Ascorbic Acid and Glucosinolate Levels in New Czech Cabbage Cultivars: Effect of Production System and Fungal Infection

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Effect of Production System on AA Contents
2.2. Contents of GSLs in Dependence on the Production System
2.3. GSL Composition Profiles of Albatros and Target cv.
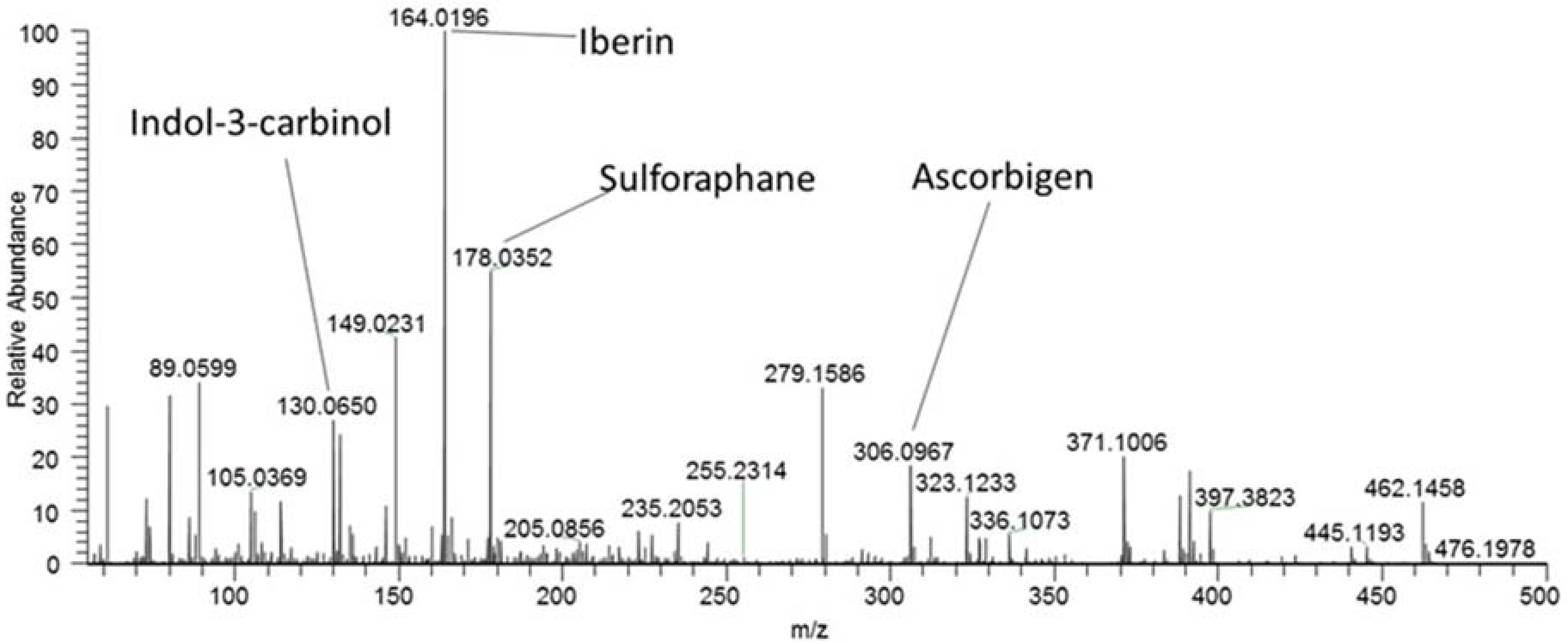
2.4. Degradation Products of the Spontaneous Enzymatic Hydrolysis of GSL
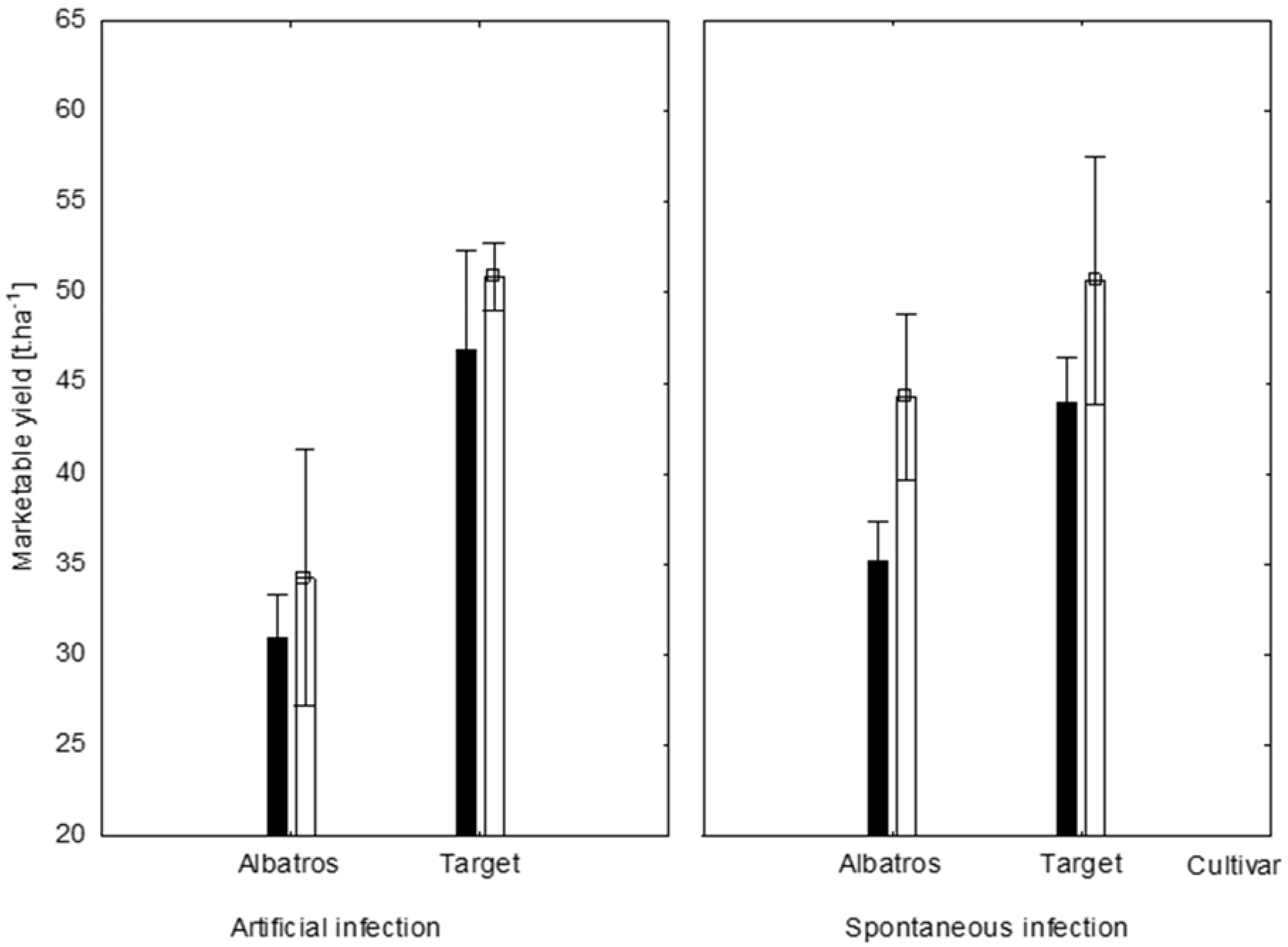
2.5. Effect of Production Systems and Fungal Infection on Albatros and Target cv. Yields
3. Materials and Methods
3.1. Biological Material and Chemicals
3.2. AA Analysis
3.3. GLS Analysis
3.4. Degradation Products Analysis
3.5. Agricultural Production Systems
3.6. Fungal Infection and Disease Assessment
3.7. Statistical Evaluation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Disclaimer
References
- Temple, N.J. Antioxidants and disease: More questions than answers. Nutr. Res. 2000, 20, 449–459. [Google Scholar] [CrossRef]
- Podsedek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT-Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Majkowska-Gadomska, J.; Wierzbicka, B. Content of Basic Nutrients and Minerals in Heads of Selected Varieties of Red Cabbage (Brasicca oleracea var. capitata f. rubra). Pol. J. Environ. Stud. 2008, 17, 295–298. [Google Scholar]
- Trdan, S.; Vali, N.; Andjus, L.; Vovk, I.; Martelanc, M.; Simonovska, B.; Jerman, J.; Vidrih, R.; Vidrih, M.; Žnidar, D. Which plant compounds influence the natural resistance of cabbage against onion thrips (Thrips tabaci Lindeman). Acta Phytopathol. Entomol. Hung. 2008, 43, 385–395. [Google Scholar] [CrossRef]
- Koudela, M.; Petrikova, K. Nutrients content and yield in selected cultivars of leaf lettuce (Lactuca sativa L. var. crispa). Hortic. Sci. 2008, 35, 99–106. [Google Scholar] [CrossRef]
- Penas, E.; Frias, J.; Matrinez-Villaluenga, C.; Vidal-Valverde, C. Bioactive compounds, myrosinase aktivity, and antioxidant capacity of white cabbages grown in different locations of Spain. J. Agric. Food Chem. 2011, 20, 3772–3779. [Google Scholar] [CrossRef] [PubMed]
- Vogtmann, H.; Matthies, K.; Kehres, B.; Meier-Ploeger, A. Enhanced food quality induced by compost applications. In Science and Engineering of Composting; Hoitnik, H.A.J., Keener, H.M., Eds.; Renaissance: Worthington, OH, USA, 1993; pp. 645–667. ISBN 09-366-45156. [Google Scholar]
- Franczuk, J.; Zaniewicz-Bajkowska, A.; Kosterna, E.; Rosa, R.; Pniewska, I.; Olszewski, W. The effect of cover plants on the yield and content of selected components of cabbage varieties. Acta Scientiarum Polonorum. Hortorum Cultus 2010, 9, 23–30. [Google Scholar]
- Bavec, M.; Turinek, M.; Mlakar, S.G.; Mikola, N.; Bavec, F. Some internal quality properties of white cabbage from different farming systems. In ISHS Acta Horticulturae 933, Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on Organic Horticulture: Productivity and Sustainability, Lisbon, Portugal, 22–27 August 2010; Mourao, I., Aksoy, U., Eds.; International Society for Horticultural Science: Leuven, Belgium, 2012; Volume 933, pp. 577–583, ISBN 978-90-66053-97-7. [Google Scholar]
- Castro, A.; Aires, A.; Rosa, E.; Bloem, E.; Stulen, I.; De Kok, L.J. Distribution of glucosinolates in Brassica oleracea cultivars. Phyton 2004, 44, 133–143. [Google Scholar]
- Haneklaus, S.; Bloem, E.; Schnug, E. Disease control by sulfur induced resistence. Asp. Appl. Biol. 2006, 79, 221–224. [Google Scholar]
- Haneklaus, S.; Bloem, E.; Schnug, E. Sulfur interactions in crop ecosystems. In Sulfur in Plants an Ecological Perspective; Hawkesford, M.J., De Kok, L.J., Eds.; Springer Science & Business Media: Berlin, Germany, 2007; pp. 17–58. ISBN 9781402058875. [Google Scholar]
- Antonious, G.; Bomford, M.; Vincelli, P. Screening Brassica species for glucosinolate content. J. Environ. Sci. Health Part B 2009, 44, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, N.M.; Tytgat, T.O.G.; Kirkegaard, J.A. Root and shoot glucosinolates: A comparison of their diversity, function and interactions in natural and managed ecosystems. Phytochem. Rev. 2009, 8, 171–186. [Google Scholar] [CrossRef]
- Yan, X.; Chen, S. Regulation of plant glucosinolate metabolism. Planta 2007, 226, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Variyar, P.S.; Banerjee, A.; Akkarakaran, J.J.; Suprasanna, P. Role of glucosinolates in plant stress tolerance. In Emerging Technologies and Management of Crop Stress Tolerance; Ahmad, P., Rasool, S., Eds.; Elsevier Inc.: New York, NY, USA, 2014; Volume 1, pp. 271–291. ISBN 978-0-12-800876-8. [Google Scholar]
- Bellostas, N.; Sorensen, A.D.; Sorensen, J.C.; Sorensen, H. Genetic variation and metabolism of glucosinolates. Adv. Bot. Res. 2007, 45, 369–415. [Google Scholar] [CrossRef]
- Burrow, M.; Zhang, Z.Y.; Ober, J.A.; Lambrix, V.M.; Wittstock, U.; Gershenzon, J.; Kliebenstein, D.J. ESP and ESM1 mediate indol-3-acetonitrile production from indol-3-ylmethyl glucosinolate in Arabidopsis. Phytochemistry 2008, 69, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Ciska, E.; Martyniak-Przybyszewska, B.; Kozlowska, H. Content of glucosinolates in cruciferous vegetables grown at the same site for two years under different climatic conditions. J. Agric. Food Chem. 2000, 48, 2862–2867. [Google Scholar] [CrossRef] [PubMed]
- Mayton, H.S.; Oliver, C.; Vaughn, S.F.; Loria, R. Correlation of fungicidal aktivity of Brassica species with allyl isothiocyanate production in macerated leaf tissue. Phytopathology 1996, 86, 267–271. [Google Scholar] [CrossRef]
- Brader, G.; Tas, E.; Palva, E.T. Jasmonate-dependent induction of indole glucosinolates in Arabidopsis by culture filtrates of the nonspecific pathogen Erwinia carotovora. Plant Physiol. 2001, 126, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Sorensen, J.L. Phytoalexin accumulation and production of antifungal compounds by the crucifer wasabi. Phytochemistry 1998, 49, 1959–1965. [Google Scholar] [CrossRef]
- Tierens, K.; Thomma, B.P.H.; Brouwer, M.; Schmidt, J.; Kistner, K.; Porzel, A.; Mauch-Mani, B.; Cammue, B.P.A.; Broekaert, W.F. Study of the role ofantimicrobial glucosinolate-derived isothiocyanates in resistence of Arabidopsis to microbial pathogens. Plant Physiol. 2001, 125, 1688–1699. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Yamada, K.; Kosemura, S.; Yamamura, S.; Hasegawa, K. Phototropic stimulation induces the conversion of glucosinolate to phototropism-regulating substances of radish hypocotyls. Phytochemistry 2000, 54, 275–279. [Google Scholar] [CrossRef]
- Mithen, R.F. Glucosinolates and their degradation products. Adv. Bot. Res. 2001, 35, 213–262. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Warman, P.R.; Havard, K.A. Yield, vitamin and mineral contents of organically and conventionally grown carrots and cabbage. Agric. Ecosyst. Environ. 1997, 61, 155–162. [Google Scholar] [CrossRef]
- Fjelkner-Modig, S.; Bengtsson, H.; Nyström, S.; Stegmark, R. The influence of organic and integrated production on nutritional, sensory and agricultural aspects of vegetable raw materials for food production. Acta Agric. Scand. Sect. B Plant Soil Sci. 2010, 50, 102–113. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Leszczynska, T. Comparison of vitamin C and ß-carotene in cruciferous vegetables grown in diversified ecological conditions. Pol. J. Environ. Stud. 2013, 22, 167–173. [Google Scholar]
- Park, S.; Arasu, M.V.; Lee, M.-K.; Chun, J.-H.; Seo, J.M.; Lee, S.-W.; Al-Dhabi, N.A.; Kim, S.-J. Quiantification of glucosinolates, anthocyanins, free amino acids, and vitamin C in inbred lines of cabbage (Brassica oleracea L.). Food Chem. 2014, 145, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Leon, A.M.; Lozano, M.; Gonzalez, D.; Ayuso, M.C.; Fernandez-Leon, M.F. Bioactive compounds content and total antioxidant activity of two Savoy cabbages. Czech J. Food Sci. 2014, 32, 549–554. [Google Scholar] [CrossRef]
- Brotman, Y.; Landau, U.; Cuadros-Inostroza, A.; Takayuki, K.; Fernie, A.R.; Chet, I.; Viterbo, A.; Willmitzer, L. Trichoderma-plant root colonization: Escaping early plant defense responses and activation of the antioxidant machinery for saline stress tolerance. PLoS Pathog. 2013, 9, e1003221. [Google Scholar] [CrossRef]
- Mastouri, F.; Bjorkman, T.; Harman, G.E. Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistence to water deficit. Mol. Plant-Microbe Interact. 2012, 25, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Farid, I.B.; Jahangir, M.; Mustafa, N.R.; Van Dam, N.M.; Van den Hondel, C.A.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Glucosinolate profiling of Brassica rapa cultivars after infection by Leptosphaeria maculans and Fusarium oxysporum. Biochem. Syst. Ecol. 2010, 38, 612–620. [Google Scholar] [CrossRef]
- Kabouw, P.; Biere, A.; Van Der Putten, W.H.; Van Dam, N.M. Intra-specific differences in root and shoot gluosinolate profiles among white cabbage (Brassica oleracea var. capitata) cultivars. J. Agric. Food Chem. 2010, 58, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Vicas, S.I.; Teusdea, A.C.; Carbunar, M.; Socaci, S.A.; Socaciu, C. Glucosinolates profile and antioxidant capacity of Romanian Brassica vegetables obtained by organic and conventional agricultural practices. Plant Food Hum. Nutr. 2013, 68, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Kusznierewicz, B.; Bartoszek, A.; Wolska, L.; Drzewiecki, J.; Gormstern, S.; Namiesnik, J. Partial characterization of white cabbages (Brassica oleracea var. capitata f. alba) from different regions by glucosinolates, bioactive compounds, total antioxidant activities and proteins. LWT-Food Sci. Technol. 2008, 41, 1–9. [Google Scholar] [CrossRef]
- Charron, C.S.; Saxton, A.M.; Sams, C.E. Relationship of climate and genotype to seasonal variation in the glucosinolate-myrosinase system. I. Glucosinolate content in ten cultivars of Brassica oleracea grown in fall and spring seasons. J. Sci. Food Agric. 2005, 85, 671–681. [Google Scholar] [CrossRef]
- Wennberg, M.; Ekvall, J.; Olsson, K.; Nyman, M. Changes in carbohydrate and glucosinolate composition in white cabbage (Brassica oleracea var. capitata) during blanching and treatment with acetic acid. Food Chem. 2006, 95, 226–236. [Google Scholar] [CrossRef]
- Menard, R.; Larue, J.P.; Siluq, D.; Thouvenot, D. Glucosinolates in cauliflower as biochemical markers for resistence against downy mildew. Phytochemistry 1999, 52, 29–35. [Google Scholar] [CrossRef]
- Buxdorf, K.; Yaffe, H.; Barda, O.; Levy, M. The effects of glucosinolates and their breakdown products on necrotrophic fungi. PLoS ONE 2013, 8, e70771. [Google Scholar] [CrossRef] [PubMed]
- Aghajanzadehdivaei, T. Sulfur Metabolism, Glucosinolates and Fungal Resistence in Brassica. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2015; doi:10.1371/journal.pone.0070771. [Google Scholar]
- Fahey, J.; Zalcmann, A.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Wagner, A.E.; Rimbach, G. Ascorbigen: Chemistry, occurrence, and biologic properties. Clin. Dermatol. 2009, 27, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Hara, M.; Fukino, N.; Kakizaki, T.M.Y. Glucosinolate metabolism, functionality and breeding for the improvement of Brassicaceae vegetables. Breed. Sci. 2014, 64, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Pavlovič, I.; Salopek-Sondi, B. White cabbage (Brassica oleracea var. capitata f. alba): Botanical, phytochemical and pharmacological overview. Phytochem. Rev. 2017, 16, 117–135. [Google Scholar] [CrossRef]
- Hrncirik, K.; Valusek, J.; Velisek, J. Investigation of ascorbigen as a breakdown product of glucobrassicin autolysis in Brassica vegetables. Eur. Food Res. Technol. 2001, 212, 576–581. [Google Scholar] [CrossRef]
- Kim, G.; Fujita, T.; Stankovic, K.M.; Welling, D.B.; Moon, I.S.; Choi, J.Y.; Yun, J.; Kang, J.S.; Lee, J.D. Sulforaphane, a natural component of broccoli, inhibits vestibular schwannoma growth in vitro and in vivo. Sci. Rep. 2016, 6, 36215. [Google Scholar] [CrossRef] [PubMed]
- Boote, K.J.; Jones, J.W.; Mishoe, J.W.; Berger, R.D. Coupling pests to crop growth simulators to predict yield reductions. Phytopathology 1983, 73, 1581–1587. [Google Scholar] [CrossRef]
- Lundrgardh, B.; Botek, P.; Schulzova, V.; Hajslova, J.; Stromberg, A.; Andersson, C. Impact of different green manures on the content of S-alk(en)yl-l-cysteine sulfoxides and L-ascorbic acid in leek (Allium porrum). J. Agric. Food Chem. 2008, 56, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, J.O. Spore production and dispersal of Alternaria dauci. Phytopathology 1977, 67, 1262–1266. [Google Scholar] [CrossRef]
- Shahin, E.A.; Shepard, J.F. An efficient technique for inducing profuse sporulation of Alternaria species. Phytopathology 1979, 69, 618–620. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available. |



{kind=link}
{kind=link}
{kind=link}
| Cultivar | Production System | Disease Severity 1 | Dry Biomass (%) | AA ± SD 2 (mg kg−1) | Total GSLs ± SD 2 (mg kg−1) |
|---|---|---|---|---|---|
| Albatros | Integrated | 3.04a * | 10.8 | 4326 ± 216 | 911 ± 36 |
| 2.71a ** | 9.7 | 1713 ± 86 | 982 ± 39 | ||
| Ecological | 3.63c * | 10.2 | 4275 ± 214 | 1470 ± 59 | |
| 3.38b ** | 10.5 | 3795 ± 190 | 1014 ± 41 | ||
| Target | Integrated | 3.29b * | 8.7 | 2933 ± 147 | 1090 ± 44 |
| 2.96a ** | 8.6 | 3271 ± 186 | 798 ± 32 | ||
| Ecological | 4.17e * | 9.2 | 2684 ± 134 | 1320 ± 53 | |
| 3.83c ** | 9.1 | 3353 ± 168 | 916 ± 37 |
| Glucosinolate | Abbreviation | Formula | Ion Type | m/z |
|---|---|---|---|---|
| Glucobrassicin | GB | C16H20N2O9S2 | [M − H]− | 447.0532 |
| 4-hydroxyglucobrassicin | HGB | C16H20N2O10S2 | [M − H]− | 463.0481 |
| Methoxyglucobrassicin/neoglucobrassicin | MGB/NGB | C17H22N2O10S2 | [M − H]− | 477.0638 |
| Sinigrin | SINI | C10H16KNO9S2 | [M − H]− | 358.0267 |
| Glucoiberin | IBER | C11H21NO10S3 | [M − H]− | 422.0249 |
| Progoitrin | PROG | C11H19NO10S2 | [M − H]− | 388.0372 |
| Glucoraphanin | RAPHA | C12H23NO10S3 | [M − H]− | 436.0406 |
| Gluconapin | NAPI | C11H19NO9S2 | [M − H]− | 372.0423 |
| Glucoibervirin | IBEV | C11H21NO9S3 | [M − H]− | 406.0300 |
| System | SINI | GB | MGB/NGB | HGB | IBEV | NAPI | PROG | IBER | RAPHA | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| I* | 241 | 289 | 53 | 19 | 10 | 59 | 109 | 106 | 26 | 912 |
| I** | 188 | 464 | 78 | 21 | 7 | 32 | 63 | 108 | 22 | 982 |
| E* | 342 | 418 | 116 | 23 | 16 | 115 | 222 | 175 | 44 | 1470 |
| E** | 337 | 278 | 92 | 14 | 11 | 36 | 59 | 156 | 30 | 1014 |
| System | SINI | GB | MGB/NGB | HGB | IBEV | NAPI | PROG | IBER | RAPHA | Total |
|---|---|---|---|---|---|---|---|---|---|---|
| I* | 294 | 507 | 98 | 13 | 8 | 20 | 34 | 107 | 9 | 1090 |
| I** | 267 | 260 | 73 | 15 | 8 | 15 | 26 | 119 | 14 | 798 |
| E* | 436 | 390 | 99 | 20 | 13 | 42 | 88 | 195 | 37 | 1320 |
| E** | 208 | 348 | 84 | 18 | 5 | 55 | 82 | 91 | 25 | 916 |
| Degradation Products | Summary Formula | Ion Type | m/z |
|---|---|---|---|
| Sulforaphane | C6H11NOS2 | [M + H]+ | 178.0360 |
| Iberin | C5H9NOS2 | [M + H]+ | 164.0204 |
| Indole-3-carbinol | C9H9NO | [M + H − H2O]+ | 130.0657 |
| Ascorbigen | C15H15NO6 | [M + H]+ | 306.0978 |
| Ionization Mode | Positive | Negative |
|---|---|---|
| Extraction solvent | EtAc 1 | MeOH 2 |
| Ionization gas temperature (He) | 450 °C | 350 °C |
| Ionization gas pressure (He) | 5.5 bar | 5.5 bar |
| Desorption time | 7 s | 7 s |
| Capillary voltage | 40 V | −50 V |
| Voltage of ion optical system | 250 V | −150 V |
| Skimmer voltage | 20 V | −20 V |
| Capillary temperature | 250 °C | 250 °C |
| Resolution | 50,000 FWHM | 50,000 FWHM |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novotny, C.; Schulzova, V.; Krmela, A.; Hajslova, J.; Svobodova, K.; Koudela, M. Ascorbic Acid and Glucosinolate Levels in New Czech Cabbage Cultivars: Effect of Production System and Fungal Infection. Molecules 2018, 23, 1855. https://doi.org/10.3390/molecules23081855
Novotny C, Schulzova V, Krmela A, Hajslova J, Svobodova K, Koudela M. Ascorbic Acid and Glucosinolate Levels in New Czech Cabbage Cultivars: Effect of Production System and Fungal Infection. Molecules. 2018; 23(8):1855. https://doi.org/10.3390/molecules23081855
Chicago/Turabian StyleNovotny, Cenek, Vera Schulzova, Ales Krmela, Jana Hajslova, Katerina Svobodova, and Martin Koudela. 2018. "Ascorbic Acid and Glucosinolate Levels in New Czech Cabbage Cultivars: Effect of Production System and Fungal Infection" Molecules 23, no. 8: 1855. https://doi.org/10.3390/molecules23081855
APA StyleNovotny, C., Schulzova, V., Krmela, A., Hajslova, J., Svobodova, K., & Koudela, M. (2018). Ascorbic Acid and Glucosinolate Levels in New Czech Cabbage Cultivars: Effect of Production System and Fungal Infection. Molecules, 23(8), 1855. https://doi.org/10.3390/molecules23081855