
Effect of Different Anthocyanidin Glucosides on Lutein Uptake by Caco-2 Cells, and Their Combined Activities on Anti-Oxidation and Anti-Inflammation In Vitro and Ex Vivo


Abstract
1. Introduction
2. Results and Discussion
2.1. Effects of Lutein-Anthocyanin Combinations on Oxidative Inhibition in Chemical and Cellular Models
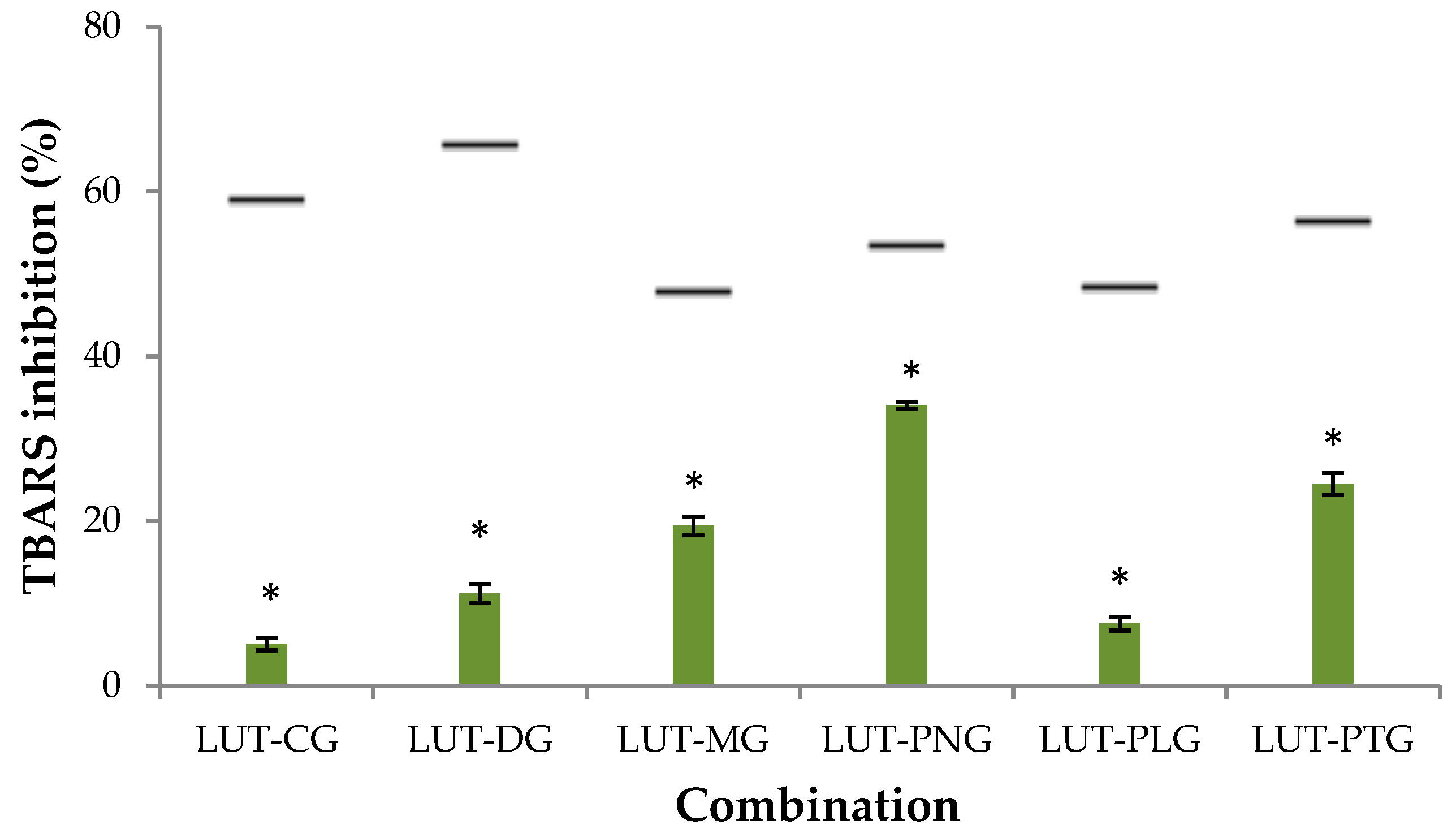
2.1.1. Inhibitory Effect on Liposome Peroxidation
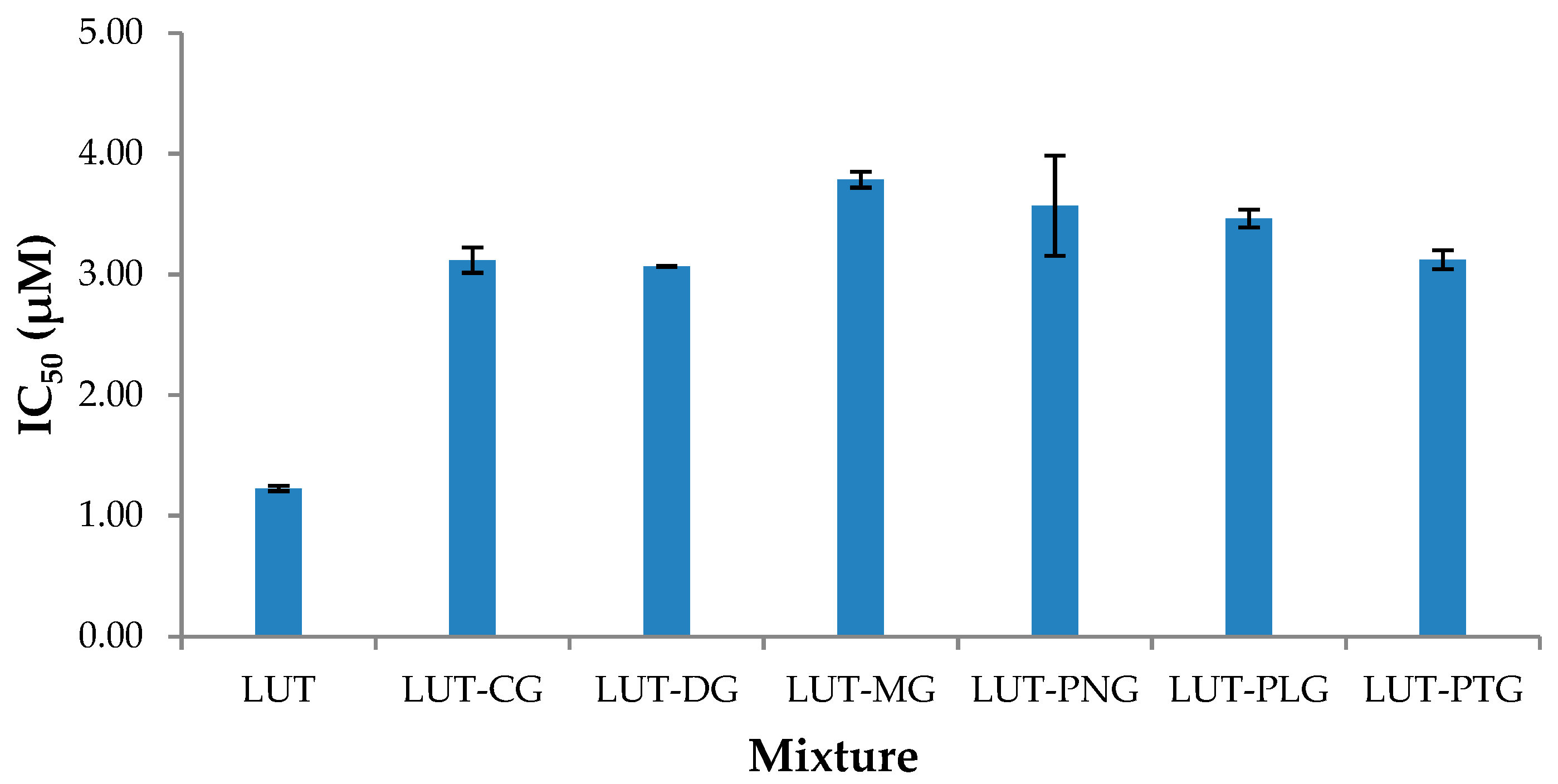
2.1.2. Cellular Antioxidant Activity (CAA)
2.2. Effects of Lutein-Anthocyanin Combinations on Anti-Inflammation in Chemical and Cellular Models
2.2.1. Lipoxygenase Inhibitory Activity
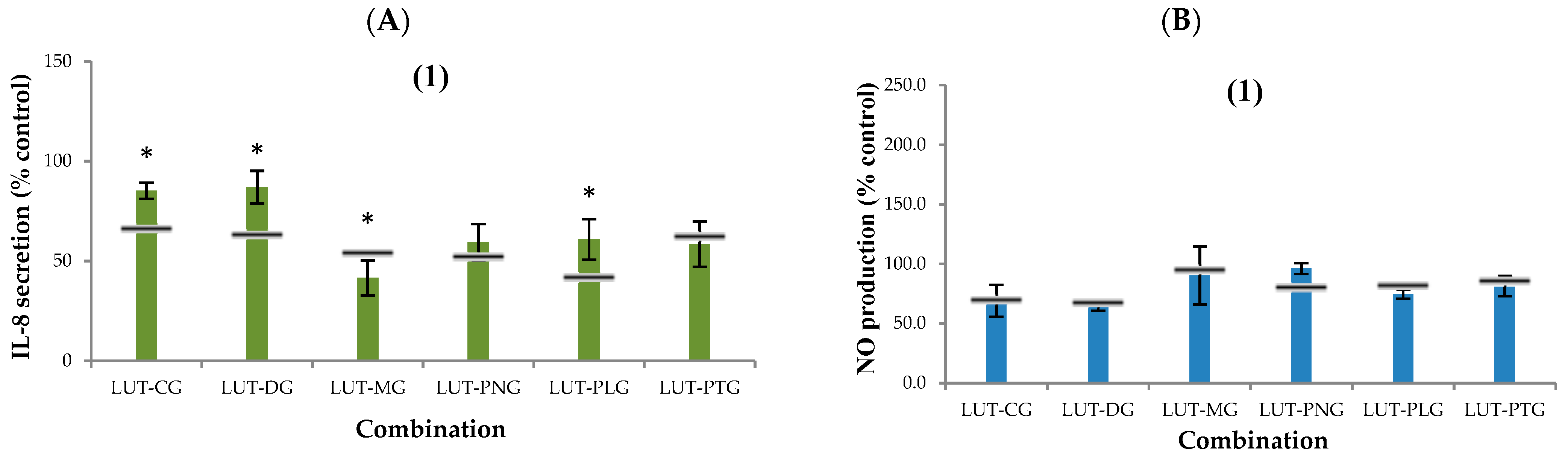
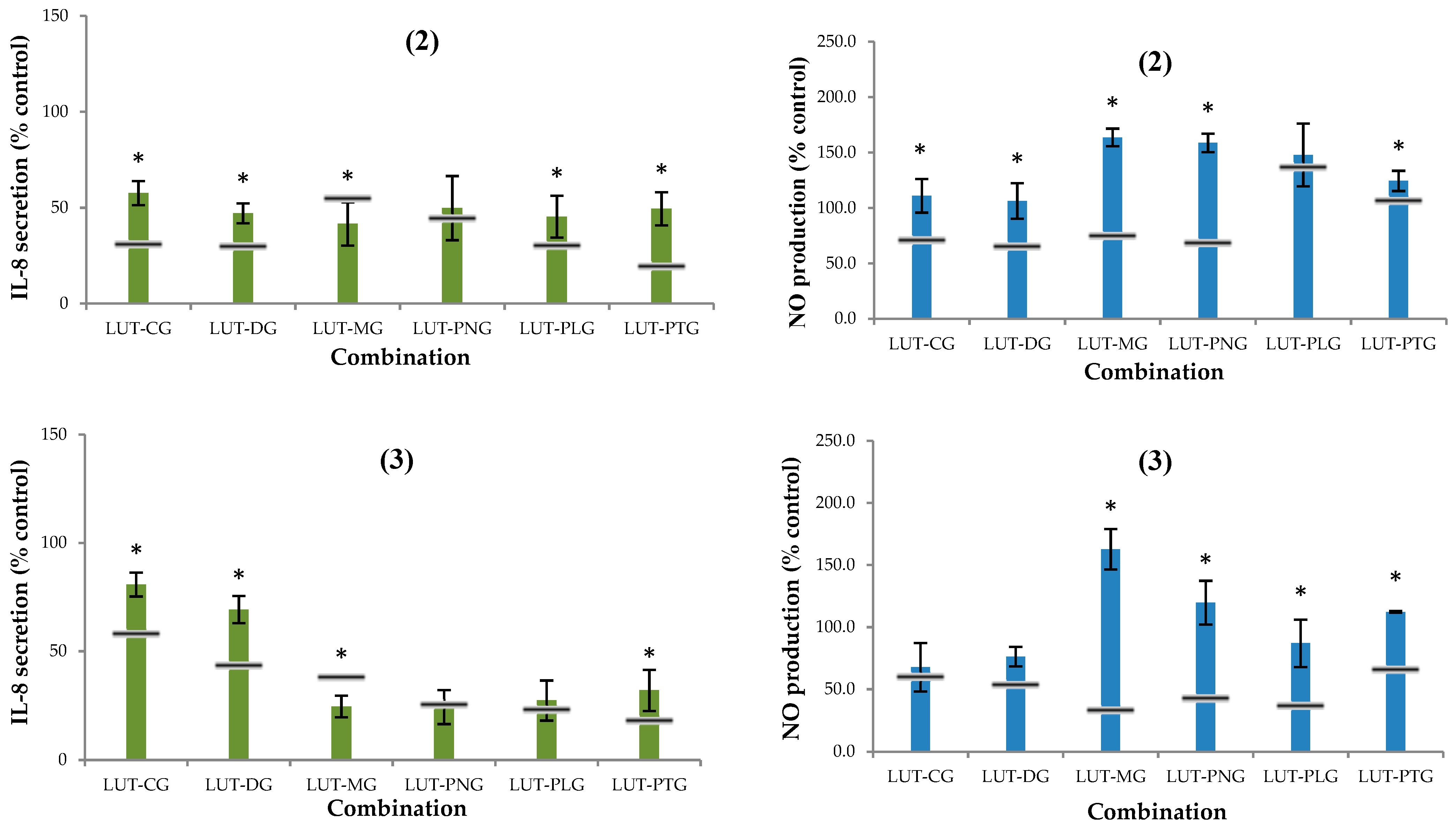
2.2.2. Secretion of Interleukin-8 (IL-8)
2.2.3. Nitric Oxide (NO) Production
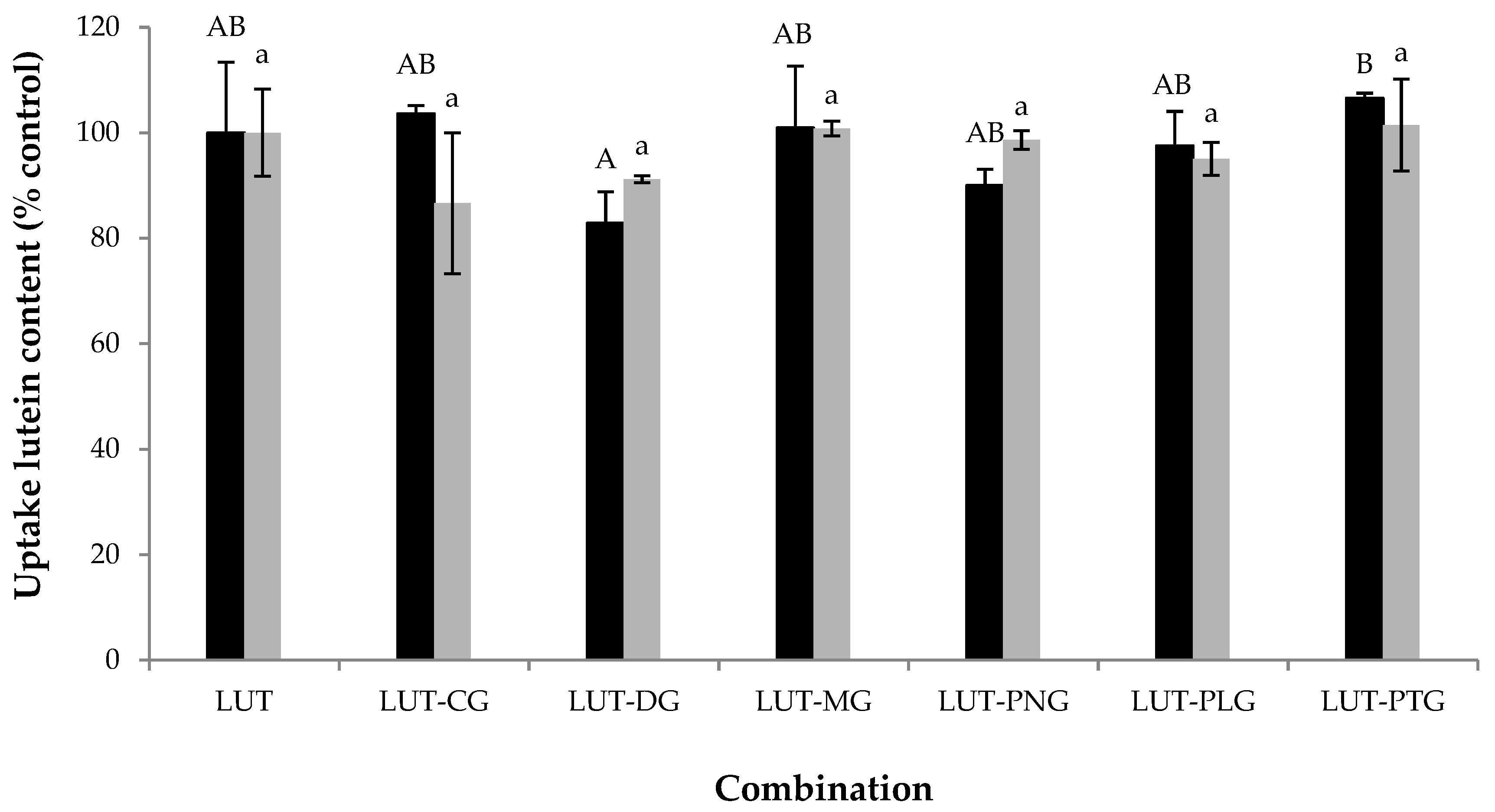
2.3. Interferences of Anthocyanins on Lutein Uptake by Caco-2 Cells
3. Materials and Methods
3.1. Materials
3.2. Phytochemical Stock Preparation
3.3. Inhibition of Liposome Peroxidation
3.4. In Vitro Anti-Inflammatory Assay: Lipoxygenase Inhibition
3.5. General Cell Culture Conditions
3.6. Cell Viability
3.7. Cellular Antioxidant Assay
- AUCs is the integrated area under the sample fluorescence versus time curve;
- AUCc is the integrated area under the control fluorescence versus time curve;
- Fluorescence excitation was measured at 485 nm and fluorescence emission was measured at 520 nm every 5 min for 12 cycles at 37 °C.
3.8. TNF-α-Induced Inflammation
3.9. Cellular Uptake of Lutein
3.10. Extraction of Lutein from Cell Lysate
3.11. Lutein Analysis by LC-MS
3.12. Mode of Interaction Determination
- Synergy: the experimental inhibitory activity is greater than the expected activity;
- Antagonism: the experimental inhibitory activity is lesser than the expected activity;
- Addition: the experimental inhibitory activity is equal to the expected activity.
3.13. Data Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reboul, E.; Thap, S.; Tourniaire, F.; André, M.; Juhel, C.; Morange, S.; Amiot, M.J.; Lairon, D.; Borel, P. Differential effect of dietary antioxidant classes (carotenoids, polyphenols, vitamins C and E) on lutein absorption. Br. J. Nutr. 2007, 97, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.Q.; Jiang, P.F.; Gao, Y.Z. Lutein prevents osteoarthritis through Nrf2 activation and downregulation of inflammation. Arch. Med. Sci. AMS 2018, 14, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Thurnham, D.I. Macular zeaxanthins and lutein—A review of dietary sources and bioavailability and some relationships with macular pigment optical density and age-related macular disease. Nutr. Res. Rev. 2007, 20, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C. A comprehensive review of eggs, choline, and lutein on cognition across the life-span. J. Am. Coll. Nutr. 2018, 37, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Bueno, J.M.; Ramos-Escudero, F.; Sáez-Plaza, P.; Muñoz, A.M.; José Navas, M.; Asuero, A.G. Analysis and antioxidant capacity of anthocyanin pigments. Part I: General Considerations Concerning Polyphenols and Flavonoids. Crit. Rev. Anal. Chem. 2012, 42, 102–125. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Paterson, J.; Bucknall, M.; Arcot, J. Interactions between phytochemicals from fruits and vegetables: Effects on bioactivities and bioavailability. Crit. Rev. Food Sci. Nutr. 2018, 58, 1310–1329. [Google Scholar] [CrossRef] [PubMed]
- Phan, M.A.T.; Bucknall, M.; Arcot, J. Interactive effects of β-carotene and anthocyanins on cellular uptake, antioxidant activity and anti-inflammatory activity in vitro and ex vivo. J. Funct. Foods 2018, 45, 129–137. [Google Scholar] [CrossRef]
- Han, R.M.; Zhang, J.P.; Skibsted, L.H. Reaction dynamics of flavonoids and carotenoids as antioxidants. Molecules 2012, 17, 2140–2160. [Google Scholar] [CrossRef] [PubMed]
- Bonarska-Kujawa, D.; Pruchnik, H.; Kleszczyńska, H. Interaction of selected anthocyanins with erythrocytes and liposome membranes. Cell. Mol. Biol. Lett. 2012, 17, 289. [Google Scholar] [CrossRef] [PubMed]
- Song, L.L.; Liang, R.; Li, D.D.; Xing, Y.D.; Han, R.M.; Zhang, J.P.; Skibsted, L.H. Beta-carotene radical cation addition to green tea polyphenols. Mechanism of antioxidant antagonism in peroxidizing liposomes. J. Agric. Food Chem. 2011, 59, 12643–12651. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Chen, C.H.; Han, R.M.; Zhang, J.P.; Skibsted, L.H. Thermodynamic versus kinetic control of antioxidant synergism between β-carotene and (iso) flavonoids and their glycosides in liposomes. J. Agric. Food Chem. 2010, 58, 9221–9227. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Volkova, N.; Rosenblat, M.; Aviram, M. Lycopene synergistically inhibits LDL oxidation in combination with vitamin E, glabridin, rosmarinic acid, carnosic acid, or garlic. Antioxid. Redox 2000, 2, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, F. Dietary antioxidant synergy in chemical and biological systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2343–2357. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhu, F.; Meckling, K.A.; Marcone, M.F. Antioxidant capacity of food mixtures is not correlated with their antiproliferative activity against MCF-7 breast cancer cells. J. Med. Food 2013, 16, 1138–1145. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, S.; Krishnaswamy, S.; Devashya, V.; Sethuraman, S.; Krishnan, U.M. Influence of membrane lipid composition on flavonoid–membrane interactions: Implications on their biological activity. Prog. Lipid Res. 2015, 58, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Knaup, B.; Oehme, A.; Valotis, A.; Schreier, P. Anthocyanins as lipoxygenase inhibitors. Mol. Nutr. Food Res. 2009, 53, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Serpen, A.; Gökmen, V. Effects of β-carotene on soybean lipoxygenase activity: Kinetic studies. Eur. Food Res. Technol. 2006, 224, 743–748. [Google Scholar] [CrossRef]
- Tarahovsky, Y.S.; Muzafarov, E.N.; Kim, Y.A. Rafts making and rafts braking: How plant flavonoids may control membrane heterogeneity. Mol. Cell. Biochem. 2008, 314, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Sampson, J.; Candeias, L.P.; Bramley, P.M.; Rice-Evans, C.A. Antioxidant activities of carotenes and xanthophylls. FEBS Lett. 1996, 384, 240–242. [Google Scholar] [CrossRef]
- Roberts, W.G.; Gordon, M.H. Determination of the total antioxidant activity of fruits and vegetables by a liposome assay. J. Agric. Food Chem. 2003, 51, 1486–1493. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Xue, J.; Abbas, S.; Feng, B.; Zhang, X.; Xia, S. Liposome as a delivery system for carotenoids: Comparative antioxidant activity of carotenoids as measured by ferric reducing antioxidant power, DPPH assay and lipid peroxidation. J. Agric. Food Chem. 2014, 62, 6726–6735. [Google Scholar] [CrossRef] [PubMed]
- Durak, A.; Gawlik-Dziki, U.; Pecio, Ł. Coffee with cinnamon—Impact of phytochemicals interactions on antioxidant and anti-inflammatory in vitro activity. Food Chem. 2014, 162, 81–88. [Google Scholar] [CrossRef] [PubMed]
- O’sullivan, S.M.; Woods, J.A.; O’brien, N.M. Use of Tween 40 and Tween 80 to deliver a mixture of phytochemicals to human colonic adenocarcinoma cell (CaCo-2) monolayers. Br. J. Nutr. 2004, 91, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Zhang, H.; Liu, R.; Mine, Y.; McCallum, J.; Kirby, C.; Tsao, R. Antioxidant and anti-inflammatory activities of pyranoanthocyanins and other polyphenols from staghorn sumac (Rhus hirta L.) in Caco-2 cell models. J. Funct. Foods 2016, 20, 139–147. [Google Scholar] [CrossRef]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Biehler, E.; Hoffmann, L.; Krause, E.; Bohn, T. Divalent minerals decrease micellarization and uptake of carotenoids and digestion products into Caco-2 cells. J. Nutr. 2011, 141, 1769–1776. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds: lutein, cyanidin-3-O-glucoside chloride, delphinidin-3-O-glucoside cholride, pelargonidin-3-O-glucoside chloride, malvidin-3-O-glucoside chloride, peonidin-3-O-glucoside chloride, and petunidin-3-O-glucoside chloride are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mixture | Lutein: Anthocyanin Ratio | |||||
|---|---|---|---|---|---|---|
| 1:3 | 1:1 | 3:1 | ||||
| Experimental Effect 1 | Expected Additive Effect 2 | Experimental Effect | Expected Additive Effect | Experimental Effect | Expected Additive Effect | |
| LUT-CG | 38.1 ± 12.5 | 52.1 | 36.9 ± 12.8 | 52.1 | 46.3 ± 7.5 | 45.2 |
| LUT-DG | 48.2 ± 12.3 | 49.4 | 45.5 ± 5.9 | 36.4 | 44.1 ± 5.3 | 44.0 |
| LUT-MG | 50.8 ± 7.1 | 48.1 | 44.0 ± 3.1 | 42.5 | 50.3 ± 6.8 | 50.4 |
| LUT-PNG | 42.9 ± 6.9 | 43.9 | 33.6 ± 8.8 | 38.1 | 48.7 ± 2.4 | 46.8 |
| LUT-PLG | 38.6 ± 4.8 | 44.3 | 32.5 ± 8.6 | 38.9 | 49.2 ± 4.0 | 45.8 |
| LUT-PTG | 38.2 ± 6.5 | 42.6 | 34.6 ± 11.5 | 39.2 | 47.6 ± 5.9 | 45.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phan, M.A.T.; Bucknall, M.; Arcot, J. Effect of Different Anthocyanidin Glucosides on Lutein Uptake by Caco-2 Cells, and Their Combined Activities on Anti-Oxidation and Anti-Inflammation In Vitro and Ex Vivo. Molecules 2018, 23, 2035. https://doi.org/10.3390/molecules23082035
Phan MAT, Bucknall M, Arcot J. Effect of Different Anthocyanidin Glucosides on Lutein Uptake by Caco-2 Cells, and Their Combined Activities on Anti-Oxidation and Anti-Inflammation In Vitro and Ex Vivo. Molecules. 2018; 23(8):2035. https://doi.org/10.3390/molecules23082035
Chicago/Turabian StylePhan, Minh Anh Thu, Martin Bucknall, and Jayashree Arcot. 2018. "Effect of Different Anthocyanidin Glucosides on Lutein Uptake by Caco-2 Cells, and Their Combined Activities on Anti-Oxidation and Anti-Inflammation In Vitro and Ex Vivo" Molecules 23, no. 8: 2035. https://doi.org/10.3390/molecules23082035
APA StylePhan, M. A. T., Bucknall, M., & Arcot, J. (2018). Effect of Different Anthocyanidin Glucosides on Lutein Uptake by Caco-2 Cells, and Their Combined Activities on Anti-Oxidation and Anti-Inflammation In Vitro and Ex Vivo. Molecules, 23(8), 2035. https://doi.org/10.3390/molecules23082035