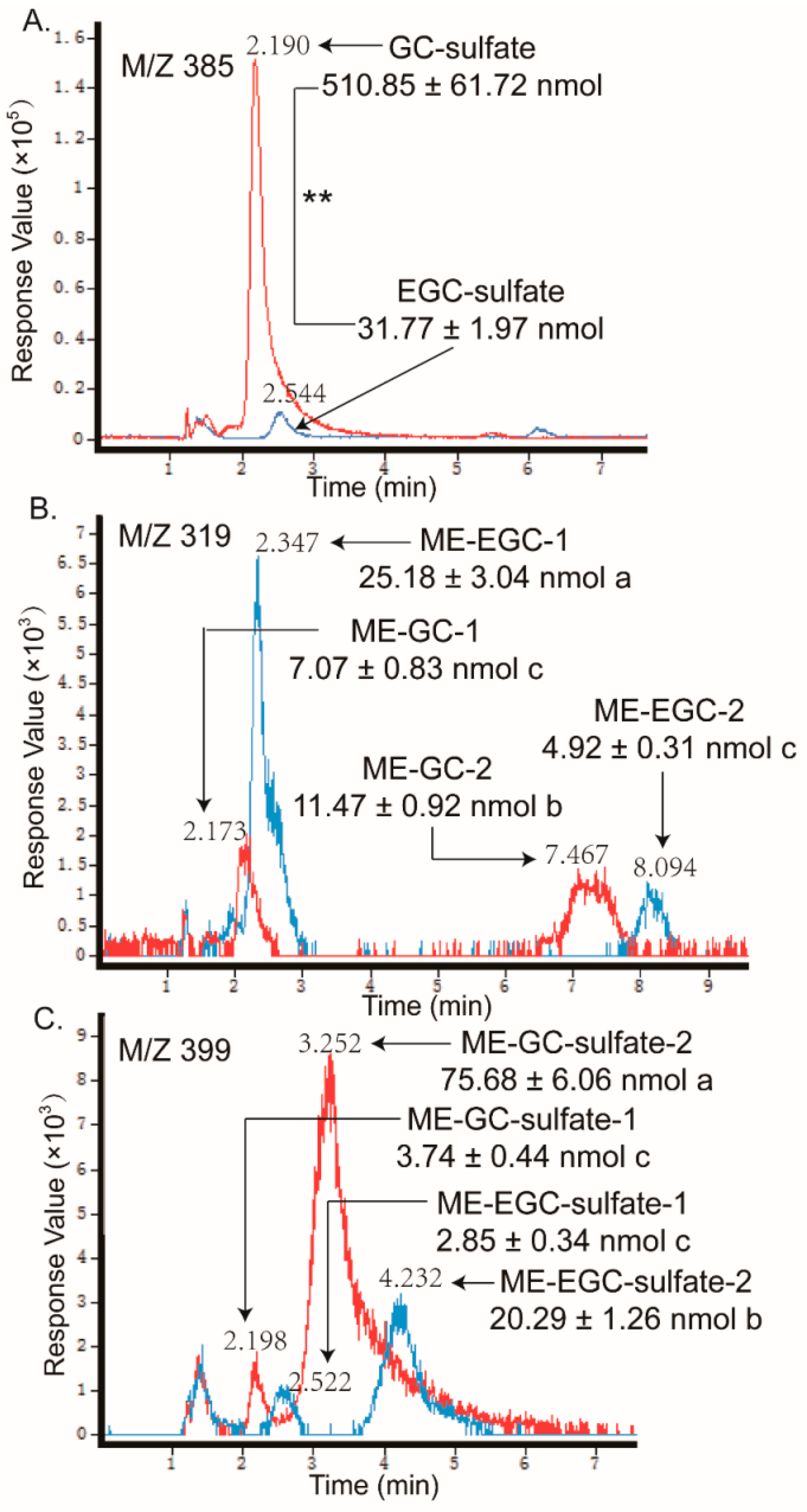
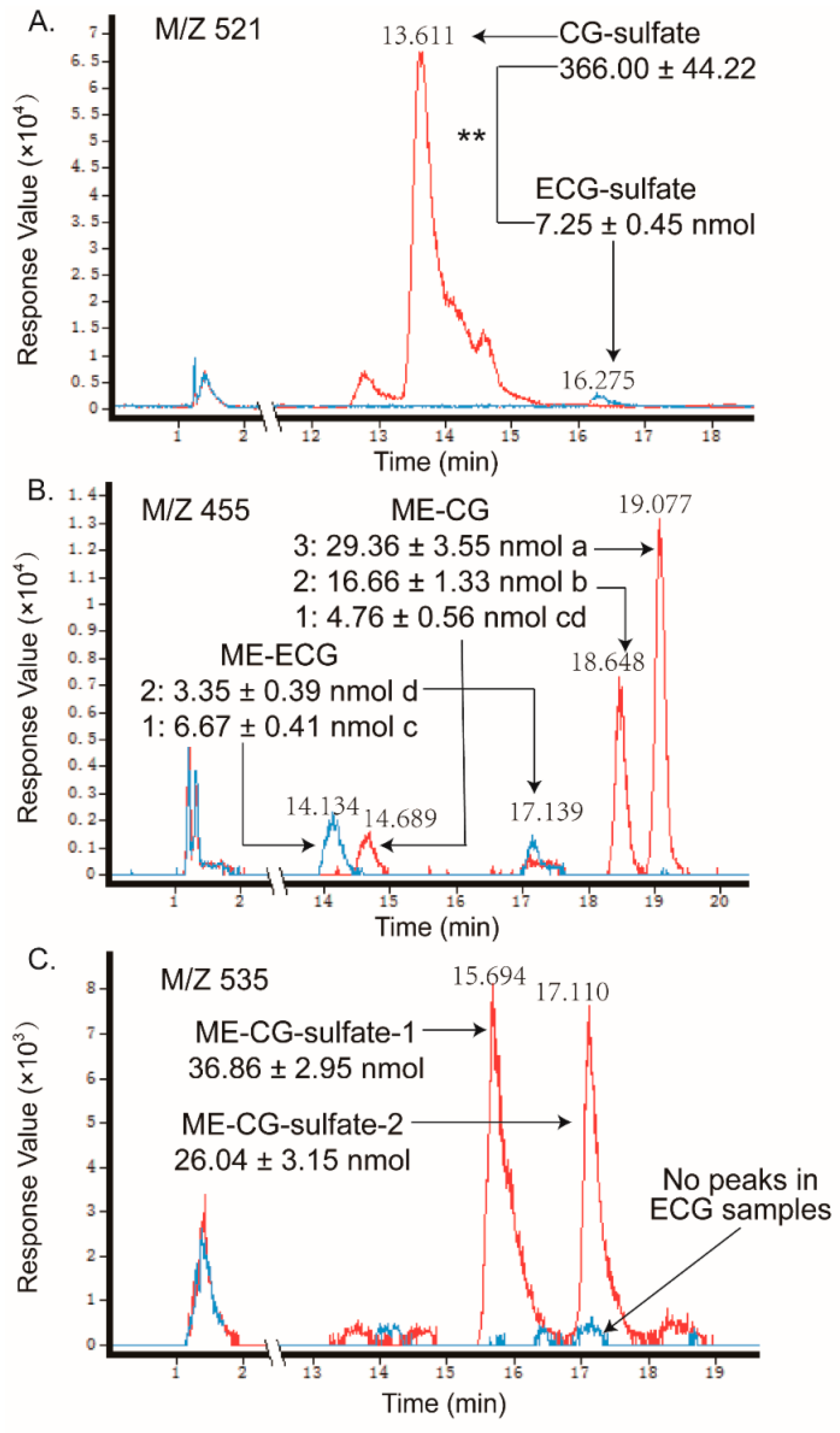
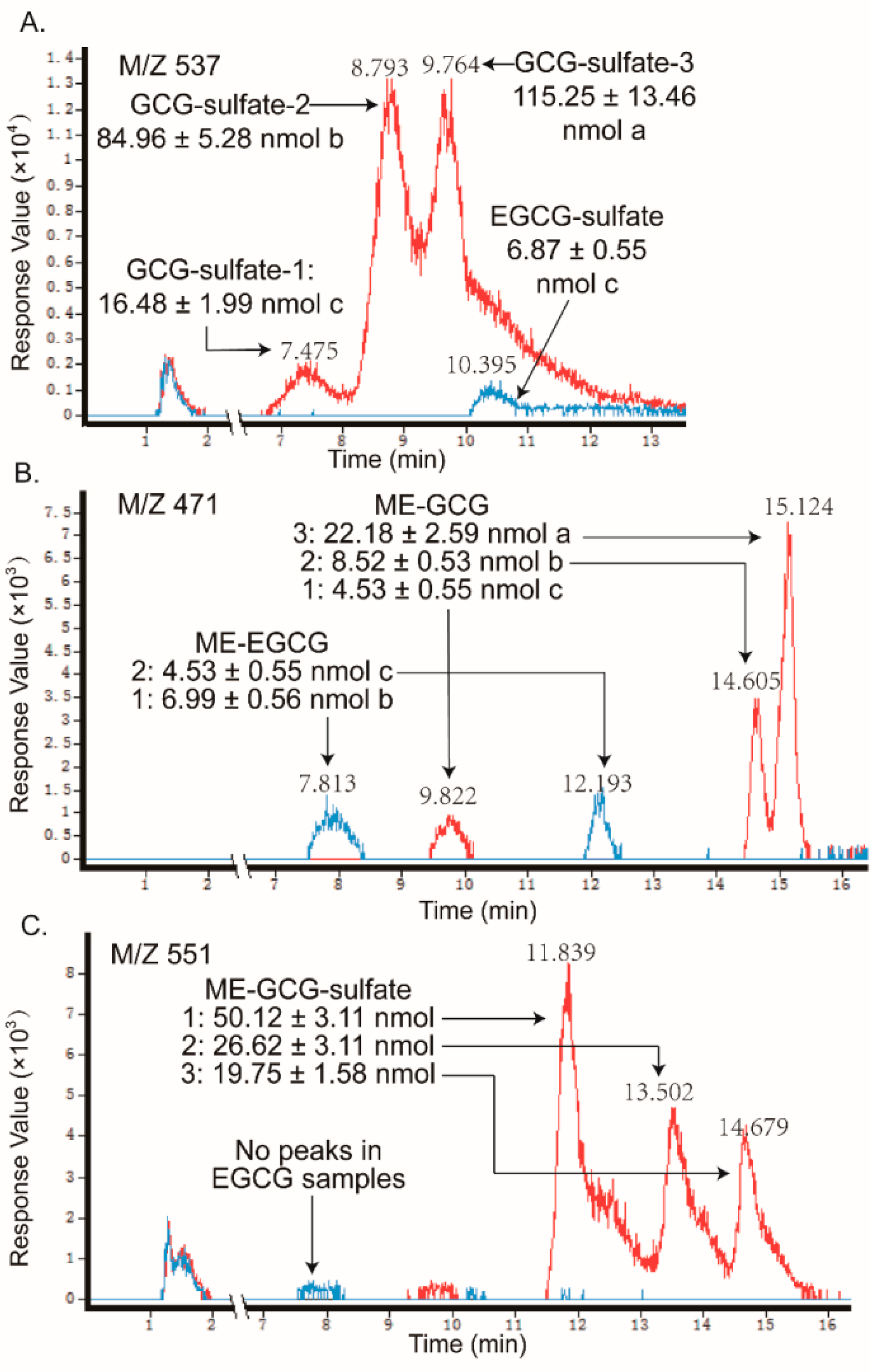
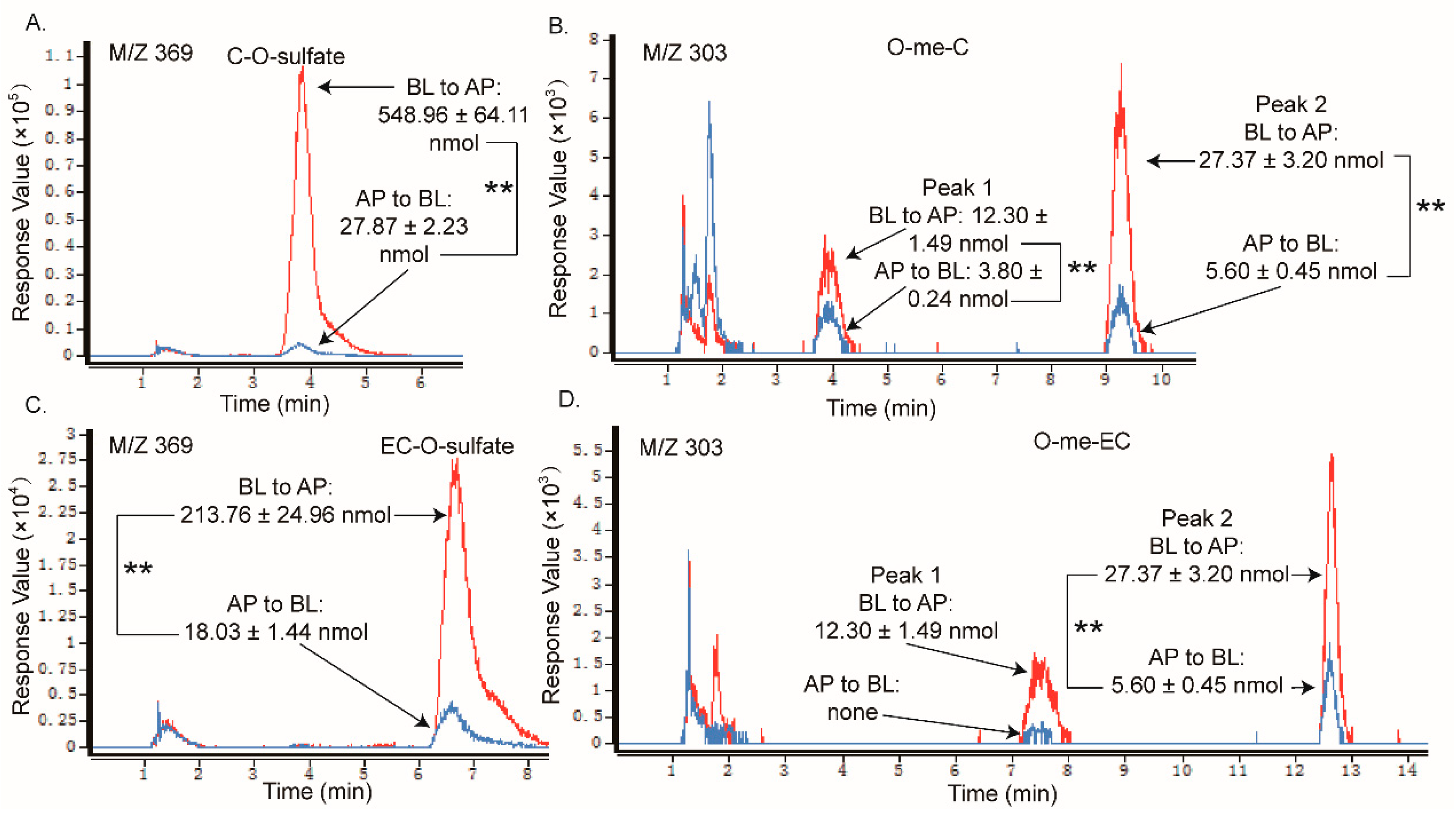
2.2. Metabolites of Eight GTCs
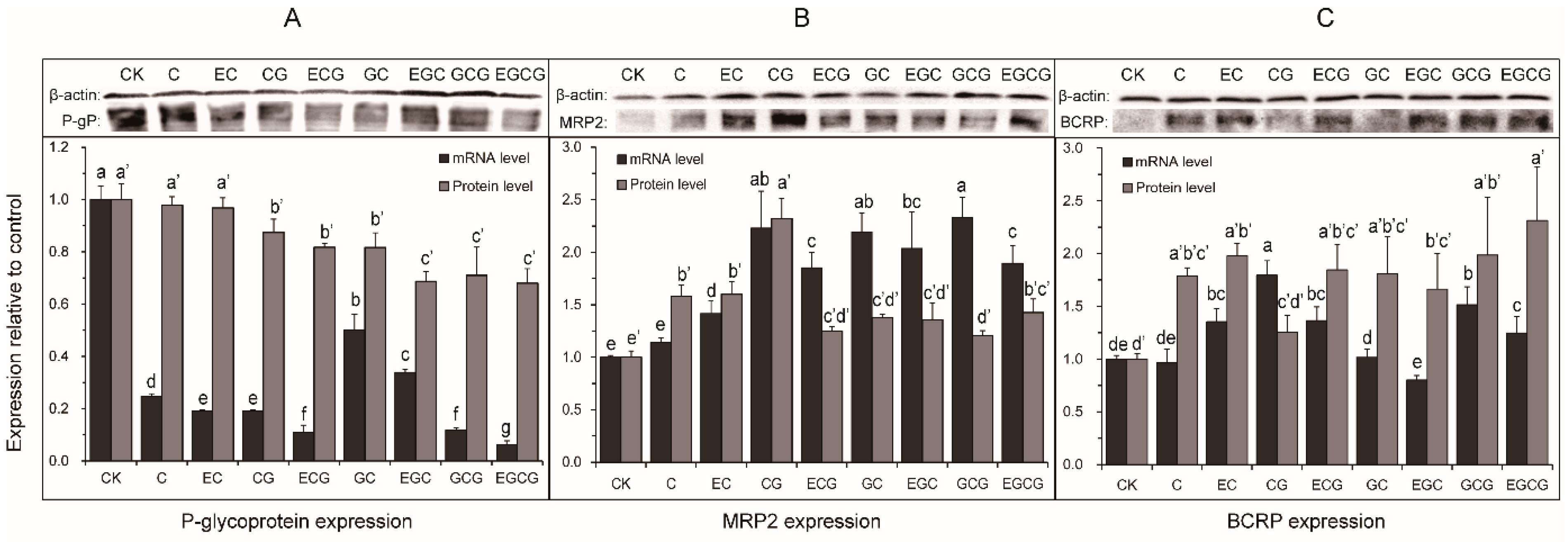
2.3. Effect of Eight GTCs on the Expression of MRP2, BCRP, and P-glycoprotein in Caco-2 Cells
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Cell Culture
3.2.2. Detection of Caco-2 Viability
3.2.3. Transepithelial Transport Experiments across Caco-2 Monolayer
3.2.4. HPLC Analysis
3.2.5. LC/MS Analysis
3.2.6. Quantitative Real Time RT-PCR
3.2.7. Western Blot
3.2.8. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yamamoto, T.; Juneja, L.R.; Chu, S.C. Chemistry and Applications of Green Tea; CRC Press: New York, NY, USA, 1997; Chapter 1. [Google Scholar]
- Huang, Y.; Xu, R.; Song, B.; Yang, S.; Zhao, L.; Wu, S. Effects of (−)-Epigallocatechin gallate on some protein factors involved in the epidermal growth factor receptor signaling pathway. J. Nanjing Med. Univ. 2009, 23, 293–299. [Google Scholar] [CrossRef]
- Huvaere, K.; Nielsen, J.H.; Bakman, M.; Hammershøj, M.; Skibsted, L.H.; Sørensen, J.; Vognsen, L.; Dalsgaard, T.K. Antioxidant properties of green tea extract protect reduced fat soft cheese against oxidation induced by light exposure. J. Agric. Food Chem. 2011, 59, 8718–8723. [Google Scholar] [CrossRef]
- Ju, J.; Lu, G.; Lambert, J.D.; Yang, C.S. Inhibition of carcinogenesis by tea constituents. Semin. Cancer Biol. 2007, 17, 395–402. [Google Scholar] [CrossRef]
- Masuda, M.; Suzui, M.; Lim, J.T.; Deguchi, A.; Soh, J.W.; Weinstein, I.B. Epigallocatechin-3-gallate decreases VEGF production in head and neck and breast carcinoma cells by inhibiting EGFR-related pathways of signal transduction. J. Exp. Ther. Oncol. 2002, 2, 350–359. [Google Scholar] [CrossRef]
- Wan, S.B.; Chen, D.; Ping Dou, Q.; Hang Chan, T. Study of the green tea polyphenols catechin-3-gallate (CG) and epicatechin-3-gallate (ECG) as proteasome inhibitors. Bioorgan. Med. Chem. 2004, 12, 3521–3527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, Y.; Chow, M.S.S.; Zuo, Z. Investigation of intestinal absorption and disposition of green tea catechins by Caco-2 monolayer model. Int. J. Pharm. 2004, 287, 1–12. [Google Scholar] [CrossRef]
- Abrahamse, S.L.; Kloots, W.J.; van Amelsvoort, J.M.M. Absorption, distribution, and secretion of epicatechin and quercetin in the rat. Nutr. Res. 2005, 25, 305–317. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, Y.; Li, R.C. Oral absorption and bioavailability of tea catechins. Planta Med. 2000, 66, 444–447. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; van der Hooft, J.J.J.; Crozier, A. Human studies on the absorption, distribution, metabolism, and excretion of tea polyphenols. Am. J. Clin. Nutr. 2013, 98, 1619S–1630S. [Google Scholar] [CrossRef]
- Warden, B.A.; Smith, L.S.; Beecher, G.R.; Balentine, D.A.; Clevidence, B.A. Catechins are bioavailable in men and women drinking black tea throughout the day. J Nutr 2001, 131, 1731–1737. [Google Scholar] [CrossRef] [PubMed]
- Kida, K.; Suzuki, M.; Matsumoto, N.; Nanjo, F.; Hara, Y. Identification of biliary metabolites of (−)-epigallocatechin gallate in rats. J. Agric. Food Chem. 2000, 48, 4151–4155. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Meng, X.; Winnik, B.; Lee, M.J.; Lu, H.; Sheng, S.; Buckley, B.; Yang, C.S. Analysis of urinary metabolites of tea catechins by liquid chromatography/electrospray ionization mass spectrometry. Chem. Res. Toxicol. 2001, 14, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Borges, G.; Momma, T.Y.; Spencer, J.P.E.; Keen, C.L.; Crozier, A.; Schroeter, H. The metabolome of [2-(14) C] (−)-epicatechin in humans: Implications for the assessment of efficacy, safety, and mechanisms of action of polyphenolic bioactives. Sci. Rep. 2016, 6, 29034. [Google Scholar] [CrossRef] [PubMed]
- Daood, M.; Tsai, C.; Ahdab-Barmada, M.; Watchko, J.F. ABC transporter (P-gp/ABCB1, MRP1/ABCC1, BCRP/ABCG2) expression in the developing human CNS. Neuropediatrics 2008, 39, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Sever, R.; Brugge, J.S. Differential selectivity of efflux transporter inhibitors in Caco-2 and MDCK-MDR1 monolayers: A strategy to assess the interaction of a new chemical entity with P-gp, BCRP, and MRP2. J. Pharm. Sci. 2012, 101, 1888–1897. [Google Scholar]
- Hong, J.; Lambert, J.D.; Lee, S.H.; Sinko, P.J.; Yang, C.S. Involvement of multidrug resistance-associated proteins in regulating cellular levels of (−)-epigallocatechin-3-gallate and its methyl metabolites. Biochem. Biophys. Res. Commun. 2003, 310, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, J.B.; Walle, T. Cellular uptake and efflux of the tea flavonoid (−) Epicatechin-3-gallate in the human intestinal cell line Caco-2. J. Pharmacol. Exp. Ther. 2003, 307, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Meunier, V.; Bourrié, M.; Berger, Y.; Fabre, G. The human intestinal epithelial cell line Caco-2; Pharmacological and pharmacokinetic applications. Cell Biol. Toxicol. 1995, 11, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Hirohashi, T.; Suzuki, H.; Chu, X.Y.; Tamai, I.; Tsuji, A.; Sugiyama, Y. Function and expression of multidrug resistance-associated protein family in human colon adenocarcinoma cells (Caco-2). J. Pharmacol. Exp. Ther. 2000, 292, 265–270. [Google Scholar] [PubMed]
- Artursson, P.; Karlsson, J. Correlation between oral drug absorption in humans and apparent drug permeability coefficients in human intestinal epithelial (Caco-2) cells. Biochem. Biophys. Res. Commun. 1991, 175, 880–885. [Google Scholar] [CrossRef]
- Wan, Y.F. Metabolism of green tea catechins: An overview. Curr. Drug Metab. 2006, 7, 755–809. [Google Scholar]
- Chen, L.; Lee, M.J.; Li, H.; Yang, C.S. Absorption, distribution, elimination of tea polyphenols in rats. Drug Metab. Dispos. 1997, 25, 1045–1050. [Google Scholar] [PubMed]
- Sugihara, N.; Kuroda, N.; Watanabe, F.; Choshi, T.; Kamishikiryo, J.; Seo, M. Effects of catechins and their related compounds on cellular accumulation and efflux transport of mitoxantrone in Caco-2 cell monolayers. J. Food Sci. 2017, 82, 1224–1230. [Google Scholar] [CrossRef]
- Ottaviani, J.I.; Momma, Y.T.; Heiss, C.; Kwik-Uribe, C.; Schroeter, H.; Keen, C. The stereochemical configuration of flavanols influences the level and metabolism of flavanols in humans and their biological activity in vivo. Free Radic. Biol. Med. 2010, 50, 237–244. [Google Scholar] [CrossRef]
- Clarke, K.A.; Dew, T.P.; Watson, R.E.B.; Farrar, M.D.; Bennett, S.; Nicolaou, A.; Rhodes, L.E.; Williamson, G. High performance liquid chromatography tandem mass spectrometry dual extraction method for identification of green tea catechin metabolites excreted in human urine. J. Chromatogr. B 2014, 972, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Kuhnle, G.; Spencer, J.P.; Schroeter, H.; Shenoy, B.; Debnam, E.S.; Srai, S.K.; Rice-Evans, C.; Hahn, U. Epicatechin and catechin are O-methylated and glucuronidated in the small intestine. Biochem. Biophys. Res. Commun. 2000, 277, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, J.B.; Walle, T. Glucuronidation and sulfation of the tea flavonoid (-)-epicatechin by the human and rat enzymes. Drug Metab. Dispos. 2002, 30, 897–903. [Google Scholar] [CrossRef]
- Hirsova, P.; Karlasova, G.; Dolezelova, E.; Cermanova, J.; Zagorova, M.; Kadova, Z.; Hroch, M.; Sispera, L.; Tomsik, P.; Lenicek, M.; et al. Cholestatic effect of epigallocatechin gallate in rats is mediated via decreased expression of Mrp2. Toxicology 2013, 303, 9–15. [Google Scholar] [CrossRef]
- Baba, S. In vivo comparison of the bioavailability of (+)-catechin, (−)-epicatechin and their mixture in orally administered rats. J. Nutr. 2001, 131, 2885–2891. [Google Scholar] [CrossRef]
- Meng, X.; Lee, M.J.; Li, C.; Sheng, S.; Zhu, N.; Sang, S.; Ho, C.T.; Yang, C.S. Formation and identification of 4′-O-methyl-(−)-epigallocatechin in humans. Drug Metab. Dispos. 2001, 29, 789–793. [Google Scholar]
- Lu, H.; Meng, X.; Yang, C.S. Enzymology of methylation of tea catechins and inhibition of catechol-O-methyltransferase by (−)-epigallocatechin gallate. Drug Metab. Dispos. 2003, 31, 572–579. [Google Scholar] [CrossRef]
- Kitagawa, S.; Nabekura, T.; Kamiyama, S. Inhibition of P-glycoprotein function by tea catechins in KB-C2 cells. J. Pharm. Pharmacol. 2004, 56, 1001–1005. [Google Scholar] [CrossRef] [PubMed]
- Gerk, P.M.; Vore, M. Regulation of expression of the multidrug resistance-associated protein 2 (MRP2) and its role in drug disposition. J. Pharmacol. Exp. Ther. 2002, 302, 407–415. [Google Scholar] [CrossRef]
- Keppler, D.; König, J. Hepatic secretion of conjugated drugs and endogenous substances. Semin. Liver Dis. 2000, 20, 265–272. [Google Scholar] [CrossRef]
- Kast, H.R.; Goodwin, B.; Tarr, P.T.; Jones, S.A.; Anisfeld, A.M.; Stoltz, C.M.; Tontonoz, P.; Kliewer, S.; Willson, T.M.; Edwards, P.A. Regulation of multidrug resistance-associated protein 2 (ABCC2) by the nuclear receptors pregnane X receptor, farnesoid X-activated receptor, and constitutive androstane receptor. J. Biol. Chem. 2002, 277, 2908–2915. [Google Scholar] [CrossRef] [PubMed]
- Farabegoli, F.; Papi, A.; Bartolini, G.; Ostan, R.; Orlandi, M. (−)-Epigallocatechin-3-gallate downregulates Pg-P and BCRP in a tamoxifen resistant MCF-7 cell line. Phytomedicine 2010, 17, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Johnston, K.; Sharp, P.; Clifford, M.; Morgan, L. Dietary polyphenols decrease glucose uptake by human intestinal Caco-2 cells. FEBS Lett. 2005, 579, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Maubon, N.; Le Vee, M.; Fossati, L.; Audry, M.; Le Ferrec, E.; Bolze, S.; Fardel, O. Analysis of drug transporter expression in human intestinal Caco-2 cells by real-time PCR. Fundam. Clin. Pharm. 2007, 21, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, J.B.; Walle, T. Transport and metabolism of the tea flavonoid (−)-epicatechin by the human intestinal cell line Caco-2. Pharm. Res. 2001, 18, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds (−)-epicatechin (EC), (+)-catechin (C), (−)-epigallocatechin (EGC), (+)-gallocatechin (GC), (−)-epicatechin gallate (ECG), (−)-catechin gallate (CG), (−)-epigallocatechin gallate (EGCG), and (−)-gallocatechin gallate (GCG)are available or not from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transport in the Absence of MK571 (10−7 cm/s) | Efflux Ratio | Transport in the Presence of MK571 (10−7 cm/s) | Efflux Ratio | |||
|---|---|---|---|---|---|---|
| AP to BL | BL to AP | AP to BL | BL to AP | |||
| C | 1.56 ± 0.11 b | 26.18 ± 1.52 a | 16.74 | 3.97 ± 0.55 a,b,* | 5.33 ± 0.45 a,# | 1.34 |
| EC | 1.52 ± 0.17 b | 16.94 ± 1.63 b | 11.16 | 4.39 ± 0.45 a,* | 4.65 ± 0.53 b,# | 1.06 |
| CG | 1.39 ± 0.12 b | 6.51 ± 0.42 d | 4.68 | 2.85 ± 0.10 c,* | 3.85 ± 0.22 c,# | 1.35 |
| ECG | 1.38 ± 0.09 b | 3.87 ± 0.31 e | 2.82 | 2.63 ± 0.22 c,* | 2.72 ± 0.30 d,# | 1.04 |
| GC | 2.64 ± 0.21 a | 8.75 ± 0.28 c | 3.32 | 3.43 ± 0.08 b,c,* | 4.45 ± 0.46 b,# | 1.30 |
| EGC | 2.46 ± 0.10 a | 6.74 ± 0.62 d | 2.74 | 3.63 ± 0.35 b,* | 3.67 ± 0.13 c,# | 1.01 |
| GCG | 1.03 ± 0.21 c | 2.93 ± 0.13 e | 2.84 | 1.73 ± 0.16 d,* | 2.14 ± 0.16 e,# | 1.24 |
| EGCG | 0.93 ± 0.08 c | 2.57 ± 0.10 e | 2.75 | 1.89 ± 0.18 d,* | 1.98 ± 0.20 e,# | 1.05 |
| Compound Name | Detected Mass (ESI-) | MS/MS Fragments (Collision Energy) | Retention Time (min) | Relative Content (nmol) | Significance |
|---|---|---|---|---|---|
| C-O-sulfate | 369.0279 | 289, 231, 109 (20 V) | 3.86 | 541.57 ± 65.43 | * |
| EC-O-sulfate | 369.0285 | 289, 245,137 (20 V) | 6.69 | 202.50 ± 12.58 | |
| O-me-C | 303.0879 | 248, 203, 159, 101, 73 (1 0V) | 3.86, 9.25 | 41.07 ± 3.66 | * |
| O-me-EC | 303.089 | 248, 159, 73 (10 V) | 7.40, 12.64 | 22.33 ± 1.85 | |
| O-me-C-O-sulfate | ND | ND | ND | ND | |
| O-me-EC-O-sulfate | ND | ND | ND | ND | |
| GC-O-sulfate | 385.024 | 305, 261, 125 (20 V) | 2.19 | 510.85 ± 61.72 | ** |
| EGC-O-sulfate | 385.0246 | 305, 219, 154 (20 V) | 2.54 | 31.77 ± 1.97 | |
| O-me-GC | 319.0523 | 303, 194, 105 (10 V) | 2.17, 7.47 | 18.54 ± 1.74 | * |
| O-me-EGC | 319.0523 | 253, 183, 93 (10V) | 2.35, 8.10 | 30.10 ± 3.35 | |
| O-me-GC-O-sulfate | 399.0395 | 385, 288, 124 (10 V) | 2.20, 3.25 | 79.42 ± 6.50 | * |
| O-me-EGC-O-sulfate | 399.0404 | 319, 253, 183, 93 (10 V) | 2.52, 4.23 | 23.14 ± 1.60 | |
| CG-O-sulfate | 521.0416 | 441, 369, 289, 249, 169, 64 (20 V) | 13.61 | 366.00 ± 44.22 | ** |
| ECG-O-sulfate | 521.0406 | 441, 331, 289, 169, 125 (20 V) | 16.28 | 7.25 ± 0.45 | |
| O-me-CG | 455.0998 | 439, 382, 271, 169, 125, 58 (20 V) | 14.69, 18.65, 19.08 | 50.78 ± 5.44 | * |
| O-me-ECG | 455.0989 | 352, 183, 109 (20 V) | 14.13, 17.14 | 10.02 ± 0.80 | |
| O-me-CG-O-sulfate | 535.0567 | 455, 397, 303, 198, 141, 89 (10 V) | 15.69, 17.11 | 62.89 ± 6.10 | |
| O-me-ECG-O-sulfate | ND | ND | ND | ND | |
| GCG-O-sulfate | 537.0362 | 457, 395, 305, 216, 169, 125 (10 V) | 7.48, 8.79, 9.76 | 216.70 ± 20.73 | ** |
| EGCG-O-sulfate | 537.0376 | 457, 287, 169, 113 (10 V) | 10.4 | 6.87 ± 0.55 | |
| O-me-GCG | 471.0931 | 402, 287, 125 (10 V) | 9.82, 14.61, 15.12 | 35.23 ± 3.67 | * |
| O-me-EGCG | 471.0954 | 305, 113 (10 V) | 7.81, 12.19 | 11.51 ± 1.11 | |
| O-me-GCG-O-sulfate | 551.0513 | 464, 399, 369, 324, 205, 157, 77 (10 V) | 11.84, 13.50, 14.68 | 96.48 ± 7.80 | |
| O-me-EGCG-O-sulfate | ND | ND | ND | ND |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| P-glycoprotein/ABCB1 | GCCAAAGCCAAAATATCAGAC | TTCCAATGTGTTCGGCAT |
| MRP2/ABCC2 | TGAGCAAGTTTGAAACGCACAT | AGCTCTTCTCCTGCCGTCTCT |
| BCRP/ABCG2 | TGCAACATGTACTGGCGAAGA | TCTTCCACAAGCCCCAGG |
| β-actin | CAAGATCATTGCTCCTCCTGA | AGTCCGCCTAGAAGCATTTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ai, Z.; Liu, S.; Qu, F.; Zhang, H.; Chen, Y.; Ni, D. Effect of Stereochemical Configuration on the Transport and Metabolism of Catechins from Green Tea across Caco-2 Monolayers. Molecules 2019, 24, 1185. https://doi.org/10.3390/molecules24061185
Ai Z, Liu S, Qu F, Zhang H, Chen Y, Ni D. Effect of Stereochemical Configuration on the Transport and Metabolism of Catechins from Green Tea across Caco-2 Monolayers. Molecules. 2019; 24(6):1185. https://doi.org/10.3390/molecules24061185
Chicago/Turabian StyleAi, Zeyi, Shuyuan Liu, Fengfeng Qu, Haojie Zhang, Yuqiong Chen, and Dejiang Ni. 2019. "Effect of Stereochemical Configuration on the Transport and Metabolism of Catechins from Green Tea across Caco-2 Monolayers" Molecules 24, no. 6: 1185. https://doi.org/10.3390/molecules24061185
APA StyleAi, Z., Liu, S., Qu, F., Zhang, H., Chen, Y., & Ni, D. (2019). Effect of Stereochemical Configuration on the Transport and Metabolism of Catechins from Green Tea across Caco-2 Monolayers. Molecules, 24(6), 1185. https://doi.org/10.3390/molecules24061185