
Anti-Obesity Effects of Medicinal and Edible Mushrooms



Abstract
1. Introduction
2. Etiology of Obesity
2.1. Diet, Lifestyle, and Societal Determinants
2.2. Genetics
2.3. Medical and Psychiatric Illness
2.4. Infectious Agents
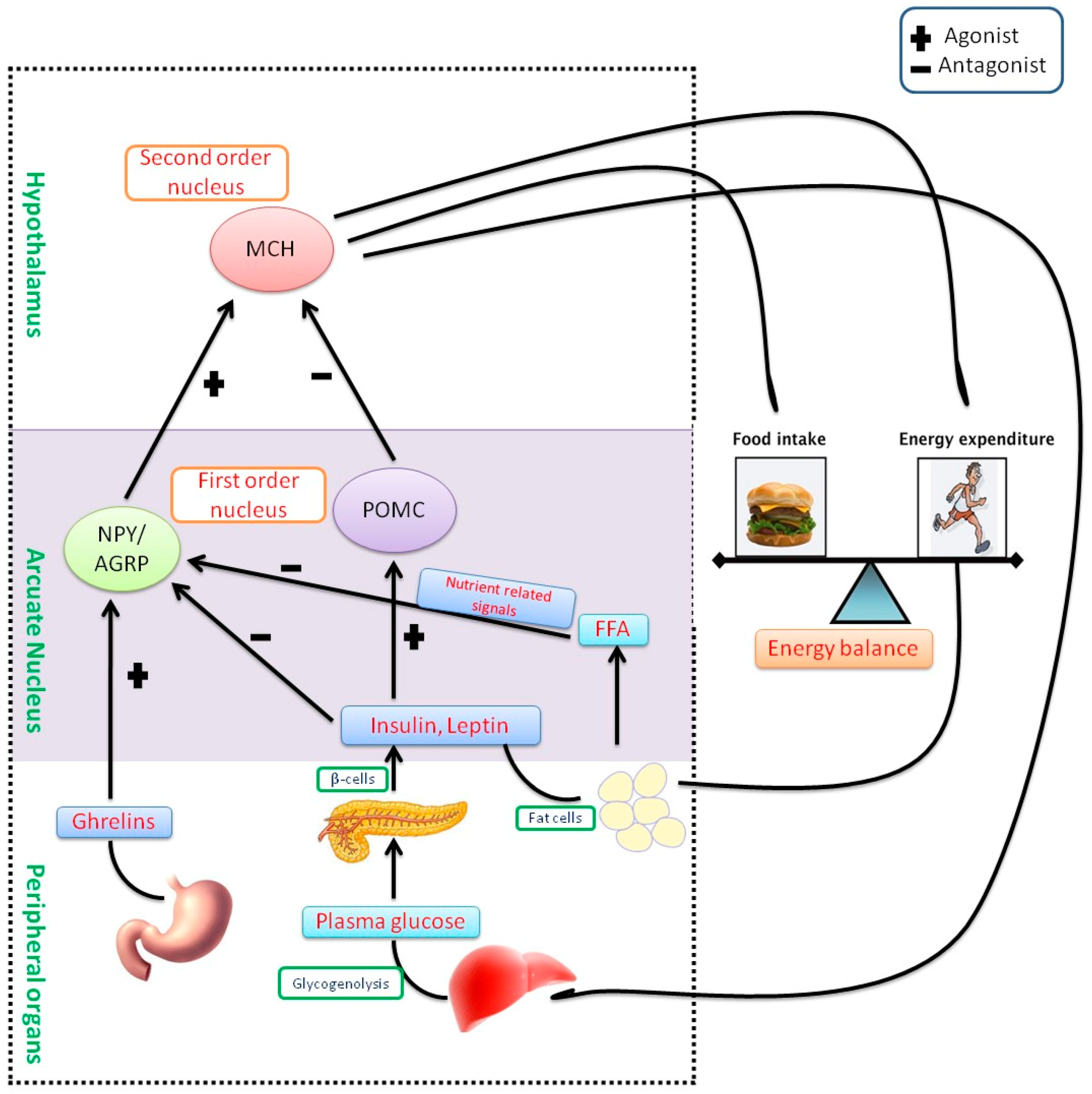
3. Pathophysiology of Obesity
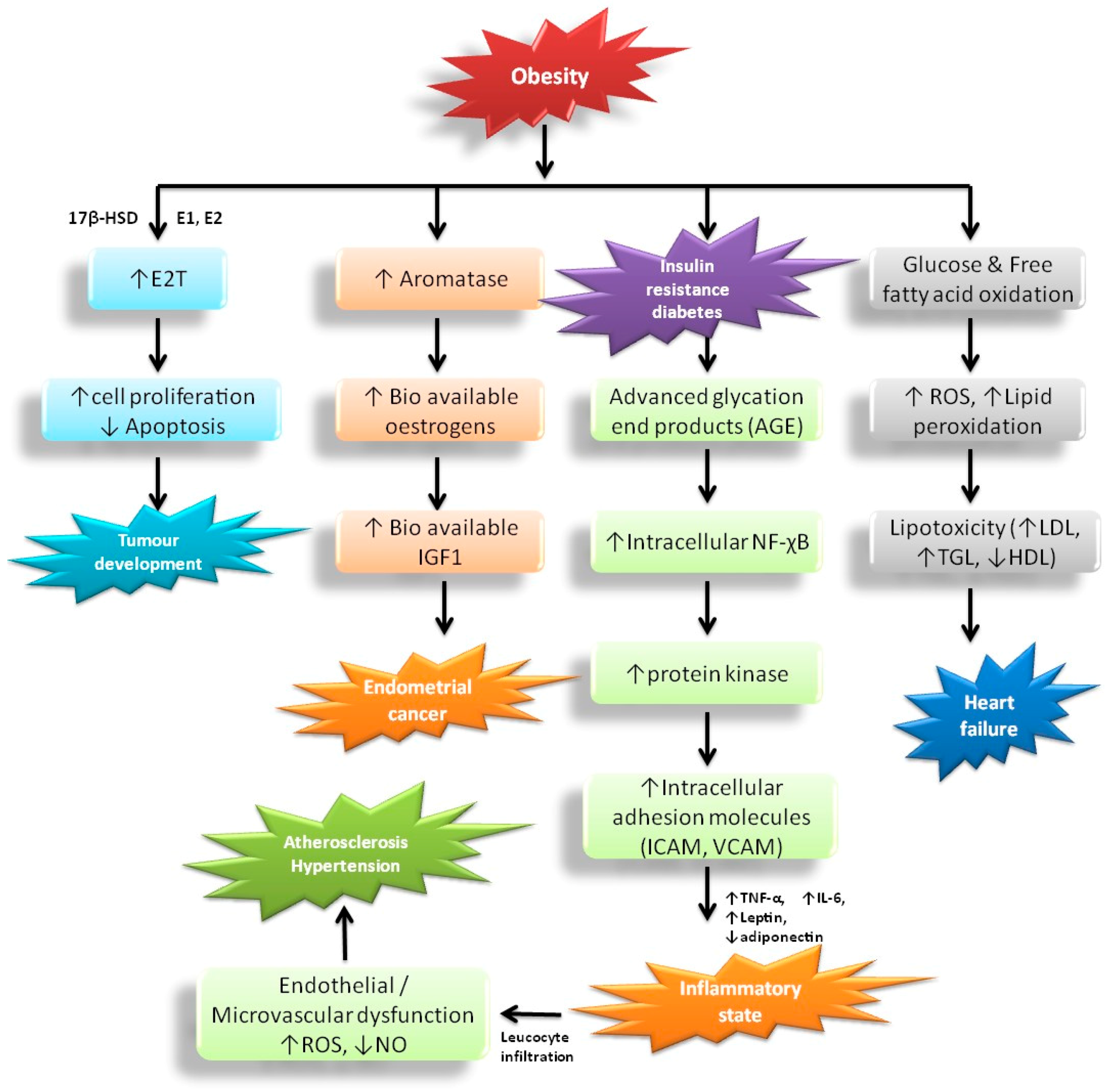
4. Pathologies Associated with Obesity and Its Effects on Health
4.1. Chronic Inflammation and Endothelial Dysfunction
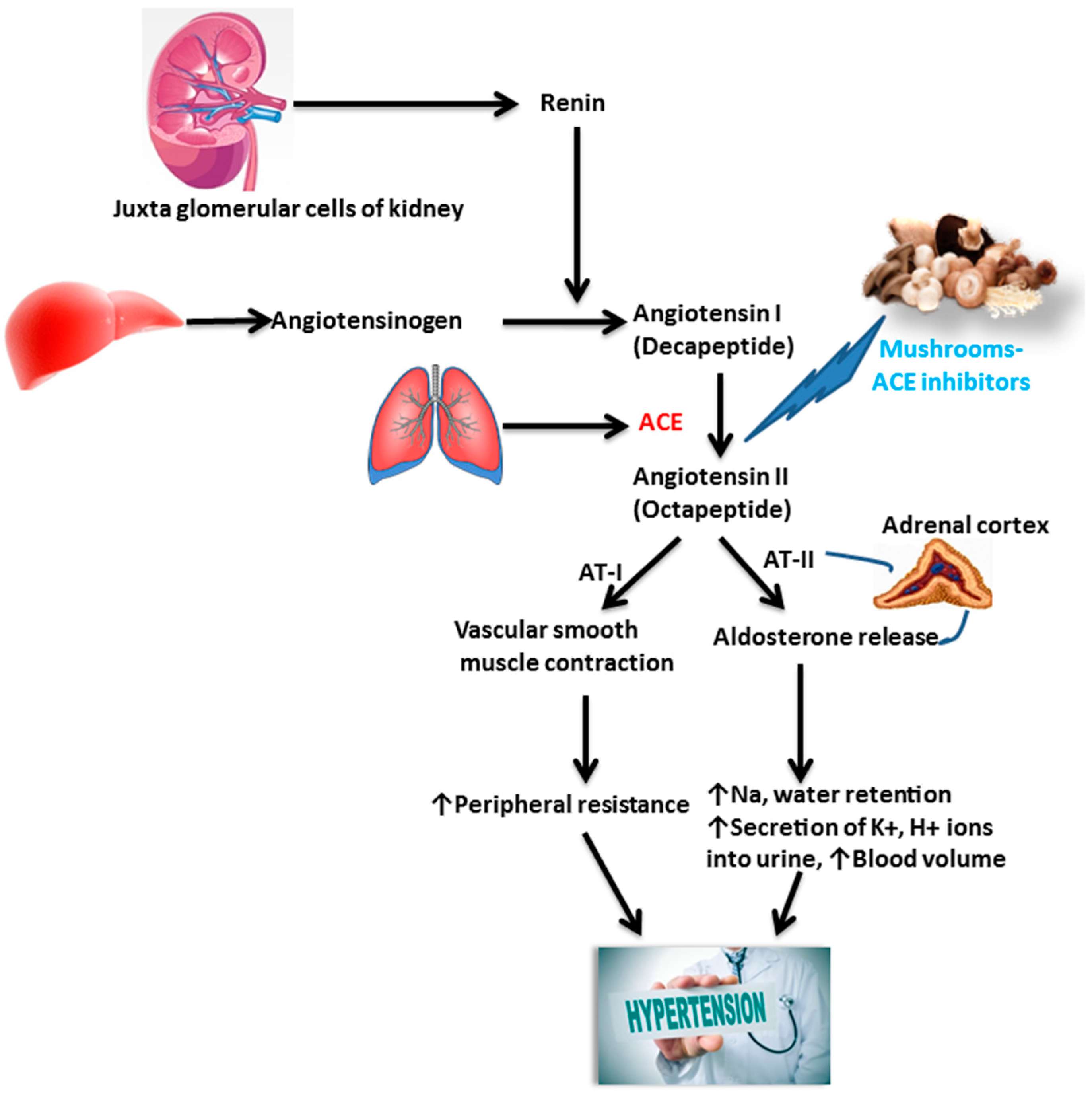
4.2. Hypertension and Atherosclerosis
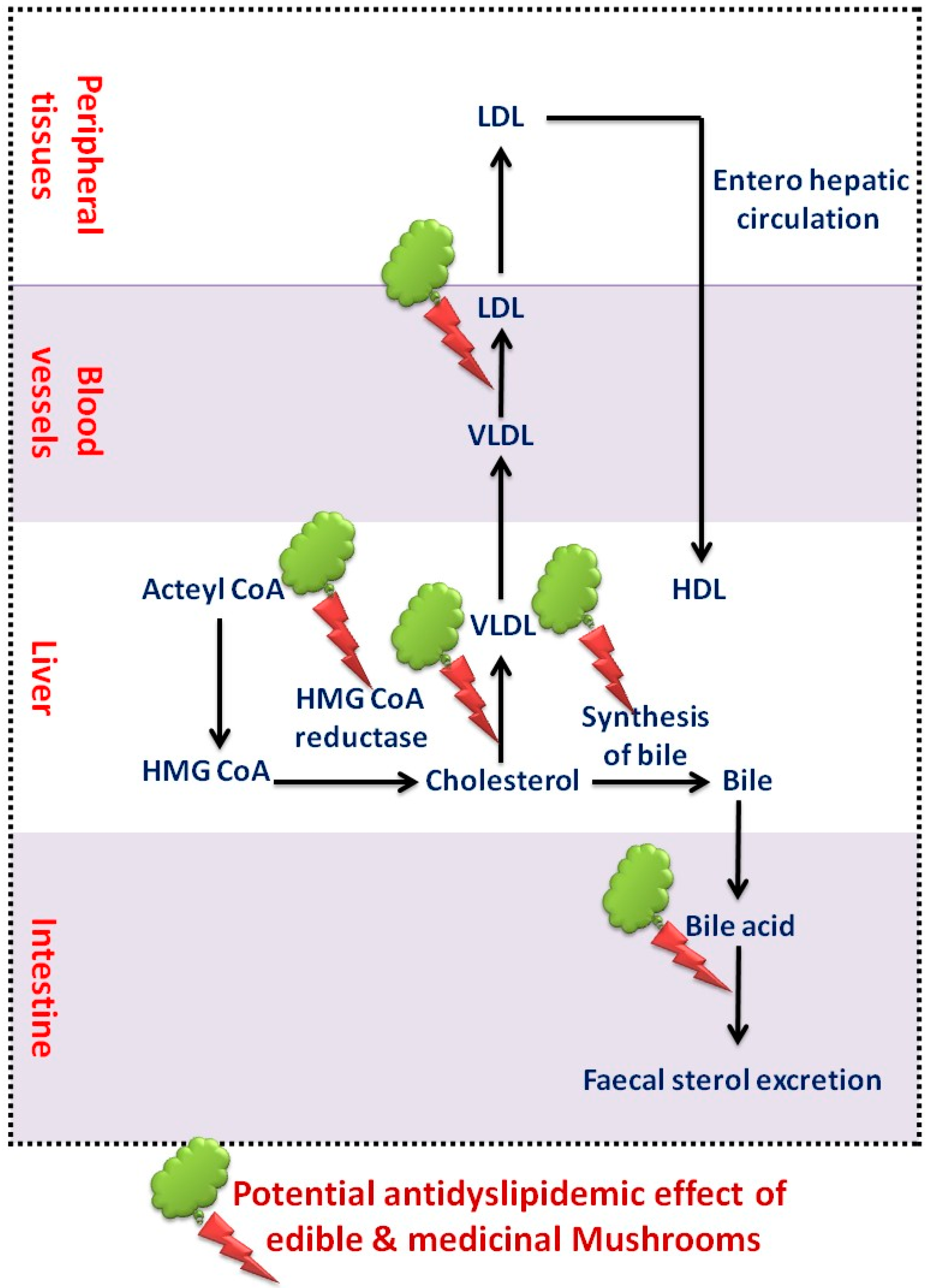
4.3. Dyslipidemia and Cardiac Alterations
4.4. Metabolic Syndrome
4.5. Cancer and Neurodegenerative Disorders
4.6. Sex Hormone Imbalance
5. Treatment of Obesity
5.1. Dietary Intervention and Diet Control
5.2. Physical Activity and Pharmacotherapy
5.3. Surgical Treatment
5.4. Natural Products
5.5. Mushrooms
6. Anti-Obesity Effects of Edible and Medicinal Mushrooms
6.1. Actions on Hypertension
6.2. Actions on Dyslipidemia
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 17β-HSD | 17β-hydroxysteroid dehydrogenase |
| ACC1 | acetyl-CoA carboxylase 1 |
| ACE | angiotensin converting enzyme |
| ACO | 1-aminocyclopropane-1-carboxylate oxidase |
| AGE | advanced glycation end products |
| AGPAT | 1-acylglycerol-3-phosphate O-acyltransferase 1 |
| AgRP | agouti-related peptide |
| ALT | alanine transaminase |
| AMPK | 5′ adenosine monophosphate-activated protein kinase |
| Ang-I | angiotensin I |
| Ang-II | angiotensin II |
| AST | aspartate transaminase |
| AT-1 | angiotensin II type 1 receptor |
| BMI | body mass index |
| BMR | basal metabolic rate |
| BUN | blood urea nitrogen |
| Ca | calcium |
| CAT | catalase |
| Cl | chloride |
| CNS | central nervous system |
| CPT1 | carnitine Palmitoyltransferase 1A |
| CRE | creatinine |
| CRP | c-reactive protein |
| CVD | cardiovascular diseases |
| DGAT | diacylglycerol O-Acyltransferase 1 |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| E1 | estrone |
| E2 | estradiol |
| FFA | free fatty acids |
| GPATR | glycerol-3-phosphate acyltransferase |
| GSH | glutathione |
| GSH-Px | glutathione peroxidase |
| HDL | high density lipoprotein |
| HFD | high fat diet |
| HMG-CoA | β-hydroxy, β methyl glutaryl COA |
| ICAM | Intercellular Adhesion Molecule |
| IGF1 | IGF binding protein 1 |
| IL-1 | interleukin 1 |
| IL-6 | interleukin 6 |
| K | potassium |
| LDL | low density lipoprotein |
| LPO | lipid peroxidation |
| MAPK | mitogen-activated protein kinases |
| MCH | melanin concentrating hormone |
| MDA | melondialdehyde |
| Mg | magnesium |
| mRNA | messenger ribonucleic acids |
| Na | sodium |
| NF-χB | nuclear factor kappa B |
| NO | nitric oxide |
| NPY | neuropeptide Y |
| OGTT | oral glucose tolerance test |
| P | phosphorus |
| POMC | pro-opiomelanocortin |
| PPAR-α | peroxisome proliferator-activated receptor alpha |
| PUFA | poly unsaturated fatty acids |
| RAS | renin-angiotensin-aldosterone system |
| ROS | reactive oxygen species |
| SCD1 | stearoyl-CoA desaturase 1 |
| SCFA | short chain fatty acids |
| SCFA | short chain fatty acids |
| SOD | superoxide dismutase |
| SREBP-1c | sterol regulatory element-binding transcription factor 1 |
| STZ | streptozotocin |
| T-AOC | total antioxidant capacity |
| TBARS | thiobarbituric acid reactive substances |
| TC | total cholesterol |
| TG | triglycerides |
| TNF-α | tumour necrosis factor-α |
| UCP2 | mitochondrial uncoupling proteins 2 |
| UCP3 | mitochondrial uncoupling proteins 3 |
| VCAM | vascular cell adhesion molecule |
| VLDL | very low density lipoprotein |
| WHO | world health organization |
| WLS | weight loss surgery |
| Δ4A | Δ4-androstenedione |
References
- Kanagasabapathy, G.; Malek, S.N.A.; Mahmood, A.A.; Chua, K.H.; Vikineswary, S.; Kuppusamy, U.R. Beta-glucan-rich extract from Pleurotus sajor-caju (Fr.) Singer prevents obesity and oxidative stress in C57BL/6J Mice fed on a high-fat diet. Evid. Based Complement. Altern. Med. 2013, 2013, 185259. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Tfayli, H.; Michaliszyn, S.F.; Arslanian, S. Impaired lipolysis diminished fat oxidation and metabolic inflexibility in obese girls with polycystic ovary syndrome. J. Clin. Endocrinol. MeTable. 2018, 103, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Bang, C.Y.; Guo, Y.R.; Choung, S.Y. Anti-obesity effects of Aster spathulifolius extract in high-fat-diet-induced obese rats. J. Med. Food 2016, 19, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Kennett, G.A.; Clifton, P.G. New approaches to the pharmacological treatment of obesity: Can they break through the efficacy barrier? Pharmacol. Biochem. Behav. 2010, 97, 63–83. [Google Scholar] [CrossRef] [PubMed]
- Bardou, M.; Barkun, A.N.; Martel, M. Obesity and colorectal cancer. Gut 2013, 62, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, W.M.T.; Panveloski-Costa, A.C.; Yokota, C.N.F.; Pereira, J.N.B.; Filho, J.M.; Torres, R.P.; Hirabara, S.M.; Curi, R.; Alba-Loureiro, T.C. Comparison of Goto-Kakizaki rats and high fat diet-induced obese rats: Are they reliable models to study Type 2 Diabetes mellitus? PLoS ONE 2017, 12, e0189622. [Google Scholar] [CrossRef] [PubMed]
- Gooda Sahib Jambocus, N.; Saari, N.; Ismail, A.; Khatib, A.; Mahomoodally, M.F.; Abdul Hamid, A. An investigation into the antiobesity effects of Morinda citrifolia L. leaf extract in high-fat diet-induced obese rats using a (1)H NMR metabolomics approach. J. Diabetes Res. 2016, 2016, 2391592. [Google Scholar] [PubMed]
- Yamada, T.; Oinuma, T.; Niihashi, M.; Mitsumata, M.; Fujioka, T.; Hasegawa, K.; Nagaoka, H.; Itakura, H. Effects of Lentinus edodes mycelia on dietary-induced atherosclerotic involvement in rabbit aorta. J. Atheroscler. Thromb. 2002, 9, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kasabri, V.; Al-Hallaq, E.K.; Bustanji, Y.K.; Abdul-Razzak, K.K.; Abaza, I.F.; Afifi, F.U. Antiobesity and antihyperglycaemic effects of Adiantum capillus-veneris extracts: In vitro and in vivo evaluations. Pharm. Biol. 2017, 55, 164–172. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity and Overweight. Fact Sheet. Updated in June 2016. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 28 June 2016).
- Rodríguez-Rodero, S.; Menéndez-Torre, E.; Fernández-Bayón, G.; Morales-Sánchez, P.; Sanz, L.; Turienzo, E.; González, J.J.; Martinez-Faedo, C.; Suarez-Gutiérrez, L.; Ares, J.; et al. Altered intragenic DNA methylation of HOOK2 gene in adipose tissue from individuals with obesity and type 2 diabetes. PLoS ONE 2017, 12, e0189153. [Google Scholar] [CrossRef] [PubMed]
- Flegal, K.M.; Carroll, M.D.; Ogden, C.L.; Curtin, L.R. Prevalence and trends in obesity among United States adults, 1999–2008. J. Am. Med. Assoc. 2010, 303, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Gooding, H.C.; Ning, H.; Gillman, M.W.; Shay, C.; Allen, N.; Goff, D.C., Jr.; Lloyd-Jones, D.; Chiuve, S. Application of a lifestyle-based tool to estimate premature cardiovascular disease events in young adults: The coronary artery risk development in young adults (CARDIA) Study. JAMA Int. Med. 2017, 177, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Elffers, T.W.; de Mutsert, R.; Lamb, H.J.; de Roos, A.; Willems van Dijk, K.; Rosendaal, F.R.; Jukema, J.W.; Trompet, S. Body fat distribution, in particular, visceral fat, is associated with cardiometabolic risk factors in obese women. PLoS ONE 2017, 12, e0185403. [Google Scholar] [CrossRef] [PubMed]
- Reales, G.; Rovaris, D.L.; Jacovas, V.C.; Hünemeier, T.; Sandoval, J.R.; Salazar-Granara, A.; Demarchi, D.A.; Tarazona-Santos, E.; Felkl, A.B.; Serafini, M.A.; et al. A tale of agriculturalists and hunter-gatherers: Exploring the thrifty genotype hypothesis in native South Americans. Am. J. Physiol. Anthropol. 2017, 163, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Provencher, M.D.; Bélanger, M.È.; Shriqui, C.; Lachance, I.; Bonneville, S. Psychoeducation for overweight patients with psychiatric disorders: The Wellness program developed in Quebec. Encephale 2016, 42, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Vandyk, A.D.; Baker, C. Qualitative descriptive study exploring schizophrenia and the everyday effect of medication-induced weight gain. Int. J. Ment. Health Nurs. 2012, 21, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Charlotte, M.; Schwartz, E.; Slade, E.; Medoff, D.; Li, L.; Dixon, L.; Kilbourne, A.M.; Kreyenbuhl, J. Gender differences in mood stabilizer medications prescribed to Veterans with serious mental illness. J. Affect. Disord. 2015, 188, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Nihalani, N.; Schwartz, T.L.; Siddiqui, U.A.; Megna, J.L. Weight gain, obesity, and psychotropic prescribing. J. Obes. 2011, 2011, 893629. [Google Scholar] [CrossRef] [PubMed]
- Wierucka-Rybak, M.; Bojanowska, E. Bacteria, viruses, and hypothalamic inflammation: Potential new players in obesity. Postepy Hig. Med. Dosw. 2014, 68, 271–279. [Google Scholar] [CrossRef]
- Karlsson, E.A.; Beck, M.A. The burden of obesity on infectious disease. Exp. Biol. Med. 2010, 235, 1412–1424. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, H.; Heppner, K.M.; Holland, J.; Kabra, D.; Tschöp, M.H.; Pfluger, P.T. Ablation of ghrelin O-acyltransferase does not improve glucose intolerance or body adiposity in mice on a leptin-deficient ob/ob background. PLoS ONE 2013, 8, e61822. [Google Scholar] [CrossRef] [PubMed]
- Palou, M.; Sanchez, J.; Rodriguez, A.M.; Priego, T.; Pico, C. Induction of NPY/AgRP orexigenic peptide expression in rat hypothalamus is an early event in fasting: Relationship with circulating leptin, insulin, and glucose. Cell Physiol. Biochem. 2009, 23, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, J.; Imai, J.; Izumi, T.; Takahashi, H.; Kawana, Y.; Takahashi, K.; Kodama, S.; Kaneko, K.; Gao, J.; Uno, K.; et al. Neuronal signals regulate obesity-induced β-cell proliferation by FoxM1 dependent mechanism. Nat. Commun. 2017, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
- Das Graças Coelho de Souza, M.; Kraemer-Aguiar, L.G.; Bouskela, E. Inflammation-induced microvascular dysfunction in obesity—A translational approach. Clin. Hemorheol. Microcirc. 2016, 64, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Sukalingam, K.; Xu, B. Impact of consumption and cooking manners of vegetable oils on cardiovascular diseases—A critical review. Trends Food Sci. Technol. 2018, 71, 132–154. [Google Scholar] [CrossRef]
- Tsubone, H.; Makimura, Y.; Hanafusa, M.; Yamamoto, Y.; Tsuru, Y.; Motoi, M.; Amano, S. Agaricus brasiliensis KA21 improves circulatory functions in spontaneously hypertensive rats. J. Med. Food 2014, 17, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Sukalingam, K.; Ganesan, K. Rhesus blood groups associated with risk to obesity and diabetes mellitus: A report on Punjabi population in Selangor, Malaysia. Int. J. Integr. Med. Sci. 2015, 2, 105–109. [Google Scholar]
- Geng, X.; Tian, G.; Zhang, W.; Zhao, Y.; Zhao, L.; Ryu, M.; Wang, H.; Ng, T.B. Isolation of an Angiotensin I-Converting Enzyme inhibitory protein with antihypertensive effect in spontaneously hypertensive rats from the edible wild mushroom Leucopaxillus tricolor. Molecules 2015, 20, 10141–10153. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.C.; Abdullah, N.; Shuib, A.S.; Aminudin, N. Novel angiotensin I-converting enzyme inhibitory peptides derived from edible mushroom Agaricus bisporus (J.E. Lange) Imbach identified by LC-MS/MS. Food Chem. 2014, 148, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Sowers, J.R. Diabetes mellitus and vascular disease. Hypertension 2013, 61, 943–947. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, M.; Nakano, M.; Morii, Y.; Ohashi, T.; Fujiwara, Y.; Sonoyama, K. Hepatic LDL receptor mRNA in rats is increased by dietary mushroom (Agaricus bisporus) fiber and sugar beet fiber. J. Nutr. 2000, 130, 2151–2156. [Google Scholar] [CrossRef] [PubMed]
- Kurukulasuriya, L.R.; Stas, S.; Lastra, G.; Manrique, C.; Sowers, J.R. Hypertension in obesity. Med. Clin. N. Am. 2011, 95, 903–917. [Google Scholar] [CrossRef] [PubMed]
- Ece, I.; Yilmaz, H.; Alptekin, H.; Yormaz, S.; Colak, B.; Yilmaz, F.; Sahin, M. Comparative effectiveness of laparoscopic sleeve gastrectomy on morbidly obese, super-obese, and super-super obese patients for the treatment of morbid obesity. Obes. Surg. 2017, 28, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.C.; Jeong, Y.T.; Yang, B.K.; Islam, R.; Koyyalamudi, S.R.; Pang, G.; Cho, K.Y.; Song, C.H. White button mushroom (Agaricus bisporus) lowers blood glucose and cholesterol levels in diabetic and hypercholesterolemic rats. Nutr. Res. 2010, 30, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Gani, S.B. Attenuation of curcumin on isoproterenol-induced alterations in cardiac marker enzymes in Wistar albino rats. Int. J. Appl. Biosci. 2014, 2, 78–83. [Google Scholar]
- Rizvi, A.A. Addressing hypertension in the patient with type 2 diabetes mellitus: Pathogenesis, goals, and therapeutic approach. Eur. Med. J. Diabetes 2017, 5, 84–92. [Google Scholar] [PubMed]
- Ganesan, K.; Gani, S.B. Relationship between ABO, Rh Blood Groups, and Diabetes Mellitus, obesity in Namakkal town, Tamilnadu. Int. J. Adv. Pharm. Biol. Chem. 2014, 3, 995–998. [Google Scholar]
- De Pergola, G.; Silvestris, F. Obesity as a major risk factor for cancer. J. Obes. 2013, 2013, 291546. [Google Scholar] [CrossRef] [PubMed]
- Allott, E.H.; Masko, E.M.; Freedland, S.J. Obesity and prostate cancer: Weighing the evidence. Eur. Urol. 2013, 63, 800–809. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Sukalingam, K.; Xu, B. Impact of consumption of repeatedly heated cooking oils on the incidence of various cancers—A critical review. Crit. Rev. Food Sci. Nutr. 2017, 58, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B. Molecular targets of vitexin and isovitexin in cancer therapy: A critical review. Ann. N.Y. Acad. Sci. 2017, 1401, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Wang, Z.; Li, Y.; Kong, D.; Ali, S. The complexities of obesity and diabetes with the development and progression of pancreatic cancer. Biochim. Biophys. Acta 2011, 1815, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Anderson, D.; Lurie-Beck, J. The relationship between abdominal obesity and depression in the general population: A systematic review and meta-analysis. Obes. Res. Clin. Pract. 2011, 5, e267–e278. [Google Scholar] [CrossRef] [PubMed]
- Hagan, K.A.; Munger, K.L.; Ascherio, A.; Grodstein, F. Epidemiology of Major Neurodegenerative Diseases in Women: Contribution of the Nurses’ Health Study. Am. J. Public Health 2016, 26, 1650–1655. [Google Scholar] [CrossRef] [PubMed]
- Brewer, C.J.; Balen, A.H. The adverse effects of obesity on conception and implantation. Reproduction 2010, 140, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Tchernof, A.; Després, J.-P. Pathophysiology of Human Visceral Obesity: An Update. Physiol. Rev. 2013, 93, 359–404. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.M.; Jones, T.H. Testosterone and obesity. Obes. Rev. 2015, 16, 581–606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.D.; Li, H.C.; Gao, M.; Wang, L.; Deng, Q.; Shi, T.; He, J.L.; Chong, T.; Wang, Z.M. Sexual development characteristics and sex hormone levels in obese male adolescents. Zhonghua Nan Ke Xue 2013, 19, 434–438. [Google Scholar] [PubMed]
- Steenackers, N.; Gesquiere, I.; Matthys, C. The relevance of dietary protein after bariatric surgery: What do we know? Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Frith, E.; Loprinzi, P.D. The protective effects of a novel fitness-fatness index on all-cause mortality among adults with cardiovascular disease. Clin. Cardiol. 2017, 40, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Gontarev, S.; Kalac, R.; Zivkovic, V.; Velickovska, L.A.; Telai, B. The association between high blood pressure, physical fitness and fatness in adolescents. Nutr. Hosp. 2017, 34, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Kissane, N.A.; Pratt, J.S.A. Medical and surgical treatment of obesity. Best Pract. Res. Clin. Anaesthesiol. 2011, 25, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Bessesen, D.H.; Van Gaal, L.F. Progress and challenges in anti-obesity pharmacotherapy. Lancet Diabetes Endocrinol. 2018, 6, 237–248. [Google Scholar] [CrossRef]
- Banu, G.S.; Kumar, G.; Pandian, M.R. Cholesterol lowering activity of the aqueous fruit extract of Trichosanthes dioica Roxb. in normal and streptozotocin diabetic rats. J. Clin. Diagn. Res. 2007, 1, 561–569. [Google Scholar]
- Kumar, G.; Murugesan, A.G. Hypolipidaemic activity of Helicteres isora L. bark extracts in streptozotocin induced diabetic rats. J. Ethnopharmacol. 2008, 116, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Ramasamy, M.; Gani, S.B. Antihyperlipideamic effect of Solanum trilobatum L. leaves extract on streptozotocin induced diabetic rats. Asian J. Biomed. Pharm. Sci. 2013, 3, 51–56. [Google Scholar]
- Kumar, G.; Banu, G.S.; Murugesan, A.G. Attenuation of Helicteres isora L. bark extracts on streptozotocin-induced alterations in glycogen and carbohydrate metabolism in albino rats. Hum. Exp. Toxicol. 2009, 28, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Banu, G.S.; Murugesan, A.G. Effect of Helicteres isora bark extracts on heart antioxidant status and lipid peroxidation in streptozotocin diabetic rats. J. Appl. Biomed. 2008, 6, 89–95. [Google Scholar]
- Jayachandran, M.; Zhang, T.; Ganesan, K.; Xu, B.; Chung, S.S.M. Isoquercetin ameliorates hyperglycemia and regulates key enzymes of glucose metabolism via insulin signaling pathway in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2018, 829, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Mushroom polysaccharides: Chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.T.; Wasser, S.P. The Role of Culinary-Medicinal Mushrooms on Human Welfare with a Pyramid Model for Human Heath. Int. J. Med. Mushrooms 2012, 14, 95–134. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Yoon, S.J.; Pyun, Y.R. Polysaccharides from edible mushroom Hinmogi (Tremella fuciformis) inhibit differentiation of 3T3-L1 adipocytes by reducing mRNA expression of PPARγ, C/EBPα, and leptin. Food Sci. Biotechnol. 2008, 17, 267–273. [Google Scholar]
- Nyam, K.L.; Chow, C.F.; Tan, C.S.; Ng, S.T. Antidiabetic Properties of the Tiger’s milk medicinal mushroom, Lignosus rhinocerotis (Agaricomycetes), in streptozotocin-induced diabetic rats. Int. J. Med. Mushrooms 2017, 19, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Bao, L.; Xiong, W.; Ma, K.; Han, J.; Wang, W.; Yin, W.; Liu, H. Lanostane triterpenes from the tibetan medicinal mushroom Ganoderma leucocontextum and their inhibitory effects on HMG-CoA reductase and α-glucosidase. J. Nat. Prod. 2015, 78, 1977–1989. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Li, Y.; Zhou, T.; Xu, D.P.; Zhang, P.; Li, S.; Li, H.B. Bioactivities and health benefits of mushrooms mainly from China. Molecules 2016, 21, 938. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Cheung, R.C.; Wong, J.H.; Chan, Y.S.; Dan, X.; Pan, W.; Wang, H.; Guan, S.; Chan, K.; Ye, X.; et al. Fungal proteinaceous compounds with multiple biological activities. Appl. Microbiol. Biotechnol. 2016, 100, 6601–6617. [Google Scholar] [CrossRef] [PubMed]
- Delzenne, N.M.; Bindels, L.B. Gut microbiota: Ganoderma lucidum, a new prebiotic agent to treat obesity? Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 553–554. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D. Obesity: Medicinal mushroom reduces obesity by modulating microbiota. Nat. Rev. Endocrinol. 2015, 11, 504. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Korivi, M.; Yang, H.T.; Huang, C.C.; Chaing, Y.Y.; Tsai, Y.C. Effect of Pleurotus tuber-regium polysaccharides supplementation on the progression of diabetes complications in obese-diabetic rats. Chin. J. Physiol. 2014, 57, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, N.; Zhang, J.; Zhao, H.; Lin, L.; Jia, S.; Jia, L. Antihyperlipidemic and hepatoprotective activities of residue polysaccharide from Cordyceps militaris SU-12. Carbohydr. Polym. 2015, 131, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Manson, J.E.; Stampfer, M.J.; Rexrode, K.M.; Willett, W.C.; Hu, F.B. A prospective study of trans fatty acids in erythrocytes and risk of coronary heart disease. Circulation 2007, 115, 1858–1865. [Google Scholar] [CrossRef] [PubMed]
- Bobek, P.; Hromadová, M.; Ozdín, L. Oyster mushroom (Pleurotus ostreatus) reduces the activity of 3-hydroxy-3-methylglutaryl CoA reductase in rat liver microsomes. Experientia 1995, 51, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Furuncuoğlu, Y.; Tulgar, S.; Dogan, A.N.; Cakar, S.; Tulgar, Y.K.; Cakiroglu, B. How obesity affects the neutrophil/lymphocyte and platelet/lymphocyte ratio, systemic immune-inflammatory index and platelet indices: A retrospective study. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1300–1306. [Google Scholar] [PubMed]
- Mao, G.H.; Ren, Y.; Feng, W.W.; Li, Q.; Wu, H.Y.; Jin, D.; Zhao, T.; Xu, C.Q.; Yang, L.Q.; Wu, X.Y. Antitumor and immunomodulatory activity of a water-soluble polysaccharide from Grifola frondosa. Carbohydr. Polym. 2015, 134, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Thyagarajan-Sahu, A.; Lane, B.; Sliva, D. ReishiMax, mushroom-based dietary supplement, inhibits adipocyte differentiation, stimulates glucose uptake and activates AMPK. BMC Complement. Altern. Med. 2011, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Cheskin, L.J.; Davis, L.M.; Lipsky, L.M.; Mitola, A.H.; Lycan, T.; Mitchell, V.; Mickle, B.; Adkins, E. Lack of energy compensation over 4 days when white button mushrooms are substituted for beef. Appetite 2008, 51, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Poddar, K.H.; Ames, M.; Hsin-Jen, C.; Feeney, M.J.; Wang, Y.; Cheskin, L.J. Positive effect of mushrooms substituted for meat on body weight, body composition, and health parameters. A 1-year randomized clinical trial. Appetite 2013, 71, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.P.; Park, S.O.; Lee, S.J.; Nam, S.H.; Friedman, M. A polysaccharide isolated from the liquid culture of Lentinus edodes (Shiitake) mushroom mycelia containing black rice bran protects mice against a Salmonella lipopolysaccharide-induced endotoxemia. J. Agric. Food Chem. 2013, 61, 10987–10994. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Hayes, M. Bioactive peptides from meat muscle and by-products: Generation, functionality, and application as functional ingredients. Meat Sci. 2014, 98, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.G.; Kim, Y.H.; Bolormaa, Z.; Kim, M.K.; Seo, G.S.; Lee, J.S. Characterization of an antihypertensive angiotensin I-converting enzyme inhibitory peptide from the edible mushroom Hypsizygus marmoreus. BioMed Res. Int. 2013, 2013, 283964. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.C.; Abdullah, N.; Shuib, A.S. Novel angiotensin I-converting enzyme inhibitory peptides derived from an edible mushroom, Pleurotus cystidiosus O.K. Miller identified by LC-MS/MS. BMC Complement. Altern. Med. 2013, 13, 313. [Google Scholar] [CrossRef] [PubMed]
- Alam, N.; Yoon, K.N.; Lee, T.S.; Lee, U.Y. Hypolipidemic activities of dietary Pleurotus ostreatus in hypercholesterolemic rats. Mycobiology 2011, 39, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Jeong, S.C.; Kim, J.H.; Lee, Y.H.; Ju, Y.C.; Lee, J.S. Characterization of a new antihypertensive angiotensin I-converting enzyme inhibitory peptide from Pleurotus cornucopiae. Food Chem. 2011, 127, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Reza, M.A.; Hossain, M.A.; Damte, D.; Jo, W.S.; Hsu, W.H.; Park, S.C. Hypolipidemic and hepatic steatosis preventing activities of the wood ear medicinal mushroom Auricularia auricula-judae (Higher Basidiomycetes) ethanol extract in vivo and in vitro. Int. J. Med. Mushrooms 2015, 17, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, M.; Ohashi, T.; Fujiwara, Y.; Sonoyama, K.; Nakano, M. Cholesterol-lowering effects of maitake (Grifola frondosa) fiber, shiitake (Lentinus edodes) fiber, and enokitake (Flammulina velutipes) fiber in rats. Exp. Biol. Med. 2001, 226, 758–765. [Google Scholar] [CrossRef]
- Bang, T.H.; Suhara, H.; Doi, K.; Ishikawa, H.; Fukami, K.; Parajuli, G.P.; Katakura, Y.; Yamashita, S.; Watanabe, K.; Adhikari, M.K. Wild mushrooms in Nepal: Some potential candidates as antioxidant and ACE-inhibition sources. J. Evid. Based Complement. Altern. Med. 2014, 2014, 195305. [Google Scholar]
- Xie, Y.; Zhang, H.; Liu, H.; Xiong, L.; Gao, X.; Jia, H.; Lian, Z.; Tong, N.; Han, T. Hypocholesterolemic effects of Kluyveromyces marxianus M3 isolated from Tibetan mushrooms on diet-induced hypercholesterolemia in rat. Braz. J. Microbiol. 2015, 46, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Cimerman, N.G.; Cimerman, A. Pleurotus fruiting bodies contain the inhibitor of 3-hydroxy-3-methylglutaryl-coenzyme A reductase—lovastatin. Exp. Mycol. 1995, 19, 1–6. [Google Scholar] [CrossRef]
- Zhao, S.; Rong, C.; Liu, Y.; Xu, F.; Wang, S.; Duan, C.; Chen, J.; Wu, X. Extraction of a soluble polysaccharide from Auricularia polytricha and evaluation of its anti-hypercholesterolemic effect in rats. Carbohydr. Polym. 2015, 122, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Vitak, T.Y.; Wasser, S.P.; Nevo, E.; Sybirna, N.O. The effect of the medicinal mushrooms Agaricus brasiliensis and Ganoderma lucidum (Higher Basidiomycetes) on the erythron system in normal and streptozotocin-induced diabetic rats. Int. J. Med. Mushrooms 2015, 17, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Yurkiv, B.; Wasser, S.P.; Nevo, E.; Sybirna, N.O. Antioxidant Effects of Medicinal Mushrooms Agaricus brasiliensis and Ganoderma lucidum (Higher Basidiomycetes): Evidence from Animal Studies. Int. J. Med. Mushrooms 2015, 17, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Tian, G.; Zhang, W.; Zhao, Y.; Zhao, L.; Wang, H.; Ng, T.B. A Tricholoma matsutake peptide with Angiotensin Converting Enzyme inhibitory and antioxidative activities and antihypertensive effects in spontaneously hypertensive rats. Sci. Rep. 2016, 6, 24130. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, D.; You, Y.; Zeng, S.; Hu, Y.; Duan, X.; Liu, A.; Chen, H.; Hu, X.; Chen, S.; et al. Structural characterization and antidiabetic activity of a glucopyranose-rich heteropolysaccharide from Catathelasma ventricosum. Carbohydr. Polym. 2016, 149, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Ma, Y.; Geng, L.; Zhao, A.; Zheng, J.; Xu, C. Fermentation characteristics in stirred-tank reactor of exopolysaccharides with hypolipidemic activity produced by Pleurotus geesteranus 5#. An. Acad. Bras. Cienc. 2013, 85, 1473–1481. [Google Scholar]
- Lin, L.; Cui, F.; Zhang, J.; Gao, X.; Zhou, M.; Xu, N.; Zhao, H.; Liu, M.; Zhang, C.; Jia, L. Antioxidative and renoprotective effects of residue polysaccharides from Flammulina velutipes. Carbohydr. Polym. 2016, 146, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lv, G.; He, W.; Shi, L.; Pan, H.; Fan, L. Effects of extraction methods on the antioxidant activities of polysaccharides obtained from Flammulina velutipes. Carbohydr. Polym. 2013, 98, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Mohamad Ansor, N.; Abdullah, N.; Aminudin, N. Anti-angiotensin converting enzyme (ACE) proteins from mycelia of Ganoderma lucidum (Curtis) P. Karst. BMC Complement. Altern. Med. 2013, 13, 256. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Cho, H.Y.; Yang, H.C.; Ra, K.S.; Suh, H.J. Angiotensin I-converting enzyme inhibitor from Grifola frondosa. Food Res. Int. 2001, 34, 177–182. [Google Scholar] [CrossRef]
- Hiwatashi, K.; Kosaka, Y.; Suzuki, N.; Hata, K.; Mukaiyama, T.; Sakamoto, K.; Shirakawa, H.; Komai, M. Yamabushitake mushroom (Hericium erinaceus) improved lipid metabolism in mice fed a high-fat diet. Biosci. Biotechnol. Biochem. 2010, 74, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.K.; Park, J.B.; Song, C.H. Hypolipidemic effect of an Exo-biopolymer produced from a submerged mycelial culture of Hericium erinaceus. Biosci. Biotechnol. Biochem. 2003, 67, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Grdović, N.; Dinić, S.; Arambašić, J.; Mihailović, M.; Uskoković, A.; Marković, J.; Poznanović, G.; Vidović, S.; Zeković, Z.; Mujić, A.; et al. The protective effect of a mix of Lactarius deterrimus and Castanea sativa extracts on streptozotocin-induced oxidative stress and pancreatic β-cell death. Br. J. Nutr. 2012, 108, 1163–1176. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Hwang, I.; Kim, S.; Hong, E.J.; Jeung, E.B. Lentinus edodes promotes fat removal in hypercholesterolemic mice. Exp. Ther. Med. 2013, 6, 1409–1413. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.N.; Lee, J.S.; Kim, H.Y.; Lee, K.R.; Shin, P.G.; Cheong, J.C.; Yoo, Y.B.; Alam, N.; Ha, T.M.; Lee, T.S. Appraisal of Antihyperlipidemic Activities of Lentinus lepideus in Hypercholesterolemic Rats. Mycobiology 2011, 39, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Spim, S.R.; de Oliveira, B.G.; Leite, F.G.; Gerenutti, M.; Grotto, D. Effects of Lentinula edodes consumption on biochemical, hematologic and oxidative stress parameters in rats receiving high-fat diet. Eur. J. Nutr. 2017, 56, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.N.; Alam, N.; Lee, J.S.; Cho, H.J.; Kim, H.Y.; Shim, M.J.; Lee, M.W.; Lee, T.S. Antihyperlipidemic effect of dietary Lentinus edodes on plasma, feces and hepatic tissues in hypercholesterolemic rats. Mycobiology 2011, 39, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Dhingra, G.S.; Shri, R. Evaluation of carbohydrate metabolism inhibition by some species of medicinal mushrooms from India. Int. J. Med. Mushrooms 2015, 17, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.C.; Lee, D.H.; Kim, J.H.; Yu, H.E.; Park, J.S.; Lee, J.S. Production and characterization of antihypertensive angiotensin I-converting enzyme inhibitor from Pholiota adiposa. J. Microbiol. Biotechnol. 2006, 16, 757–763. [Google Scholar]
- Zheng, L.; Zhai, G.; Zhang, J.; Wang, L.; Ma, Z.; Jia, M.; Jia, L. Antihyperlipidemic and hepatoprotective activities of mycelia zinc polysaccharide from Pholiota nameko SW-02. Int. J. Biol. Macromol. 2014, 70, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, L.; Fang, J.C.; Wong, J.H.; Ng, T.B.; Jiang, Y.; Wang, C.R.; Zhang, N.Y.; Wen, T.Y.; Qu, L.Y. Isolation and identification of a novel polysaccharide-peptide complex with antioxidant, antiproliferative and hypoglycaemic activities from the abalone mushroom. Biosci. Rep. 2012, 32, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Meng, G.; Zhang, C.; Lin, L.; Xu, N.; Liu, M.; Cui, F.; Jia, L. The antioxidative effects of acidic-, alkalic-, and enzymatic-extractable mycelium zinc polysaccharides by Pleurotus djamor on liver and kidney of streptozocin-induced diabetic mice. BMC Complement. Altern. Med. 2015, 15, 440. [Google Scholar] [CrossRef] [PubMed]
- Alam, N.; Yoon, K.N.; Lee, J.S.; Cho, H.J.; Shim, M.J.; Lee, T.S. Dietary effect of Pleurotus eryngii on biochemical function and histology in hypercholesterolemic rats. Saudi J. Biol. Sci. 2011, 18, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Alam, N.; Yoon, K.N.; Lee, T.S. Antihyperlipidemic activities of Pleurotus ferulae on biochemical and histological function in hypercholesterolemic rats. J. Res. Med. Sci. 2011, 16, 776–786. [Google Scholar] [PubMed]
- Ibadallah, B.X.; Abdullah, N.; Shuib, A.S. Identification of angiotensin-converting enzyme inhibitory proteins from mycelium of Pleurotus pulmonarius (oyster mushroom). Planta Med. 2015, 81, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.N.; Alam, N.; Shim, M.J.; Lee, T.S. Hypolipidemic and antiatherogenesis effect of culinary-medicinal pink oyster mushroom, Pleurotus salmoneostramineus L. Vass. (higher Basidiomycetes), in hypercholesterolemic rats. Int. J. Med. Mushrooms 2012, 14, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Xu, B. Antidiabetic and antioxidant activities of eight medicinal mushroom species from China. Int. J. Med. Mushrooms 2015, 17, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Hyoung Lee, D.; Ho Kim, J.; Sik Park, J.; Jun Choi, Y.; Soo Lee, J. Isolation and characterization of a novel angiotensin I-converting enzyme inhibitory peptide derived from the edible mushroom Tricholoma giganteum. Peptides 2004, 25, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Bobek, P.; Ozdín, L.; Kuniak, L.; Hromadová, M. Regulation of cholesterol metabolism with dietary addition of oyster mushrooms (Pleurotus ostreatus) in rats with hypercholesterolemia. Cas Lek Cesk 1997, 136, 186–190. [Google Scholar] [PubMed]
- Yang, B.K.; Kim, G.N.; Jeong, Y.T.; Jeong, H.; Mehta, P.; Song, C.H. Hypoglycemic effects of exo-biopolymers produced by five different medicinal mushrooms in STZ-induced diabetic rats. Mycobiology 2008, 36, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Yang, U.K.; Jeong, S.C.; Lee, H.J.; Sohn, D.H.; Song, C.H. Antidiabetic and hypolipidemic effects of Collybia confluens mycelia produced by submerged culture in streptozotocin—Diabetic rats. Arch. Pharmacol. Res. 2006, 29, 73–79. [Google Scholar] [CrossRef]
- Chen, X.; Wu, G.; Huang, Z. Structural analysis and antioxidant activities of polysaccharides from cultured Cordyceps militaris. Int. J. Biol. Macromol. 2013, 58, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Yeh, M.Y.; Ko, W.C.; Lin, L.Y. Hypolipidemic and antioxidant activity of enoki mushrooms (Flammulina velutipes). BioMed Res. Int. 2014, 2014, 352385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.N.; Lin, Z.B. Hypoglycemic effect of Ganoderma lucidum polysaccharides. Acta Pharmacol. Sin. 2004, 25, 191–195. [Google Scholar] [PubMed]
- Li, F.; Zhang, Y.; Zhong, Z. Antihyperglycemic effect of Ganoderma lucidum polysaccharides on streptozotocin-induced diabetic mice. Int. J. Mol. Sci. 2011, 12, 6135–6145. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Yang, B.; Yu, Y.; Chen, Q.; Huang, T.; Li, D. Ganoderma lucidum polysaccharides exert anti-hyperglycemic effect on streptozotocin-induced diabetic rats through affecting β-cells. Comb. Chem. High Throughput Screen 2012, 15, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.K.; Kim, D.H.; Jeong, S.C.; Das, S.; Choi, Y.S.; Shin, J.S.; Lee, S.C.; Song, C.H. Hypoglycemic effect of a Lentinus edodes exo-polymer produced from a submerged mycelial culture. Biosci. Biotechnol. Biochem. 2002, 66, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Kim, S.W.; Lim, J.M.; Joo, J.H.; Kim, H.O.; Kim, H.M.; Yun, J.W. Hypoglycemic effect of crude exopolysaccharides produced by a medicinal mushroom Phellinus baumii in streptozotocin-induced diabetic rats. Life Sci. 2005, 76, 3069–3080. [Google Scholar] [CrossRef] [PubMed]
- Rony, K.A.; Ajith, T.A.; Mathew, J.; Janardhanan, K.K. The medicinal cracked-cap polypore mushroom Phellinus rimosus (higher Basidiomycetes) attenuates alloxan-induced hyperglycemia and oxidative stress in rats. Int. J. Med. Mushrooms 2013, 15, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Rony, K.A.; Ajith, T.A.; Janardhanan, K.K. Hypoglycemic and hypolipidemic effects of the cracked-cap medicinal mushroom Phellinus rimosus (Higher Basidiomycetes) in streptozotocin-induced diabetic rats. Int. J. Med. Mushrooms 2015, 17, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.J.; Hwang, H.J.; Kim, S.W.; Oh, J.Y.; Baek, Y.M.; Choi, J.W.; Bae, S.H.; Yun, J.W. Hypoglycemic effects of exopolysaccharides produced by mycelial cultures of two different mushrooms Tremella fuciformis and Phellinus baumii in ob/ob mice. Appl. Microbiol. Biotechnol. 2007, 75, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lee, H.S.; Kim, S.H.; Moon, B.; Lee, C. Antioxidant and anti-inflammatory activities of methanol extracts of Tremella fuciformis and its major phenolic acids. J. Food Sci. 2014, 79, C460–C468. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.K.; Jung, Y.S.; Song, C.H. Hypoglycemic effects of Ganoderma applanatum and Collybia confluens exo-polymers in streptozotocin-induced diabetic rats. Phytother. Res. 2007, 21, 1066–1069. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Edible/Medicinal Mushroom | Botanical Name | Study Model/Methods | Bioactive Compounds | References |
|---|---|---|---|---|
| Edible | Agaricus campestris | Hypercholesterolemic diet and STZ induced rats-plasma glucose, TG, TC, ALT, AST and LDL | Vitamin C, D, B12, folates, and polyphenols | [63] |
| Edible | Agaricus bisporus | High-fat diet in rats-serum cholesterol and hepatic LDL receptor mRNA | Fibers | [32] |
| Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [30] | ||
| Edible | Agaricus brasiliensis | STZ-induced diabetic rats-plasma glucose, TG, TC, glycated hemoglobin, TBARS | Polyphenols and flavonoids | [94,95] |
| Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [27] | ||
| Edible | Boletus bicolor | Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [96] |
| Edible | Leucopaxillus tricolor | Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [29] |
| Edible | Catathelasma ventricosum | STZ induced diabetic rats-plasma glucose, total TC, TG | Heteropolysaccharide | [97] |
| Edible | Pleurotus geesteranus | STZ induced diabetic rats-plasma glucose, total TC, TG | Polysaccharides | [98] |
| Edible | Flammulina velutipes | STZ-induced diabetic rats-SOD, GSH-Px, CAT, MDA, ALT, AST, BUN, CRE, TC, LDL-C and HDL-C | Polysaccharides | [99] |
| In vitro-DPPH free radical & Hydroxy radical scavenging, in vitro α-glycosidase, aldose reductase inhibitory assays | Polysaccharides | [100] | ||
| Edible | G. lucidum | Hypertensive rats-ACE assay | Oligopeptide | [101] |
| Edible | Gloeostereum incarnatum | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Edible | Grifola frondosa | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Hypertensive rats-ACE assay | Oligopeptide | [102] | ||
| Edible | Hericium erinaceus | C57BL/6J mice model-serum and hepatic TG levels | Flavonoids | [103] |
| Hyperlipidemic rats-plasma total cholesterol, LDL, HDL, cholesterol, triglyceride, phospholipid, atherogenic index, and hepatic HMG-CoA reductase | Exo-polymer | [104] | ||
| Edible | Hypsizygus marmoreus | Hypertensive rats-ACE assay | Oligopeptide | [84] |
| Edible | Lactarius deterrimus | STZ-induced diabetic rats-plasma glucose, TG, glycated hemoglobin, glycated serum protein, and AGE, SOD, CAT, GSH levels | Polyphenols and flavonoids | [105] |
| Edible | Lentinula edodes | High-fat diet in rats-TG, TC, LDL, cholesterol 7-α-hydroxylase 1 | Lentinan KS-2 | [106] |
| High-fat diet in rats-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio, BIL, CRE, Urea, BUN, Uric acid, Total protein, Na, Ca, Cl, K, albumin, P, Mg | Lentinan KS-2 | [107] | ||
| High-fat diet in rabbits-TC, histological, immunohistochemical and morphometrically analysis | Lentinan KS-2 | [8] | ||
| High-fat diet in rats-TG, TC, ALT, AST, Urea, glucose, malondialdehyde | Lentinan KS-2 | [108] | ||
| Edible | Lentinus lepideus | High-fat diet in rats-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio, BIL, CRE, Urea, BUN, Uric acid, Total protein, Na, Ca, Cl, K, albumin, P, Mg | Lentinan KS-2, flavonoids | [109] |
| Edible | Lenzites elegans | in vitro enzymatic starch digestion assay | Polyphenols and flavonoids | [110] |
| Edible | Morchella vulgaris | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Edible | Oudemansiella radicata | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Edible | Pholiota adipose | Hypertensive rats-ACE assay | Oligopeptide | [111] |
| Edible | Pholiota nameko SW-02 | mice hyperlipidemic models-blood lipid levels (TC, TG, HDL-C, LDL-C, and VLDL-C), liver lipid levels (TC and TG) and antioxidant status (SOD, T-AOC, MDA, and LPO) | Mycelia zinc polysaccharide | [112] |
| Edible | Pleurotus abalonus | Diabetic mice-Inhibition of the proliferation of hepatoma HepG2 cells and breast cancer MCF7 cells, antioxidant activity in erythrocyte hemolysis, blood glucose and TG | Polysaccharide-peptide complex LB-1b | [113] |
| Edible | Pleurotus cornucopiae | Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [87] |
| Edible | Pleurotus cystidiosus O.K. Miller | Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [85] |
| Edible | Pleurotus djamor | STZ-induced diabetic rats-SOD, GSH-Px, CAT, MDA, ALT, AST, BUN, CRE, TC, LDL-C and HDL-C | Mycelium zinc polysaccharides | [114] |
| Edible | Pleurotus eryngii | High-fat diet in rats-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio, BIL, CRE, Urea, BUN, Uric acid, Total protein, Na, Ca, Cl, K, albumin, P, Mg | Polysaccharides | [115] |
| Edible | Pleurotus ferulae | High-fat diet in rats-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio, BIL, CRE, Urea, BUN, Uric acid, Total protein, Na, Ca, Cl, K, albumin, P, Mg | Polysaccharides | [116] |
| Edible | Pleurotus ostreatus | High-fat diet in rats-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio, BIL, CRE, Urea, BUN, Uric acid, Total protein, Na, Ca, Cl, K, albumin, P, Mg | Polysaccharides | [86] |
| Edible | Pleurotus pulmonarius | Hypertensive rats-Angiotensin I-Converting Enzyme assay | Oligopeptide | [117] |
| Edible | Pleurotus salmoneostramineus L. Vass | High-fat diet in rats-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio, BIL, CRE, Urea, BUN, Uric acid, Total protein, Na, Ca, Cl, K, albumin, P, Mg | Polysaccharides | [118] |
| Edible | Pleurotus tuber-regium | Ob diabetic rats-TC, TG, LDL, HDL, and PPAR-α mRNA expression | Polysaccharides | [72] |
| Edible | Ramaria botrytoides | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Edible | Russula aeruginea | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Edible | Tremella fuciformis | in vitro α-glycosidase, aldose reductase inhibitory assays, DPPH free radical scavenging | Polyphenols and flavonoids | [119] |
| Edible | Tricholoma giganteum | Hypertensive rats-ACE assay | Oligopeptide | [120] |
| Edible | Tricholoma matsutake | Hypertensive rats-ACE assay | Oligopeptide | [96] |
| Edible | Tuber micheli | Hypertensive rats-ACE assay | Oligopeptide | [29] |
| Edible | Pleurotus ostreatus | TC content in serum, lipoproteins in the liver, and HMG-CoA reductase in liver microsomes | Polysaccharides | [121] |
| Inhibition of HMG CoA reductase-lovastatin | Polysaccharides | [92] | ||
| Edible | Adiantum capillus-veneris L. | High cholesterol diet fed Wistar rats-Pancreatic triacylglycerol lipase and α-amylase/α-glucosidase, OGTT, TC, TG | Polyphenols | [9] |
| Edible | Aster spathulifolius Maxim | High-fat diet fed Wistar rats-body weight gain, visceral fat pad weights, serum lipid levels, as well as hepatic lipid levels, numbers of lipid droplets, expression of fat intake-related gene ACC2 and lipogenesis-related genes (e.g., SREBP-1c, ACC1, FAS, SCD1, GPATR, AGPAT, and DGAT), fatty acid oxidation and thermogenesis-related genes (e.g., PPAR-α, ACO, CPT1, UCP2, and UCP3), phosphorylated AMPKα, phosphorylated ACC | Polysaccharides | [3] |
| Edible | Kluyveromyces marxianus | The high-fat diet fed Wistar rats-TC, TG, HDL-C, LDL-C, levels in the serum and liver, atherogenic index | Polysaccharides | [91] |
| Edible and Medicinal | Collybia peronata, Ganoderma australe, Ganoderma lingzhi, Heterobasidion linzhiense, Heterobasidion linzhiense, Inocybe sp., Inonotus andersonii, Lactarius hatsudake, Lenzites betulina, Panellus sp., Phellinus conchatus, Phellinus gilvus, Phlebia tremellosa, Postia stiptica, Rigidoporus sp., Trametes versicolor, Tricholoma caligatum | ACE Inhibitory Assay | Polyphenol | [90] |
| Medicinal | Armillariella mellea | STZ-induced diabetic rats-plasma glucose, TG, TC, ALT, AST | exo-biopolymers | [122] |
| Medicinal | Auricularia auricula-judae | High-fat diet in mice-phospholipids, liver enzymes, TG, glycerol, glycerol-3-phosphate dehydrogenase | Phenolic compound | [88] |
| Medicinal | Collybia confluens | STZ induced animal model-plasma glucose, total TC, TG, ALT, AST | Exo-polymer | [123] |
| Medicinal | Cordyceps militaris | In vitro-Superoxide anion, DPPH free radical & Hydroxy radical scavenging, In vitro-HMG-CoA reductase and α-glucosidase | Polysaccharides | [124] |
| mice hyperlipidemic models-blood lipid levels (TC, TG, HDL-C, LDL-C, and VLDL-C), liver lipid levels (TC and TG) and antioxidant status (SOD, T-AOC, MDA, and LPO) | Polysaccharides | [73] | ||
| Medicinal | Cordyceps sinensis | STZ-induced diabetic rats-plasma glucose, TG, TC, ALT, AST | exo-biopolymers | [122] |
| Medicinal | Coriolus versicolor | STZ-induced diabetic rats-plasma glucose, TG, TC, ALT, AST | exo-biopolymers | [122] |
| Medicinal | Flammulina velutipes | High-fat diet in rats-TC, LDL, body weight, food intake, liver weight, cecum weight, cecum pH, Cecal acetic acid, butyric acid, and total SCFA | Fibers | [89] |
| High-fat diet in male hamsters-TG, TC, LDL, total lipids, phospholipids, LDL/HDL ratio | Dietary fiber, polysaccharide, and mycosterol, | [125] | ||
| Medicinal | Fomes fomentarius | STZ-induced diabetic rats-plasma glucose, TG, TC, ALT, AST | Exo-biopolymers | [89] |
| Medicinal | Ganoderma leucocontextum | In vitro-HMG-CoA reductase and α-glucosidase | Lanostane (Triterpenes) | [65] |
| Medicinal | Ganoderma lucidum | STZ-induced diabetic rats-plasma glucose, TC, TG, glycated hemoglobin, TBARS | Polysaccharides | [94,95] |
| STZ-induced diabetic rats-plasma glucose, TG, TC, NO, SOD, CAT, GPx | Polysaccharides | [126,127,128] | ||
| in vitro α-glycosidase, aldose reductase inhibitory assays, DPPH free radical scavenging | Polyphenols and flavonoids | [119] | ||
| Medicinal | Ganoderma philippii | in vitro enzymatic starch digestion assay | Appanoxidic acid A | [110] |
| Medicinal | Grifola frondosa | High-fat diet in rats-TC, LDL, body weight, food intake, liver weight, cecum weight, cecum pH, Cecal acetic acid, butyric acid, and total short-chain fatty acid | Fiber | [89] |
| Medicinal | Lentinus edodes | STZ induced animal model-plasma glucose, total cholesterol, and triglyceride | Exo-polymer | [129] |
| High-fat diet in rats-TC, LDL, body weight, food intake, liver weight, cecum weight, cecum pH, Cecal acetic acid, butyric acid, and total short-chain fatty acid | Fibers | [89] | ||
| Medicinal | Paecilomyces japonica | STZ-induced diabetic rats-plasma glucose, TG, TC, ALT, AST | exo-biopolymers | [122] |
| Medicinal | Phellinus baumii | STZ induced diabetic rats-plasma glucose, TG, TC, ALT, AST | Heteropolysaccharides and two proteoglycans | [130] |
| Medicinal | Phellinus rimosus | Alloxan-induced diabetic rats-plasma glucose, OGTT, TC, TG, SOD, CAT, GPx, and GSH | Polysaccharides | [131] |
| STZ-induced diabetic rats-plasma glucose, lipid profile, ALT, AST, serum insulin, liver glycogen | Polysaccharides | [132] | ||
| Medicinal | Rigidoporus ulmarius | in vitro enzymatic starch digestion assay | Polysaccharides | [110] |
| Medicinal | Tremella fuciformis | ob/ob mice-Plasma glucose, OGTT, TG | Exopolysaccarides | [133] |
| In vitro-ABTS radical scavenging activity, DPPH radical scavenging activity, LDL oxidation; NO synthase expression in RAW 264.7 cells | Polyphenols and flavonoids | [134] | ||
| Medicinal | Ganoderma applanatum and Collybia confluens | STZ-induced diabetic rats-Plasma glucose, TC, TG | Exo-polymer | [135] |
| Medicinal | Auricularia polytricha | Serum total lipids and TC | Polysaccharides | [93] |
| Medicinal | Pleurotus sajor-caju (Fr.) Singer | C57BL/6J mice fed on a high-fat diet-body weight, serum lipid, and liver enzymes, protein carbonyl and lipid hydroperoxide levels, enzymic antioxidants (SOD, CAT, and GPx) activities, Expression of hormone-sensitive lipase, adipose triglyceride lipase, peroxisome proliferator-activated receptor gamma, sterol regulatory binding protein-1c, and lipoprotein lipase | β-glucan | [1] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganesan, K.; Xu, B. Anti-Obesity Effects of Medicinal and Edible Mushrooms. Molecules 2018, 23, 2880. https://doi.org/10.3390/molecules23112880
Ganesan K, Xu B. Anti-Obesity Effects of Medicinal and Edible Mushrooms. Molecules. 2018; 23(11):2880. https://doi.org/10.3390/molecules23112880
Chicago/Turabian StyleGanesan, Kumar, and Baojun Xu. 2018. "Anti-Obesity Effects of Medicinal and Edible Mushrooms" Molecules 23, no. 11: 2880. https://doi.org/10.3390/molecules23112880
APA StyleGanesan, K., & Xu, B. (2018). Anti-Obesity Effects of Medicinal and Edible Mushrooms. Molecules, 23(11), 2880. https://doi.org/10.3390/molecules23112880