
Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide


 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Method
2.1. Compounds and Peptide
2.2. Cell Lines
2.3. MTT Cell Viability Assay for the Individual Compounds
2.4. MTT Cell Viability Assay for the Co-Administration
2.5. Dual-Fluorescence for Live/Dead Nucleated Cell Assay
2.6. Molecular Docking
2.7. Molecular Dynamics Simulation
2.8. Statistical Analysis
3. Results
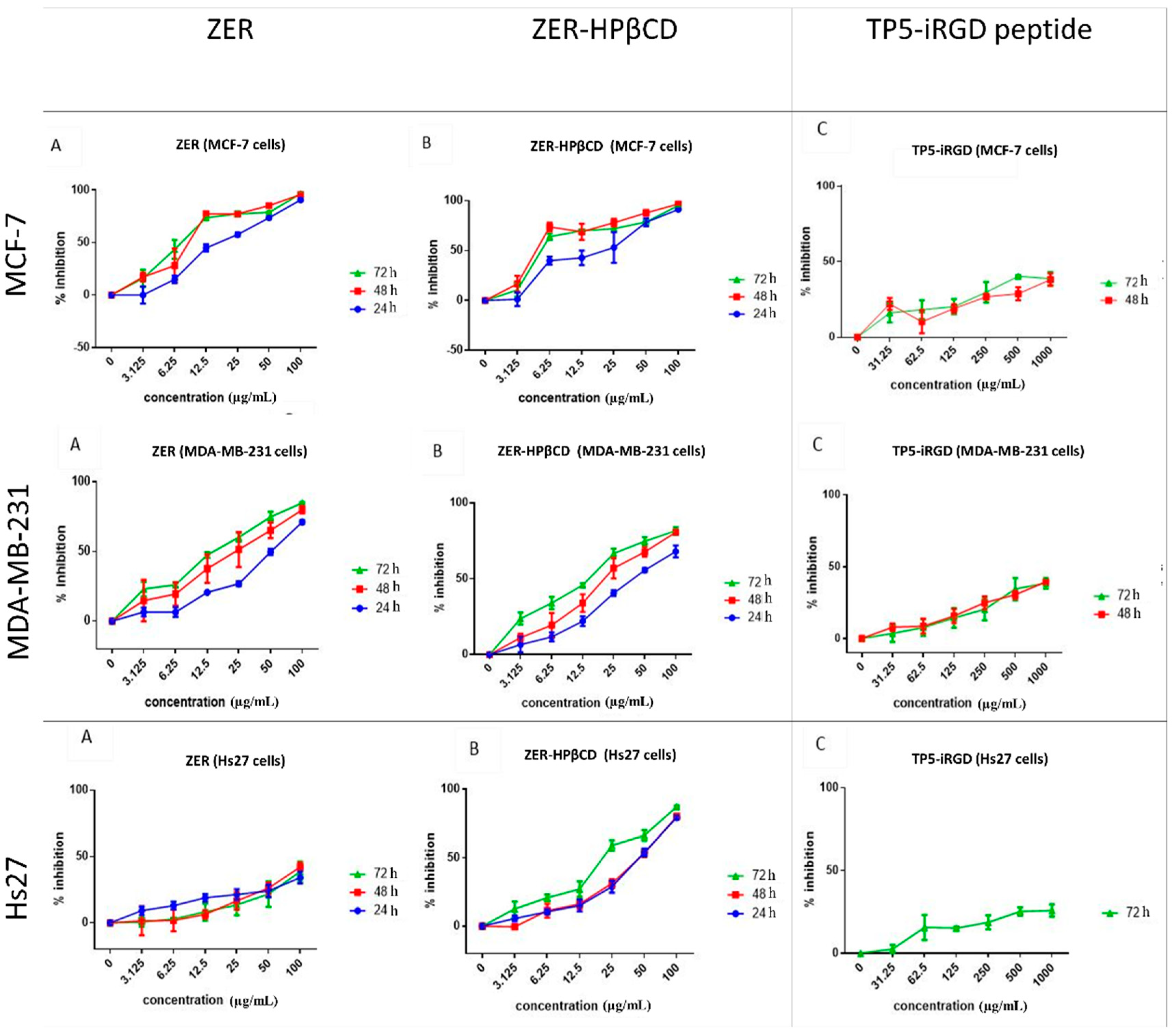
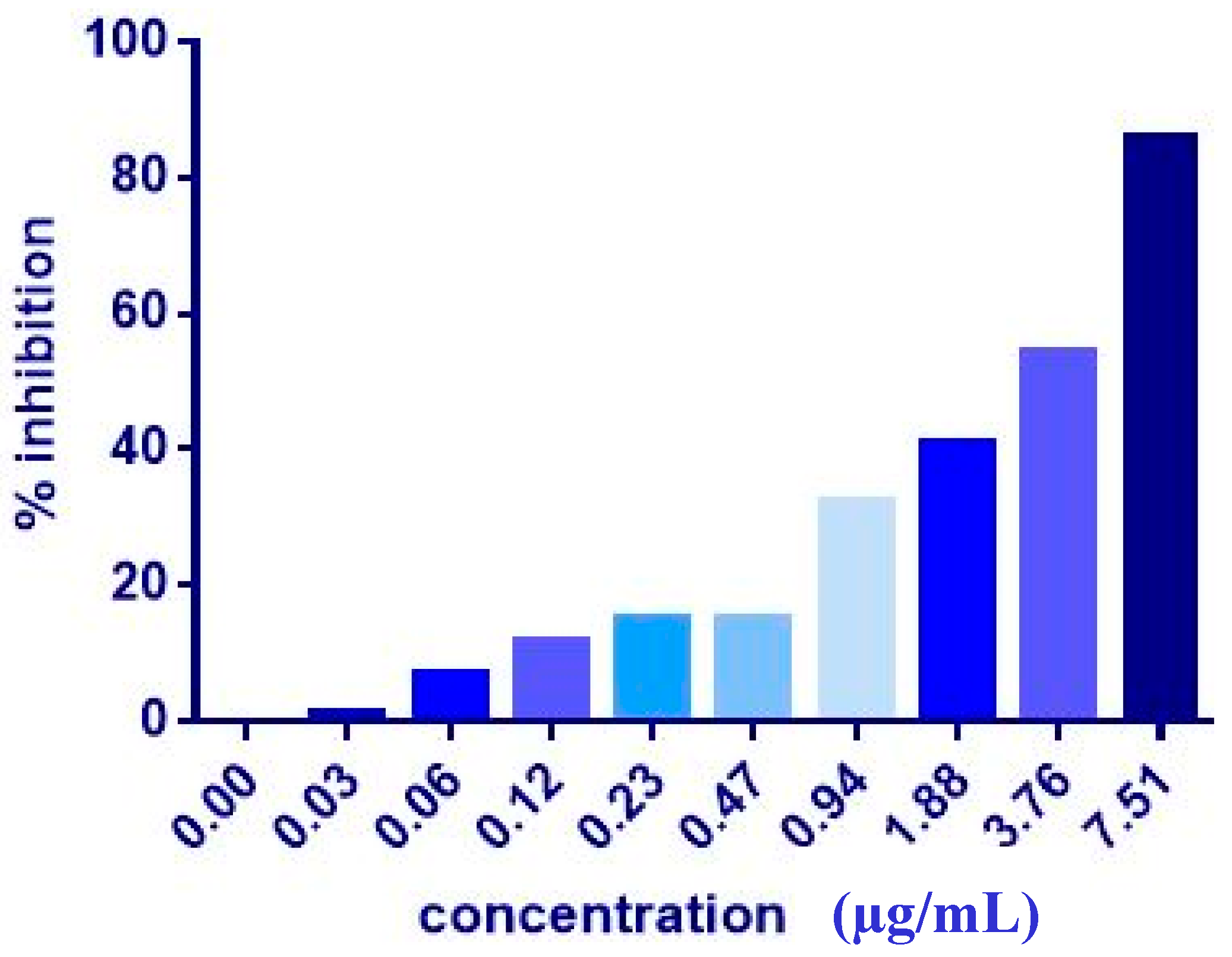
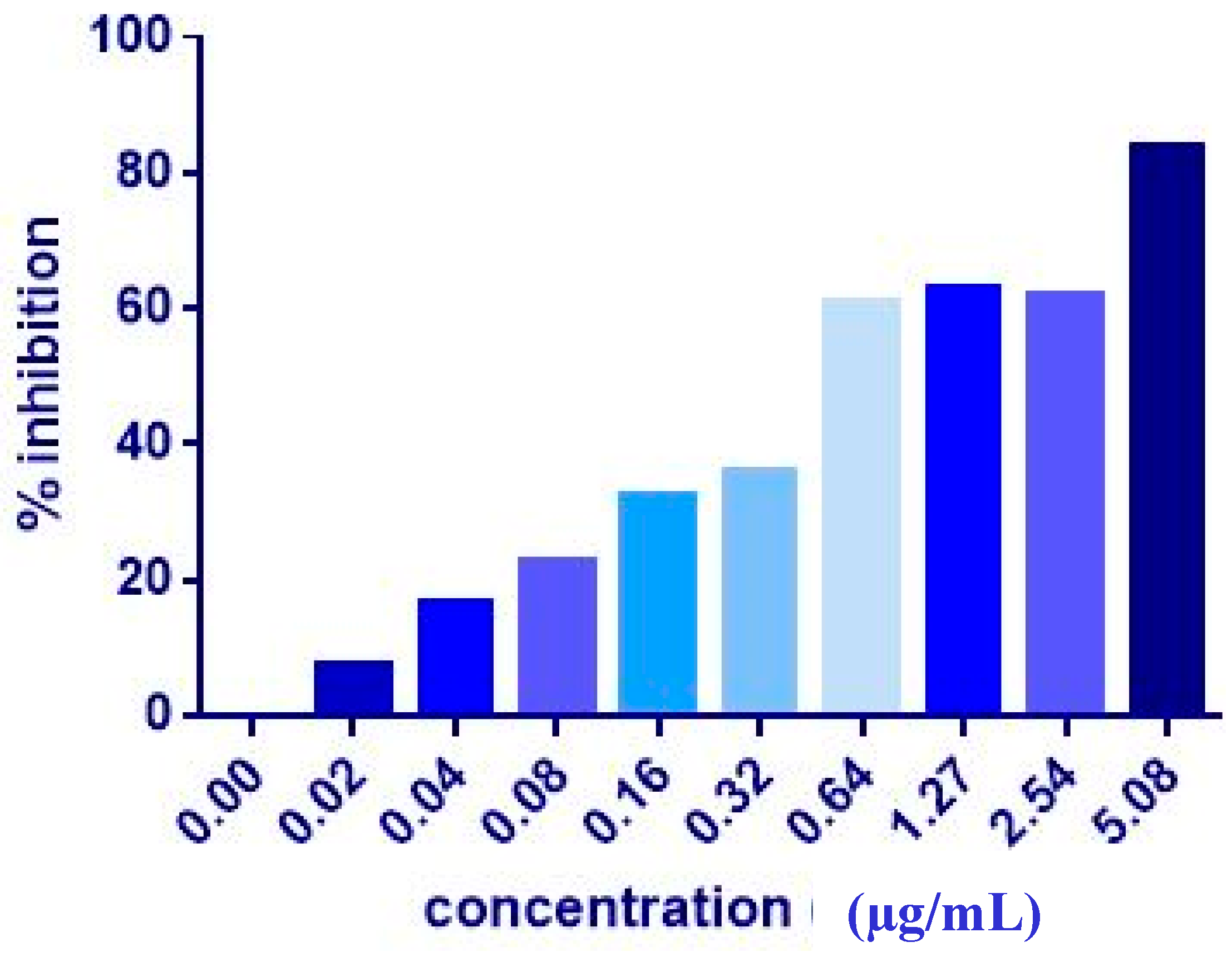
3.1. MTT Assay of Individual Compounds
3.2. MTT Assay of the Co-Administration
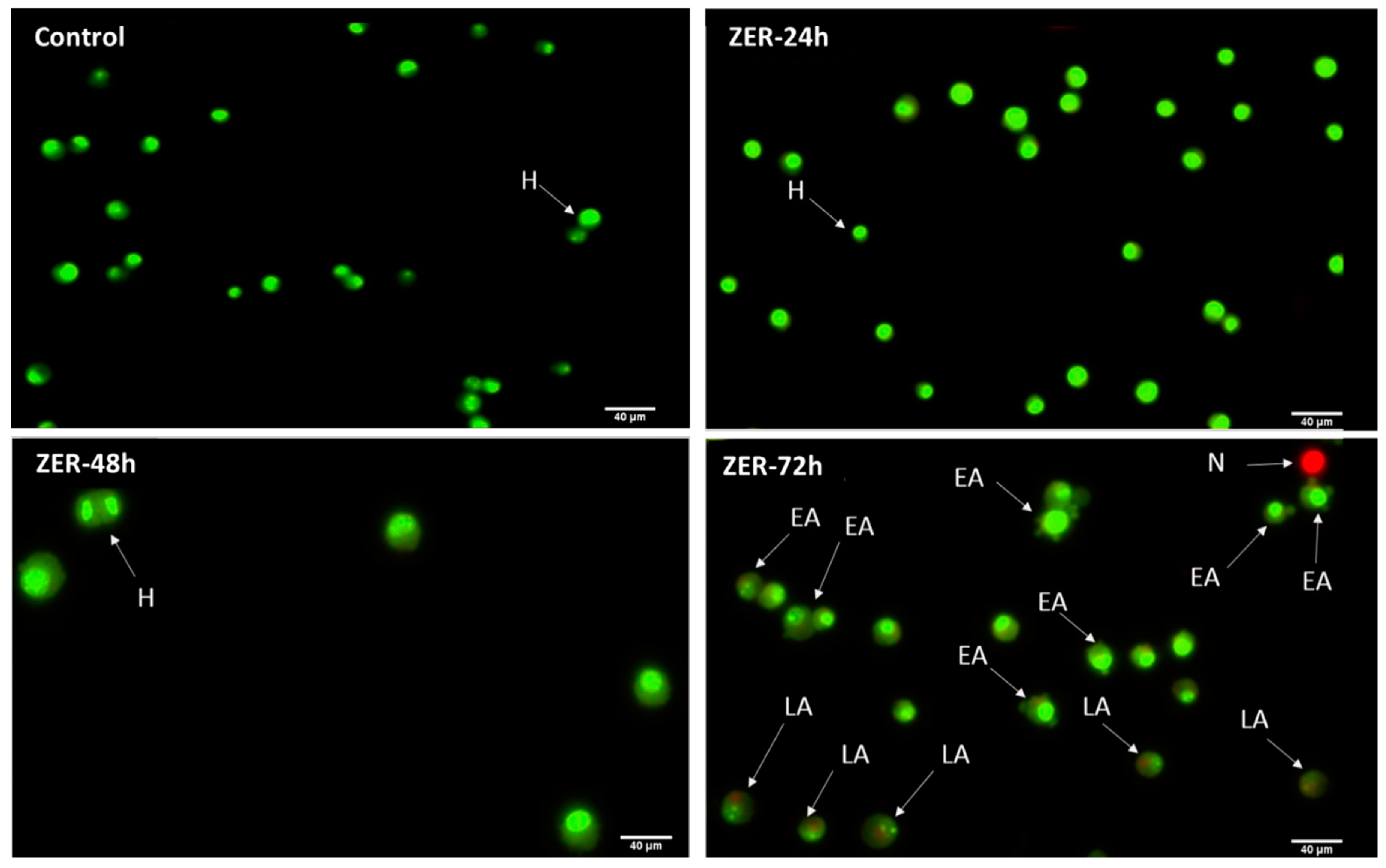
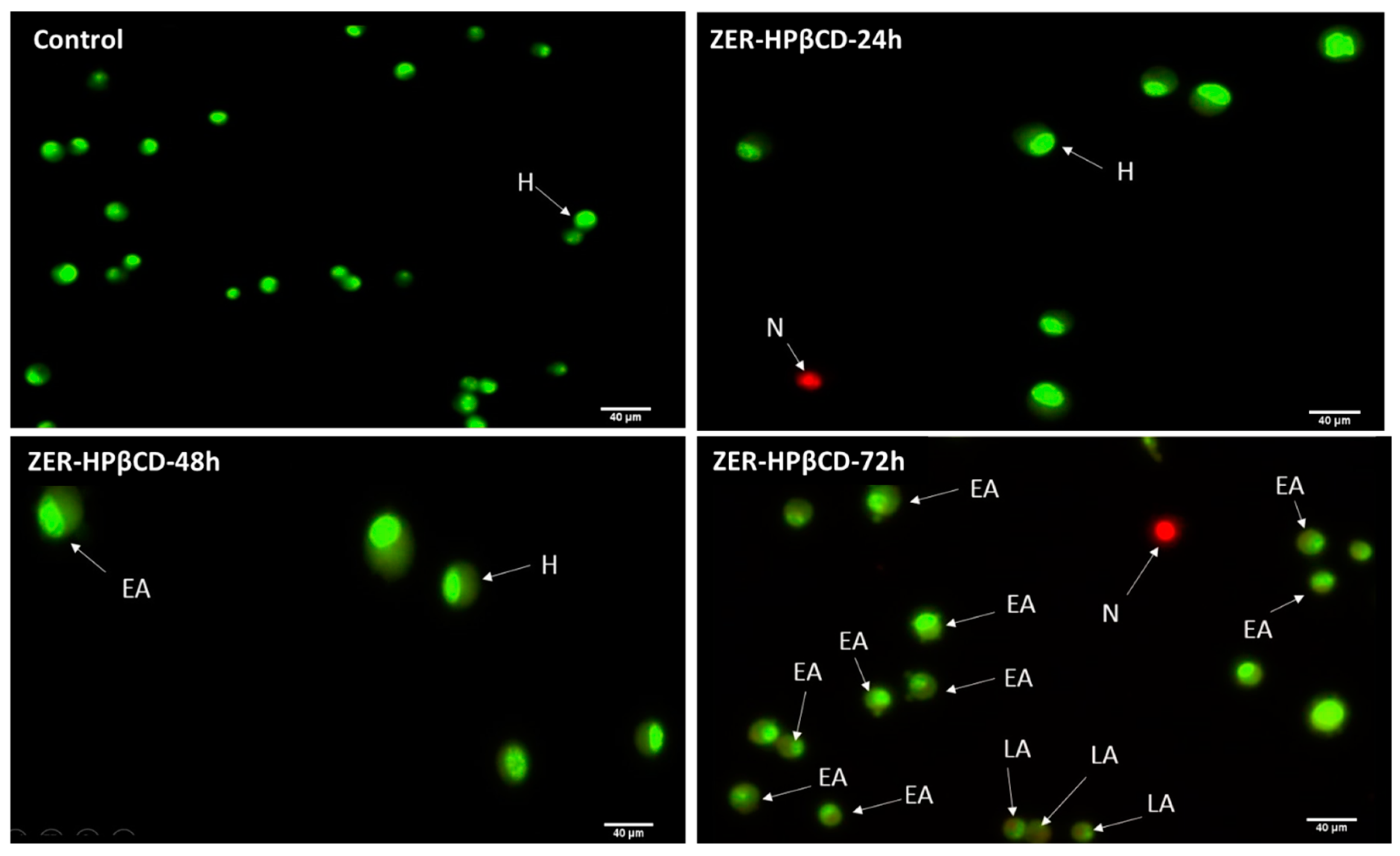
3.3. Apoptosis Detection by Dual Staining
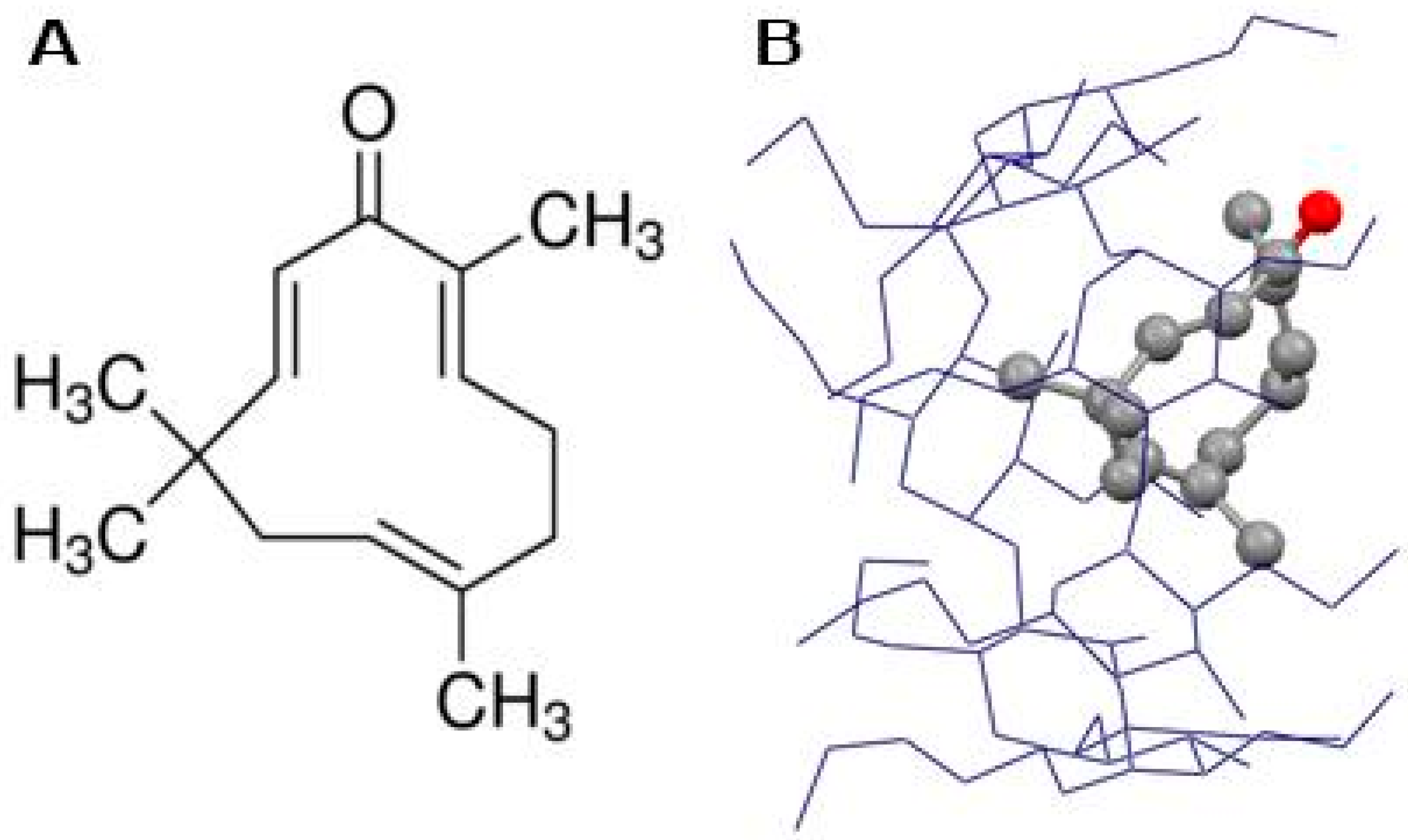
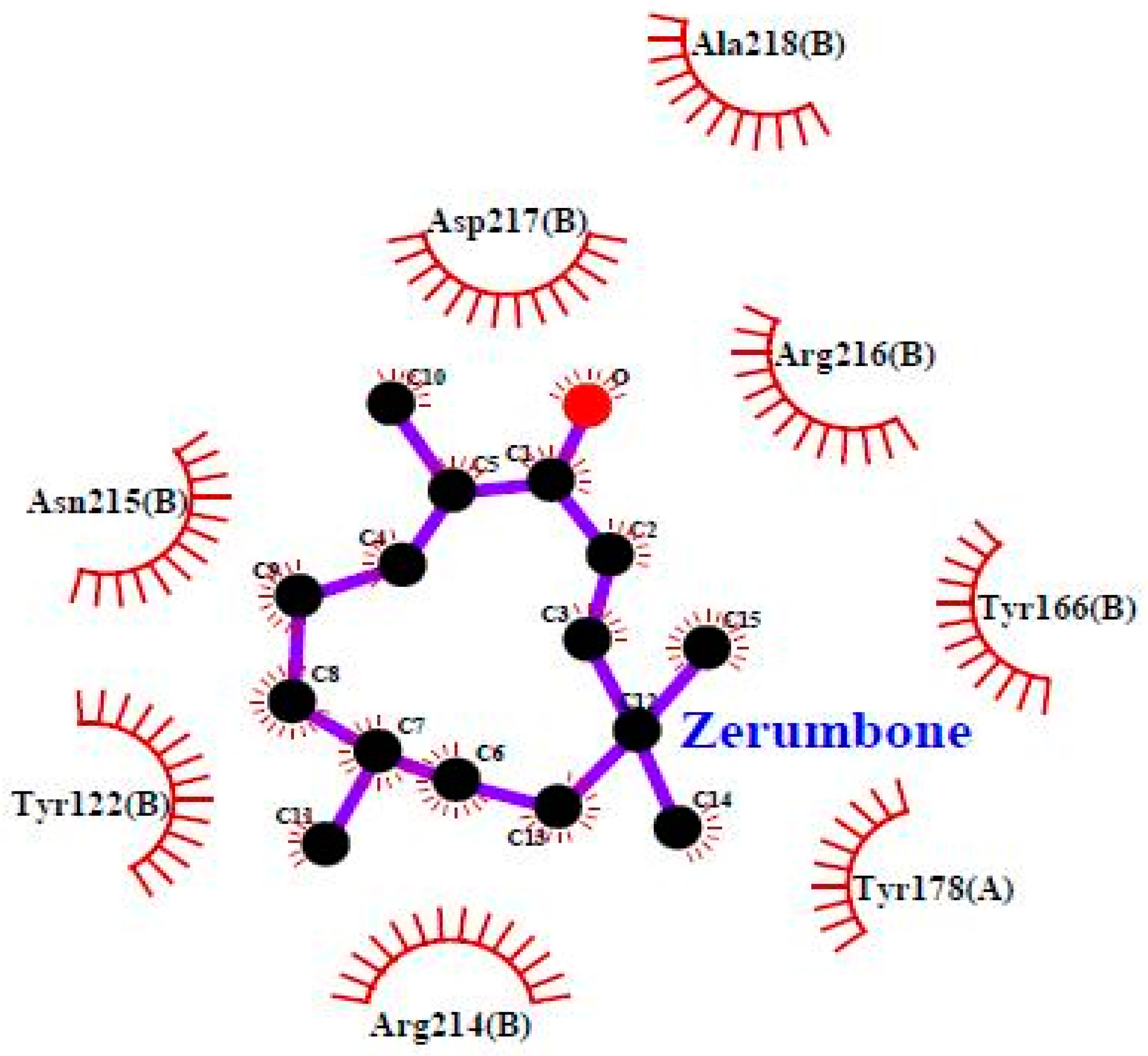
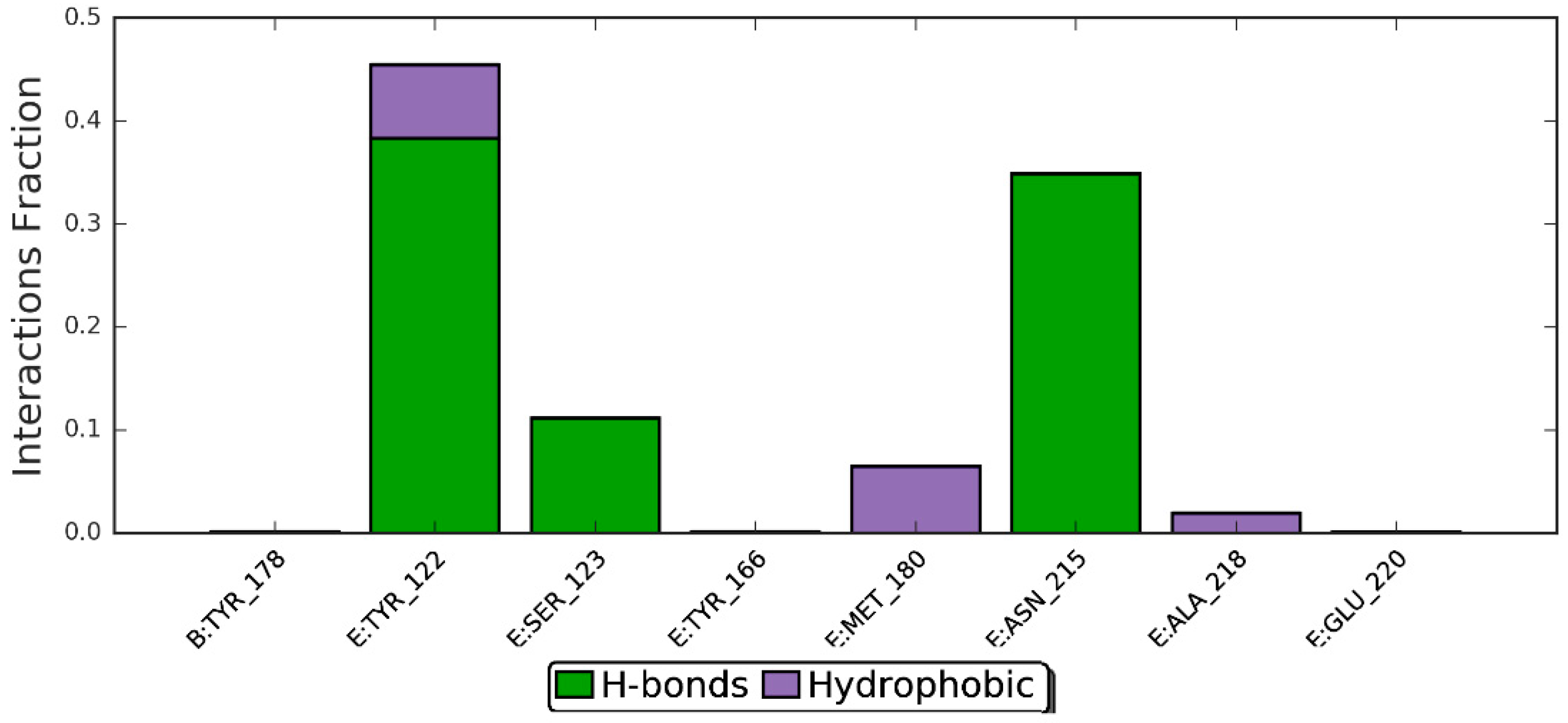
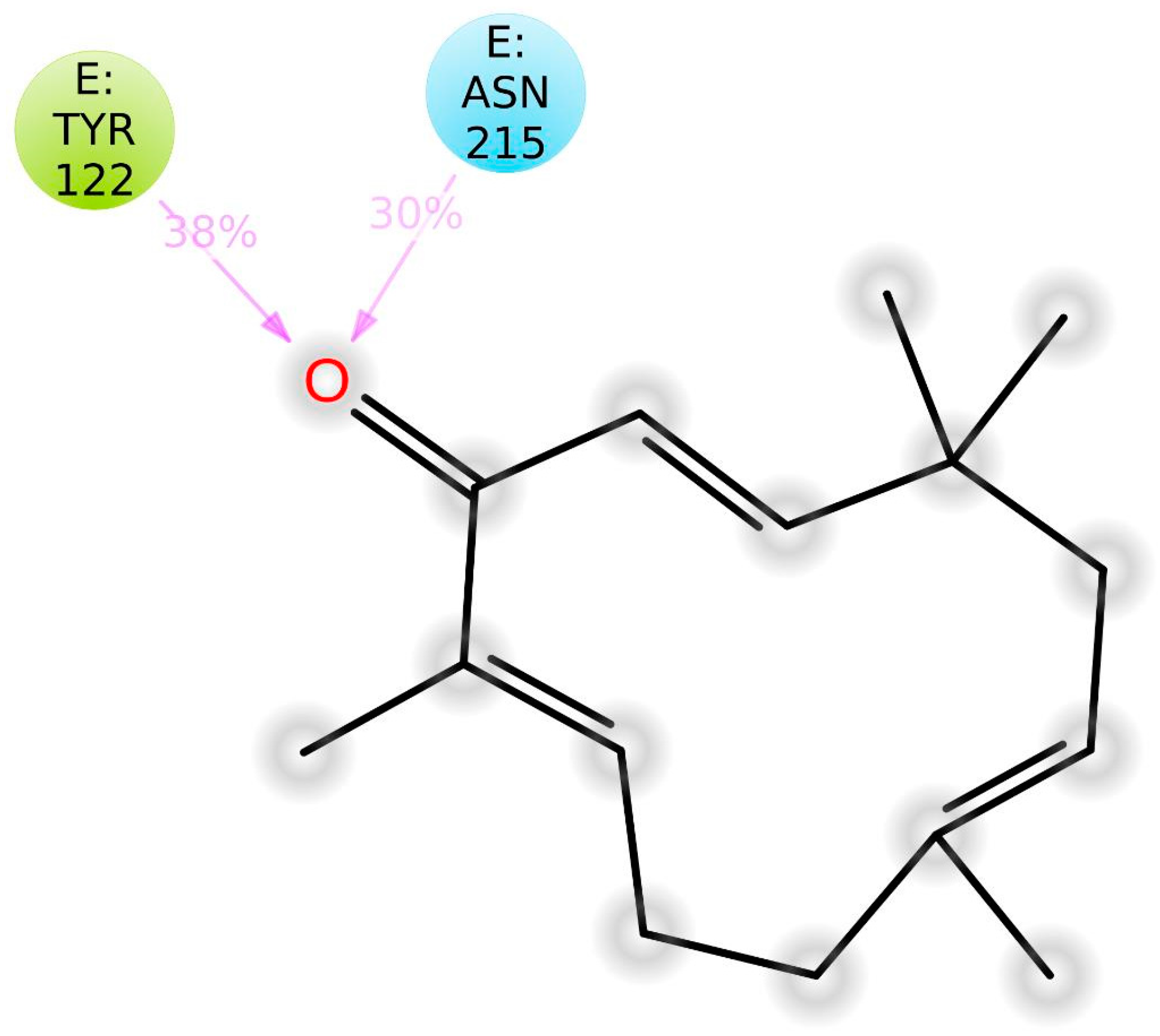
3.4. Molecular Docking
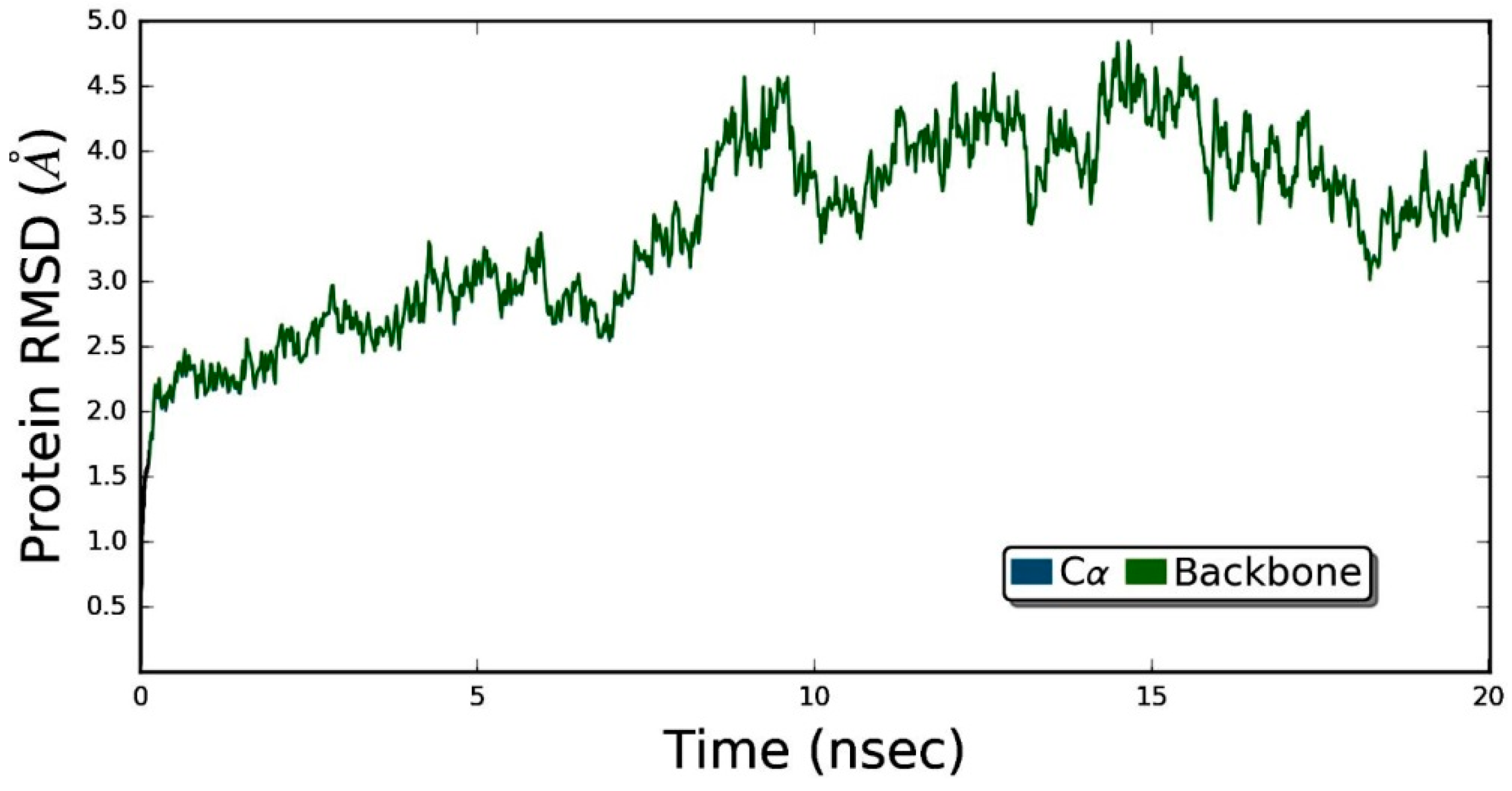
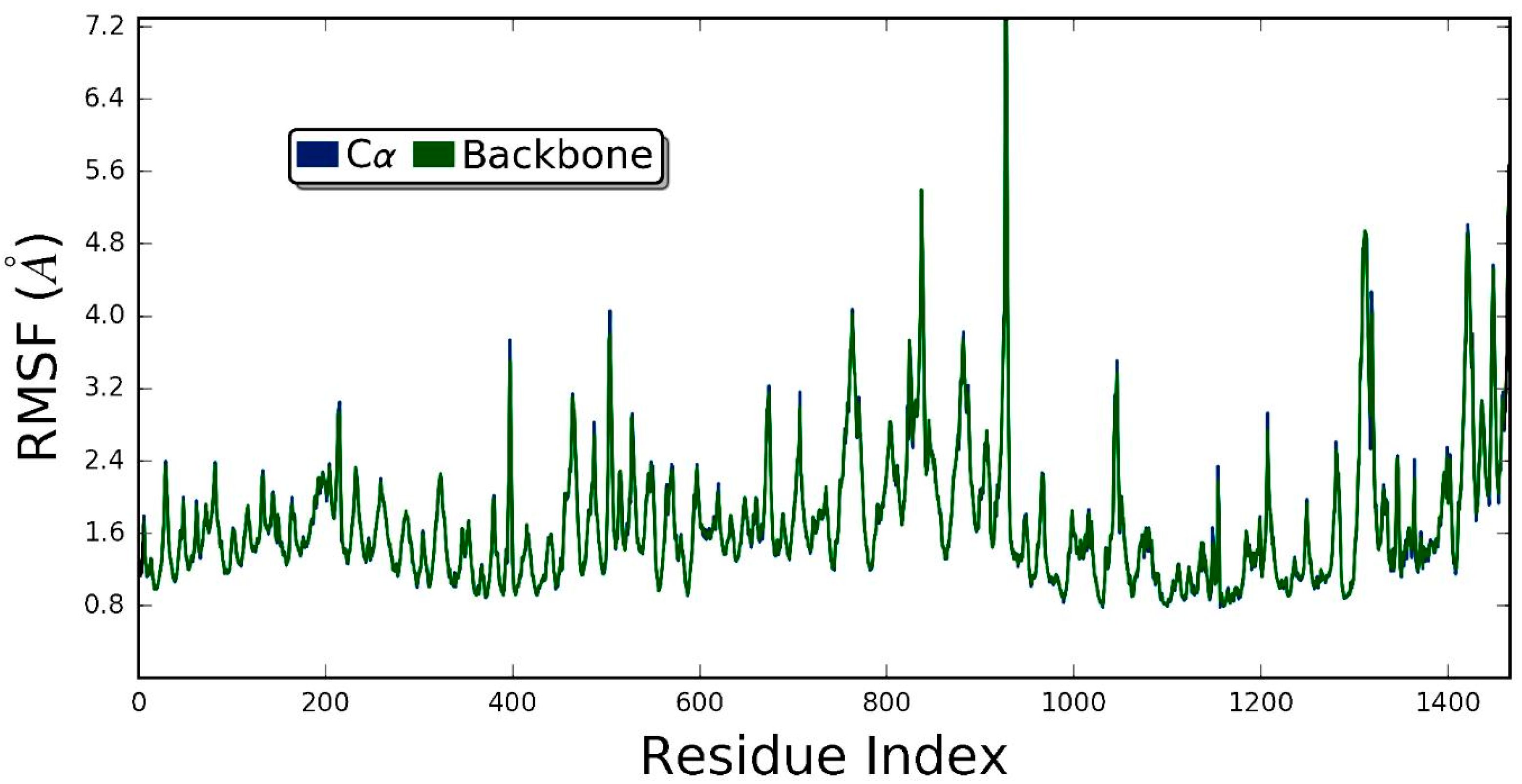
3.5. Molecular Dynamics Simulation Studies of ZER in Complex with TP5-iRGD/Integrin αvβ3
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Ca: A Cancer J. Clin. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Yang, J.; Zhang, Q.; Yang, J.; Wang, H.; Xu, J.; Zheng, J. iRGD as a tumorpenetrating peptide for cancer therapy (Review). Mol. Med. Rep. 2017, 15, 2925–2930. [Google Scholar] [CrossRef] [PubMed]
- Minchinton, A.I.; Tannock, I.F. Drug penetration in solid tumours. Nat. Rev. Cancer 2006, 6, 583. [Google Scholar] [CrossRef] [PubMed]
- Hambley, T.W.; Hait, W.N. Is anticancer drug development heading in the right direction? Cancer Res. 2009, 69, 1259–1262. [Google Scholar] [CrossRef] [PubMed]
- Hai-Tao, Z.; Hui-Cheng, L.; Zheng-Wu, L.; Chang-Hong, G. A tumor-penetrating peptide modification enhances the antitumor activity of endostatin in vivo. Anti-Cancer Drugs 2011, 22, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Rabbani-Chadegani, A.; Paydar, P.; Amirshenava, M.; Aramvash, A. An in vitro study on the effect of vinca alkaloid, vinorelbine, on chromatin histone, HMGB proteins and induction of apoptosis in mice non-adherent bone marrow cells. Drug Chem. Toxicol. 2015, 38, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Böhme, D.; Beck-Sickinger, A.G. Drug delivery and release systems for targeted tumor therapy. J. Pept. Sci. 2015, 21, 186–200. [Google Scholar]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef]
- Guo, W.; Giancotti, F.G. Integrin signalling during tumour progression. Nat. Rev. Mol. Cell Biol. 2004, 5, 816. [Google Scholar] [CrossRef]
- Xiong, J.-P.; Stehle, T.; Zhang, R.; Joachimiak, A.; Frech, M.; Goodman, S.L.; Arnaout, M.A. Crystal structure of the extracellular segment of integrin αVβ3 in complex with an Arg-Gly-Asp ligand. Science 2002, 296, 151–155. [Google Scholar] [CrossRef]
- Dutta, S.; Mahalanobish, S.; Saha, S.; Ghosh, S.; Sil, P.C. Natural products: An upcoming therapeutic approach to cancer. Food Chem. Toxicol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Khazir, J.; Mir, B A.; Mir, S.A.; Cowan, D. Natural products as lead compounds in drug discovery. Taylor Fr. 2013. [Google Scholar]
- Eid, E.E.; Abdul, A.B.; Suliman, F.E.O.; Sukari, M.A.; Rasedee, A.; Fatah, S.S. Characterization of the inclusion complex of zerumbone with hydroxypropyl-β-cyclodextrin. Carbohydr. Polym. 2011, 83, 1707–1714. [Google Scholar] [CrossRef]
- Sung, B.; Murakami, A.; Oyajobi, B.O.; Aggarwal, B.B. Zerumbone abolishes RANKL-induced NF-κB activation, inhibits osteoclastogenesis, and suppresses human breast cancer–induced bone loss in athymic nude mice. Cancer Res. 2009, 69, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, A.; Arlotti, J.A.; Murakami, A.; Singh, S.V. Zerumbone causes Bax-and Bak-mediated apoptosis in human breast cancer cells and inhibits orthotopic xenograft growth in vivo. Breast Cancer Res. Treat. 2012, 136, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Bae, S.Y.; Oh, S.J.; Lee, J.; Lee, J.H.; Lee, H.C.; Lee, S.K.; Kil, W.H.; Kim, S.W.; Nam, S.J. Zerumbone Suppresses IL-1β-induced Cell Migration and Invasion by Inhibiting IL-8 and MMP-3 Expression in Human Triple-negative Breast Cancer Cells. Phytother. Res. 2014, 28, 1654–1660. [Google Scholar] [CrossRef]
- Kim, S.; Kil, W.H.; Lee, J.; Oh, S.-J.; Han, J.; Jeon, M.; Jung, T.; Lee, S.K.; Bae, S.Y.; Lee, H.C. Zerumbone suppresses EGF-induced CD44 expression through the inhibition of STAT3 in breast cancer cells. Oncol. Rep. 2014, 32, 2666–2672. [Google Scholar] [CrossRef]
- Eid, E.E.; Bustamam Abdul, A.; Rasedee, A.; Suliman, F.E.O.; Sukari, M.A.; Fatah, S.A. Liquid chromatography–tandem mass spectroscopic method for the determination of zerumbone in human plasma and its application to pharmacokinetics. J. Mass Spectrom. 2011, 46, 772–781. [Google Scholar] [CrossRef]
- Snyder, E.L.; Dowdy, S.F. Cell penetrating peptides in drug delivery. Pharm. Res. 2004, 21, 389–393. [Google Scholar] [CrossRef]
- Karagiannis, E.D.; Urbanska, A.M.; Sahay, G.; Pelet, J.M.; Jhunjhunwala, S.; Langer, R.; Anderson, D.G. Rational design of a biomimetic cell penetrating peptide library. Acs Nano 2013, 7, 8616–8626. [Google Scholar] [CrossRef][Green Version]
- Yamashita, H.; Kato, T.; Oba, M.; Misawa, T.; Hattori, T.; Ohoka, N.; Tanaka, M.; Naito, M.; Kurihara, M.; Demizu, Y. Development of a Cell-penetrating Peptide that Exhibits Responsive Changes in its Secondary Structure in the Cellular Environment. Sci. Rep. 2016, 6, 33003. [Google Scholar] [CrossRef] [PubMed]
- Lundin, P.; Johansson, H.; Guterstam, P.; Holm, T.; Hansen, M.; Langel, Ü.; El Andaloussi, S. Distinct uptake routes of cell-penetrating peptide conjugates. Bioconjugate Chem. 2008, 19, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Tünnemann, G.; Martin, R.M.; Haupt, S.; Patsch, C.; Edenhofer, F.; Cardoso, M.C. Cargo-dependent mode of uptake and bioavailability of TAT-containing proteins and peptides in living cells. Faseb J. 2006, 20, 1775–1784. [Google Scholar]
- Raucher, D.; Ryu, J.S. Cell-penetrating peptides: Strategies for anticancer treatment. Trends Mol. Med. 2015, 21, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Regberg, J.; Srimanee, A.; Langel, Ü. Applications of cell-penetrating peptides for tumor targeting and future cancer therapies. Pharmaceuticals 2012, 5, 991–1007. [Google Scholar] [CrossRef] [PubMed]
- Heitz, F.; Morris, M.C.; Divita, G. Twenty years of cell-penetrating peptides: From molecular mechanisms to therapeutics. Br. J. Pharmacol. 2009, 157, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Sarko, D.; Beijer, B.; Boy, R.G.; Nothelfer, E.-M.; Leotta, K.; Eisenhut, M.; Altmann, A.; Haberkorn, U.; Mier, W. The pharmacokinetics of cell-penetrating peptides. Mol. Pharm. 2010, 7, 2224–2231. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, D.M.; Langel, K.; Eriste, E.; Langel, U. Cell-penetrating peptides: Design, synthesis, and applications. Acs Nano 2014, 8, 1972–1994. [Google Scholar] [CrossRef]
- Bolhassani, A. Potential efficacy of cell-penetrating peptides for nucleic acid and drug delivery in cancer. Biochim. Et Biophys. Acta (Bba)-Rev. Cancer 2011, 1816, 232–246. [Google Scholar] [CrossRef]
- Lehto, T.; Kurrikoff, K.; Langel, Ü. Cell-penetrating peptides for the delivery of nucleic acids. Expert Opin. Drug Deliv. 2012, 9, 823–836. [Google Scholar] [CrossRef]
- Sugahara, K.N.; Teesalu, T.; Karmali, P.P.; Kotamraju, V.R.; Agemy, L.; Greenwald, D.R.; Ruoslahti, E. Coadministration of a tumor-penetrating peptide enhances the efficacy of cancer drugs. Science 2010, 328, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Bechara, C.; Sagan, S. Cell-penetrating peptides: 20 years later, where do we stand? Febs Lett. 2013, 587, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, Q.; Yu, Z.; He, Q. Cell-penetrating peptide-based intelligent liposomal systems for enhanced drug delivery. Curr. Pharm. Biotechnol. 2014, 15, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Kandela, I.; Chou, J.; Chow, K. Registered report: Coadministration of a tumor-penetrating peptide enhances the efficacy of cancer drugs. eLife 2015, 4, e06959. [Google Scholar] [CrossRef] [PubMed]
- Lao, X.; Li, B.; Liu, M.; Chen, J.; Gao, X.; Zheng, H. Increased antitumor activity of tumor-specific peptide modified thymopentin. Biochimie 2014, 107, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Shi, N.-Q.; Gao, W.; Xiang, B.; Qi, X.-R. Enhancing cellular uptake of activable cell-penetrating peptide-doxorubicin conjugate by enzymatic cleavage. Int. J. Nanomed. 2012, 7, 1613–1621. [Google Scholar]
- Aroui, S.; Brahim, S.; De Waard, M.; Kenani, A. Cytotoxicity, intracellular distribution and uptake of doxorubicin and doxorubicin coupled to cell-penetrating peptides in different cell lines: A comparative study. Biochem. Biophys. Res. Commun. 2010, 391, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Cun, X.; Chen, J.; Ruan, S.; Zhang, L.; Wan, J.; He, Q.; Gao, H. A novel strategy through combining iRGD peptide with tumor-microenvironment-responsive and multistage nanoparticles for deep tumor penetration. Acs Appl. Mater. Interfaces 2015, 7, 27458–27466. [Google Scholar] [CrossRef]
- Hu, C.; Yang, X.; Liu, R.; Ruan, S.; Zhou, Y.; Xiao, W.; Yu, W.; Yang, C.; Gao, H. Coadministration of iRGD with multistage responsive nanoparticles enhanced tumor targeting and penetration abilities for breast cancer therapy. Acs Appl. Mater. Interfaces 2018, 10, 22571–22579. [Google Scholar] [CrossRef]
- Tang, E.L.; Rajarajeswaran, J.; Fung, S.Y.; Kanthimathi, M. Antioxidant activity of Coriandrum sativum and protection against DNA damage and cancer cell migration. BMC Complement. Altern. Med. 2013, 13, 347. [Google Scholar] [CrossRef]
- Ciapetti, G.; Granchi, D.; Savarino, L.; Cenni, E.; Magrini, E.; Baldini, N.; Giunti, A. In vitro testing of the potential for orthopedic bone cements to cause apoptosis of osteoblast-like cells. Biomaterials 2002, 23, 617–627. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Huey, R.; Olson, A.J. Distributed automated docking of flexible ligands to proteins: Parallel applications of AutoDock 2.4. J. Comput.-Aided Mol. Des. 1996, 10, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.J.; Chow, D.E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D. Scalable Algorithms for Molecular Dynamics Simulations on Commodity Clusters. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006. [Google Scholar]
- Desmond Molecular Dynamics System. Maestro-Desmond Interoperability Tools; Schrödinger: New York, NY, USA, 2016. [Google Scholar]
- Harder, E.; Damm, W.; Maple, J.; Wu, C.; Reboul, M.; Xiang, J.Y.; Wang, L.; Lupyan, D.; Dahlgren, M.K.; Knight, J.L. OPLS3: A force field providing broad coverage of drug-like small molecules and proteins. J. Chem. Theory Comput. 2015, 12, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695. [Google Scholar] [CrossRef]
- Martyna, G.J.; Tobias, D.J.; Klein, M.L. Constant pressure molecular dynamics algorithms. J. Chem. Phys. 1994, 101, 4177–4189. [Google Scholar] [CrossRef]
- Tuckerman, M.; Berne, B.J.; Martyna, G.J. Reversible multiple time scale molecular dynamics. J. Chem. Phys. 1992, 97, 1990–2001. [Google Scholar] [CrossRef]
- Cheng, Y.; Ji, Y. RGD-modified polymer and liposome nanovehicles: Recent research progress for drug delivery in cancer therapeutics. Eur. J. Pharm. Sci. 2018, 128, 8–17. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Mao, X.; Liu, J.; Gong, Z.; Zhang, H.; Lu, Y.; Zou, H.; Yu, Y.; Chen, Y.; Sun, Z.; Li, W. iRGD-conjugated DSPE-PEG2000 nanomicelles for targeted delivery of salinomycin for treatment of both liver cancer cells and cancer stem cells. Nanomedicine 2015, 10, 2677–2695. [Google Scholar] [CrossRef]
- Eid, E.E.; Azam, F.; Hassan, M.; Taban, I.M.; Halim, M.A. Zerumbone binding to estrogen receptors: An in-silico investigation. J. Recept. Signal. Transduct. 2018, 38, 342–351. [Google Scholar] [CrossRef]
- Azam, F.; Alabdullah, N.H.; Ehmedat, H.M.; Abulifa, A.R.; Taban, I.; Upadhyayula, S. NSAIDs as potential treatment option for preventing amyloid β toxicity in Alzheimer’s disease: An investigation by docking, molecular dynamics, and DFT studies. J. Biomol. Struct. Dyn. 2018, 36, 2099–2117. [Google Scholar] [CrossRef] [PubMed]
- Maiorov, V.N.; Crippen, G.M. Significance of root-mean-square deviation in comparing three-dimensional structures of globular proteins. J. Mol. Biol. 1994, 235, 625–634. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | ZER | ZER-HPβCD | TP5-iRGD Peptide | |||||
|---|---|---|---|---|---|---|---|---|
| 24 h IC50 (µg/mL) | 48 h IC50 (µg/mL) | 72 h IC50 (µg/mL) | 24 h IC50 (µg/mL) | 48 h IC50 (µg/mL) | 72 h IC50 (µg/mL) | 48 h IC50 (µg/mL) | 72 h IC50 (µg/mL) | |
| MCF7 | 17.36 ± 2.3 | 9.06 ± 1.0 | 7.51 ± 0.2 | 20.81 ± 9.3 | 4.43 ± 0.2 | 5.08 ± 0.2 | >1000 | >1000 |
| MDA-MB-231 | 50.93 ± 3.0 | 23.44 ± 10.0 | 14.96 ± 1.5 | 40.73 ± 1.9 | 21.1 ± 6.7 | 12.18 ± 0.7 | >1000 | >1000 |
| Hs27 | >100 | >100 | >100 | 42.99 ± 2.3 | 46.14 ± 2.4 | 21.45 ± 1.3 | >1000 | >1000 |
| Co-administration Ratio | MCF-7 (% of Inhibition) | MDA-MB-231 (% of Inhibition) | ||
|---|---|---|---|---|
| Compound (IC50): TP5-iRGD | ZER | ZER-HPβCD | ZER | ZER-HPβCD |
| 1:10 X | 91.4 ± 3.0 | 90.4 ± 4.2 | 58.1 ± 8.8 | 41.5 ± 20.4 |
| 1:20 X | 67.6 ± 3.9 | 85.8 ± 7.0 | 44.0 ± 8.6 | 72.8 ± 2.3 |
| 1:30 X | 72.1 ± 8.6 | 68.5 ± 13.2 | 48.3 ± 5.4 | 79.1 ± 1.8 |
| Co-Administration | MCF-7–IC50 (µg/mL) |
|---|---|
| ZER with TP5-iRGD | 3.13 ± 0.7 |
| ZER-HPβCDwith TP5-iRGD | 0.49 ± 0.004 |
| Parameters | ZER | ZER in the Presence of TP5-iRGD |
|---|---|---|
| Binding Energy, ΔGb (kcal/mol) | −6.77 | −8.13 |
| Predicted Ki (µM) | 10.97 | 1.10 |
| RMSD | 4.39 | 4.16 |
| Residues involved in hydrophobic interactions | Met-180(B), Arg-214(B) | Tyr-122(B), Tyr-166(B), Tyr-178(A), Arg-214(B), Arg-216(B), Ala-218(B) |
| Residues involved in hydrophilic interactions | Nil | Nil |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
E. M. Eid, E.; S. Alanazi, A.; Koosha, S.; A. Alrasheedy, A.; Azam, F.; M. Taban, I.; Khalilullah, H.; Sadiq Al-Qubaisi, M.; A. Alshawsh, M. Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules 2019, 24, 2554. https://doi.org/10.3390/molecules24142554
E. M. Eid E, S. Alanazi A, Koosha S, A. Alrasheedy A, Azam F, M. Taban I, Khalilullah H, Sadiq Al-Qubaisi M, A. Alshawsh M. Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules. 2019; 24(14):2554. https://doi.org/10.3390/molecules24142554
Chicago/Turabian StyleE. M. Eid, Eltayeb, Abdulrahman S. Alanazi, Sanaz Koosha, Alian A. Alrasheedy, Faizul Azam, Ismail M. Taban, Habibullah Khalilullah, Mothanna Sadiq Al-Qubaisi, and Mohammed A. Alshawsh. 2019. "Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide" Molecules 24, no. 14: 2554. https://doi.org/10.3390/molecules24142554
APA StyleE. M. Eid, E., S. Alanazi, A., Koosha, S., A. Alrasheedy, A., Azam, F., M. Taban, I., Khalilullah, H., Sadiq Al-Qubaisi, M., & A. Alshawsh, M. (2019). Zerumbone Induces Apoptosis in Breast Cancer Cells by Targeting αvβ3 Integrin upon Co-Administration with TP5-iRGD Peptide. Molecules, 24(14), 2554. https://doi.org/10.3390/molecules24142554