Phytochemical and Biological Profile of Moricandia arvensis (L.) DC.: An Inhibitor of Pancreatic Lipase

 ,
, 

 and
and
Abstract
1. Introduction
2. Results
2.1. Phytochemical Profile
2.2. Antioxidant Activity
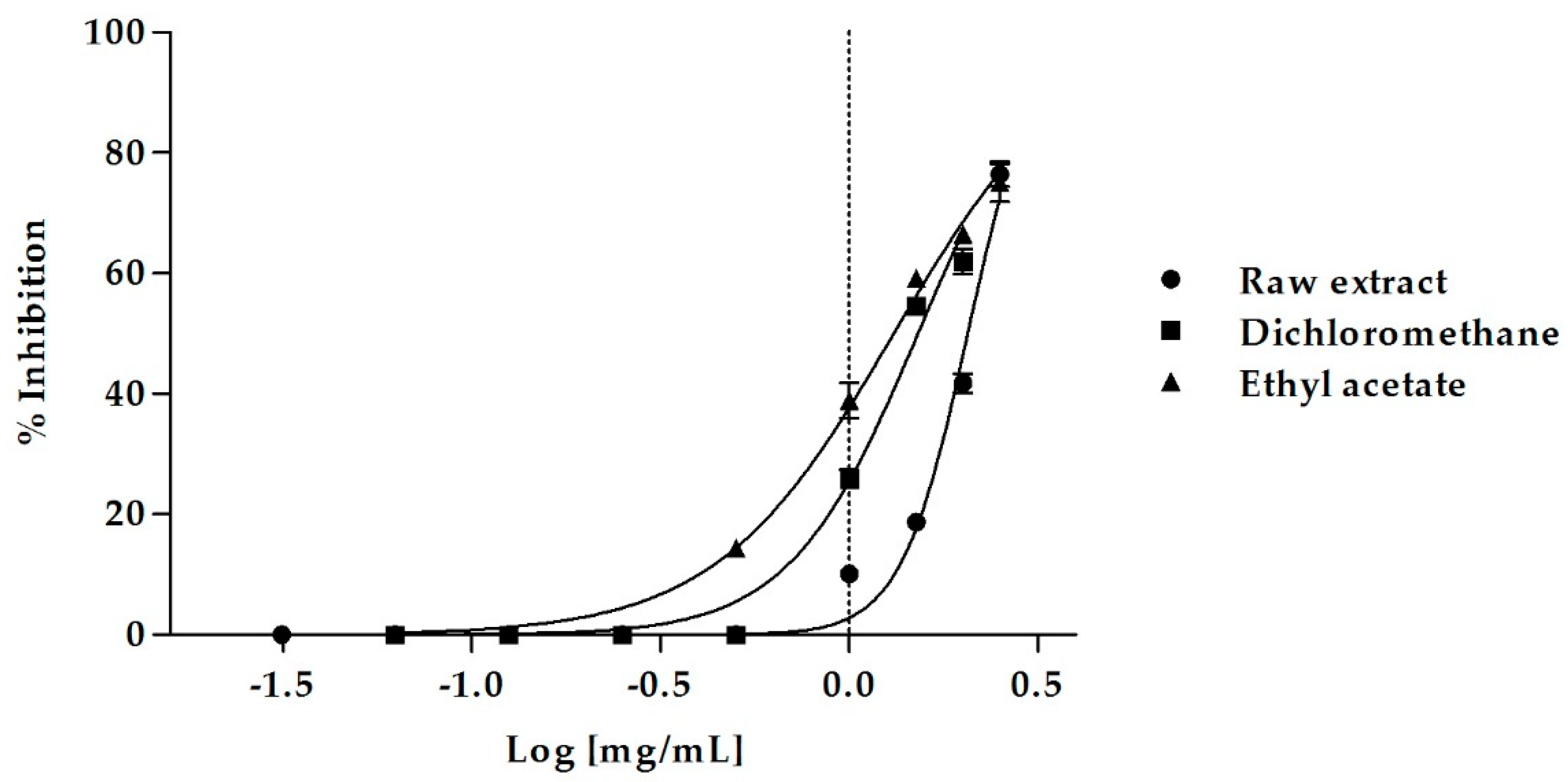
2.3. Pancreatic Lipase Inhibition
3. Discussion
4. Materials and Methods
4.1. Collection of Plant Material and Preparation of the Extracts
4.2. GC–MS Analysis
4.3. Determination of Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
4.4. Glucosinolates Extraction and Desulfation
4.5. Chemical Analysis and Quantitation of Glucosinolates
4.6. ESI-MS/MS Analyses of Glucosinolates
4.7. HPLC of Phenolics
4.8. HPLC-HRMS Analyses of Phenolics
4.9. Free Radical Scavenging Activity (FRSA) Assay
4.10. β-Carotene Bleaching/Linoleic Co-Oxidation Assay
4.11. In Vitro Pancreatic Lipase Assay
4.12. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Thurairajah, P.H.; Syn, W.K.; Neil, D.A.; Stell, D.; Haydon, G. Orlistat (xenical) induced subacute liver failure. Eur. J. Gastroenterol. Hepatol. 2005, 17, 1437–1438. [Google Scholar]
- Bray, G.A. Medical therapy for obesity. Mt. Sinai J. Med. 2010, 77, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Perri, V.; Menichini, F.; Marrelli, M.; Uzunov, D.; Statti, G.A.; Menichini, F. Wild Mediterranean dietary plants as inhibitors of pancreatic lipase. Phytother. Res. 2012, 26, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Fassina, P.; Scherer Adami, F.; Terezinha Zani, V.; Kasper Machado, I.C.; Garavaglia, J.; Quevedo Grave, M.T.; Ramos, R.; Morelo Dal Bosco, S. The effect of Garcinia cambogia as coadjuvant in the weight loss process. Nutr. Hosp. 2015, 32, 2400–2408. [Google Scholar] [PubMed]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar] [PubMed]
- Sun, N.-N.; Wu, T.-Y.; Chau, C.-F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef] [PubMed]
- Pignatti, S. Flora d’Italia; Edizione Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Skandrani, I.; Leloup, L.; Kovacic, H.; Dijoux-Franca, M.G.; Ghedira, K.; Ghedira, L.C. Antioxidant, antimutagenic, tanning and calpain induction activities of methanolic extract of Tunisian Plant (Moricandia arvensis). Iran. J. Pharm. Res. 2017, 16, 119–134. [Google Scholar] [PubMed]
- Belkhiri, A.; Lockwood, B.G. An indole derivative and glucosinolates from Moricandia arvensis. Phytochemistry 1990, 29, 1315–1316. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Screening crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.R.; Tsunoda, S. Fatty acid spectrum of mediterranean wild cruciferae. J. Am. Oil Chem. 1978, 55, 320–323. [Google Scholar] [CrossRef]
- Tatsuzawa, F.; Kato, K.; Sato, M.; Ito, S.; Muraoka, H.; Takahata, Y.; Ogawa, S. Acylated cyanidin 3-sophoroside-5-glucoside in purple-violet flowers of Moricandia arvensis (Brassicaceae). Nat. Prod. Commun. 2015, 10, 457–459. [Google Scholar] [CrossRef] [PubMed]
- Braham, H.; Mighri, Z.; Jannet, H.B.; Matthew, S.; Abreu, P.M. Antioxidant Phenolic Glycosides from Moricandia arvensis. J. Nat. Prod. 2005, 68, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Berreghioua, A.; Cheriti, A. Phytochemical investigation of the medicinal plant Moricandia arvensis L. from Algerian Sahara. Asian J. Pharm. Clin. Res. 2018, 5, 450–453. [Google Scholar] [CrossRef]
- Skandrani, I.; Boubaker, J.; Bouhlel, I.; Limem, I.; Ghedira, K.; Chekir-Ghedira, L. Leaf and root extracts of Moricandia arvensis protect against DNA damage in human lymphoblast cell K562 and enhance antioxidant activity. Environ. Toxicol. Pharmacol. 2010, 30, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Limem, I.; Neffati, A.; Boubaker, J.; Sghaier, M.B.; Bhouri, W.; Bouhlel, I.; Kilani, S.; Ghedira, K.; Chekir-Ghedira, L. Assessment of phenolic content, free-radical-scavenging capacity genotoxic and anti-genotoxic effect of aqueous extract prepared from Moricandia arvensis leaves. Food Chem. Toxicol. 2010, 48, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Bouhlel, I.; Limem, I.; Boubaker, J.; Bhouri, W.; Neffati, A.; Ben Sghaier, M.; Kilani, S.; Ghedira, K.; Ghedira-Chekir, L. Moricandia arvensis extracts protect against DNA damage, mutagenesis in bacteria system and scavenge the superoxide anion. Toxicol. In Vitro 2009, 23, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Pinon, A.; Simon, A.; Ghedira, K.; Chekir-Ghedira, L. Chloroform extract from Moricandia arvensis inhibits growth of B16-F0 melanoma cells and promotes differentiation in vitro. Cell Prolif. 2010, 43, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Boubaker, J.; Bhouri, W.; Limem, I.; Kilani, S.; Ben Sghaier, M.; Neffati, A.; Bouhlel, I.; Ghedira, K.; Chekir-Ghedira, L. Leaf extracts from Moricandia arvensis promote antiproliferation of human cancer cells, induce apoptosis, and enhance antioxidant activity. Drug Chem. Toxicol. 2010, 33, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Zeraib, A.; Ramdani, M.; Lograda, T.; Chalard, P.; Figueredo, G. Chemical Composition and Antimicrobial Activity of Essential Oil of Moricandia arvensis L. (DC.). Asian J. Plant Sci. 2011, 10, 342–346. [Google Scholar] [CrossRef]
- Wagner, H.; Bladt, S. Plant Drug Analysis: A Thin Layer Chromatography Atlas, 2nd ed.; Springer: Berlin, Germany, 2009. [Google Scholar]
- Avato, P.; Argentieri, M.P. Brassicaceae a rich source of health improving phytochemicals. Phytochem. Rev. 2015, 14, 1019–1033. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef] [PubMed]
- Marzouk, M.M.; Al-Nowaihi, A.-S.M.; Kawashty, S.A.; Saleh, N.A.M. Chemosystematic studies on certain species of the family Brassicaceae (Cruciferae) in Egypt. Biochem. System. Ecol. 2010, 38, 680–685. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Harnly, J.M. Phenolic component profiles of mustard greens, Yu Choy, and 15 other Brassica vegetables. J. Agric. Food Chem. 2010, 58, 6850–6857. [Google Scholar] [CrossRef] [PubMed]
- Velasco, P.; Francisco, M.; Moreno, D.; Ferreres, F.; Garcia-Viguera, C.; Cartea, M.E. Phytochemical fingerprinting of vegetable Brassica oleracea and Brassica napus by simultaneous identification of glucosinolates and phenolics. Phytochem. Anal. 2011, 22, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Identification of complex, naturally occurring flavonoid glycosides in kale (Brassica oleraceae var. sabellica) by high-performance liquid chromatography diode-array detection/electrospray ionization multi-stage mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2009–2022. [Google Scholar] [PubMed]
- Llorach, R.; Gil-Izquierdo, A.; Ferreres, F.; Tomas-Braberan, F.A. HPLC-DAD-MS/MS ESI characterization of unusual highly glycosylated acylated flavonoids from cauliflower (Brassica oleracea L. var. botrytis) agroindustrial byproducts. J. Agric. Food Chem. 2003, 51, 3895–3899. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Fernandes, F.; Oliveira, J.M.A.; Valentao, P.; Pereira, J.A. Metabolic profiling and biological capacity of Pieris brassicae fed with kale (Brassica oleracea L. var. acephala). Food Chem Toxicol. 2009, 47, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Skandrani, I.; Ben Sghaier, M.; Neffati, A.; Boubaker, J.; Bouhlel, I.; Kilani, S.; Mahmoud, A.; Gedira, K.; Chekir-Ghedira, L. Antigenotoxic and free radical scavenging activities of extracts from Moricandia arvensis. Drug Chem. Toxicol. 2007, 30, 361–382. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Conforti, F.; Araniti, F.; Statti, G.A. Effects of saponins on lipid metabolism: A review of potential health benefits in the treatment of obesity. Molecules 2016, 21, 1404. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Suresh, S.; Bayineni, V.K.; Kaeppagari, R.K. Lipase inhibitors from plants and their medical applications. Int. J. Pharm. Pharm. Sci. 2015, 7, 1–5. [Google Scholar]
- Birari, R.B.; Bhutani, K.K. Pancreatic lipase inhibitors from natural sources: Unexplored potential. Drug Discov. Today 2007, 129, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Daxenbilchler, M.E.; Spencer, G.F.; Carlson, D.G.; Rose, G.B.; Brinker, A.M.; Powell, R.G. Glucosinolate composition of seeds from 297 species of wild plants. Phytochemistry 1991, 30, 2623–2638. [Google Scholar] [CrossRef]
- Fiorentino, A.; Ricci, A.; D’Abrosca, B.; Golino, A.; Izzo, A.; Pascarella, M.T.; Picolella, S.; Esposito, A. Kempferol glycosides from Lobularia maritima and their potentail role in plant interactions. Chem. Biodivers. 2009, 6, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, G. Quercetin: A flavonol with multifaceted therapeutic applications? Fitoterapia 2015, 106, 256–271. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Lee, H.; Kim, S.; Park, J.; Ha, T. The anti-obesity effect of quercetin is mediated by the AMPK and MAPK signaling pathways. Biochem. Biophys. Res. Commun. 2008, 373, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Sharma, A.; Tuli, H.S.; Sak, K.; Punia, S.; Mukherjee, T.K. Kaempferol—A dietary anticancer molecule with multiple mechanisms of action: Recent trends and advancements. J. Funct. Food. 2017, 30, 203–219. [Google Scholar] [CrossRef]
- Bucholz, T.; Melzig, M.F. Polyphenolics compound as pancreatic lipase inhibitors. Planta Med. 2015, 81, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Argentieri, M.P.; Avato, P.; Menichini, F.; Conforti, F. Inhibitory effect on lipid absorption and variability of chemical constituents from Capparis sicula subsp. sicula and Capparis orientalis. Chem. Biodivers. 2016, 13, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Menichini, F.; Conforti, F. Hypolipidemic and Antioxidant Properties of Hot Pepper Flower (Capsicum annuum L.). Plant Foods Hum. Nutr. 2016, 71, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Menichini, G.; Alfano, C.; Marrelli, M.; Toniolo, C.; Provenzano, E.; Statti, G.A.; Nicoletti, M.; Menichini, F.; Conforti, F. Hypericum perforatum L. subsp. perforatum induces inhibition of free radicals and enhanced phototoxicity in human melanoma cells under ultraviolet light. Cell Prolif. 2013, 46, 193–202. [Google Scholar] [PubMed]
- Marrelli, M.; Menichini, F.; Conforti, F. A comparative study of Zingiber officinale Roscoe pulp and peel: Phytochemical composition and evaluation of antitumour activity. Nat. Prod. Res. 2015, 29, 2045–2049. [Google Scholar] [CrossRef] [PubMed]
- Argentieri, M.P.; Accogli, R.; Fanizzi, F.P.; Avato, P. Glucosinolates profile of “mugnolo” a variety of Brassica oleracea L. native to Southern Italy (Salento). Planta Med. 2011, 77, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Argentieri, M.P.; Macchia, F.; Papadia, P.; Fanizzi, F.P.; Avato, P. bioactive compounds from Capparis spinosa subsp. rupestris. Ind. Crop Prod. 2012, 36, 65–69. [Google Scholar] [CrossRef]
- Conforti, F.; Marrelli, M.; Statti, G.; Menichini, F. Antioxidant and cytotoxic activities of methanolic extract and fractions from Senecio gibbosus subsp. gibbosus (GUSS) DC. Nat. Prod. Res. 2006, 20, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Conforti, F.; Marrelli, M.; Statti, A.; Menichini, F.; Uzunov, D.; Solimene, U.; Menichini, F. Comparative chemical composition and antioxidant activity of Calamintha nepeta (L.) Savi subsp. glandulosa (Req.) Nyman and Calamintha grandiflora (L.) Moench (Labiatae). Nat. Prod. Res. 2012, 26, 91–97. [Google Scholar] [PubMed]
Sample Availability: Samples of the extracts are available from the authors. |


{kind=link}
| Compound 1 | RT 2 | RAP 3 |
|---|---|---|
| n-Hex | ||
| Fatty acids | ||
| Capric acid | 13.073 | 0.3 |
| Lauric acid | 15.027 | tr 4 |
| Palmitoleic acid | 15.119 | tr |
| Myristic acid | 16.799 | 0.6 |
| Stearic acid | 19.834 | 0.7 |
| Palmitic acid | 20.199 | 0.9 |
| Terpenes | ||
| Neophytadiene | 17.445 | 1.0 |
| Phytosterols | ||
| β-Sitosterol | 33.796 | tr |
| 22,24-Dimethylcholesterol | 33.893 | tr |
| Stigmasta-3,5-dien-7-one | 36.054 | 2.1 |
| DCM | ||
| Vanillin | 13.570 | tr |
| 2,4-Di-tert-butylphenol | 14.627 | 3.4 |
| Dihydroactinolide | 14.970 | 1.8 |
| Loliolide | 17.107 | 6.1 |
| Citronellyl acetone | 17.496 | tr |
| Hexahydrofarnesyl acetone | 17.525 | 3.4 |
| Number | Rt (min) | Name | UV (λmax, nm) | [M + H]+, m/z (%) | |
|---|---|---|---|---|---|
| 1 | 14.08 | Kaempferol-3-O-β-(2″-O-glucosyl)-rutinoside | 266.6, 319.9 sh, 338.9 | 757 (100) | 611 (1.0) [(M + H)-146]+; 595 (46) [(M + H)-162]+; 449 (4) [(M + H)-308]+; 287 (1) [(M + H)-308-162]+, [Aglycone + H]+ |
| 2 | 14.60 | Kaempferol-3-O-β-(2″-O-xylosyl-6″-O-rhamnosyl)-glucoside | 266.6, 319.9 sh, 338.9 | 727 (100) | 595 (37) [(M + H)-132]+; 581 (2) [(M + H)-146]+; 433 (6) [(M + H)-162-132]+; 287 (2) [(M + H)-132-162-146]+, [Aglycone + H]+ |
| 3 | 16.47 | Quercetin-3-O-β-sophoroside-7-O-α-rhamnoside | 257.2, 268.0 sh, 352.0 | 773 (100) | 611 (2) [(M + H)-162-]+; 449 (64) [(M + H)-162-162]+; 303 (18) [(M + H)-162-162-146]+, [Aglycone + H]+ |
| 4 | 16.81 | Quercetin-3-O-β-glucosyl-7-O-α-rhamnoside | 256.0, 268.0 sh, 354.4 | 611 (100) | 449 (71) [(M + H)-162]+; 303 (19) [(M + H)-162-146]+, [Aglycone + H]+ |
| 5 | 18.26 | Kaempferol-3-O-β-sophoroside-7-O-α-rhamnoside | 266.6, 319.0 sh, 343.6 | 757 (100) | 595 (2) [(M + H)-162]+; 433.1 (81) [(M + H)-162-162]+; 287.0 (28) [(M + H)-162-162-146]+, [Aglycone + H]+ |
| 6 | 18.28 | Kaempferol-3-O-β-glucosyl-7-O-α-rhamnoside | 266.6, 320.0 sh, 342.4 | 595 (100) | 433 (78) [(M + H)-162]+; 287 (23) [(M + H)-162-146]+, [Aglycone + H]+ |
| 7 | 18.95 | Kaempferol-3-O-α-arabinosyl-7-O-α-rhamnoside | 266.6, 344.8 sh, 347.2 | 565 (100) | 433 (100) [(M + H)-132]+; 419 (4) [(M + H)-146]+; 287 (23) [(M + H)-132-146]+, [Aglycone + H]+ |
| Compound | [M + Na]+ m/z | MS/MS (%) |
|---|---|---|
| 3-Hydroxypropyl-GLS | 320 | 219 (2) [(M + Na)-RCNOH]+; 100 (100) [CH2CHCH2NCS + H]+; 72 (10) [CH2NCS]+ |
| 3-Hydroxybutyl-GLS | 334 | 219 (35) [(M + Na)-RCNOH]+; 100 (100) [CH2CHCH2NCS + H]+ |
| Gluconapin (3-Butenyl-GLS) | 316 | 185 (3) [(M + Na)-RCNOH-H2S]+; 154 (2) [(M + Na)-Glu]+; 72 (10) [CH2NCS]+; 58 (19) [NCS]+ |
| Isobutyl-GLS | 318 | 186 (2) [(M + Na)-RCNOH-H2S]+; 100 (100) [CH2CHCH2NCS + H]+; 72 (10) [CH2NCS]+ |
| Glucoviorylin (2-(Methylthio)-ethyl-GLS) | 336 | 220 (38) [(M + Na)-RCNOH]+; 174 (2) [(M + Na)-Glu]+; 100 (100) [CH2CHCH2NCS + H]+; 72 (10) [CH2NCS]+ |
| Glucoiberverin (3-(Methylthio)-propyl-GLS) | 350 | 219 (15) [(M + Na)-RCNOH]+; 100 (100) [CH2CHCH2NCS + H]+; 58 (13) [NCS]+ |
| Glucotropaeolin (Benzyl-GLS) | 352 | 220 (2) [(M + Na)-RCNOH]+; 100 (100) [CH2CHCH2NCS + H]+; 72 (10) [CH2NCS]+ |
| Sample | IC50 (μg/mL) | ||
|---|---|---|---|
| DPPH Test | β-Carotene Bleaching Test | ||
| 30 min | 60 min | ||
| MeOH extract | 355.5 ± 7.9 c | 37.36 ± 3.06 b | >100 |
| n-Hex | >1000 | >100 | >100 |
| DCM | 870.7 ± 15.9 d | >100 | >100 |
| EtOAc | 171.9 ± 1.0 b | 35.69 ± 2.30 b | 63.92 ± 2.51 c |
| Ascorbic acid * | 2.00 ± 0.01 a | - | - |
| Propyl gallate * | - | 1.00 ± 0.02 a | 1.00 ± 0.02 a |
| Sample | IC50 (mg/mL) |
|---|---|
| MeOH extract | 2.06 ± 0.02 d |
| n-Hex | >10 |
| DCM | 1.52 ± 0.02 c |
| EtOAc | 1.31 ± 0.02 b |
| Orlistat * | 0.018 ± 0.001 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marrelli, M.; Morrone, F.; Argentieri, M.P.; Gambacorta, L.; Conforti, F.; Avato, P. Phytochemical and Biological Profile of Moricandia arvensis (L.) DC.: An Inhibitor of Pancreatic Lipase. Molecules 2018, 23, 2829. https://doi.org/10.3390/molecules23112829
Marrelli M, Morrone F, Argentieri MP, Gambacorta L, Conforti F, Avato P. Phytochemical and Biological Profile of Moricandia arvensis (L.) DC.: An Inhibitor of Pancreatic Lipase. Molecules. 2018; 23(11):2829. https://doi.org/10.3390/molecules23112829
Chicago/Turabian StyleMarrelli, Mariangela, Federica Morrone, Maria Pia Argentieri, Lucia Gambacorta, Filomena Conforti, and Pinarosa Avato. 2018. "Phytochemical and Biological Profile of Moricandia arvensis (L.) DC.: An Inhibitor of Pancreatic Lipase" Molecules 23, no. 11: 2829. https://doi.org/10.3390/molecules23112829
APA StyleMarrelli, M., Morrone, F., Argentieri, M. P., Gambacorta, L., Conforti, F., & Avato, P. (2018). Phytochemical and Biological Profile of Moricandia arvensis (L.) DC.: An Inhibitor of Pancreatic Lipase. Molecules, 23(11), 2829. https://doi.org/10.3390/molecules23112829