
Metabolomics of Solanum lycopersicum Infected with Phytophthora infestans Leads to Early Detection of Late Blight in Asymptomatic Plants

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
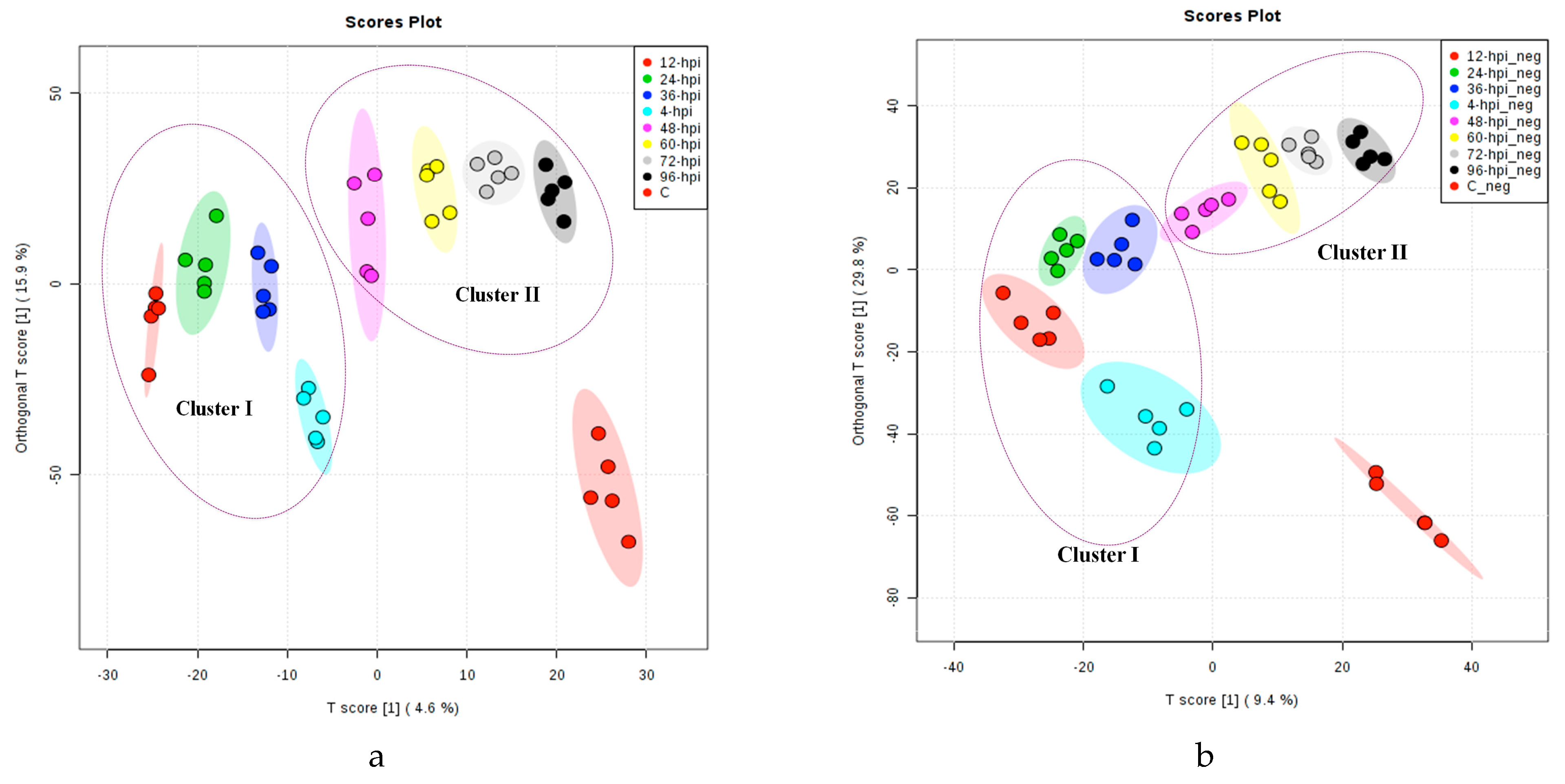
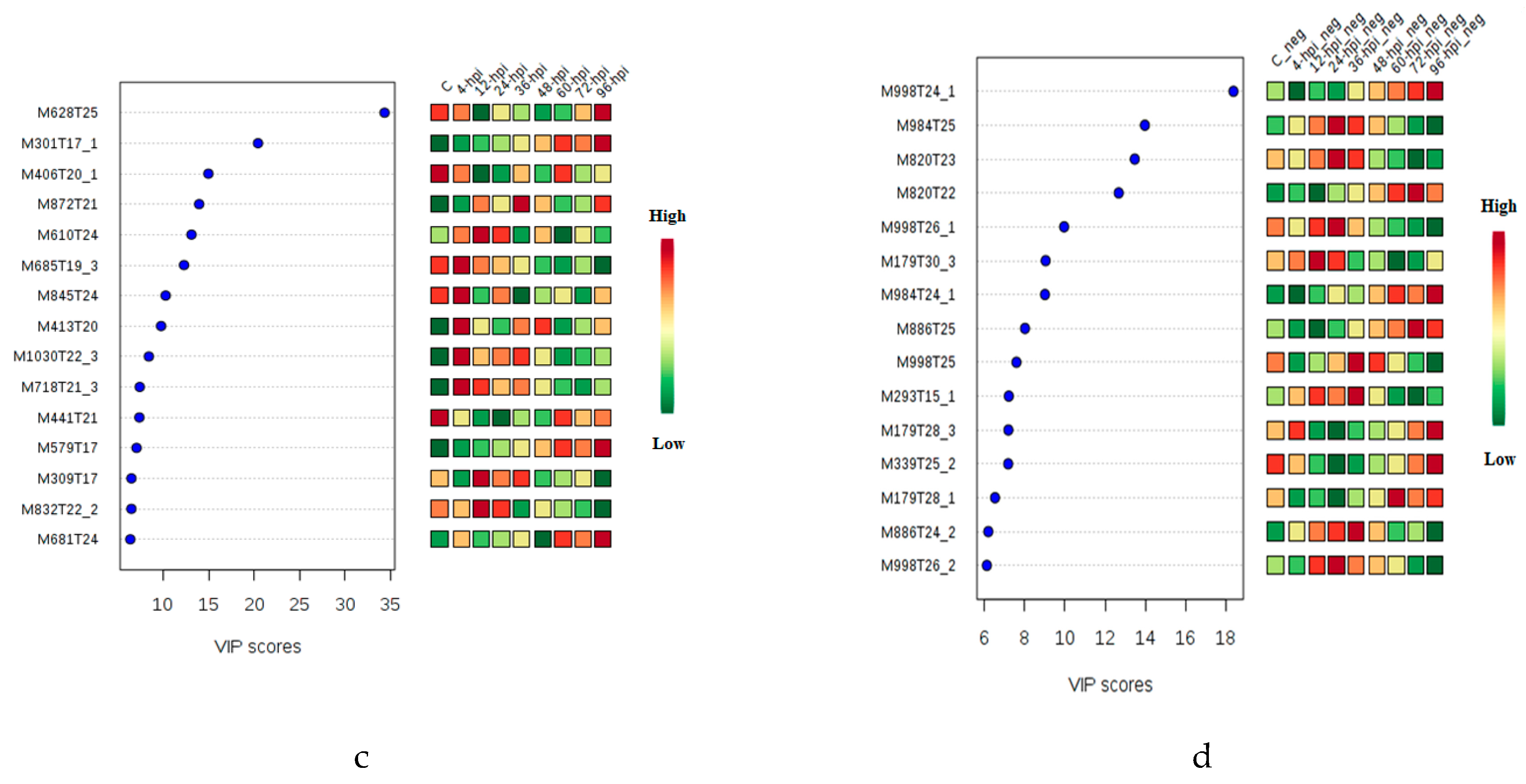
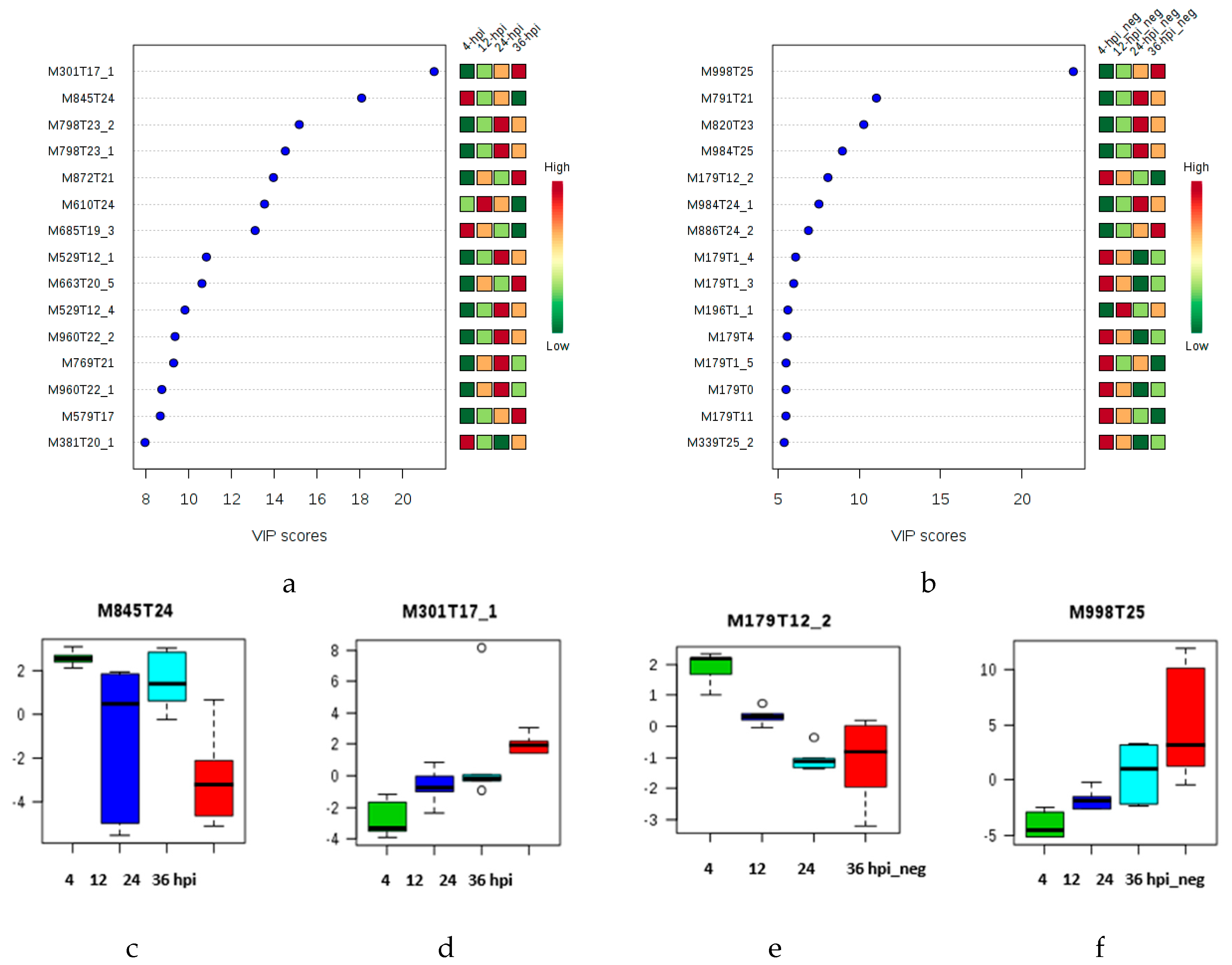
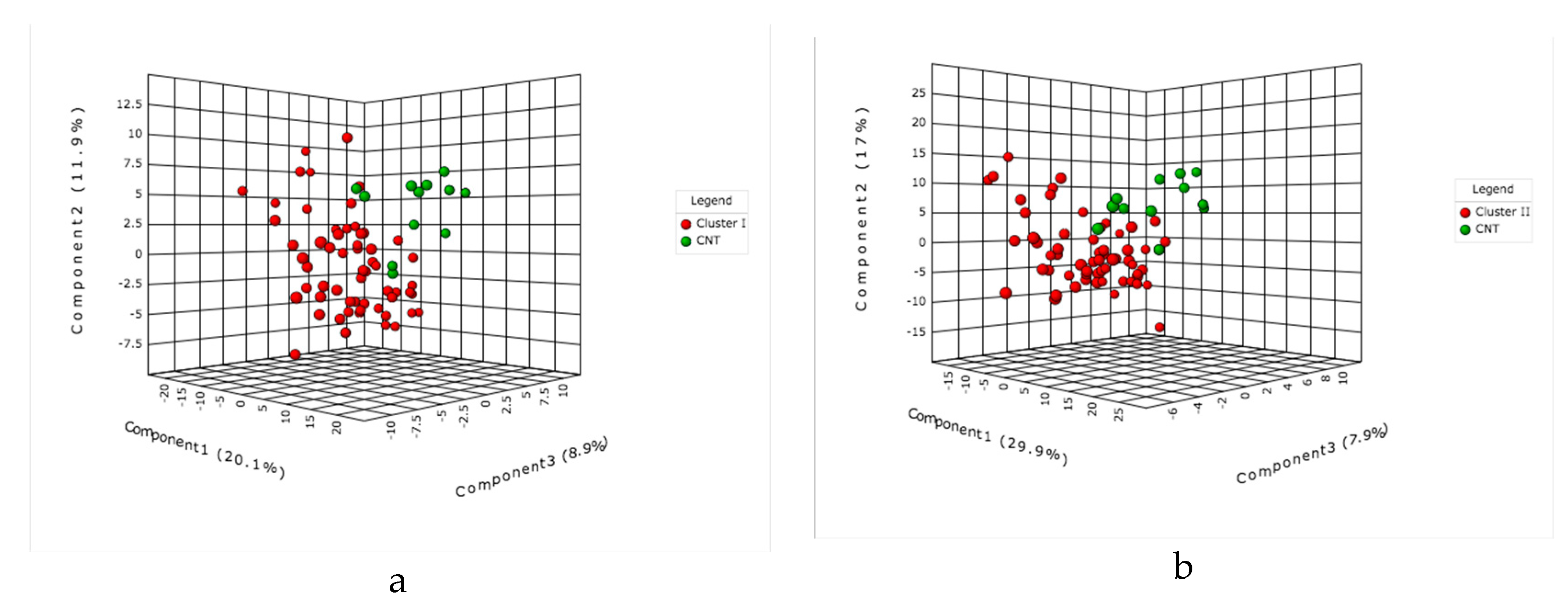
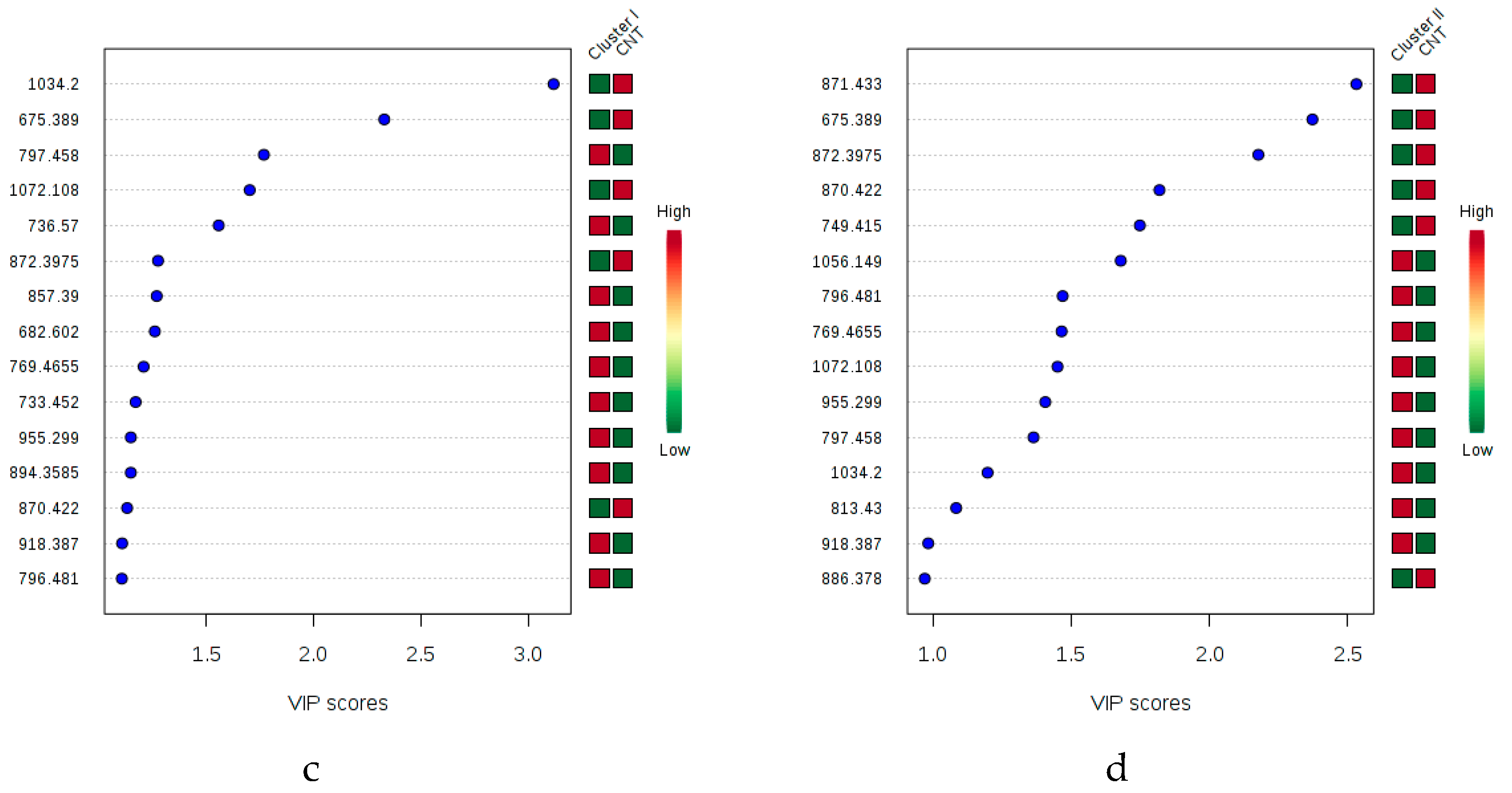
2.1. Multivariate Statistical Analysis of LC-MS Metabolomics Data
2.2. MALDI-MS Protocol for Analysis of Infected Tomato
2.3. Multivariate Data Analysis of MALDI-MS
3. Materials and Methods
3.1. Chemicals
3.2. Tomato Plant Samples
3.3. Pathogen Strain
3.4. Infection of Tomato Plants with Phytophthora Infestans
3.5. Sample Preparation
3.6. Untargeted Analysis of Metabolites
3.6.1. UHPLC-Q-TOF-MS Analysis
3.6.2. MALDI-MS Profile Analysis
3.7. Multivariate Data Analysis
3.7.1. LC-MS-Based Metabolomics Data
3.7.2. MALDI-MS Profile Data
3.8. Annotation of Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ballester, A.-R.; Tikunov, Y.; Molthoff, J.; Grandillo, S.; Viquez-Zamora, M.; de Vos, R.; de Maagd, R.A.; van Heusden, S.; Bovy, A.G. Identification of loci affecting accumulation of secondary metabolites in tomato fruit of a solanum lycopersicum × solanum chmielewskii introgression line population. Front. Plant Sci. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- de Vos, R.C.H.; Hall, R.D.; Moing, A. Metabolomics of a model fruit: Tomato. In Annual Plant Reviews; Wiley-Blackwell: Oxford, UK, 2011; Volume 43, pp. 109–155. ISBN 9781444339956. [Google Scholar]
- Cichon, M.J.; Riedl, K.M.; Schwartz, S.J. A metabolomic evaluation of the phytochemical composition of tomato juices being used in human clinical trials. Food Chem. 2017, 228, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Bovy, A.; Schijlen, E.; Hall, R.D. Metabolic engineering of flavonoids in tomato (Solanum lycopersicum): The potential for metabolomics. Metabolomics 2007, 3, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Fry, W. Phytophthora infestans: The plant (and R gene) destroyer. Mol. Plant Pathol. 2008, 9, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.M.; Ocampo, L.M.Q.; Céspedes, M.C.; Carreño, N.; González, A.; Rojas, A.; Zuluaga, A.P.; Myers, K.; Fry, W.E.; Jiménez, P.; et al. Characterization of Phytophthora infestans populations in Colombia: First report of the A2 mating type. Phytopathology 2009, 99, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Laurindo, B.S.; Laurindo, R.D.F.; Fontes, P.P.; Vital, C.E.; Delazari, F.T.; Baracat-Pereira, M.C.; da Silva, D.J.H. Comparative analysis of constitutive proteome between resistant and susceptible tomato genotypes regarding to late blight. Funct. Integr. Genomics 2018, 18, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Nick, C.; Aguilera, J.G.; Laurindo, B.S.; de Souza Almeida, V.; de Freitas, R.D.; Cruz, C.D.; da Silva, D.J.H. Genetic parameters and correlation between tomato late blight field resistance and fruit quality. Am. J. Plant Sci. 2014, 5, 2418–2425. [Google Scholar] [CrossRef]
- Abreu, F.B.; da Silva, D.J.H.; Cruz, C.D.; Mizubuti, E.S.G. Inheritance of resistance to Phytophthora infestans (Peronosporales, Pythiaceae) in a new source of resistance in tomato (Solanum sp. (formerly Lycopersicon sp.), Solanales, Solanaceae). Genet. Mol. Biol. 2008, 31, 493–497. [Google Scholar] [CrossRef]
- Elsayed, A.Y.; da Silva, D.J.H.; Carneiro, P.C.S.; Mizubuti, E.S.G. The inheritance of late blight resistance derived from Solanum habrochaites. Crop Breed. Appl. Biotechnol. 2012, 12, 199–205. [Google Scholar] [CrossRef]
- Pushpa, D.; Yogendra, K.N.; Gunnaiah, R.; Kushalappa, A.C.; Murphy, A. Identification of late blight resistance-related metabolites and genes in potato through nontargeted metabolomics. Plant Mol. Biol. Rep. 2014, 32, 584–595. [Google Scholar] [CrossRef]
- Lee, S.-J.; Rose, J.K.C. Mediation of the transition from biotrophy to necrotrophy in hemibiotrophic plant pathogens by secreted effector proteins. Plant Signal. Behav. 2010, 5, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Li, Y.; Yang, X.; Chen, J.; Hua, J.; Gao, Y. DNA barcoding identification of Pseudococcidae (Hemiptera: Coccoidea) using the mitochondrial COI gene. Mitochondrial DNA Part B 2018, 3, 419–423. [Google Scholar] [CrossRef]
- Ahmad, F.; Babalola, O.O.; Tak, H.I. Potential of MALDI-TOF mass spectrometry as a rapid detection technique in plant pathology: Identification of plant-associated microorganisms. Anal. Bioanal. Chem. 2012, 404, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Babalola, O.O.; Kirby, B.M.; Le Roes-Hill, M.; Cook, A.E.; Cary, S.C.; Burton, S.G.; Cowan, D.A. Phylogenetic analysis of actinobacterial populations associated with Antarctic Dry Valley mineral soils. Environ. Microbiol. 2009, 11, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Trout, C.L.; Ristaino, J.B.; Madritch, M.; Wangsomboondee, T. Rapid detection of Phytophthora infestans in late blight-infected potato and tomato using PCR. Plant Dis. 1997, 81, 1042–1048. [Google Scholar] [CrossRef]
- Tooley, P.W.; Bunyard, B.A.; Carras, M.M.; Hatziloukas, E. Development of PCR primers from internal transcribed spacer region 2 for detection of Phytophthora species infecting potatoes. Appl. Environ. Microbiol. 1997, 63, 1467–1475. [Google Scholar] [PubMed]
- Klein, A.T.; Yagnik, G.B.; Hohenstein, J.D.; Ji, Z.; Zi, J.; Reichert, M.D.; MacIntosh, G.C.; Yang, B.; Peters, R.J.; Vela, J.; et al. Investigation of the chemical interface in the soybean-aphid and rice-bacteria interactions using MALDI-mass spectrometry imaging. Anal. Chem. 2015, 87, 5294–5301. [Google Scholar] [CrossRef]
- Gupta, R.; Lee, S.E.; Agrawal, G.K.; Rakwal, R.; Park, S.; Wang, Y.; Kim, S.T. Understanding the plant-pathogen interactions in the context of proteomics-generated apoplastic proteins inventory. Front. Plant Sci. 2015, 6, 532. [Google Scholar] [CrossRef]
- Lodha, T.D.; Hembram, P.; Basak, N.; Tep, J. Proteomics: A successful approach to understand the molecular mechanism of plant-pathogen interaction. Am. J. Plant Sci. 2013, 4, 1212–1226. [Google Scholar] [CrossRef]
- Que, Y.; Xu, L.; Lin, J.; Ruan, M.; Zhang, M.; Chen, R. Differential protein expression in sugarcane during sugarcane- sporisorium scitamineum interaction revealed by 2-DE and MALDI-TOF-TOF/MS. Comp. Funct. Genom. 2011, 2011, 1–10. [Google Scholar] [CrossRef]
- Kim, J.Y.; Wu, J.; Kwon, S.J.; Oh, H.; Lee, S.E.; Kim, S.G.; Wang, Y.; Agrawal, G.K.; Rakwal, R.; Kang, K.Y.; et al. Proteomics of rice and Cochliobolus miyabeanus fungal interaction: Insight into proteins at intracellular and extracellular spaces. Proteomics 2014, 14, 2307–2318. [Google Scholar] [CrossRef] [PubMed]
- Abu-Nada, Y.; Kushalappa, A.C.; Marshall, W.D.; Al-Mughrabi, K.; Murphy, A. Temporal dynamics of pathogenesis-related metabolites and their plausible pathways of induction in potato leaves following inoculation with Phytophthora infestans. Eur. J. Plant Pathol. 2007, 118, 375–391. [Google Scholar] [CrossRef]
- Aliferis, K.A.; Faubert, D.; Jabaji, S. A metabolic profiling strategy for the dissection of plant defense against fungal pathogens. PLoS ONE 2014, 9, e111930. [Google Scholar] [CrossRef] [PubMed]
- Hijaz, F.M.; Manthey, J.A.; Folimonova, S.Y.; Davis, C.L.; Jones, S.E.; Reyes-De-Corcuera, J.I. An HPLC-MS characterization of the changes in sweet orange leaf metabolite profile following infection by the bacterial pathogen Candidatus Liberibacter asiaticus. PLoS ONE 2013, 8, e79485. [Google Scholar] [CrossRef] [PubMed]
- Jasiński, M.; Kachlicki, P.; Rodziewicz, P.; Figlerowicz, M.; Stobiecki, M. Changes in the profile of flavonoid accumulation in Medicago truncatula leaves during infection with fungal pathogen Phoma medicaginis. Plant Physiol. Biochem. 2009, 47, 847–853. [Google Scholar] [CrossRef]
- Wojakowska, A.; Muth, D.; Narożna, D.; Mądrzak, C.; Stobiecki, M.; Kachlicki, P. Changes of phenolic secondary metabolite profiles in the reaction of narrow leaf lupin (Lupinus angustifolius) plants to infections with Colletotrichum lupini fungus or treatment with its toxin. Metabolomics 2013, 9, 575–589. [Google Scholar] [CrossRef]
- Augustyn, W.A.; Regnier, T.; Combrinck, S.; Botha, B.M. Metabolic profiling of mango cultivars to identify biomarkers for resistance against Fusarium infection. Phytochem. Lett. 2014, 10, civ–cx. [Google Scholar] [CrossRef]
- Cajka, T.; Vaclavikova, M.; Dzuman, Z.; Vaclavik, L.; Ovesna, J.; Hajslova, J. Rapid LC-MS-based metabolomics method to study the Fusarium infection of barley. J. Sep. Sci. 2014, 37, 912–919. [Google Scholar] [CrossRef]
- Rivero, J.; Gamir, J.; Aroca, R.; Pozo, M.J.; Flors, V. Metabolic transition in mycorrhizal tomato roots. Front. Microbiol. 2015, 6, 598. [Google Scholar] [CrossRef]
- Wolfender, J.-L.; Marti, G.; Thomas, A.; Bertrand, S. Current approaches and challenges for the metabolite profiling of complex natural extracts. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef]
- Wolfender, J.; Rudaz, S.; Choi, Y.H.; Kim, H.K. Plant metabolomics: From holistic data to relevant biomarkers. Curr. Med. Chem. 2013, 20, 1056–1090. [Google Scholar] [CrossRef] [PubMed]
- López-Gresa, M.P.; Lisón, P.; Kim, H.K.; Choi, Y.H.; Verpoorte, R.; Rodrigo, I.; Conejero, V.; Bellés, J.M. Metabolic fingerprinting of Tomato Mosaic Virus infected Solanum lycopersicum. J. Plant Physiol. 2012, 169, 1586–1596. [Google Scholar] [CrossRef] [PubMed]
- Sade, D.; Shriki, O.; Cuadros-Inostroza, A.; Tohge, T.; Semel, Y.; Haviv, Y.; Willmitzer, L.; Fernie, A.R.; Czosnek, H.; Brotman, Y. Comparative metabolomics and transcriptomics of plant response to Tomato yellow leaf curl virus infection in resistant and susceptible tomato cultivars. Metabolomics 2015, 11, 81–97. [Google Scholar] [CrossRef]
- Bagherian, S.A.A.; Hamzehzarghani, H.; Izadpanah, K.; Djavaheri, M. Effects of potato spindle tuber viroid infection on tomato metabolic profile. J. Plant Physiol. 2016, 201, 42–53. [Google Scholar] [CrossRef]
- Eloh, K.; Sasanelli, N.; Maxia, A.; Caboni, P. Untargeted metabolomics of tomato plants after root-knot nematode infestation. J. Agric. Food Chem. 2016, 64, 5963–5968. [Google Scholar] [CrossRef] [PubMed]
- Camañes, G.; Scalschi, L.; Vicedo, B.; González-Bosch, C.; García-Agustín, P. An untargeted global metabolomic analysis reveals the biochemical changes underlying basal resistance and priming in Solanum lycopersicum, and identifies 1-methyltryptophan as a metabolite involved in plant responses to Botrytis cinerea and Pseudomonas sy. Plant J. 2015, 84, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Errard, A.; Ulrichs, C.; Kühne, S.; Mewis, I.; Drungowski, M.; Schreiner, M.; Baldermann, S. Single versus multiple-pest infestation affects differently the biochemistry of tomato (Solanum lycopersicum ’Ailsa Craig’). J. Agric. Food Chem. 2015, 63, 10103–10111. [Google Scholar] [CrossRef]
- Kant, M.R.; Ament, K.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Differential timing of spider mite-induced direct and indirect defenses in tomato plants. Plant Physiol. 2004, 135, 483–495. [Google Scholar] [CrossRef]
- Gromski, P.S.; Muhamadali, H.; Ellis, D.I.; Xu, Y.; Correa, E.; Turner, M.L.; Goodacre, R. A tutorial review: Metabolomics and partial least squares-discriminant analysis–A marriage of convenience or a shotgun wedding. Anal. Chim. Acta 2015, 879, 10–23. [Google Scholar] [CrossRef]
- Ivanisevic, J.; Benton, H.P.; Rinehart, D.; Epstein, A.; Kurczy, M.E.; Boska, M.D.; Gendelman, H.E.; Siuzdak, G. An interactive cluster heat map to visualize and explore multidimensional metabolomic data. Metabolomics 2015, 11, 1029–1034. [Google Scholar] [CrossRef]
- Banerjee, P.; Ghosh, S.; Dutta, M.; Subramani, E.; Khalpada, J.; RoyChoudhury, S.; Chakravarty, B.; Chaudhury, K. Identification of key contributory factors responsible for vascular dysfunction in idiopathic recurrent spontaneous miscarriage. PLoS ONE 2013, 8, e80940. [Google Scholar] [CrossRef]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [PubMed]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef]
- Bouarab, K.; Melton, R.; Peart, J.; Baulcombe, D.; Osbourn, A. A saponin-detoxifying enzyme mediates suppression of plant defences. Nature 2002, 418, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Faizal, A.; Geelen, D. Saponins and their role in biological processes in plants. Phytochem. Rev. 2013, 12, 877–893. [Google Scholar] [CrossRef]
- Osbourn, A.E. Preformed antimicrobial compounds and plant defense against fungal attack. Plant Cell Online 1996, 8, 1821–1831. [Google Scholar] [CrossRef]
- Toljamo, A.; Blande, D.; Kärenlampi, S.; Kokko, H. Reprogramming of strawberry (Fragaria vesca) root transcriptome in response to Phytophthora cactorum. PLoS ONE 2016, 11, 1–21. [Google Scholar] [CrossRef]
- Friedman, M. Tomato glycoalkaloids: Role in the Plant and in the diet. J. Agric. Food Chem. 2002, 50, 5751–5780. [Google Scholar] [CrossRef]
- Ito, S.; Eto, T.; Tanaka, S.; Yamauchi, N.; Takahara, H.; Ikeda, T. Tomatidine and lycotetraose, hydrolysis products of α-tomatine by Fusarium oxysporum tomatinase, suppress induced defense responses in tomato cells. FEBS Lett. 2004, 571, 31–34. [Google Scholar] [CrossRef]
- Itkin, M.; Rogachev, I.; Alkan, N.; Rosenberg, T.; Malitsky, S.; Masini, L.; Meir, S.; Iijima, Y.; Aoki, K.; de Vos, R.; et al. Glycoalkaloid metabolim 1 is Required for steroidal alkaloid glycosylation and prevention of phytotoxicity in tomato. Plant Cell 2011, 23, 4507–4525. [Google Scholar] [CrossRef]
- Dahlin, P.; Müller, M.C.; Ekengren, S.; McKee, L.S.; Bulone, V. The impact of steroidal glycoalkaloids on the physiology of Phytophthora infestans, the causative agent of potato late blight. Mol. Plant-Microbe Interact. 2017, 30, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Al Sinani, S.S.S.; Eltayeb, E.A. The steroidal glycoalkaloids solamargine and solasonine in Solanum plants. South African J. Bot. 2017, 112, 253–269. [Google Scholar] [CrossRef]
- Carere, J.; Colgrave, M.L.; Stiller, J.; Liu, C.; Manners, J.M.; Kazan, K.; Gardiner, D.M. Enzyme-driven metabolomic screening: A proof-of-principle method for discovery of plant defence compounds targeted by pathogens. New Phytol. 2016, 212, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Ökmen, B.; Etalo, D.W.; Joosten, M.H.A.J.; Bouwmeester, H.J.; de Vos, R.C.H.; Collemare, J.; de Wit, P.J.G.M. Detoxification of α-tomatine by Cladosporium fulvum is required for full virulence on tomato. New Phytol. 2013, 198, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Martin-Hernandez, M.; Dufresne, M.; Hugouvieux, V.; Melton, R.; Osbourn, A. Effects of targeted replacement of the tomatinase gene on the interaction of septoria lycopersici with tomato plants. Mol. Plant-Microbe Interact. 2000, 13, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- Blades, A.T.; Ikonomou, M.G.; Kebarle, P. Mechanism of electrospray mass spectrometry. Electrospray as an electrolysis cell. Anal. Chem. 1991, 63, 2109–2114. [Google Scholar] [CrossRef]
- Jeandet, P.; Clément, C.; Courot, E.; Cordelier, S. Modulation of phytoalexin biosynthesis in engineered plants for disease resistance. Int. J. Mol. Sci. 2013, 14, 14136–14170. [Google Scholar] [CrossRef]
- Li, R.; Tee, C.-S.; Jiang, Y.-L.; Jiang, X.-Y.; Venkatesh, P.N.; Sarojam, R.; Ye, J. A terpenoid phytoalexin plays a role in basal defense of Nicotiana benthamiana against Potato virus X. Sci. Rep. 2015, 5, 1–6. [Google Scholar] [CrossRef]
- Jadhav, S.J.; Mazza, G.; Salunkhe, D.K. Terpenoid phytoalexins in potatoes: A review. Food Chem. 1991, 41, 195–217. [Google Scholar] [CrossRef]
- Coxon, D.T.; Curtis, R.F.; Price, K.R.; Howard, B. Phytuberin: A novel antifungal terpenoid from potato. Tetrahedron Lett. 1974, 27, 2363–2366. [Google Scholar] [CrossRef]
- Andreu, A.; Oliva, C.; Distel, S.; Daleo, G. Production of phytoalexins, glycoalkaloids and phenolics in leaves and tubers of potato cultivars with different degrees of field resistance after infection with Phytophthora infestans. Potato Res. 2001, 44, 1–9. [Google Scholar] [CrossRef]
- Burka, L.T. 1-(3′-furyl)-6,7-dihydroxy-4,8-dimethylnonan-1-one, a stress metabolite from sweet potatoes (Ipomoea batatas). Phytochemistry 1978, 17, 317–318. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.M.; Nadira, U.A.; Bibi, N.; Cao, F.B.; He, X.Y.; Zhang, G.P.; Wu, F.B. Secondary metabolism and antioxidants are involved in the tolerance to drought and salinity, separately and combined, in Tibetan wild barley. Environ. Exp. Bot. 2015, 111, 1–12. [Google Scholar] [CrossRef]
- Yogendra, K.N.; Pushpa, D.; Mosa, K.A.; Kushalappa, A.C.; Murphy, A.; Mosquera, T. Quantitative resistance in potato leaves to late blight associated with induced hydroxycinnamic acid amides. Funct. Integr. Genomics 2014, 14, 285–298. [Google Scholar] [CrossRef] [PubMed]
- López-Gresa, M.P.; Maltese, F.; Bellés, J.M.; Conejero, V.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Metabolic response of tomato leaves upon different plant-pathogen interactions. Phytochem. Anal. 2010, 21, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Tomita, S.; Ikeda, S.; Tsuda, S.; Someya, N.; Asano, K.; Kikuchi, J.; Chikayama, E.; Ono, H.; Sekiyama, Y. A survey of metabolic changes in potato leaves by NMR-based metabolic profiling in relation to resistance to late blight disease under field conditions. Magn. Reson. Chem. 2017, 55, 120–127. [Google Scholar] [CrossRef]
- Cui, J.; Luan, Y.; Jiang, N.; Bao, H.; Meng, J. Comparative transcriptome analysis between resistant and susceptible tomato allows the identification of lncRNA16397 conferring resistance to Phytophthora infestans by co-expressing glutaredoxin. Plant J. 2017, 89, 577–589. [Google Scholar] [CrossRef]
- Li, J.; Luan, Y.; Liu, Z. SpWRKY1 mediates resistance to Phytophthora infestans and tolerance to salt and drought stress by modulating reactive oxygen species homeostasis and expression of defense-related genes in tomato. Plant Cell Tissue Organ Cult. 2015, 123, 67–81. [Google Scholar] [CrossRef]
- Cui, J.; Xu, P.; Meng, J.; Li, J.; Jiang, N.; Luan, Y. Transcriptome signatures of tomato leaf induced by Phytophthora infestans and functional identification of transcription factor SpWRKY3. Theor. Appl. Genet. 2018, 131, 787–800. [Google Scholar] [CrossRef]
- Sumner, L.W.; Mendes, P.; Dixon, R.A. Plant metabolomics: Large-scale phytochemistry in the functional genomics era. Phytochemistry 2003, 62, 817–836. [Google Scholar] [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- McCombie, G.; Knochenmuss, R. Small-molecule MALDI using the matrix suppression effect to reduce or eliminate matrix background interferences. Anal. Chem. 2004, 76, 4990–4997. [Google Scholar] [CrossRef] [PubMed]
- Popkova, Y.; Schiller, J. Addition of CsCl reduces ion suppression effects in the matrix-assisted laser desorption/ionization mass spectra of triacylglycerol/phosphatidylcholine mixtures and adipose tissue extracts. Rapid Commun. Mass Spectrom. 2017, 31, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Süß, R.; Schiller, J. An update of MALDI-TOF mass spectrometry in lipid research. Prog. Lipid Res. 2010, 49, 450–475. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, D.R.; Wang, Q.; Friedt, W.; Spengler, B.; Gottwald, S.; Römpp, A. High resolution mass spectrometry imaging of plant tissues: Towards a plant metabolite atlas. Analyst 2015, 140, 7696–7709. [Google Scholar] [CrossRef]
- Swarbick, P.J.; Schulze-Lefert, P.; Scholes, J.D. Metabolic consequences of susceptibility and resistance (race-specific and broad-spectrum) in barley leaves challenged with powdery mildew. Plant, Cell Environ. 2006, 29, 1061–1076. [Google Scholar] [CrossRef]
- Scholes, J.; Rolfe, S. Photosynthesis in localised regions of oat leaves infected with crown rust (Puccinia coronata): Quantitative imaging of chlorophyll fluorescence. Planta 1996, 199, 573–582. [Google Scholar] [CrossRef]
- BILGIN, D.D.; ZAVALA, J.A.; ZHU, J.; CLOUGH, S.J.; ORT, D.R.; DeLUCIA, E.H. Biotic stress globally downregulates photosynthesis genes. Plant. Cell Environ. 2010, 33, 1597–1613. [Google Scholar] [CrossRef]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front. Plant Sci. 2014, 5, 17. [Google Scholar] [CrossRef] [PubMed]
- Amarquaye, A.; Che, C.; Bejar, E.; Malone, M.; Fong, H. A new glycolipid from Byrsonima crassifolia. Planta Med. 1994, 60, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Endo, K.; Kobayashi, K.; Wada, H. Sulfoquinovosyldiacylglycerol has an essential role in thermosynechococcus elongatus BP-1 under phosphate-deficient conditions. Plant Cell Physiol. 2016, 57, 2461–2471. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Saito, K. Roles of lipids as signaling molecules and mitigators during stress response in plants. Plant J. 2014, 79, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Boudière, L.; Michaud, M.; Petroutsos, D.; Rébeillé, F.; Falconet, D.; Bastien, O.; Roy, S.; Finazzi, G.; Rolland, N.; Jouhet, J.; et al. Glycerolipids in photosynthesis: Composition, synthesis and trafficking. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Kim, Y.; Wi, S.J.; Seo, J.B.; Kwon, J.; Chung, J.H.; Park, K.Y.; Nam, M.H. Nontargeted Metabolite profiling in compatible pathogen-inoculated tobacco (Nicotiana tabacum L. cv. Wisconsin 38) using UPLC-Q-TOF/MS. J. Agric. Food Chem. 2012, 60, 11015–11028. [Google Scholar] [CrossRef]
- Furey, A.; Moriarty, M.; Bane, V.; Kinsella, B.; Lehane, M. Ion suppression; A critical review on causes, evaluation, prevention and applications. Talanta 2013, 115, 104–122. [Google Scholar] [CrossRef]
- Dong, Y.; Li, B.; Malitsky, S.; Rogachev, I.; Aharoni, A.; Kaftan, F.; Svatoš, A.; Franceschi, P. Sample preparation for mass spectrometry imaging of plant tissues: A Review. Front. Plant Sci. 2016, 7, 60. [Google Scholar] [CrossRef]
- Thunig, J.; Hansen, S.H.; Janfelt, C. Analysis of secondary plant metabolites by indirect desorption electrospray ionization imaging mass spectrometry. Anal. Chem. 2011, 83, 3256–3259. [Google Scholar] [CrossRef]
- Scanu, B.; Linaldeddu, B.T.; Deidda, A.; Jung, T. Diversity of phytophthora species from declining mediterranean maquis vegetation, including two new species, Phytophthora crassamura and P. ornamentata sp. nov. PLoS ONE 2015, 10, e0143234. [Google Scholar] [CrossRef]
- dos Santos, F.N.; Tata, A.; Belaz, K.R.A.; Magalhães, D.M.A.; Luz, E.D.M.N.; Eberlin, M.N. Major phytopathogens and strains from cocoa (Theobroma cacao L.) are differentiated by MALDI-MS lipid and/or peptide/protein profiles. Anal. Bioanal. Chem. 2017, 409, 1765–1777. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.F.; Chevallier, O.P.; Kumar, P.; Türkoğlu, O.; Bahado-Singh, R.O. High resolution metabolomic analysis of ASD human brain uncovers novel biomarkers of disease. Metabolomics 2016, 12, 62. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for comprehensive metabolomics data analysis. Curr. Protoc. Bioinforma. 2016, 55, 14.10.1–14.10.91. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Mandal, R.; Sinelnikov, I.V.; Broadhurst, D.; Wishart, D.S. MetaboAnalyst 2.0- a comprehensive server for metabolomic data analysis. Nucleic Acids Res. 2012, 40, W127–W133. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, G.B.; Jayasinghe, N.S.; Natera, S.H.A.; Inutan, E.D.; Peteros, N.P.; Roessner, U. From common to rare Zingiberaceae plants–A metabolomics study using GC-MS. Phytochemistry 2017, 140, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Musharraf, S.G.; Siddiqui, A.J.; Shamsi, T.; Choudhary, M.I.; Rahman, A. Serum metabonomics of acute leukemia using nuclear magnetic resonance spectroscopy. Sci. Rep. 2016, 6, 30693. [Google Scholar] [CrossRef]
- Zhang, L.-N.; Wang, L.; Shi, Z.-Q.; Li, P.; Li, H.-J. A metabolomic strategy based on integrating headspace gas chromatography-mass spectrometry and liquid chromatography-mass spectrometry to differentiate the five cultivars of Chrysanthemum flower. RSC Adv. 2018, 8, 9074–9082. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen rearrangement rules: computational MS/MS fragmentation and structure elucidation using MS-FINDER software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Moco, S.; Bino, R.J.; Vorst, O.; Verhoeven, H.A.; de Groot, J.; van Beek, T.A.; Vervoort, J.; de Vos, R. A liquid chromatography-mass spectrometry- based metabolome database for tomato. Plant Physiol. 2006, 141, 1205–1218. [Google Scholar] [CrossRef]
- Mintz-Oron, S.; Mandel, T.; Rogachev, I.; Feldberg, L.; Lotan, O.; Yativ, M.; Wang, Z.; Jetter, R.; Venger, I.; Adato, A.; et al. Gene expression and metabolism in tomato fruit surface tissues. Plant Physiol. 2008, 147, 823–851. [Google Scholar] [CrossRef] [PubMed]
- Iijima, Y.; Fujiwara, Y.; Tokita, T.; Ikeda, T.; Nohara, T.; Aoki, K.; Shibata, D. Involvement of ethylene in the accumulation of esculeoside a during fruit ripening of tomato (Solanum lycopersicum). J. Agric. Food Chem. 2009, 57, 3247–3252. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecular Feature | RT (min) | Metabolite | Molecular Formula | Annotation Level | Theor. (m/z) | Found (m/z) | AME (ppm) | Adducts |
|---|---|---|---|---|---|---|---|---|
| M609T9 | 9.06 | Quercetin 7-(rhamnosylglucoside) | C27H30O16 | 2 | 609.14555 | 609.1464 | 1.4 | [M − H]− |
| M1095T10 | 9.85 | Hydroxytomatine isomer I | C50H83NO22 | 3 | 1094.53832 | 1094.5382 | 0.1 | [M + FA − H]− |
| M1077T12 | 11.52 | Dehydrotomatine | C50H81NO21 | 2 | 1076.52776 | 1076.5277 | 0.1 | [M + FA − H]− |
| M1109T12 | 11.58 | UGA11 | C52H87NO24 | 3 | 1108.55397 | 1108.5541 | 0.1 | [M − H]− |
| M1121T12_1 | 11.72 | UGA28 | C52H83NO25 | 3 | 1120.51759 | 1120.5120 | 5.0 | [M − H]− |
| M1079T12 | 11.94 | Tomatine | C50H83NO21 | 2 | 1078.5440 | 1078.5436 | 0.4 | [M + FA − H]− |
| M433T13_2 | 12.86 | Naringenin-hexose I | C21H22O10 | 3 | 433.11347 | 433.1115 | 4.5 | [M − H]− |
| M317T13_2 | 13.09 | Catechin 7,4′-dimethyl ether | C17H18O6 | 2 | 317.10251 | 317.1020 | 1.6 | [M − H]− |
| M293T15_1 | 14.51 | Phytuberin | C17H26O4 | 2 | 293.17528 | 293.1774 | 7.2 | [M − H]− |
| M221T16_2 | 16.30 | Rishitin | C14H22O2 | 2 | 221.15415 | 221.1561 | 8.8 | [M − H]− |
| M737T20 | 19.55 | Tuberoside J | C39H64O14 | 2 | 737.41121 | 737.4186 | 10.0 | [M − H]− |
| M414T20 | 19.59 | Tomatidine | C27H45NO2 | 2 | 414.3372 | 414.3385 | 3.1 | [M − H]− |
| M791T21 | 21.46 | Triterpenoid saponins- Sapimukoside J | C44H72O12 | 2 | 791.49455 | 791.4955 | 1.2 | [M − H]− |
| M832T22_2 | 22.21 | Peonidin 3-(4-sinapoylgentiobioside) | C39H43O20 | 2 | 832.24259 | 832.2387 | 4.7 | [M + H]+ |
| M794T22_1 | 22.24 | 1,2-Di-O-palmitoyl-3-O-(6-sulfoquinovopyranosyl)glycerol | C41H78O12S | 2 | 793.51357 | 793.5109 | 3.4 | [M − H]− |
| M798T23_2 | 22.58 | Triterpene saponins- UNPD101109 | C43H72O13 | 2 | 797.50511 | 797.5049 | 0.3 | [M + H]+ |
| M820T23 | 22.69 | Sulfoquinovosyldiacylglycerols | C43H80O12S | 3 | 819.5289 | 819.5269 | 2.4 | [M − H]− |
| M743T23_3 | 22.83 | 3-O-a-l-Arabinopyranosylproanthocyanidin A5 | C35H32O16 | 2 | 743.13788 | 743.1395 | 2.2 | [M + Cl]− |
| M610T24 | 24.07 | Apigenin 7-[rhamnosyl-(1->2)-galacturonide] | C27H28O15 | 2 | 610.17719 | 610.1758 | 2.3 | [M + NH4]+ |
| M845T24 | 24.40 | Triterpene saponins- Cyclopassifloside III | C43H72O16 | 2 | 845.48986 | 845.4816 | 9.8 | [M + H]+ |
| M984T24_1 | 24.49 | Triterpene saponins | C48H84O18 | 3 | 983.53461 | 983.5393 | 4.8 | [M + Cl]− |
| M984T25 | 24.87 | Triterpene saponins | C48H84O18 | 3 | 983.53461 | 983.5392 | 4.7 | [M + Cl]− |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galeano Garcia, P.; Neves dos Santos, F.; Zanotta, S.; Eberlin, M.N.; Carazzone, C. Metabolomics of Solanum lycopersicum Infected with Phytophthora infestans Leads to Early Detection of Late Blight in Asymptomatic Plants. Molecules 2018, 23, 3330. https://doi.org/10.3390/molecules23123330
Galeano Garcia P, Neves dos Santos F, Zanotta S, Eberlin MN, Carazzone C. Metabolomics of Solanum lycopersicum Infected with Phytophthora infestans Leads to Early Detection of Late Blight in Asymptomatic Plants. Molecules. 2018; 23(12):3330. https://doi.org/10.3390/molecules23123330
Chicago/Turabian StyleGaleano Garcia, Paula, Fábio Neves dos Santos, Samantha Zanotta, Marcos Nogueira Eberlin, and Chiara Carazzone. 2018. "Metabolomics of Solanum lycopersicum Infected with Phytophthora infestans Leads to Early Detection of Late Blight in Asymptomatic Plants" Molecules 23, no. 12: 3330. https://doi.org/10.3390/molecules23123330
APA StyleGaleano Garcia, P., Neves dos Santos, F., Zanotta, S., Eberlin, M. N., & Carazzone, C. (2018). Metabolomics of Solanum lycopersicum Infected with Phytophthora infestans Leads to Early Detection of Late Blight in Asymptomatic Plants. Molecules, 23(12), 3330. https://doi.org/10.3390/molecules23123330