Reading More than Histones: The Prevalence of Nucleic Acid Binding among Reader Domains


Abstract
1. Introduction
1.1. Chromatin and Reader Domains
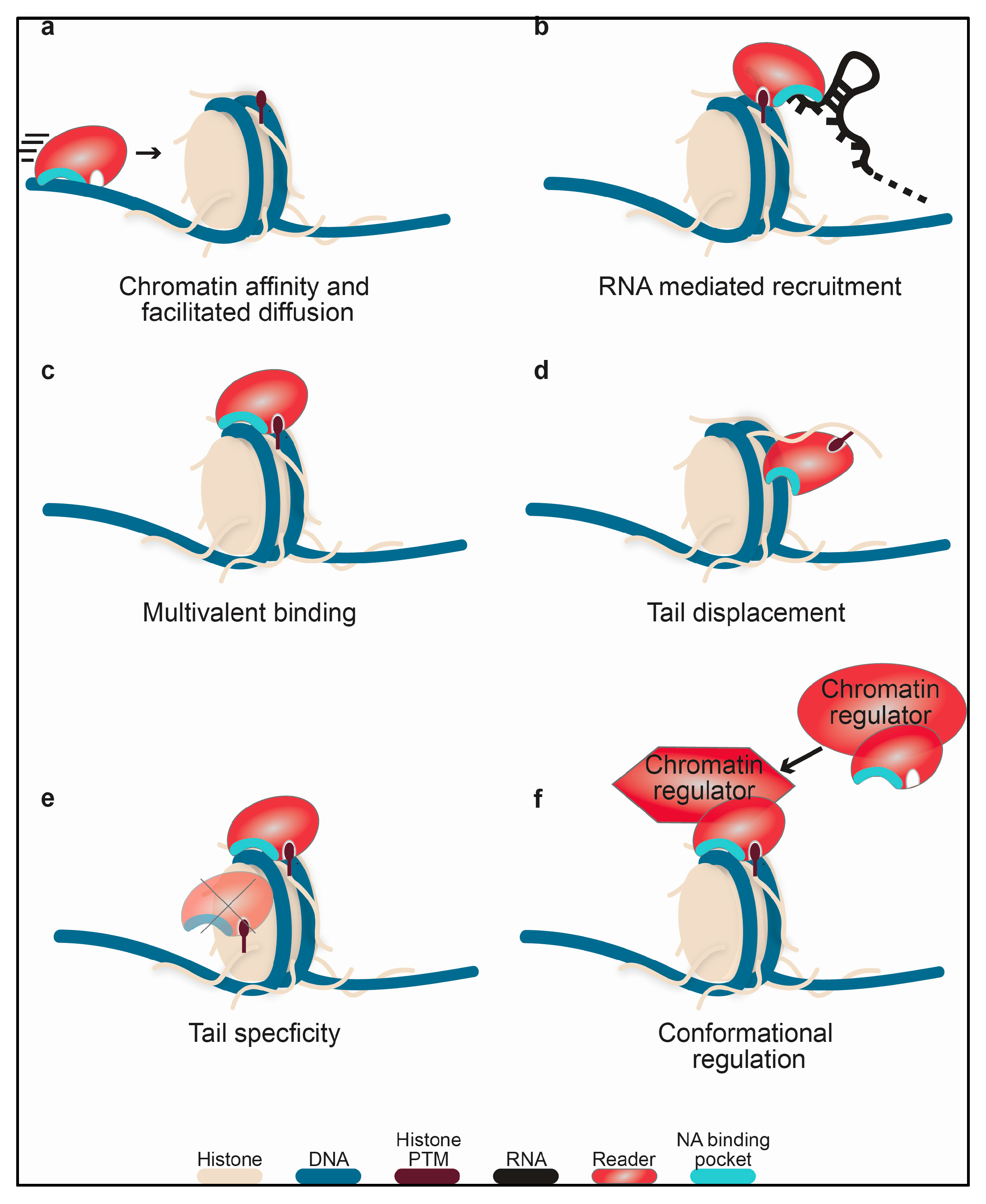
1.2. General Mechanisms of Nucleic Acid Binding
2. Overview and Importance in Function
2.1. Prevalence of Nucleic Acid Binding among Histone Readers and Interplay with Histone Binding
2.2. Comparison to Previously Characterized Nucleic Acid-Binding Domains
2.3. Functional Implications
3. Nucleic Acid-Binding Reader Domains
3.1. Bromodomains
3.2. PHD Fingers
3.3. PWWP Domains
3.4. Chromodomains
3.5. Tudor Domains
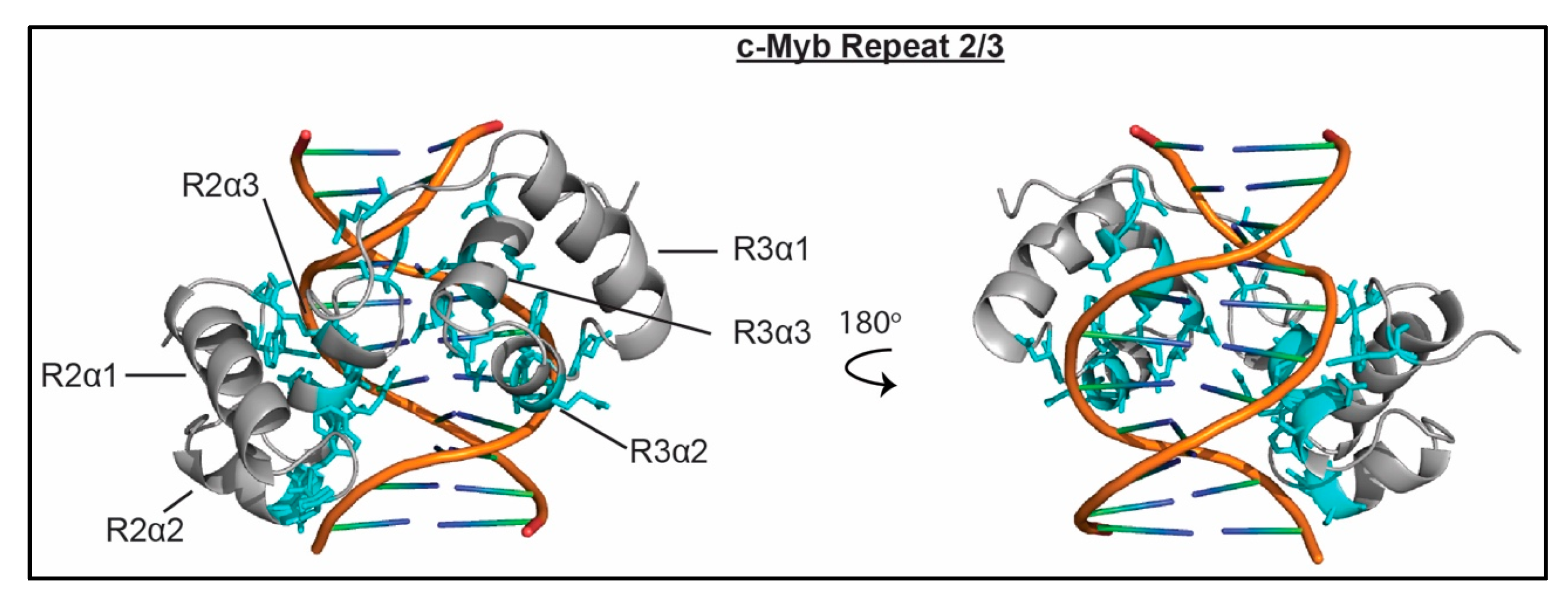
3.6. SANT Domains
3.7. Integrated Domains
4. Auxiliary Nucleic Acid Binding Domains
5. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Allfrey, V.G.; Faulkner, R.; Mirsky, A.E. Acetylation and methylation of histones and their possible role in the regulation of RNA synthesis. Proc. Natl. Acad. Sci. USA 1964, 51, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Winston, F.; Allis, C.D. The bromodomain: A chromatin-targeting module? Nat. Struct. Biol. 1999, 6, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Dhalluin, C.; Carlson, J.E.; Zeng, L.; He, C.; Aggarwal, A.K. Structure and ligand of a histone acetyltransferase bromodomain. Nature 1999, 399, 491–496. [Google Scholar] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Musselman, C.A.; Lalonde, M.-E.; Côté, J.; Kutateladze, T.G. Perceiving the epigenetic landscape through histone readers. Nat. Struct. Mol. Biol. 2012, 19, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.J.; Wang, Z. Readout of Epigenetic Modifications. Annu. Rev. Biochem. 2013, 82, 81–118. [Google Scholar] [CrossRef] [PubMed]
- Rothbart, S.B.; Strahl, B.D. Interpreting the language of histone and DNA modifications. Biochim. Biophys. Acta 2014, 1839, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Andrews, F.H.; Strahl, B.D.; Kutateladze, T.G. Insights into newly discovered marks and readers of epigenetic information. Nat. Chem. Biol. 2016, 12, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Filippakopoulos, P.; Knapp, S. The bromodomain interaction module. FEBS Lett. 2012, 586, 2692–2704. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Zhou, M.-M. Structure and mechanisms of lysine methylation recognition by the chromodomain in gene transcription. Biochemistry 2011, 50, 1966–1980. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Min, J. Structure and function of the nucleosome-binding PWWP domain. Trends Biochem. Sci. 2014, 39, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Rohs, R.; Jin, X.; West, S.M.; Joshi, R.; Honig, B.; Mann, R.S. Origins of Specificity in Protein-DNA Recognition. Annu. Rev. Biochem. 2010, 79, 233–269. [Google Scholar] [CrossRef] [PubMed]
- Dickey, T.H.; Altschuler, S.E.; Wuttke, D.S. Single-Stranded DNA-Binding Proteins: Multiple Domains for Multiple Functions. Structure 2013, 21, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Helder, S.; Blythe, A.J.; Bond, C.S.; Mackay, J.P. Determinants of affinity and specificity in RNA-binding proteins. Curr. Opin. Struct. Biol. 2016, 38, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Privalov, P.L.; Dragan, A.I.; Crane-Robinson, C.; Breslauer, K.J.; Remeta, D.P.; Minetti, C.A. What Drives Proteins into the Major or Minor Grooves of DNA? J. Mol. Biol. 2007, 365, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Geertz, M.; Shore, D.; Maerkl, S.J. Massively parallel measurements of molecular interaction kinetics on a microfluidic platform. Proc. Nat. Acad. Sci. USA 2012, 109, 16540–16545. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.C.; Simon, B.; Rybin, V.; Grötsch, H.; Curtet, S.; Khochbin, S.; Carlomagno, T.; Müller, C.W. A bromodomain-DNA interaction facilitates acetylation-dependent bivalent nucleosome recognition by the BET protein BRDT. Nat. Commun. 2016, 7, 13855. [Google Scholar] [CrossRef] [PubMed]
- Kaustov, L.; Ouyang, H.; Amaya, M.; Lemak, A.; Nady, N.; Duan, S.; Wasney, G.A.; Li, Z.; Vedadi, M.; Schapira, M.; et al. Recognition and specificity determinants of the human cbx chromodomains. J. Biol. Chem. 2011, 286, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, E.; Duncan, E.M.; Masui, O.; Gil, J.; Heard, E.; Allis, C.D. Mouse polycomb proteins bind differentially to methylated histone H3 and RNA and are enriched in facultative heterochromatin. Mol. Cell. Biol. 2006, 26, 2560–2569. [Google Scholar] [CrossRef] [PubMed]
- Musselman, C.A.; Gibson, M.D.; Hartwick, E.W.; North, J.A.; Gatchalian, J.; Poirier, M.G.; Kutateladze, T.G. Binding of PHF1 Tudor to H3K36me3 enhances nucleosome accessibility. Nat. Commun. 2013, 4, 2969. [Google Scholar] [CrossRef] [PubMed]
- Charier, G.; Couprie, J.; Alpha-Bazin, B.; Meyer, V.; Quéméneur, E.; Guérois, R.; Callebaut, I.; Gilquin, B.; Zinn-Justin, S. The Tudor tandem of 53BP1: A new structural motif involved in DNA and RG-rich peptide binding. Structure 2004, 12, 1551–1562. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, K.; Basu, B.P.; Kysela, B.; Kurihara, T.; Shibata, M.; Guan, D.; Cao, Y.; Hamada, T.; Imamura, K.; Jeggo, P.A.; et al. Potential role for 53BP1 in DNA end-joining repair through direct interaction with DNA. J. Biol. Chem. 2003, 278, 36487–36495. [Google Scholar] [CrossRef] [PubMed]
- Morrison, E.A.; Sanchez, J.C.; Ronan, J.L.; Farrell, D.P.; Varzavand, K.; Johnson, J.K.; Gu, B.X.; Crabtree, G.R.; Musselman, C.A. DNA binding drives the association of BRG1/hBRM bromodomains with nucleosomes. Nat. Commun. 2017, 8, 16080. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Popowicz, G.M.; Krajewski, M.; Holak, T.A. Structural Ramification for Acetyl-Lysine Recognition by the Bromodomain of Human BRG1 Protein, a Central ATPase of the SWI/SNF Remodeling Complex. ChemBioChem 2007, 8, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Xu, C.; Huang, W.; Zhang, J.; Carlson, J.E.; Tu, X.; Wu, J.; Shi, Y. Solution Structure of Human Brg1 Bromodomain and Its Specific Binding to Acetylated Histone Tails. Biochemistry 2007, 46, 2100–2110. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Qin, S.; Zhang, J.; Ji, P.; Shi, Y.; Wu, J. Solution structure of an atypical PHD finger in BRPF2 and its interaction with DNA. J. Struct. Biol. 2012, 180, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, F.; Ruan, K.; Zhang, J.; Mei, Y.; Wu, J.; Shi, Y. Structural and Functional Insights into the Human Borjeson-Forssman-Lehmann Syndrome-associated Protein PHF6. J. Biol. Chem. 2014, 289, 10069–10083. [Google Scholar] [CrossRef] [PubMed]
- Dhayalan, A.; Rajavelu, A.; Rathert, P.; Tamas, R.; Jurkowska, R.Z.; Ragozin, S.; Jeltsch, A. The Dnmt3a PWWP domain reads histone 3 lysine 36 trimethylation and guides DNA methylation. J. Biol. Chem. 2010, 285, 26114–26120. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, W.; Zhao, C.; Wang, Y.; Wang, W.; Zhang, J.; Zhang, Z.; Li, G.; Shi, Y.; Tu, X.; et al. Solution structure of the Pdp1 PWWP domain reveals its unique binding sites for methylated H4K20 and DNA. Biochem. J. 2012, 442, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Lukasik, S.M.; Cierpicki, T.; Borloz, M.; Grembecka, J.; Everett, A.; Bushweller, J.H. High resolution structure of the HDGF PWWP domain: A potential DNA binding domain. Protein Sci. 2006, 15, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Everett, A.D. Hepatoma-derived growth factor binds DNA through the N-terminal PWWP domain. BMC Mol. Biol. 2007, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Mao, G.; Tong, D.; Huang, J.; Gu, L.; Yang, W.; Li, G.M. The histone mark H3K36me3 regulates human DNA mismatch repair through its interaction with MutSα. Cell 2013, 153, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Laguri, C.; Duband-Goulet, I.; Friedrich, N.; Axt, M.; Belin, P.; Callebaut, I.; Gilquin, B.; Zinn-Justin, S.; Couprie, J. Human mismatch repair protein MSH6 contains a PWWP domain that targets double stranded DNA. Biochemistry 2008, 47, 6199–6207. [Google Scholar] [CrossRef] [PubMed]
- Van Nuland, R.; van Schaik, F.M.; Simonis, M.; van Heesch, S.; Cuppen, E.; Boelens, R.; Timmers, H.M.; van Ingen, H. Nucleosomal DNA binding drives the recognition of H3K36-methylated nucleosomes by the PSIP1-PWWP domain. Epigenetics Chromatin 2013, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Eidahl, J.O.; Crowe, B.L.; North, J.A.; McKee, C.J.; Shkriabai, N.; Feng, L.; Plumb, M.; Graham, R.L.; Gorelick, R.J.; Hess, S.; et al. Structural basis for high-affinity binding of LEDGF PWWP to mononucleosomes. Nucleic Acids Res. 2013, 41, 3924–3936. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.R.; Nietlispach, D.; Buscaino, A.; Warner, R.J.; Akhtar, A.; Murzin, A.G.; Murzina, N.V.; Laue, E.D. Structure of the chromo barrel domain from the MOF acetyltransferase. J. Biol. Chem. 2005, 280, 32326–32331. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Blus, B.J.; Chandra, V.; Huang, P.; Rastinejad, F.; Khorasanizadeh, S. Corecognition of DNA and a methylated histone tail by the MSL3 chromodomain. Nat. Struct. Mol. Biol. 2010, 17, 1027–1029. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.A.; Ferhatoglu, Y.; Jia, Y.; Al-Jiab, R.A.; Scott, M.J. Structural and biochemical studies on the chromo-barrel domain of male specific lethal 3 (MSL3) reveal a binding preference for mono- or dimethyllysine 20 on histone H4. J. Biol. Chem. 2010, 285, 40879–40890. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Machida, S.; Kurumizaka, H.; Tagami, H.; Nakayama, J.-I. Phosphorylation of CBX2 controls its nucleosome-binding specificity. J. Biochem. 2017, 162, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.L.; Li, S.; Muñoz-Cabello, A.M.; Raguz, S.; Zeng, L.; Mujtaba, S.; Gil, J.; Walsh, M.J.; Zhou, M.M. Molecular Interplay of the Noncoding RNA ANRIL and Methylated Histone H3 Lysine 27 by Polycomb CBX7 in Transcriptional Silencing of INK4a. Mol. Cell 2010, 38, 662–674. [Google Scholar] [CrossRef] [PubMed]
- Zhen, C.Y.; Tatavosian, R.; Huynh, T.N.; Duc, H.N.; Das, R.; Kokotovic, M.; Grimm, J.B.; Lavis, L.D.; Lee, J.; Mejia, F.J.; et al. Live-cell single-molecule tracking reveals co-recognition of H3K27me3 and DNA targets polycomb Cbx7-PRC1 to chromatin. Elife 2016, 5, e17667. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, C.; Liu, Y.; Fan, K.; Li, Z.; Sun, X.; Ouyang, H.; Zhang, X.; Zhang, J.; Li, Y.; et al. Crystal structure of the human SUV39H1 chromodomain and its recognition of histone H3K9me2/3. PLoS ONE 2012, 7, e52977. [Google Scholar] [CrossRef] [PubMed]
- Shirai, A.; Kawaguchi, T.; Shimojo, H.; Muramatsu, D.; Ishida-Yonetani, M.; Nishimura, Y.; Kimura, H.; Nakayama, J.I.; Shinkai, Y. Impact of nucleic acid and methylated H3K9 binding activities of Suv39h1 on its heterochromatin assembly. Elife 2017, 6, e25317. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.L.; Yewdell, W.T.; Bell, J.C.; McNulty, S.M.; Duda, Z.; O′Neill, R.J.; Sullivan, B.A.; Straight, A.F. RNA-dependent stabilization of SUV39H1 at constitutive heterochromatin. Elife 2017, 6, e25299. [Google Scholar] [CrossRef] [PubMed]
- Schalch, T.; Job, G.; Noffsinger, V.J.; Shanker, S.; Kuscu, C.; Joshua-Tor, L.; Partridge, J.F. High-affinity binding of Chp1 chromodomain to K9 methylated histone H3 is required to establish centromeric heterochromatin. Mol. Cell 2009, 34, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M.; Shimojo, H.; Hayashi, A.; Kawaguchi, R.; Ohtani, Y.; Uegaki, K.; Nishimura, Y.; Nakayama, J.I. Intrinsic nucleic acid-binding activity of Chp1 chromodomain is required for heterochromatic gene silencing. Mol. Cell 2012, 47, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Bouazoune, K.; Mitterweger, A.; Längst, G.; Imhof, A.; Akhtar, A.; Becker, P.B.; Brehm, A. The dMi-2 chromodomains are DNA binding modules important for ATP-dependent nucleosome mobilization. EMBO J. 2002, 21, 2430–2440. [Google Scholar] [CrossRef] [PubMed]
- Musselman, C.A.; Avvakumov, N.; Watanabe, R.; Abraham, C.G.; Lalonde, M.E.; Hong, Z.; Allen, C.; Roy, S.; Nuñez, J.K.; Nickoloff, J.; et al. Molecular basis for H3K36me3 recognition by the Tudor domain of PHF1. Nat. Struct. Mol. Biol. 2012, 19, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Botuyan, M.V.; Lee, J.; Ward, I.M.; Kim, J.E.; Thompson, J.R.; Chen, J.; Mer, G. Structural basis for the methylation state-specific recognition of histone H4-K20 by 53BP1 and Crb2 in DNA repair. Cell 2006, 127, 1361–1373. [Google Scholar] [CrossRef] [PubMed]
- Shimojo, H.; Sano, N.; Moriwaki, Y.; Okuda, M.; Horikoshi, M.; Nishimura, Y. Novel structural and functional mode of a knot essential for RNA binding activity of the Esa1 presumed chromodomain. J. Mol. Biol. 2008, 378, 987–1001. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, J.; Yasukawa, T.; Enari, M.; Ogata, K.; Nishimura, Y.; Ishii, S.; Sarai, A. Recognition of specific DNA sequences by the c-myb protooncogene product: Role of three repeat units in the DNA-binding domain. Proc. Natl. Acad. Sci. USA 1993, 90, 9320–9324. [Google Scholar] [CrossRef] [PubMed]
- Mo, X.; Kowenz-Leutz, E.; Laumonnier, Y.; Xu, H.; Leutz, A. Histone H3 tail positioning and acetylation by the c-Myb but not the v-Myb DNA-binding SANT domain. Genes Dev. 2005, 19, 2447–2457. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.R.; Ko, D.; Chen, C.; Lipsick, J.S. A conserved acidic patch in the Myb domain is required for activation of an endogenous target gene and for chromatin binding. Mol. Cancer 2008, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Fuglerud, B.M.; Lemma, R.B.; Wanichawan, P.; Sundaram, A.Y.; Eskeland, R.; Gabrielsen, O.S. A c-Myb mutant causes deregulated differentiation due to impaired histone binding and abrogated pioneer factor function. Nucleic Acids Res. 2017, 45, 7681–7696. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Frouws, T.D.; Angst, B.; Fitzgerald, D.J.; DeLuca, C.; Schimmele, K.; Sargent, D.F.; Richmond, T.J. Structure and mechanism of the chromatin remodelling factor ISW1a. Nature 2011, 472, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Grüne, T.; Brzeski, J.; Eberharter, A.; Clapier, C.R.; Corona, D.F.; Becker, P.B.; Müller, C.W. Crystal structure and functional analysis of a nucleosome recognition module of the remodeling factor ISWI. Mol. Cell 2003, 12, 449–460. [Google Scholar] [CrossRef]
- Stokes, D.G.; Perry, R.P. DNA-binding and chromatin localization properties of CHD1. Mol. Cell. Biol. 1995, 15, 2745–2753. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Jenkins, K.R.; Heroux, A.; Bowman, G.D. Crystal structure of the chromodomain helicase DNA-binding protein 1 (Chd1) DNA-binding domain in complex with DNA. J. Biol. Chem. 2011, 286, 42099–42104. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Sundaramoorthy, R.; Martin, D.; Singh, V.; Owen-Hughes, T. The DNA-binding domain of the Chd1 chromatin-remodelling enzyme contains SANT and SLIDE domains. EMBO J. 2011, 30, 2596–2609. [Google Scholar] [CrossRef] [PubMed]
- Pilotto, S.; Speranzini, V.; Tortorici, M.; Durand, D.; Fish, A.; Valente, S.; Forneris, F.; Mai, A.; Sixma, T.K.; Vachette, P.; et al. Interplay among nucleosomal DNA, histone tails, and corepressor CoREST underlies LSD1-mediated H3 demethylation. Proc. Natl. Acad. Sci. USA 2015, 112, 2752–2757. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Gocke, C.B.; Luo, X.; Borek, D.; Tomchick, D.R.; Machius, M.; Otwinowski, Z.; Yu, H. Structural basis for CoREST-dependent demethylation of nucleosomes by the human LSD1 histone demethylase. Mol. Cell 2006, 23, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Savitsky, P.; Krojer, T.; Fujisawa, T.; Lambert, J.P.; Picaud, S.; Wang, C.Y.; Shanle, E.K.; Krajewski, K.; Friedrichsen, H.; Kanapin, A.; et al. Multivalent Histone and DNA Engagement by a PHD/BRD/PWWP Triple Reader Cassette Recruits ZMYND8 to K14ac-Rich Chromatin. Cell Rep. 2016, 17, 2724–2737. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Morikawa, S.; Nakamura, H.; Sekikawa, A.; Inoue, T.; Kanai, H.; Sarai, A.; Ishii, S.; Nishimura, Y. Solution structure of a specific DNA complex of the Myb DNA-binding domain with cooperative recognition helices. Cell 1994, 79, 639–648. [Google Scholar] [CrossRef]
- Tahirov, T.H.; Sato, K.; Ichikawa-Iwata, E.; Sasaki, M.; Inoue-Bungo, T.; Shiina, M.; Kimura, K.; Takata, S.; Fujikawa, A.; Morii, H.; et al. Mechanism of c-Myb-C/EBP beta cooperation from separated sites on a promoter. Cell 2002, 108, 57–70. [Google Scholar] [CrossRef]
- Biedenkapp, H.; Borgmeyer, U.; Sippel, A.E.; Klempnauer, K.H. Viral myb oncogene encodes a sequence-specific DNA-binding activity. Nature 1988, 335, 835–837. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.M.; Watson, R.J. Nucleotide preferences in sequence-specific recognition of DNA by c-myb protein. Nucleic Acids Res. 1991, 19, 3913–3919. [Google Scholar] [CrossRef] [PubMed]
- Weston, K. Extension of the DNA binding consensus of the chicken c-Myb and v-Myb proteins. Nucleic Acids Res. 1992, 20, 3043–3049. [Google Scholar] [CrossRef] [PubMed]
- Van Royen, M.E.; Zotter, A.; Ibrahim, S.M.; Geverts, B.; Houtsmuller, A.B. Nuclear proteins: Finding and binding target sites in chromatin. Chromosome Res. 2011, 19, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qin, S.; Li, F.; Li, S.; Zhang, W.; Peng, J.; Zhang, Z.; Gong, Q.; Wu, J.; Shi, Y. Crystal structure of human BS69 Bromo-ZnF-PWWP reveals its role in H3K36me3 nucleosome binding. Cell Res. 2014, 24, 890–893. [Google Scholar] [CrossRef] [PubMed]
- Cutter, A.R.; Hayes, J.J. A brief review of nucleosome structure. FEBS Lett. 2015, 589, 2914–2922. [Google Scholar] [CrossRef] [PubMed]
- Morrison, E.A.; Bowerman, S.; Sylvers, K.L.; Wereszczynski, J.; Musselman, C.A. The conformation of the histone H3 tail inhibits association of the BPTF PHD finger with the nucleosome. Elife 2018, 7, e31481. [Google Scholar] [CrossRef] [PubMed]
- Gatchalian, J.; Wang, X.; Ikebe, J.; Cox, K.L.; Tencer, A.H.; Zhang, Y.; Burge, N.L.; Di, L.; Gibson, M.D.; Musselman, C.A.; et al. Accessibility of the histone H3 tail in the nucleosome for binding of paired readers. Nat. Commun. 2017, 8, 1489. [Google Scholar] [CrossRef] [PubMed]
- Stützer, A.; Liokatis, S.; Kiesel, A.; Schwarzer, D.; Sprangers, R.; Söding, J.; Selenko, P.; Fischle, W. Modulations of DNA Contacts by Linker Histones and Post-translational Modifications Determine the Mobility and Modifiability of Nucleosomal H3 Tails. Mol. Cell 2016, 61, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Schall, K.; Kallis, E.; Eustermann, S.; Guariento, M.; Moldt, M.; Hopfner, K.P.; Michaelis, J. Single-molecule nucleosome remodeling by INO80 and effects of histone tails. FEBS Lett. 2018, 592, 318–331. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, R.H.; Ladurner, A.G.; King, D.S.; Tjian, R. Structure and function of a human TAFII250 double bromodomain module. Science 2000, 288, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Rahnamoun, H.; Lee, J.; Sun, Z.; Lu, H.; Ramsey, K.M.; Komives, E.A.; Lauberth, S.M. RNAs interact with BRD4 to promote enhanced chromatin engagement and transcription activation. Nat. Struct. Mol. Biol. 2018, 25, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.; Zhou, M.-M. The PHD finger: A versatile epigenome reader. Trends Biochem. Sci. 2011, 36, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Sawada, K.; Zhang, X.; Cheng, X. The PWWP domain of mammalian DNA methyltransferase Dnmt3b defines a new family of DNA-binding folds. Nat. Struct. Biol. 2002, 9, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Nameki, N.; Tochio, N.; Koshiba, S.; Inoue, M.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Fujikura, Y.; Saito, M.; et al. Solution structure of the PWWP domain of the hepatoma-derived growth factor family. Protein Sci. 2005, 14, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Laue, K.; Daujat, S.; Crump, J.G.; Plaster, N.; Roehl, H.H.; Kimmel, C.B.; Schneider, R.; Hammerschmidt, M. The multidomain protein Brpf1 binds histones and is required for Hox gene expression and segmental identity. Development 2008, 135, 1935–1946. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Reddy, B.; Thompson, J.; Wang, H.; Noma, K.I.; Yates, J.R., III; Jia, S. Regulation of Set9-mediated H4K20 methylation by a PWWP domain protein. Mol. Cell 2009, 33, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tsujimoto, N.; Li, E. The PWWP Domain of Dnmt3a and Dnmt3b Is Required for Directing DNA Methylation to the Major Satellite Repeats at Pericentric Heterochromatin. Mol. Cell. Biol. 2004, 24, 9048–9058. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Huang, Y.C.; Huang, S.T.; Hsieh, Y.C.; Guan, H.H.; Chen, N.C.; Chuankhayan, P.; Yoshimura, M.; Tai, M.H.; Chen, C.J. Domain swapping and SMYD1 interactions with the PWWP domain of human hepatoma-derived growth factor. Sci. Rep. 2018, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.; Cherepanov, P. The lentiviral integrase binding protein LEDGF/p75 and HIV-1 replication. PLoS Pathog. 2008, 4, e1000046. [Google Scholar] [CrossRef] [PubMed]
- Shun, M.C.; Botbol, Y.; Li, X.; Di Nunzio, F.; Daigle, J.E.; Yan, N.; Lieberman, J.; Lavigne, M.; Engelman, A. Identification and characterization of PWWP domain residues critical for LEDGF/p75 chromatin binding and human immunodeficiency virus type 1 infectivity. J. Virol. 2008, 82, 11555–11567. [Google Scholar] [CrossRef] [PubMed]
- Blus, B.J.; Wiggins, K.; Khorasanizadeh, S. Epigenetic virtues of chromodomains. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 507–526. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, A.; Zink, D.; Becker, P.B. Chromodomains are protein-RNA interaction modules. Nature 2000, 407, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Zocco, M.; Marasovic, M.; Pisacane, P.; Bilokapic, S.; Halic, M. The Chp1 chromodomain binds the H3K9me tail and the nucleosome core to assemble heterochromatin. Cell Discov. 2016, 2, 16004. [Google Scholar] [CrossRef] [PubMed]
- Meller, V.H.; Rattner, B.P. The roX genes encode redundant male-specific lethal transcripts required for targeting of the MSL complex. EMBO J. 2002, 21, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Buscaino, A.; Köcher, T.; Kind, J.H.; Holz, H.; Taipale, M.; Wagner, K.; Wilm, M.; Akhtar, A. MOF-regulated acetylation of MSL-3 in the Drosophila dosage compensation complex. Mol. Cell 2003, 11, 1265–1277. [Google Scholar] [CrossRef]
- Buscaino, A.; Legube, G.; Akhtar, A. X-chromosome targeting and dosage compensation are mediated by distinct domains in MSL-3. EMBO Rep. 2006, 7, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Sural, T.H.; Peng, S.; Li, B.; Workman, J.L.; Park, P.J.; Kuroda, M.I. The MSL3 chromodomain directs a key targeting step for dosage compensation of the Drosophila melanogaster X chromosome. Nat. Struct. Mol. Biol. 2008, 15, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Kovač, K.; Sauer, A.; Mačinković, I.; Awe, S.; Finkernagel, F.; Hoffmeister, H.; Fuchs, A.; Müller, R.; Rathke, C.; Längst, G.; et al. Tumour-associated missense mutations in the dMi-2 ATPase alters nucleosome remodelling properties in a mutation-specific manner. Nat. Commun. 2018, 9, 2112. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Wang, G.G. Tudor: A versatile family of histone methylation ‘readers’. Trends Biochem. Sci. 2013, 38, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Boyer, L.A.; Latek, R.R.; Peterson, C.L. The SANT domain: A unique histone-tail-binding module? Nat. Rev. Mol. Cell Biol. 2004, 5, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011, 66, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Hanaoka, S.; Nagadoi, A.; Nishimura, Y. Comparison between TRF2 and TRF1 of their telomeric DNA-bound structures and DNA-binding activities. Protein Sci. 2005, 14, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.A.; Wuttke, D.S. Telomerase and telomere-associated proteins: Structural insights into mechanism and evolution. Structure 2012, 20, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Okamura, H.; Nagadoi, A.; König, P.; Rhodes, D.; Nishimura, Y. Solution structure of a telomeric DNA complex of human TRF1. Structure 2001, 9, 1237–1251. [Google Scholar] [CrossRef]
- Prouse, M.B.; Campbell, M.M. The interaction between MYB proteins and their target DNA binding sites. Biochim. Biophys. Acta 2012, 1819, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.L.; Ishii, S.; Sarai, A. Binding site analysis of c-Myb: Screening of potential binding sites by using the mutation matrix derived from systematic binding affinity measurements. Nucleic Acids Res. 1996, 24, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Farnung, L.; Vos, S.M.; Wigge, C.; Cramer, P. Nucleosome–Chd1 structure and implications forchromatin remodelling. Nature 2017, 550, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Li, Y.; Xi, Y.; Jiang, S.; Stratton, S.; Peng, D.; Tanaka, K.; Ren, Y.; Xia, Z.; Wu, J.; et al. ZMYND11 links histone H3.3K36me3 to transcription elongation and tumour suppression. Nature 2014, 508, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Gong, F.; Chiu, L.Y.; Cox, B.; Aymard, F.; Clouaire, T.; Leung, J.W.; Cammarata, M.; Perez, M.; Agarwal, P.; Brodbelt, J.S.; et al. Screen identifies bromodomain protein ZMYND8 in chromatin recognition of transcription-associated DNA damage that promotes homologous recombination. Genes Dev. 2015, 29, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, Y.; Lv, J.; Zheng, X.; Wen, H.; Shen, H.; Zhu, G.; Chen, T.Y.; Dhar, S.S.; Kan, P.Y.; et al. ZMYND8 Reads the Dual Histone Mark H3K4me1-H3K14ac to Antagonize the Expression of Metastasis-Linked Genes. Mol. Cell 2016, 63, 470–484. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Passner, J.M.; Rohs, R.; Jain, R.; Sosinsky, A.; Crickmore, M.A.; Jacob, V.; Aggarwal, A.K.; Honig, B.; Mann, R.S. Functional Specificity of a Hox Protein Mediated by the Recognition of Minor Groove Structure. Cell 2007, 131, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Rohs, R.; West, S.M.; Sosinsky, A.; Liu, P.; Mann, R.S.; Honig, B. The role of DNA shape in protein-DNA recognition. Nature 2009, 461, 1248–1253. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, R.; Mishra, R.K. Novel motifs distinguish multiple homologues of Polycomb in vertebrates: Expansion and diversification of the epigenetic toolkit. BMC Genom. 2009, 10, 549. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Landsman, D. AT-hook motifs identified in a wide variety of DNA-binding proteins. Nucleic Acids Res. 1998, 26, 4413–4421. [Google Scholar] [CrossRef] [PubMed]
- Filarsky, M.; Zillner, K.; Araya, I.; Villar-Garea, A.; Merkl, R.; Längst, G.; Németh, A. The extended AT-hook is a novel RNA binding motif. RNA Biol. 2015, 12, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Turlure, F.; Maertens, G.; Rahman, S.; Cherepanov, P.; Engelman, A. A tripartite DNA-binding element, comprised of the nuclear localization signal and two AT-hook motifs, mediates the association of LEDGF/p75 with chromatin in vivo. Nucleic Acids Res. 2006, 34, 1653–1665. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reader Domain | Protein | Organism | Histone Substrate | ssRNA | dsRNA | ssDNA | dsDNA | Nucleosome |
|---|---|---|---|---|---|---|---|---|
| BD | BRM [23] | H. sapiens | H3K14ac | NT | NT | NT | 600 μM (10 bp) | NT |
| BRG1 [23,24,25] | H. sapiens | H3K14ac | NT | NT | NT | + | NT | |
| BRDT1 BD1 [17] | H. sapiens | H4K5acK8ac | NT | NT | NT | 52 μM (25 bp) | 2 μM | |
| PHD finger | BRPF2 PHD2 [26] | H. sapiens | none | NT | NT | 9 μM (14 nt) | 4 μM (14 bp) | NT |
| PHF6 ePHD2 [27] | H. sapiens | none | NT | NT | NT | 13 μM (14 bp) | NT | |
| PWWP | DNMT3b [28] | H. sapiens | H3K36me3 | NT | NT | NT | 0.23 μM (30 bp) | NT |
| Pdp1 [29] | S. pombe | H4K20me3 | NT | NT | NT | 4.9–5.3 μM (33 bp) | + | |
| HDGF [30,31] | H. sapiens | none | NT | NT | − | + | NT | |
| MSH6 [32,33] | H. sapiens | H3K36me3 | NT | NT | 2.5 μM (35nt) | 0.13 μM (35 bp) | NT | |
| PSIP1/LEDGF [34,35] | H. sapiens | H3K36me3 | NT | NT | NT | 150 μM (10 bp), 1.6 μM (40 bp) | 1.5 μM, 48 nM | |
| CD/ Chromobarrel | MOF [36] | D. melanogaster | none | + | NT | NT | NT | NT |
| MSL-3 [37,38] | D. melanogaster | H4K20me1 | + | + | NT | 0.4 μM (18 bp) | NT | |
| MSL-3 [37,38] | H. sapiens | H4K20me1 | − | − | + | 0.3 μM (18 bp) | NT | |
| CBX2 [18,39] | H. sapiens | H3K27me3 | 3.5 μM (130 nt) | NT | NT | 8.1 μM (130 bp) | NT | |
| CBX4 [19] | M. musculus | H3K9me3 | + | NT | NT | NT | NT | |
| CBX6 [19] | M. musculus | H3K27me3 | + | NT | NT | NT | NT | |
| CBX7 [19] | M. musculus | H3K9me3 | 100 μM (500 nt) | + | NT | + | NT | |
| CBX7 [18,40,41] | H. sapiens | H3K9me3 | 51 μM (13 nt) | NT | NT | − | NT | |
| CBX8 [19] | M. musculus | H3K27me3 | + | NT | NT | NT | NT | |
| SUV39H1 [42,43] | M. musculus | H3K9me3 | 0.35 μM (130 nt) | NT | NT | + | NT | |
| SUV39H1 [42,44] | H. sapiens | H3K9me3 | 2.3 μM (19 nt) | + | + | + | NT | |
| Clr4 [45,46] | S. pombe | H3K9me3 | + | NT | + | + | NT | |
| Chp1 [45,46] | S. pombe | H3K9me3 | 3.9 μM (75 nt) | NT | + | + | NT | |
| MI-2 [47] | D. melanogaster | none | NT | NT | NT | + | + | |
| Tudor | PHF1 [20,48] | H. sapiens | H3K36me3 | NT | NT | − | 201 μM (10 bp) | + |
| 53BP1 [21,49] | M. musculus | H4K20me2 | NT | NT | NT | + | NT | |
| 53BP1 [22,49] | H. sapiens | H4K20me2 | NT | NT | + | + | NT | |
| Esa1 [50] | S. cerevisiae | none | NT | 21–112 μM (varying) | NT | 70–241 μM (varying) | NT | |
| SANT/Myb | c-Myb R2R3 [51,52,53,54] | H. sapiens | unmod H3, unmod H4 | NT | NT | NT | 3.8 nM (16 bp) | + |
| ISW1a [55] | S. cerevisiae | none | NT | NT | NT | + | + | |
| ISWI [56] | D. melanogaster | none | NT | NT | NT | + | + | |
| Chd1 [57,58,59] | S. cerevisiae | none | − | NT | − | + | + | |
| CoRest1 [60,61] | H. sapiens | none | NT | NT | NT | 81 μM (18 bp) | + | |
| Integrated Domains | ZMYND11 | H. sapiens | H3K36me3 | NT | NT | NT | 31 μM (33 bp), 4.6 μM (22 bp) | 0.95 μM (unmod), 0.16 μM (H3K36me3) |
| ZMYND8 [62] | H. sapiens | H3K4me0, H3K14ac, H3K36me0 | NT | NT | NT | + | NT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weaver, T.M.; Morrison, E.A.; Musselman, C.A. Reading More than Histones: The Prevalence of Nucleic Acid Binding among Reader Domains. Molecules 2018, 23, 2614. https://doi.org/10.3390/molecules23102614
Weaver TM, Morrison EA, Musselman CA. Reading More than Histones: The Prevalence of Nucleic Acid Binding among Reader Domains. Molecules. 2018; 23(10):2614. https://doi.org/10.3390/molecules23102614
Chicago/Turabian StyleWeaver, Tyler M., Emma A. Morrison, and Catherine A. Musselman. 2018. "Reading More than Histones: The Prevalence of Nucleic Acid Binding among Reader Domains" Molecules 23, no. 10: 2614. https://doi.org/10.3390/molecules23102614
APA StyleWeaver, T. M., Morrison, E. A., & Musselman, C. A. (2018). Reading More than Histones: The Prevalence of Nucleic Acid Binding among Reader Domains. Molecules, 23(10), 2614. https://doi.org/10.3390/molecules23102614