
Differential Cytotoxic Potential of Acridocarpus orientalis Leaf and Stem Extracts with the Ability to Induce Multiple Cell Death Pathways

 ,
,  , ,
, , 

Abstract
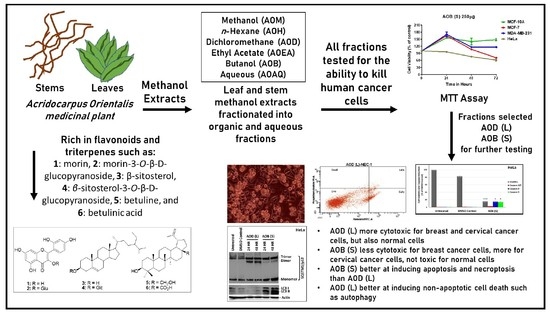
1. Introduction
2. Results
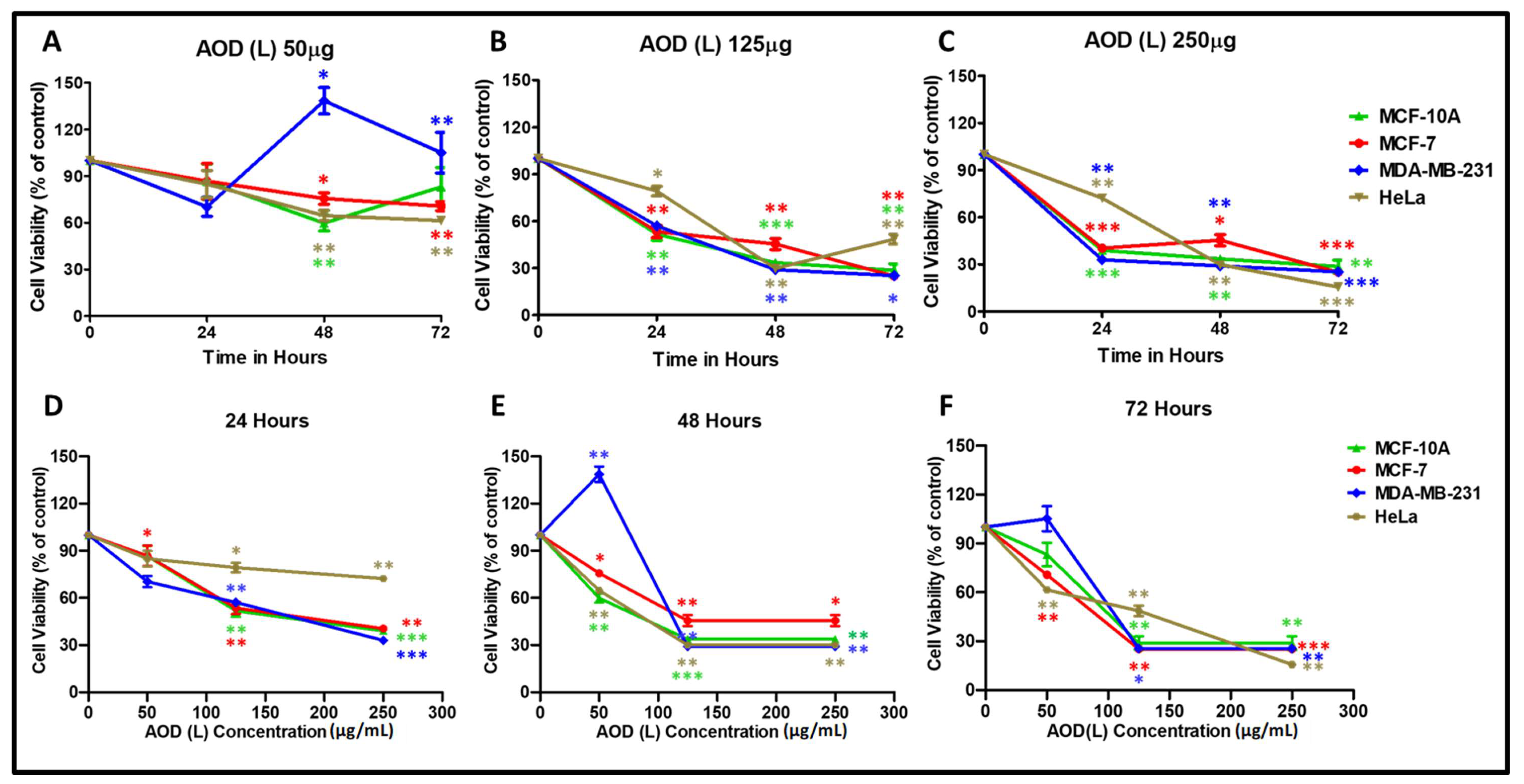
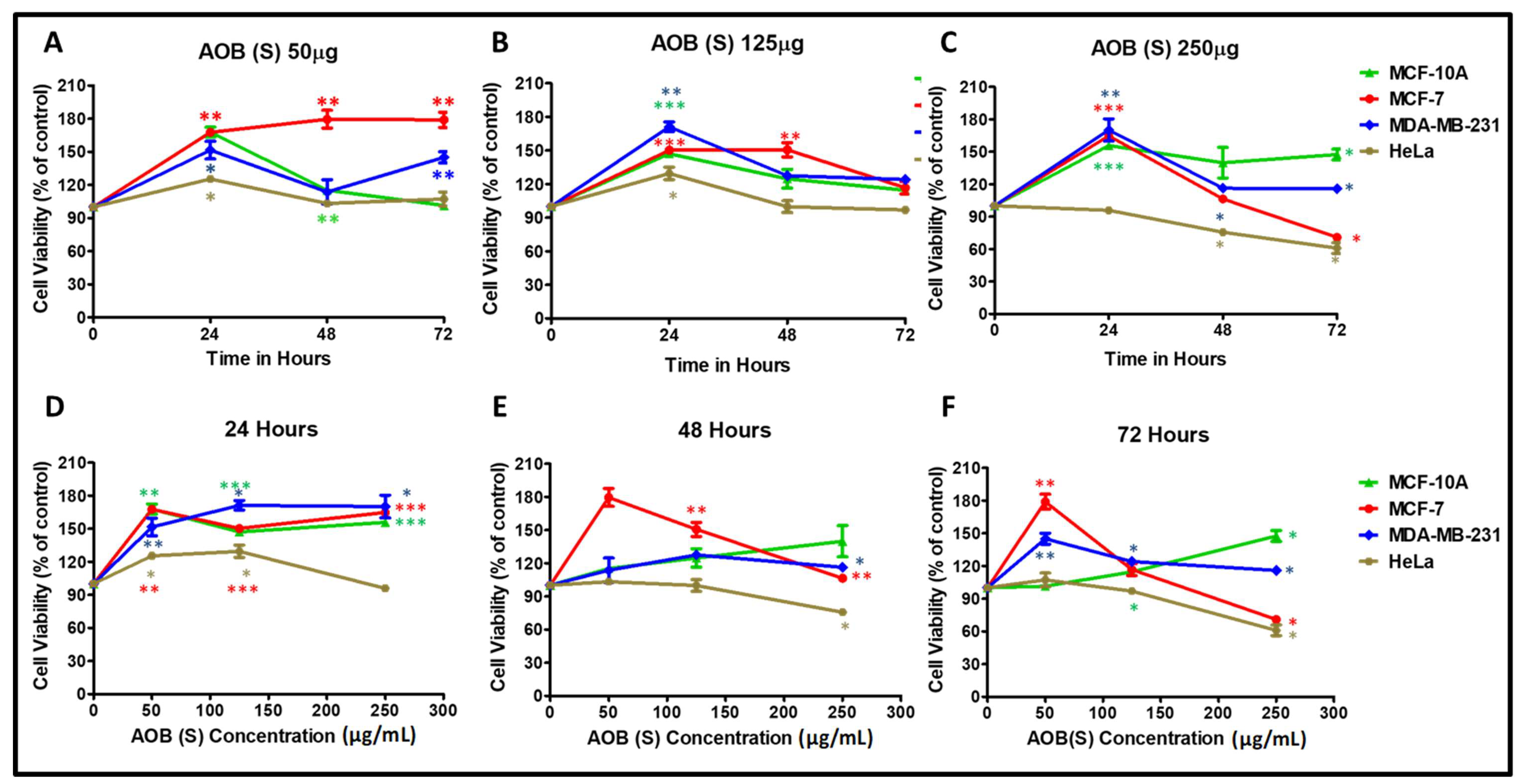
2.1. Effect of Different Leaf (L) and Stem (S) Crude Fractions of A. Orientalis on Cancer Cell Proliferation
2.2. AOD (L) and AOB (S) Reveal Different Anticancer Potentials
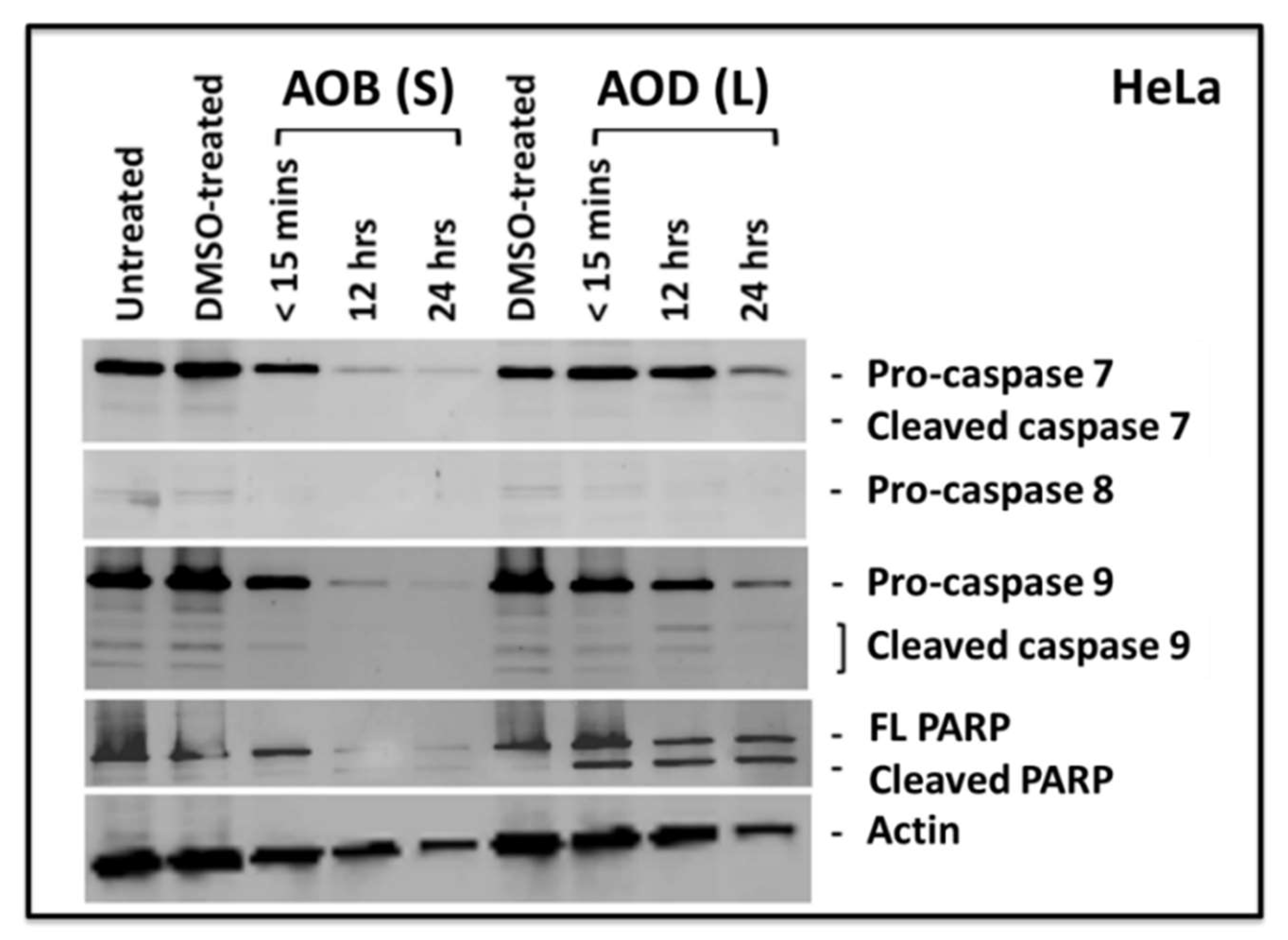
2.3. AOB (S) can Activate Significant Caspase Activity in HeLa Cells
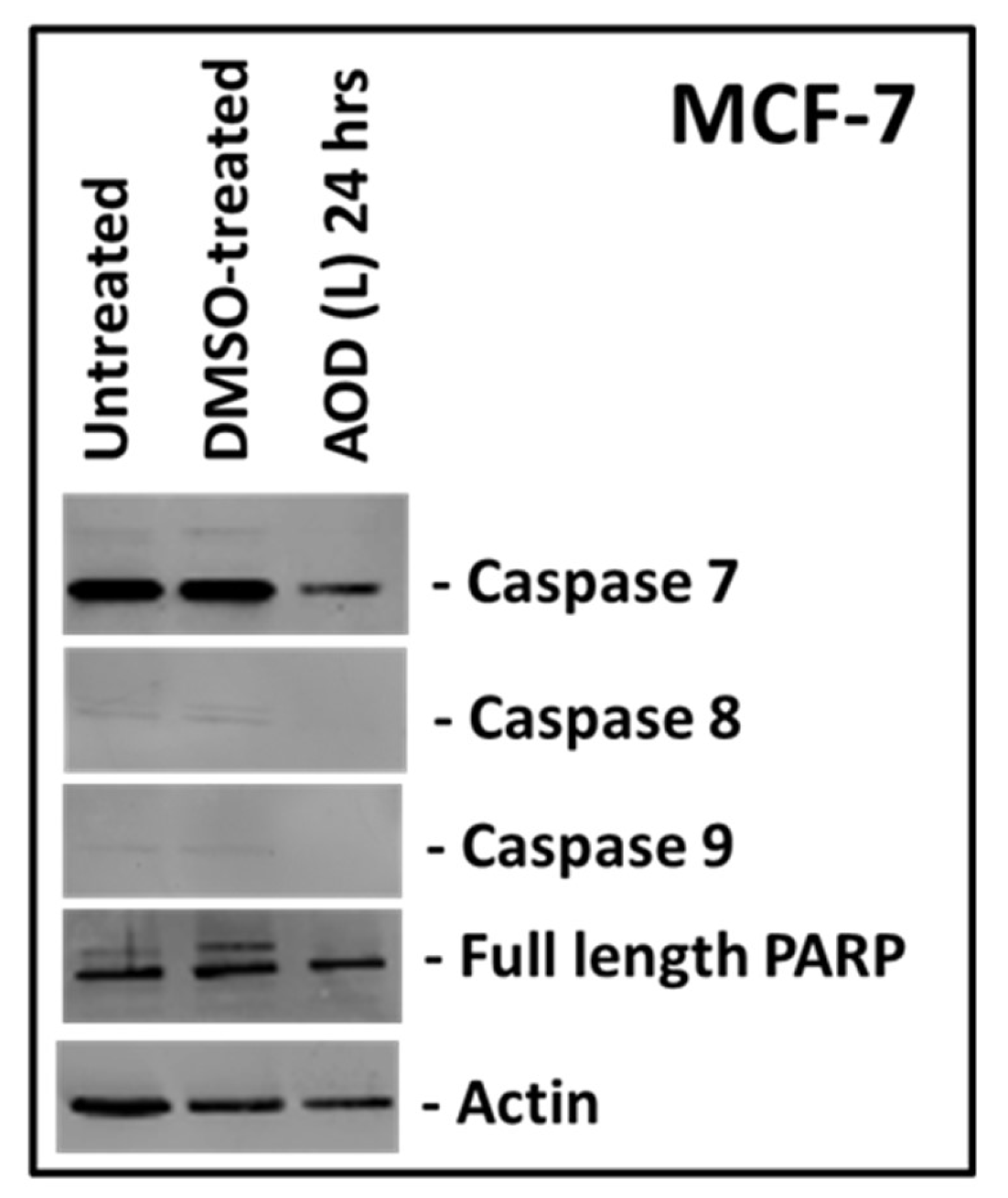
2.4. Both AOD (L) and AOB (S) can Lead to PARP Cleavage
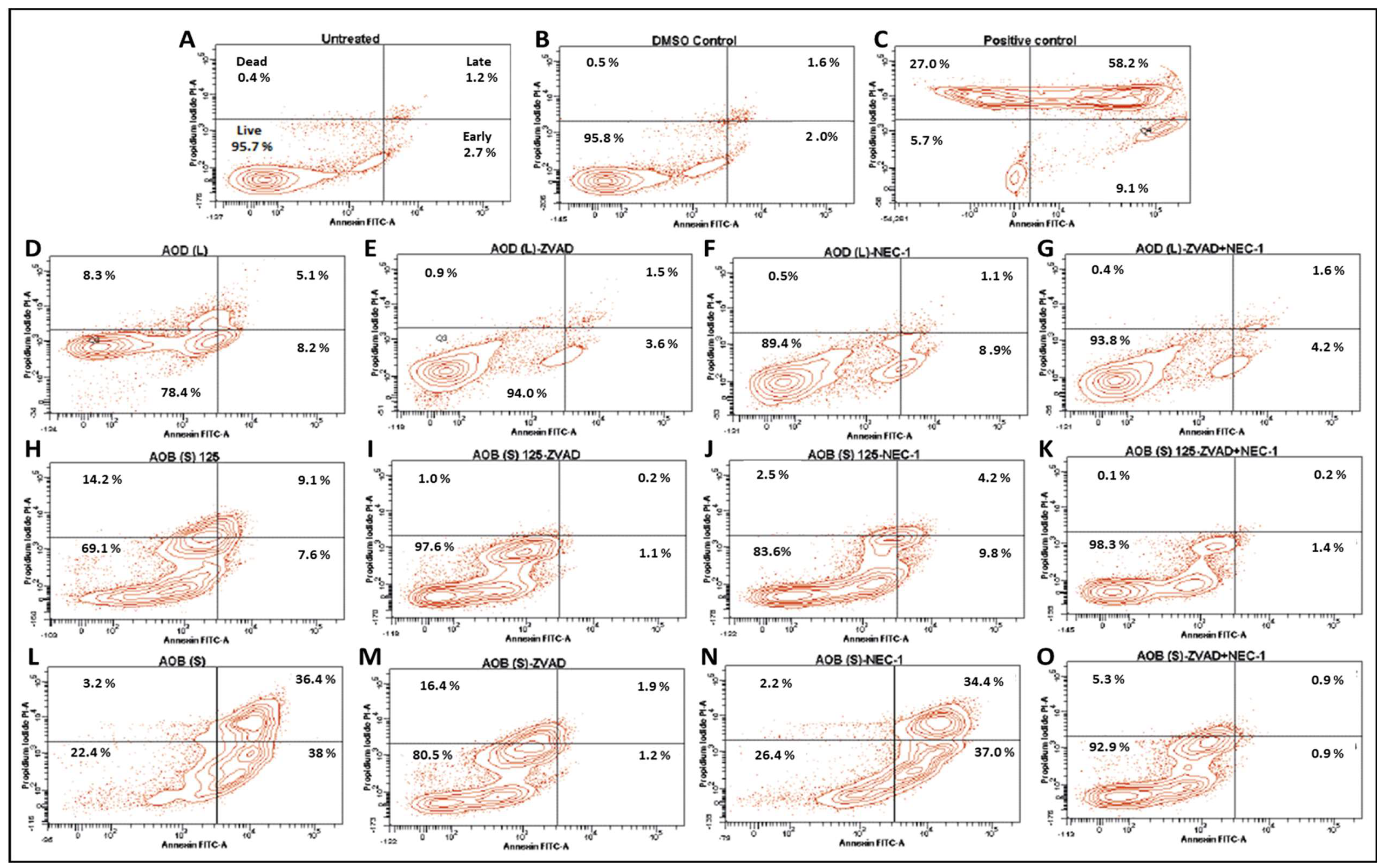
2.5. Test of the Potential of AOD (L) and AOB (S) to Induce Early Signs of Apoptosis in HeLa Cells
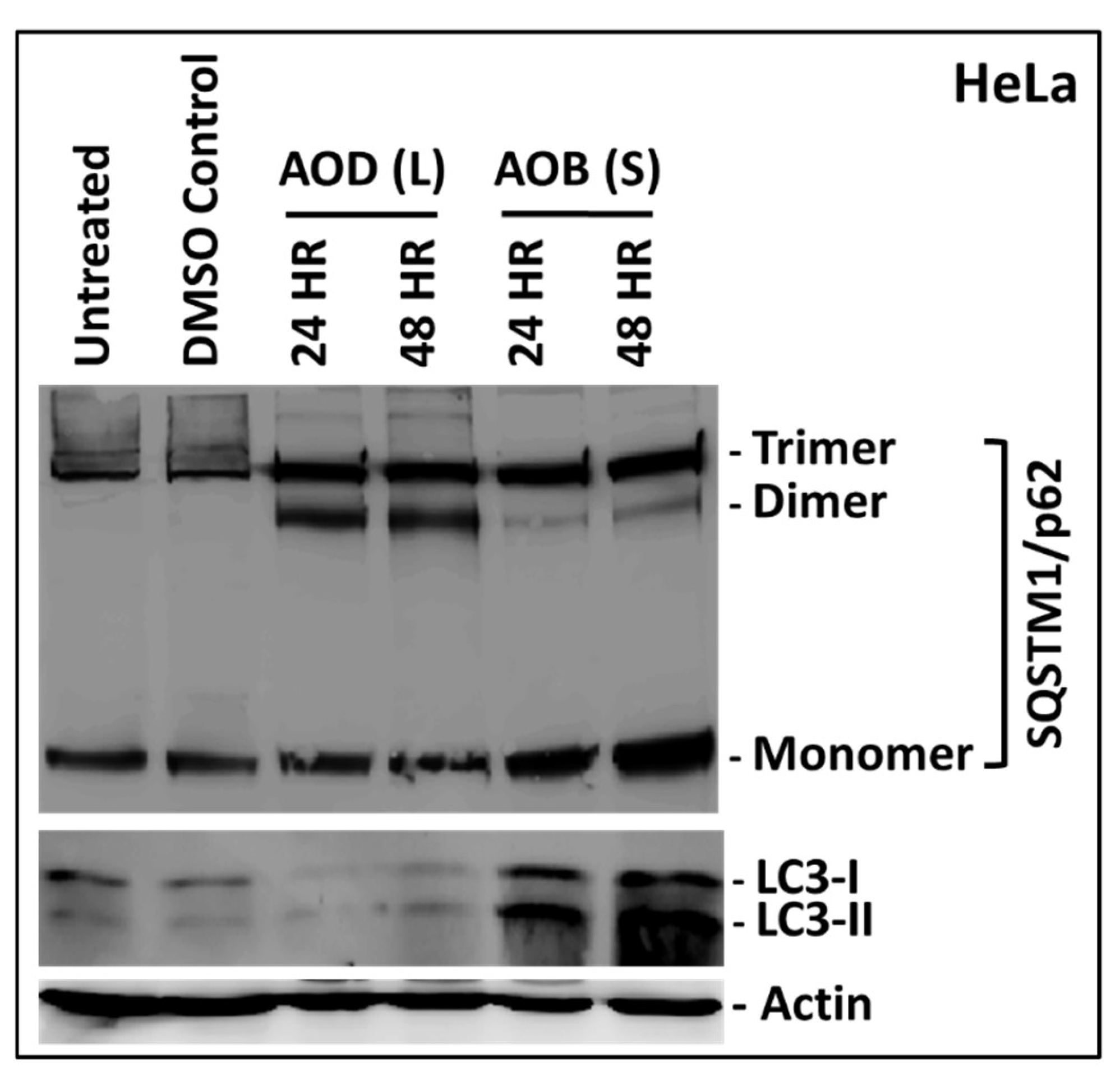
2.6. AOD (L) and AOB (S) Perturb Autophagy
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Plant Collection
4.3. Extraction and Fractionation
4.4. Plant Extract/Fractions Preparation for Cytotoxicity Studies
4.5. MTT Cell Viability Assay
4.6. Luminescent Cell Viability and Caspase-Glo® 3/7, 8 and 9 Assays
4.7. Western Blot Analysis
4.8. Flow Cytometry of Annexin V/Propidium Iodide-Stained Cells
4.9. Morphological Studies
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DMEM | Dulbecco’s Modified Eagles Medium |
| DMSO | Dimethyl Sulfoxide |
| FBS | Fetal Bovine Serum |
| EGF | Epidermal Growth Factor |
| FCS | Fetal Calf Serum |
| MTT | [3-(4,5-dimethylthiazol-2-yl)-2-5-diphenyltetrazolium bromide] |
| PARP | Poly (ADP-ribose) Polymerase |
| PBS | Phosphate Buffered Saline |
| PBST | Phosphate Buffered Saline with 0.1% Tween |
| PMSF | Phenylmethyl Sulfonyl Fluoride |
| RIPA | Radioimmunoprecipitation Assay Buffer |
| SDS | Sodium Dodecyl Sulfate |
References
- Feulner, G.R.; Boer, B.; Western, A.R. The Comprehensive Guide to the Wildflowers of the United Arab Emirates; Environmental Research and Wildlife Development Agency: Abu Dhabi, UAE, 2003. [Google Scholar]
- Winbow, C. The Native Plants of Oman: An introduction; The Environment Society of Oman, Oman Printers and Stationers: Muttrah, Sultanate of Oman, 2008. [Google Scholar]
- Hellyer, P.; Aspinall, S. (Eds.) The Emirates: A Natural History; Trident Press Limited: London, UK, 2005. [Google Scholar]
- Ghazanfar, S.; Fisher, M. Vegetation of the Arabian Peninsula; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998. [Google Scholar]
- Ghazanfar, S.A. Vegetation Structure and Phytogeography of Jabal Shams, an Arid Mountain in Oman. J. Biogeogr. 1991, 18, 299. [Google Scholar] [CrossRef]
- Thulin, M. Flora of Somalia; Kew Royal Botanic Gardens: Kew, UK, 1993. [Google Scholar]
- Cullen, J.; Knees, S.; Cubey, H. Dicotyledons, Resedaceae to Cyrillaceae; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Ghazanfar, S.A.; Al-Al-Sabahi, A.M. Medicinal plants of Northern and Central Oman (Arabia). Econ. Bot. 1993, 47, 89–98. [Google Scholar] [CrossRef]
- Mothana, R.A.; Lindequist, U.; Gruenert, R.; Bednarski, P.J. Studies of the in vitro anticancer, antimicrobial and antioxidant potentials of selected Yemeni medicinal plants from the island Soqotra. BMC Complement. Altern. Med. 2009, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Hammiche, V.; Maiza, K. Traditional medicine in Central Sahara: Pharmacopoeia of Tassili N’ajjer. J. Ethnopharmacol. 2006, 105, 358–367. [Google Scholar] [CrossRef]
- Dalziel, J.; Hutchinson, J. The Useful Plants of West Tropical Africa; Published under the Authority of the Secretary of State for the Colonies by the Crown Agents for the Colonies: London, UK, 1937. [Google Scholar]
- Watt, J.; Breyer-Brandwijk, M. The Medicinal and Poisonous Plants of Southern and Eastern Africa; Livingstone: Edinburgh, UK, 1962. [Google Scholar]
- Ghazanfar, S.A. Handbook of Arabian Medicinal Plant; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Malebo, H.M.; Tanja, W.; Cal, M.; Swaleh, S.A.M.; Omolo, M.O.; Hassanali, A.; Séquin, U.; Hamburger, M.; Brun, R.; Ndiege, I.O. Antiplasmodial, anti-trypanosomal, anti-leishmanial and cytotoxicity activity of selected Tanzanian medicinal plants. Tanzan. J. Health Res. 2009, 11, 226–234. [Google Scholar] [CrossRef]
- Malebo, H.M.; Tanja, W.; Cal, M.; Swaleh, S.A.M.; Hassanali, A.; Sequin, U.; Machocho, A.K.; Haeussinger, D.; Dalsgaard, P.; Omolo, M.O.; et al. Anti-protozoal and structure-activity relationships of chemical constituents of Acridocarpus chloropterus Oliver (Malpighiaceae) from Tanzania. Int. J. Nat. Prod. Res. 2013, 3, 74–81. [Google Scholar]
- Cao, S.; Guza, R.; Miller, J.; Andriantsiferana, R.; Rasamison, V.; Kingston, D. Cytotoxic Triterpenoids from Acridocarpus vivy from the Madagascar rain forest. J. Nat. Prod. 2004, 67, 986–989. [Google Scholar] [CrossRef]
- Sarker, S.; Latif, Z.; Gray, A. Methods in Biotechnology, Natural Products Isolation; Humana Press: Totowa, NJ, USA, 2006; pp. 323–352. [Google Scholar]
- Rehman, N.U.; Hussain, H.; Ali, L.; Khan, A.; Mabood, F.; Shinwari, Z.K.; Hussain, J.; Al-Harrasi, A. Chemical Constituents of Acridocarpus orientalis and Their Chemotaxonomic Significance. Chem. Nat. Compd. 2019, 55, 586–588. [Google Scholar] [CrossRef]
- Ksiksi, T.; Hamza, A.A. Antioxidant, lipoxygenase and histone deacetylase inhibitory activities of Acridocarpus orientalis from Al Ain and Oman. Molecules 2012, 17, 12521–12532. [Google Scholar] [CrossRef]
- Hussain, J.; Ali, L.; Khan, A.L.; Rehman, N.U.; Jabeen, F.; Kim, J.-S.; Al-Harrasi, A. Isolation and Bioactivities of the Flavonoids Morin and Morin-3-O-β-D-glucopyranoside from Acridocarpus orientalis—A Wild Arabian Medicinal Plant. Molecules 2014, 19, 17763–17772. [Google Scholar] [CrossRef]
- Devasagayam, T.P.A.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Physicians India 2004, 52, 794–804. [Google Scholar] [PubMed]
- Rehman, N.U.; Mabood, F.; Khan, A.L.; Ali, L.; Gillani, S.A.; Abbas, G.; Khan, A.; Al-Harrasi, A.; Hussain, J. Evaluation of biological potential and physicochemical properties of Acridocarpus orientalis (Malpighiaceae). Pak. J. Bot. 2019, 51, 3. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Gali-Muhtasib, H.; Hmadi, R.; Kareh, M.; Tohme, R.; Darwiche, N. Cell death mechanisms of plant-derived anticancer drugs: Beyond apoptosis. Apoptosis 2015, 20, 1531–1562. [Google Scholar] [CrossRef] [PubMed]
- Soule, H.D.; Vazquez, J.; Long, A.; Albert, S.; Brennan, M. A Human Cell Line from a Pleural Effusion Derived from a Breast Carcinoma 2. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef]
- Scherer, W.F.; Syverton, J.T.; Gey, G.O. Studies on the propagation in vitro of poliomyelitis viruses. J. Exp. Med. 1953, 97, 695–710. [Google Scholar] [CrossRef]
- Soule, H.D.; Maloney, T.M.; Wolman, S.R.; Peterson, W.D.; Brenz, R.; McGrath, C.M.; Russo, J.; Pauley, R.J.; Jones, R.F.; Brooks, S.C. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990, 50, 6075–6086. [Google Scholar]
- Cailleau, R.; Reeves, W.J.; Young, R.; Olivé, M. Breast Tumor Cell Lines from Pleural Effusions 2. J. Natl. Cancer Inst. 1974, 53, 661–674. [Google Scholar] [CrossRef]
- Panicker, N.G.; Balhamar, S.; Akhlaq, S.; Qureshi, M.M.; Rizvi, T.S.; Al-Harrasi, A.; Hussain, J.; Mustafa, F. Identification and characterization of the caspase-mediated apoptotic activity of Teucrium mascatense and its isolated compound in human cancer cells. Molecules 2019, 24, 977. [Google Scholar] [CrossRef]
- Suhail, M.M.; Wu, W.; Cao, A.; Mondalek, F.G.; Fung, K.-M.; Shih, P.-T.; Fang, Y.-T.; Woolley, C.; Young, G.; Lin, H.-K. Boswellia sacra essential oil induces tumor cell-specific apoptosis and suppresses tumor aggressiveness in cultured human breast cancer cells. BMC Complement. Altern. Med. 2011, 11, 129. [Google Scholar] [CrossRef]
- Aburto, M.R.; Hurle, J.M.; Varela-Nieto, I.; Magariños, M. Autophagy During Vertebrate Development. Cells 2012, 1, 428–448. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, K.; Hu, C.; Lin, L.; Qin, S.; Cai, X. Elemene Injection Induced Autophagy Protects Human Hepatoma Cancer Cells from Starvation and Undergoing Apoptosis. Evid. Complement. Altern. Med. 2014, 2014, 637528. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, M.; Gunning, P.W.; Stehn, J.R. The actin cytoskeleton as a sensor and mediator of apoptosis. BioArchitecture 2012, 2, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Stennicke, H.R.; Salvesen, G.S. Biochemical Characteristics of Caspases-3, -6, -7, and -8. J. Boil. Chem. 1997, 272, 25719–25723. [Google Scholar] [CrossRef] [PubMed]
- Vermes, I.; Haanen, C.; Reutelingsperger, C. Flow cytometry of apoptotic cell death. J. Immunol. Methods 2000, 243, 167–190. [Google Scholar] [CrossRef]
- Christofferson, D.E.; Yuan, J. Necroptosis as an alternative form of programmed cell death. Curr. Opin. Cell Boil. 2010, 22, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Boil. 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef]
- Guamán-Ortiz, L.M.; Orellana, M.I.R.; Ratovitski, E.A. Natural Compounds as Modulators of Non-Apoptotic Cell Death in Cancer Cells. Curr. Genom. 2017, 18, 132–155. [Google Scholar] [CrossRef]
- Carroll, B.; Otten, E.G.; Manni, D.; Stefanatos, R.; Menzies, F.M.; Smith, G.R.; Jurk, D.; Kenneth, N.; Wilkinson, S.; Passos, J.F.; et al. Oxidation of SQSTM1/p62 mediates the link between redox state and protein homeostasis. Nat. Commun. 2018, 9, 256. [Google Scholar] [CrossRef]
- Suffredini, I.B.; Paciencia, M.L.B.; AFrana, S.; Varella, A.D.; Younes, R.N. In vitro breast cancer cell lethality of Brazilian plant extracts. Die Pharm. 2007, 62, 798–800. [Google Scholar]
- Fouche, G.; Cragg, G.; Pillay, P.; Kolesnikova, N.; Maharaj, V.; Senabe, J. In vitro anticancer screening of South African plants. J. Ethnopharmacol. 2008, 119, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Abbasi, B.A.; Mahmood, T.; Kanwal, S.; Ali, B.; Shah, S.A.; Khalil, A.T. Plant-derived anticancer: Agents: A green anticancer approach. Asian Pac. J. Trop. Biomed. 2017, 7, 1129–1150. [Google Scholar] [CrossRef]
- Jaszczyszyn, A.; Gąsiorowski, K. Limitations of the MTT assay in cell viability testing. Adv. Clin. Exp. Med. 2008, 17, 5, 525–529. [Google Scholar]
- Mahmoodi, N.; Motamed, N.; Paylakhi, S.K.; Mahmoodi, N.O. Comparing the effect of Silybin and Silybin Advanced™ on viability and HER2 expression on the human breast cancer SKBR3 cell line by no serum starvation. Iran J. Pharm. Res. 2015, 14, 521–530. [Google Scholar]
- Provinciali, M.; Papalini, F.; Orlando, F.; Pierpaoli, S.; Donnini, A.; Morazzoni, P.; Riva, A.; Smorlesi, A. Effect of the Silybin-Phosphatidylcholine Complex (IdB 1016) on the Development of Mammary Tumors in HER-2/neu Transgenic Mice. Cancer Res. 2007, 67, 2022–2029. [Google Scholar] [CrossRef]
- Galluzzi, L.; Pietrocola, F.; Pedro, J.M.B.-S.; Amaravadi, R.K.; Baehrecke, E.H.; Cecconi, F.; Codogno, P.; Debnath, J.; AGewirtz, D.; Karantza, V.; et al. Autophagy in malignant transformation and cancer progression. EMBO J. 2015, 34, 856–880. [Google Scholar]
- Chen, N.; Karantza, V. Autophagy as a therapeutic target in cancer. Cancer Boil. Ther. 2011, 11, 157–168. [Google Scholar]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Arozena, A.A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar]
- Tanida, I.; Waguri, S. Measurement of Autophagy in Cells and Tissues. Adv. Struct. Saf. Stud. 2010, 648, 193–214. [Google Scholar]
- Mildenberger, J.; Johansson, I.; Sergin, I.; Kjøbli, E.; Damås, J.K.; Razani, B.; Flo, T.H.; Bjørkøy, G. N-3 PUFAs induce inflammatory tolerance by formation of KEAP1-containing SQSTM1/p62-bodies and activation of NFE2L2. Autophagy 2017, 13, 1664–1678. [Google Scholar] [CrossRef]
- Das, G.; Shravage, B.V.; Baehrecke, E.H. Regulation and Function of Autophagy during Cell Survival and Cell Death. Cold Spring Harb. Perspect. Boil. 2012, 4, a008813. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P.E.; Rice-Evans, C. Flavonoids: Antioxidants or signalling molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Soobrattee, M.; Neergheen, V.; Luximon-Ramma, A.; Aruoma, O.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. Mol. Mech. Mutagen. 2005, 579, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Okubo, T.; Kano, I. Studies on estrogenic activities of food additives with human breast cancer MCF-7 cells and mechanism of estrogenicity by BHA and OPP. Yakugaku Zasshi 2003, 123, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Lee, W.S.; Eun, S.Y.; Jung, J.H.; Park, H.-S.; Kim, G.; Choi, Y.H.; Ryu, C.H.; Jung, J.M.; Hong, S.C.; et al. Morin, a flavonoid from Moraceae, suppresses growth and invasion of the highly metastatic breast cancer cell line MDA-MB-231 partly through suppression of the Akt pathway. Int. J. Oncol. 2014, 45, 1629–1637. [Google Scholar] [CrossRef]
- Maicheen, C.; Jittikoon, J.; Vajragupta, O.; Ungwitayatorn, J. Synthesis, topoisomerase I inhibitory and cytotoxic activities of chromone derivatives. Med. Chem. 2013, 9, 329–339. [Google Scholar] [CrossRef]
- Tang, S.W.; Sukari, M.A.; Neoh, B.K.; Yeap, Y.S.Y.; Abdul, A.B.; Kifli, N.; Ee, G.C.L. Phytochemicals from Kaempferia angustifolia Rosc. and their cytotoxic and antimicrobial activities. BioMed Res. Int. 2014, 2014, 417674. [Google Scholar] [CrossRef]
- Dilshara, M.G.; Karunarathne, W.A.; Molagoda, I.M.; Kang, C.H.; Choi, Y.H.; Kim, G.Y. Morin suppresses cellular invasion by decreasing nuclear factor-κB-mediated matrix metalloproteinase-9 activity. Lat. Am. J. Pharm. 2018, 37, 1264–1271. [Google Scholar]
- Ju, Y.H.; Clausen, L.M.; Allred, K.F.; Almada, A.L.; Helferich, W.G. β-sitosterol, β-sitosterol glucoside, and a mixture of β-sitosterol and β-sitosterol glucoside modulate the growth of estrogen-responsive breast cancer cells in vitro and in ovariectomized athymic mice. J. Nutr. 2004, 134, 1145–1151. [Google Scholar] [CrossRef]
- Wimmerová, M.; Siglerová, V.; Šaman, D.; Šlouf, M.; Kaletová, E.; Wimmer, Z. Improved enzyme-mediated synthesis and supramolecular self-assembly of naturally occurring conjugates of β-sitosterol. Steroids 2017, 117, 38–43. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y.; Han, B.; Li, Z.; Wang, B.; Jiang, P.; Zhang, J.; Ma, W.; Zhou, D.; Li, X.; et al. Anti-breast-Cancer Activity Exerted by β-Sitosterol-d-glucoside from Sweet Potato via Upregulation of MicroRNA-10a and via the PI3K–Akt Signaling Pathway. J. Agric. Food Chem. 2018, 66, 9704–9718. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Yao, Y.; Shi, Z.; Everaert, N.; Ren, G. Synergistic Effect of Bioactive Anticarcinogens from Soybean on Anti-Proliferative Activity in MDA-MB-231 and MCF-7 Human Breast Cancer Cells In Vitro. Molecules 2018, 23, 1557. [Google Scholar] [CrossRef] [PubMed]
- Alam, P.; Alhowiriny, T.A.; Siddiqui, N.A.; Alqasoumi, S.I.; Basudan, O.A.; Khan, A.A.; Alhowiriny, A.T.; Alam, N. Interspecies estimation of β-sitosterol by a validated high-performance thin-layer chromatography method in genus Ficus and cytotoxic activity against HepG2, HEK-293, MCF-7, and MDA-MB-231 cell lines. J. Planar Chromatogr. Mod. TLC 2018, 31, 213–219. [Google Scholar] [CrossRef]
- Rustaie, A.; Hadjiakhoondi, A.; Akbarzadeh, T.; Safavi, M.; Samadi, N.; Sabourian, R.; Khanavi, M. Phytochemical constituents and biological activities of Salvia suffruticosa. Res. J. Pharmacogn. 2018, 5, 25–32. [Google Scholar]
- Shahat, A.A.; Hidayathulla, S.; Khan, A.A.; Alanazi, A.M.; Al Meanazel, O.T.; Alqahtani, A.S.; Alsaid, M.S.; Hussein, A.A. Phytochemical profiling, antioxidant and anticancer activities of Gastrocotyle hispida growing in Saudi Arabia. Acta Trop. 2019, 191, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Dehelean, C.A.; Feflea, S.; Molnár, J.; Zupko, I.; Soica, C. Betulin as an antitumor agent tested in vitro on A431, HeLa and MCF7, and as an angiogenic inhibitor in vivo in the CAM assay. Nat. Prod. Commun. 2012, 7, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Damle, A.A.; Pawar, Y.P.; Narkar, A.A. Anticancer activity of betulinic acid on MCF-7 tumors in nude mice. Indian J. Exp. Boil. 2013, 51, 485–491. [Google Scholar]
- Tiwari, R.; Puthli, A.; Balakrishnan, S.; Sapra, B.K.; Mishra, K.P. Betulinic Acid-Induced Cytotoxicity in Human Breast Tumor Cell Lines MCF-7 and T47D and its Modification by Tocopherol. Cancer Investig. 2014, 32, 402–408. [Google Scholar] [CrossRef]
- Król, S.K.; Kiełbus, M.; Rivero-Müller, A.; Stepulak, A. Comprehensive Review on Betulin as a Potent Anticancer Agent. BioMed Res. Int. 2015, 2015, 1–11. [Google Scholar] [CrossRef]
- Saeed, M.E.M.; Mahmoud, N.; Sugimoto, Y.; Efferth, T.; Abdel-Aziz, H. Betulinic Acid Exerts Cytotoxic Activity Against Multidrug-Resistant Tumor Cells via Targeting Autocrine Motility Factor Receptor (AMFR). Front. Pharmacol. 2018, 9, 481. [Google Scholar] [CrossRef]
- Hordyjewska, A.; Ostapiuk, A.; Horecka, A. Betulin and betulinic acid in cancer research. J. Pre-Clin. Clin. Res. 2018, 12, 72–75. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds 1–6 are available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts/Fractions | MCF-7 (µg/mL) Breast Cancer Cells | HeLa (µg/mL) Cervical Cancer Cells |
|---|---|---|
| AOH (L) | X | X |
| AOD (L) | √ (250; 72 h) | √ (125; 72 h/250; 72 h) |
| AOEA (L) | X | √ (125; 72 h/250; 72 h) |
| AOB (L) | X | X |
| AOM (L) | X | X |
| AOAQ (L) | X | X |
| AOH (S) | X | X |
| AOD (S) | X | √ (125; 72 h) |
| AOEA (S) | X | X |
| AOB (S) | X | √ (250; 72 h) |
| AOM (S) | X | X |
| AOAQ (S) | X | X |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balhamar, S.O.M.S.; Panicker, N.G.; Akhlaq, S.; Qureshi, M.M.; Ahmad, W.; Rehman, N.U.; Ali, L.; Al-Harrasi, A.; Hussain, J.; Mustafa, F. Differential Cytotoxic Potential of Acridocarpus orientalis Leaf and Stem Extracts with the Ability to Induce Multiple Cell Death Pathways. Molecules 2019, 24, 3976. https://doi.org/10.3390/molecules24213976
Balhamar SOMS, Panicker NG, Akhlaq S, Qureshi MM, Ahmad W, Rehman NU, Ali L, Al-Harrasi A, Hussain J, Mustafa F. Differential Cytotoxic Potential of Acridocarpus orientalis Leaf and Stem Extracts with the Ability to Induce Multiple Cell Death Pathways. Molecules. 2019; 24(21):3976. https://doi.org/10.3390/molecules24213976
Chicago/Turabian StyleBalhamar, Sameera Omar Mohammed Saeed, Neena Gopinathan Panicker, Shaima Akhlaq, Mohammed Mansoor Qureshi, Waqar Ahmad, Najeeb Ur Rehman, Liaqat Ali, Ahmed Al-Harrasi, Javid Hussain, and Farah Mustafa. 2019. "Differential Cytotoxic Potential of Acridocarpus orientalis Leaf and Stem Extracts with the Ability to Induce Multiple Cell Death Pathways" Molecules 24, no. 21: 3976. https://doi.org/10.3390/molecules24213976
APA StyleBalhamar, S. O. M. S., Panicker, N. G., Akhlaq, S., Qureshi, M. M., Ahmad, W., Rehman, N. U., Ali, L., Al-Harrasi, A., Hussain, J., & Mustafa, F. (2019). Differential Cytotoxic Potential of Acridocarpus orientalis Leaf and Stem Extracts with the Ability to Induce Multiple Cell Death Pathways. Molecules, 24(21), 3976. https://doi.org/10.3390/molecules24213976