
Inhibitory Effect of Camptothecin against Rice Bacterial Brown Stripe Pathogen Acidovorax avenae subsp. avenae RS-2


Abstract
:
1. Introduction
2. Results and Discussion
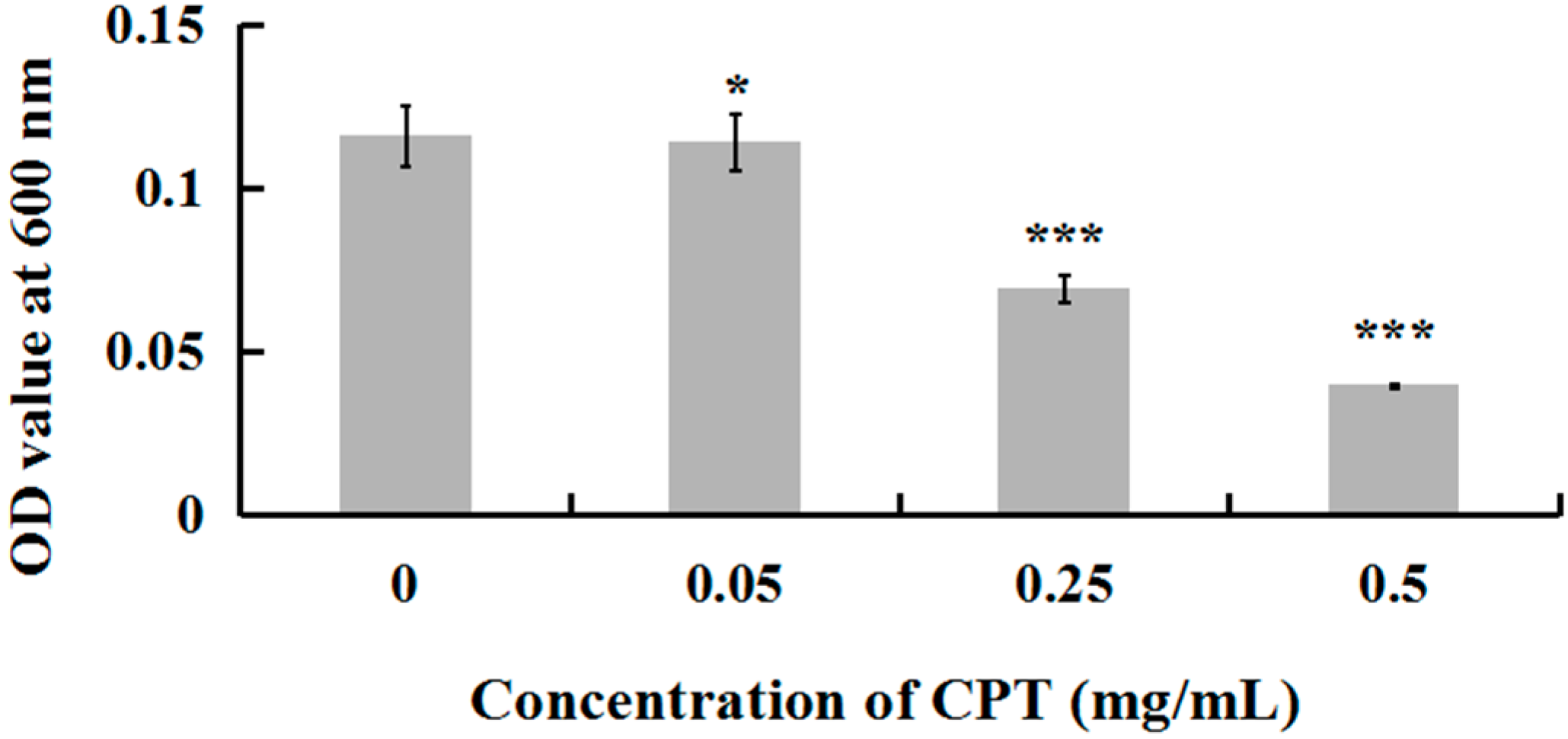
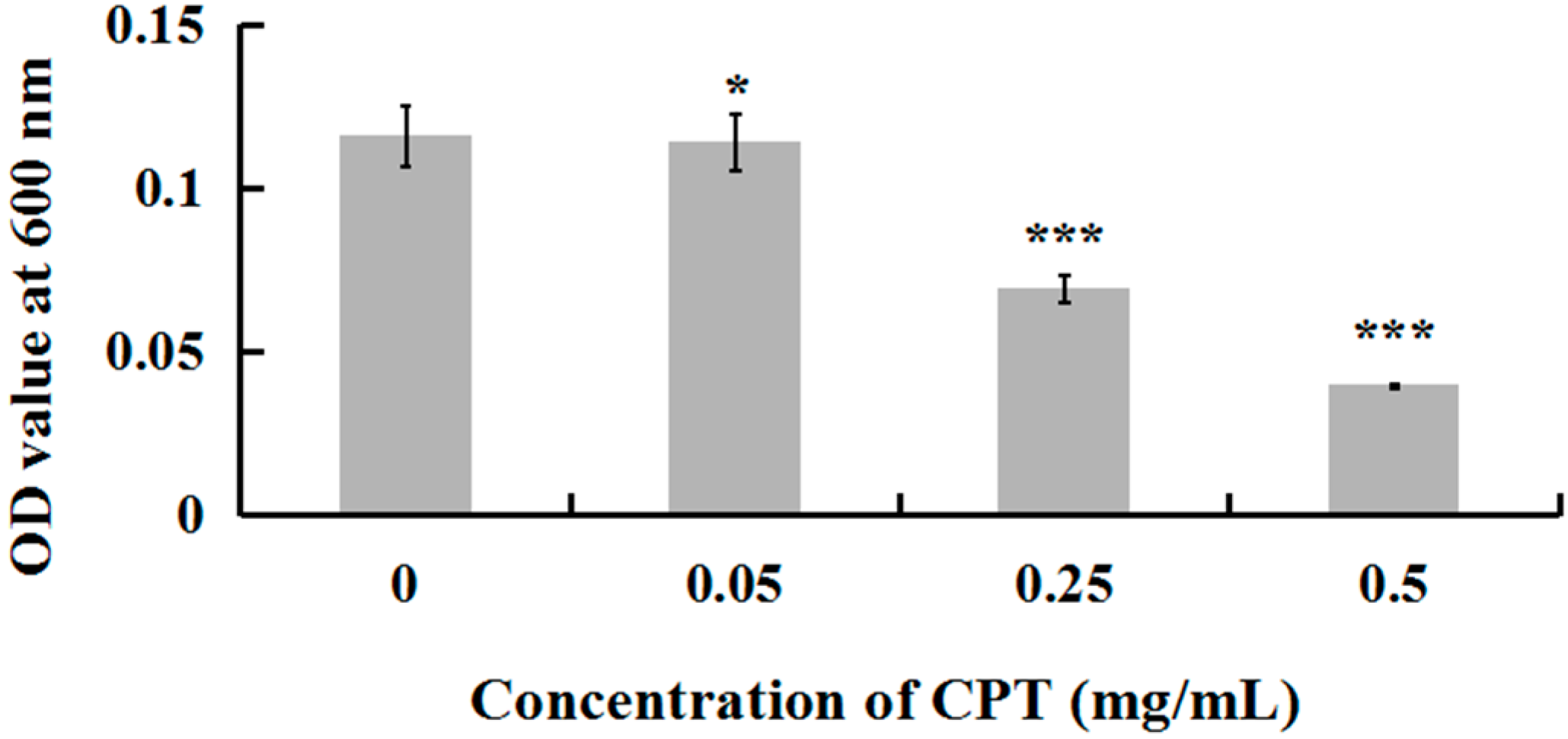
2.1. Antibacterial Activity of CPT
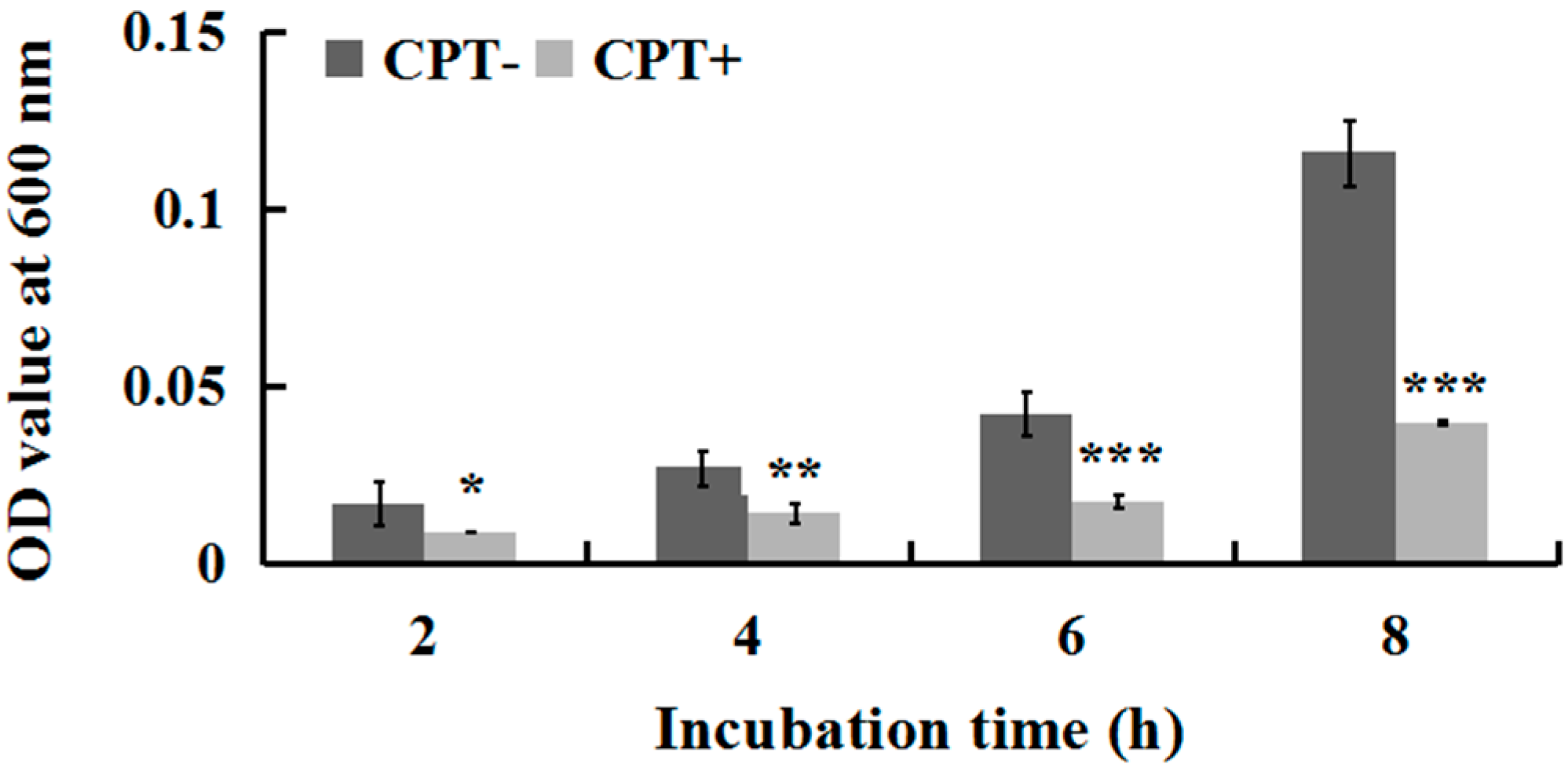
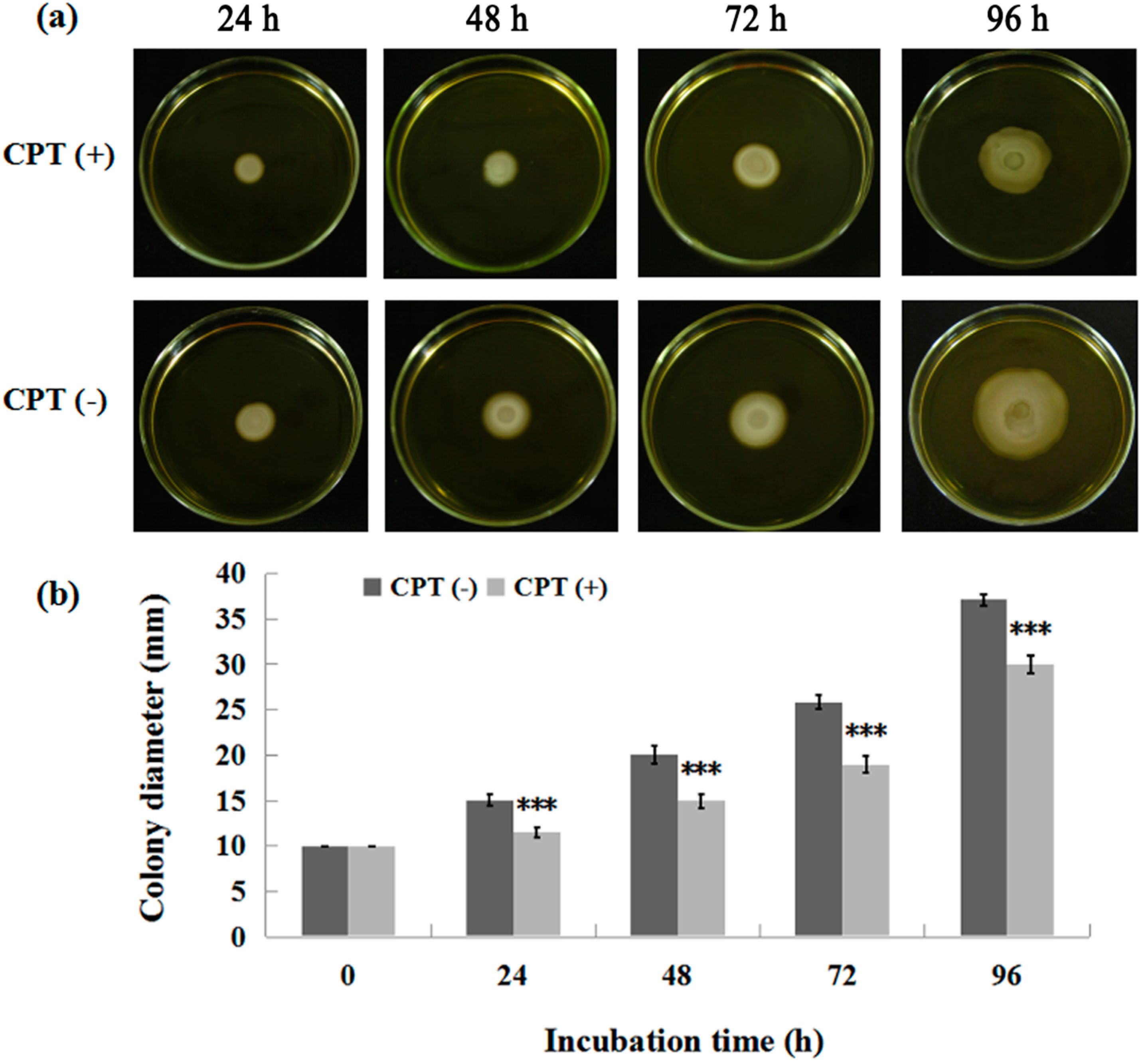
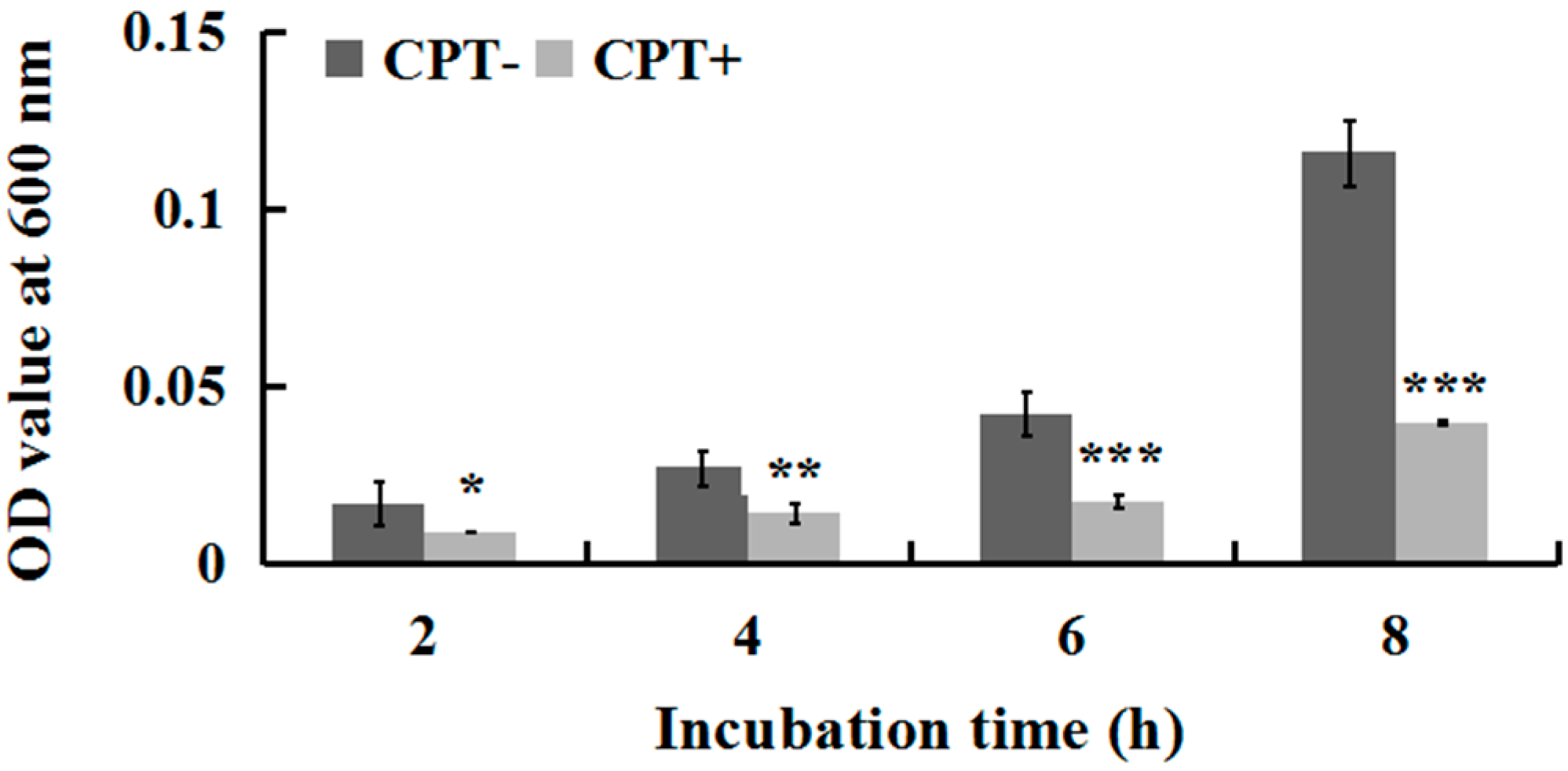
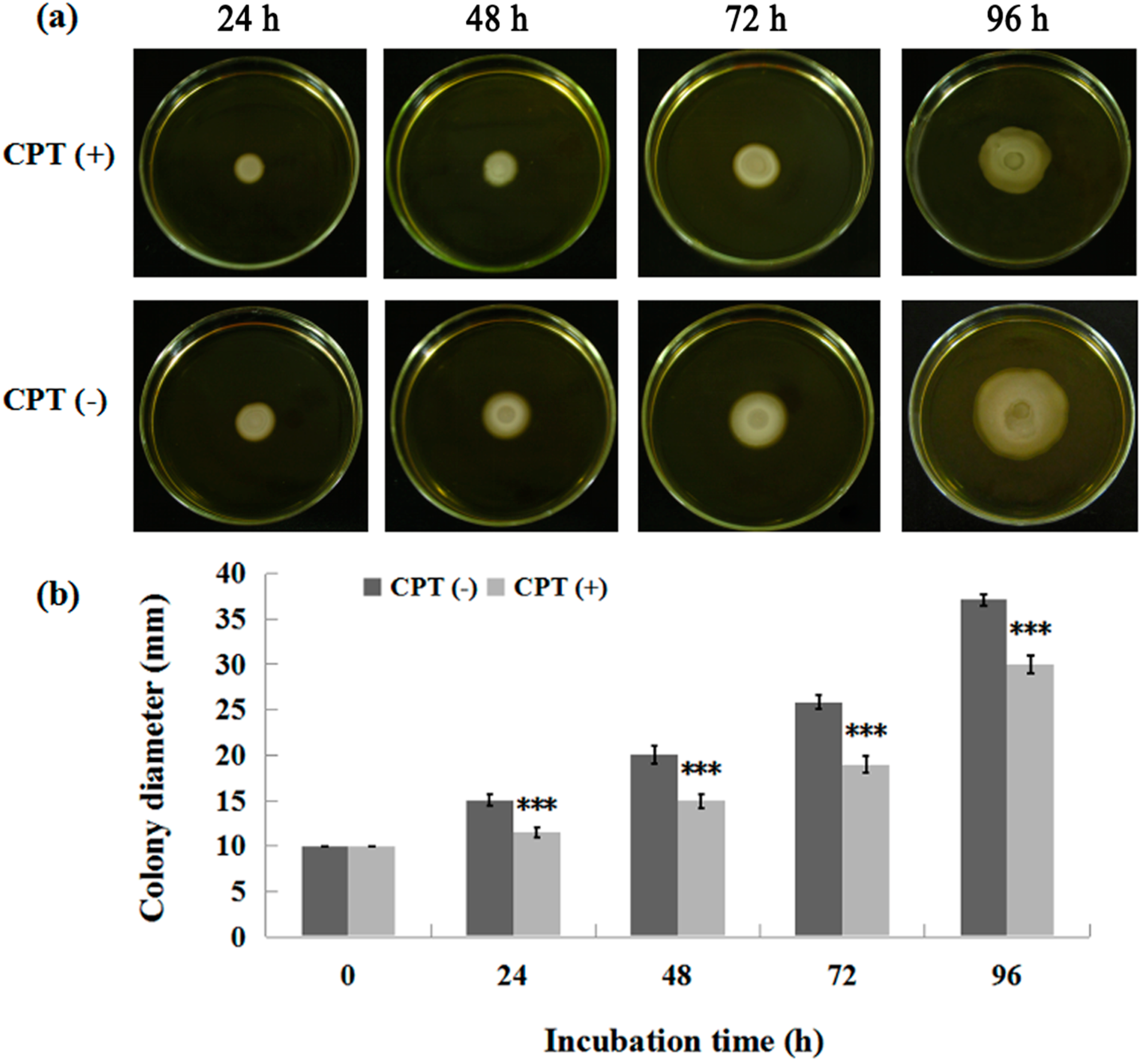
2.2. Effect of Incubation Time on Antibacterial Activity
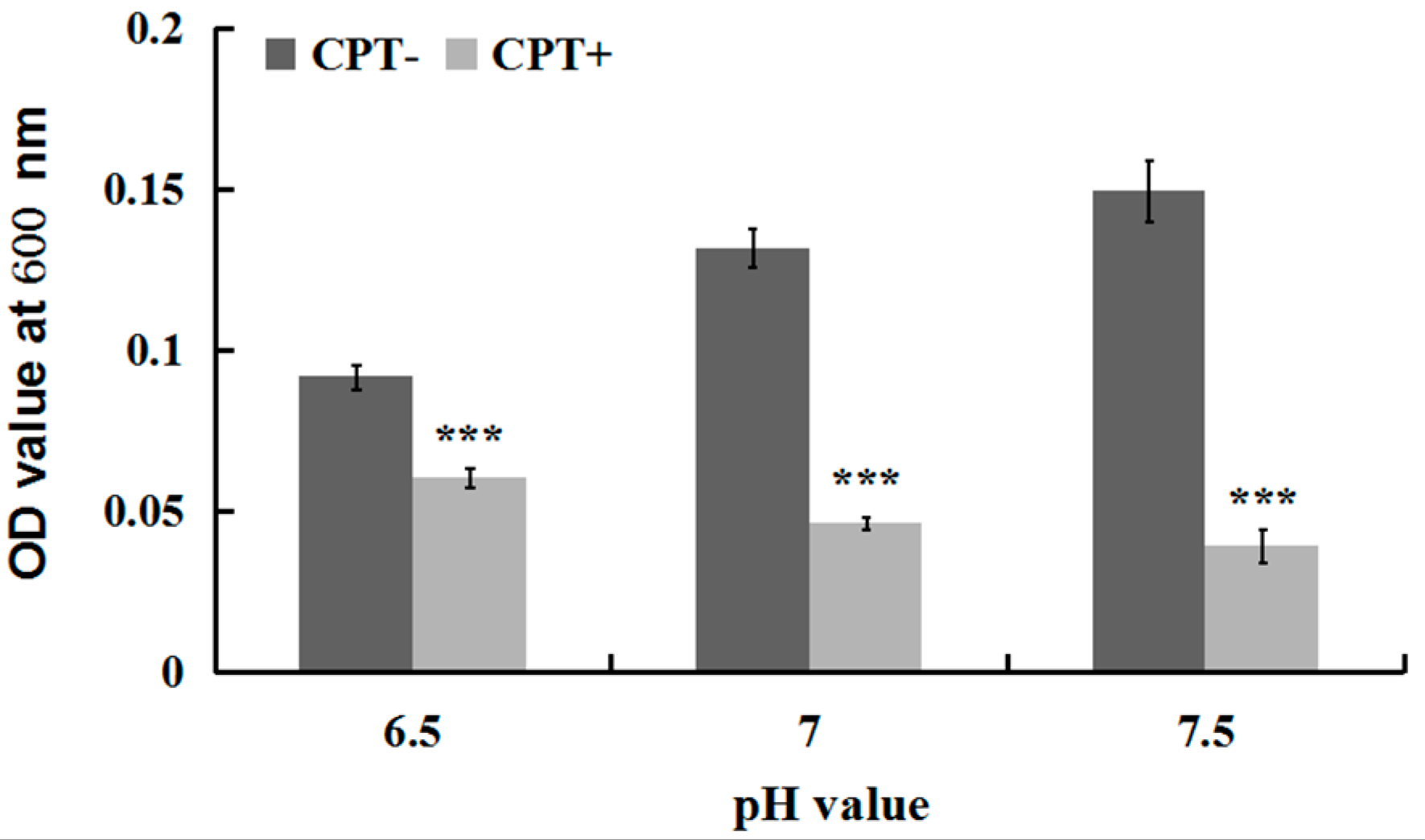
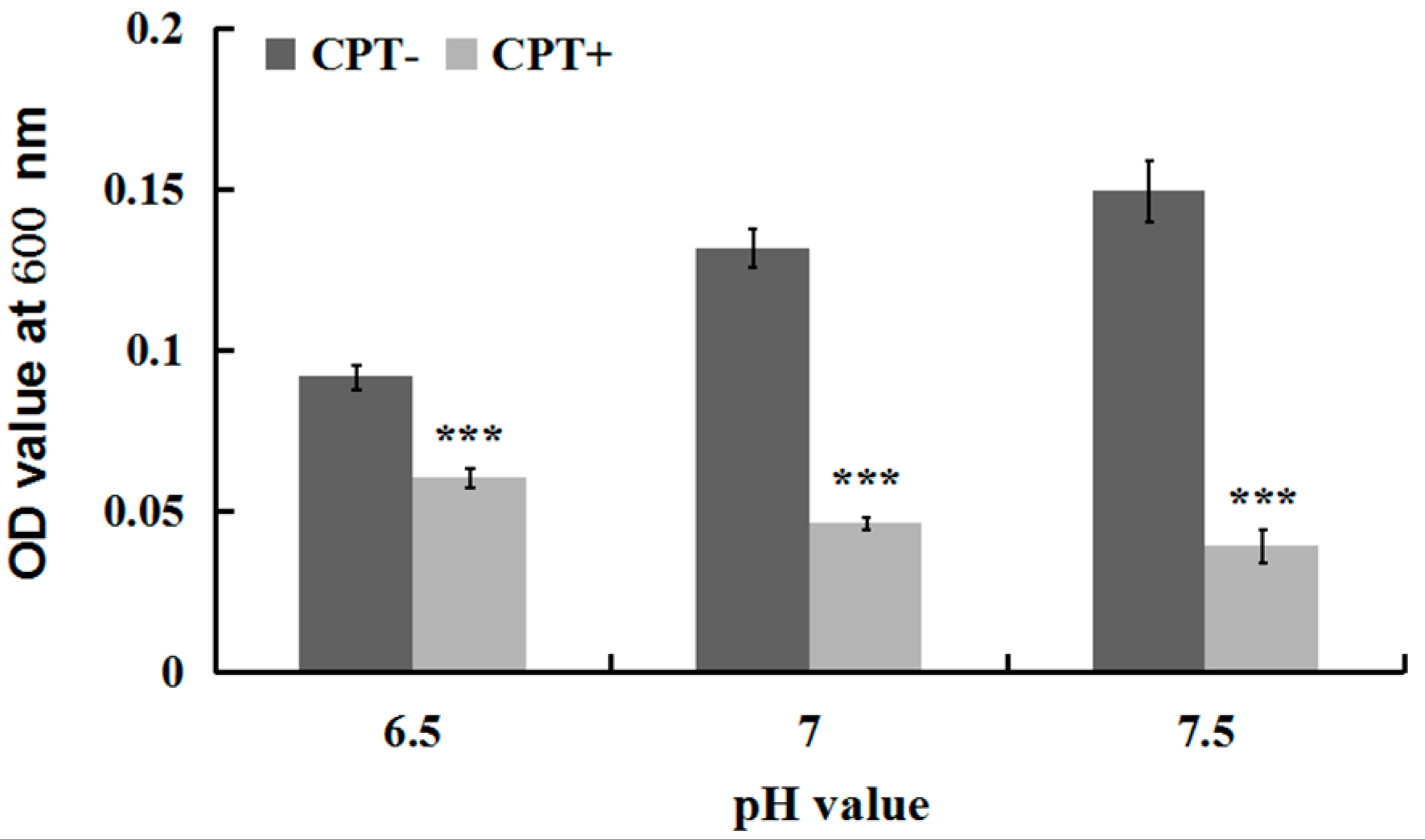
2.3. Effect of the pH Value on Antibacterial Activity
2.4. Swarming Motility
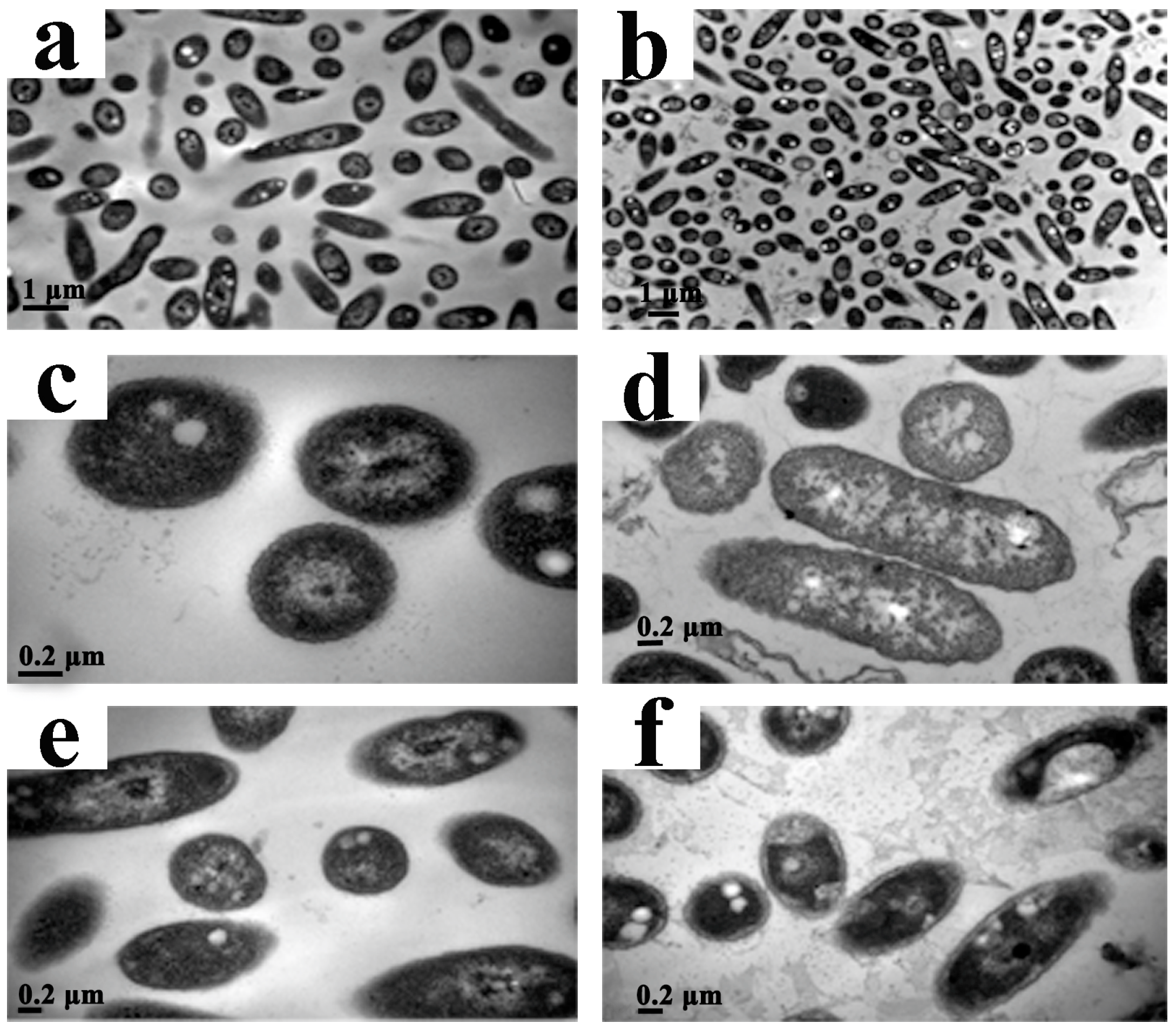
2.5. Live/Dead Cell Staining
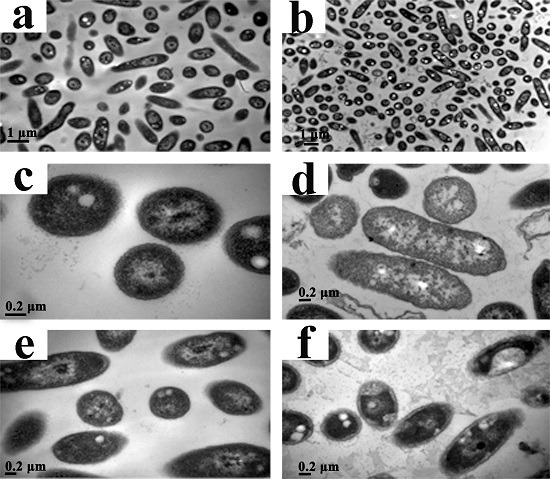
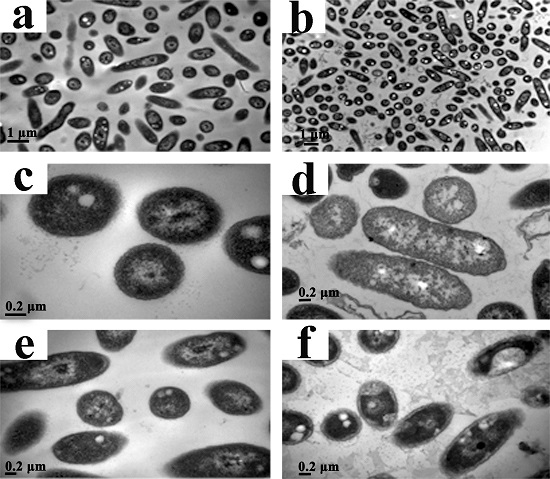
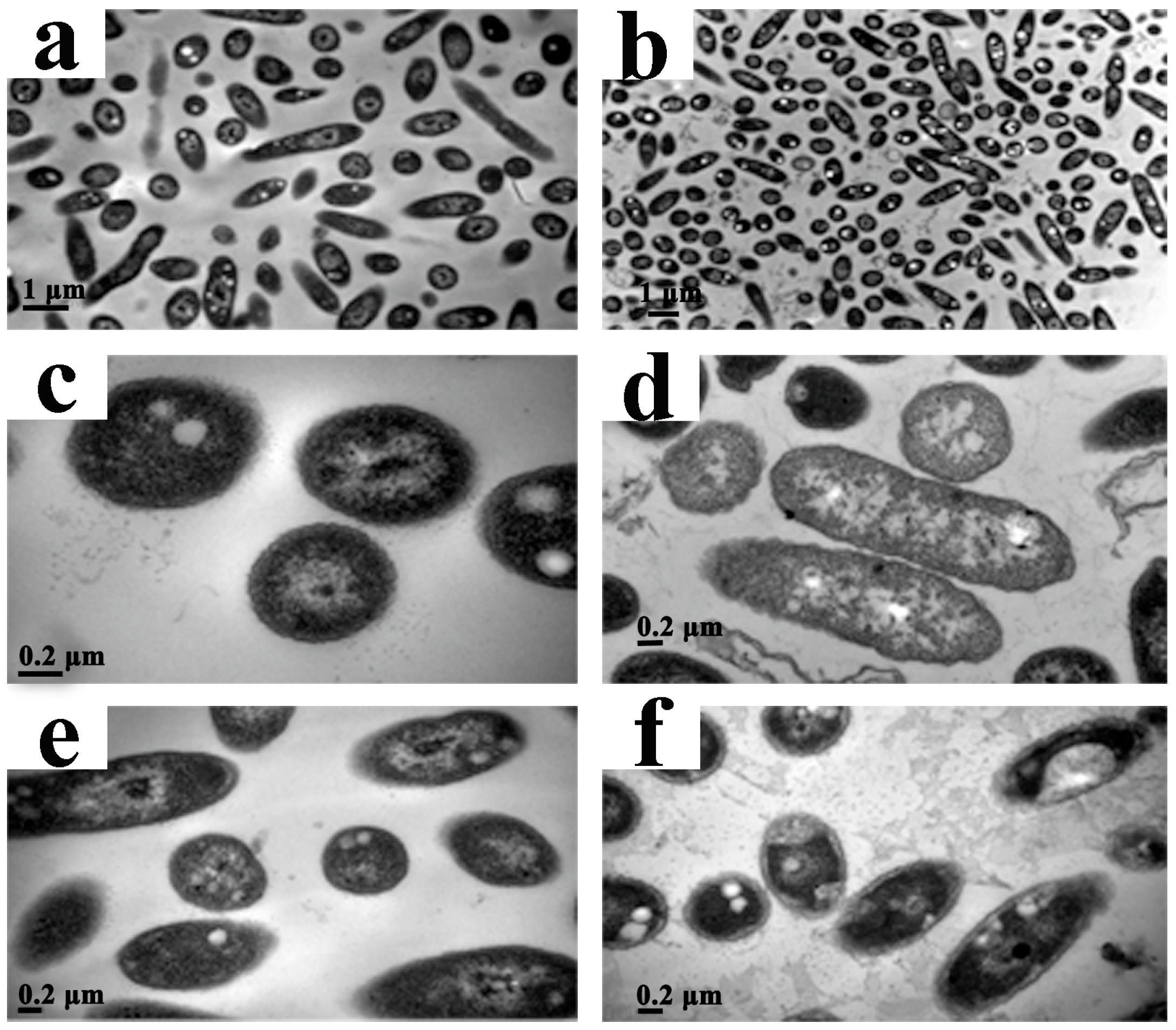
2.6. Transmission Electron Microscopy (TEM)
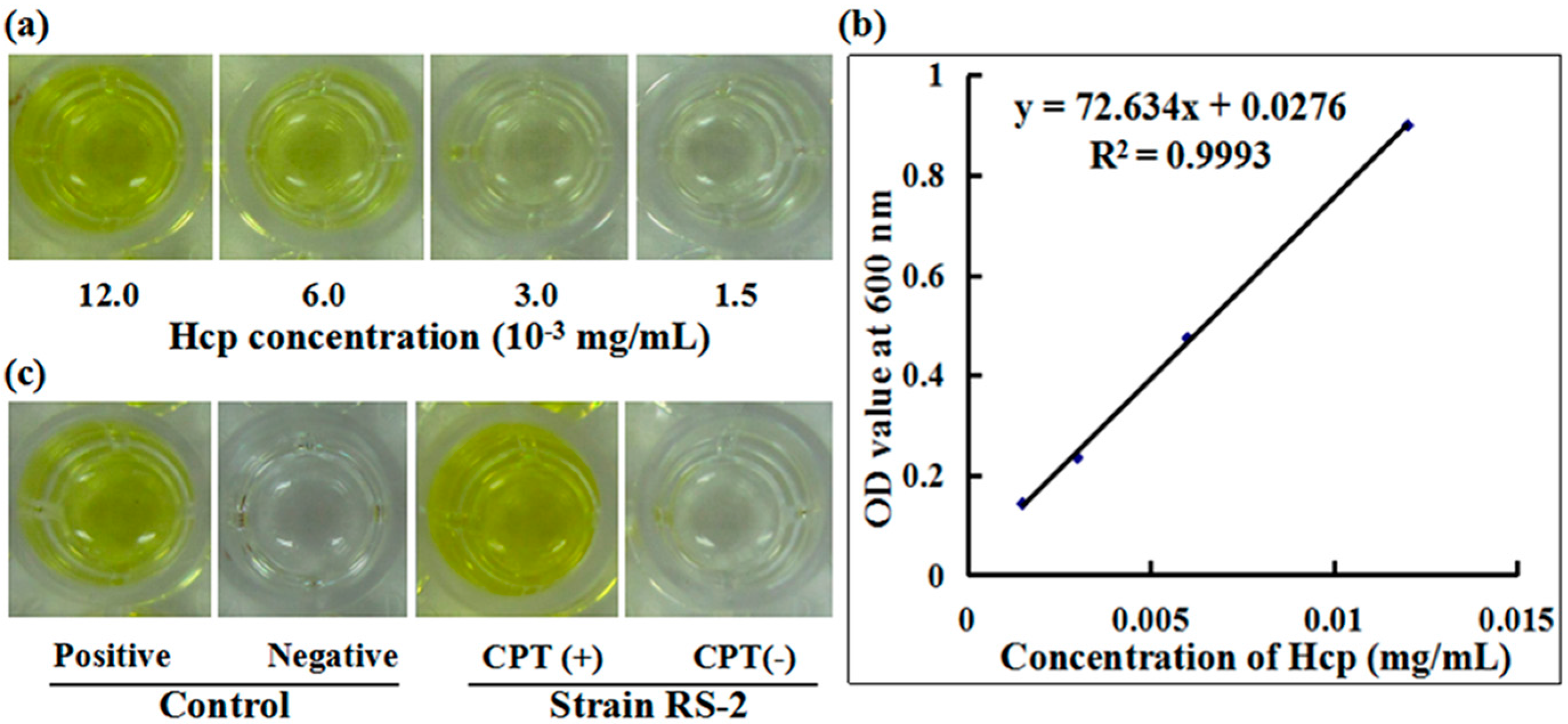
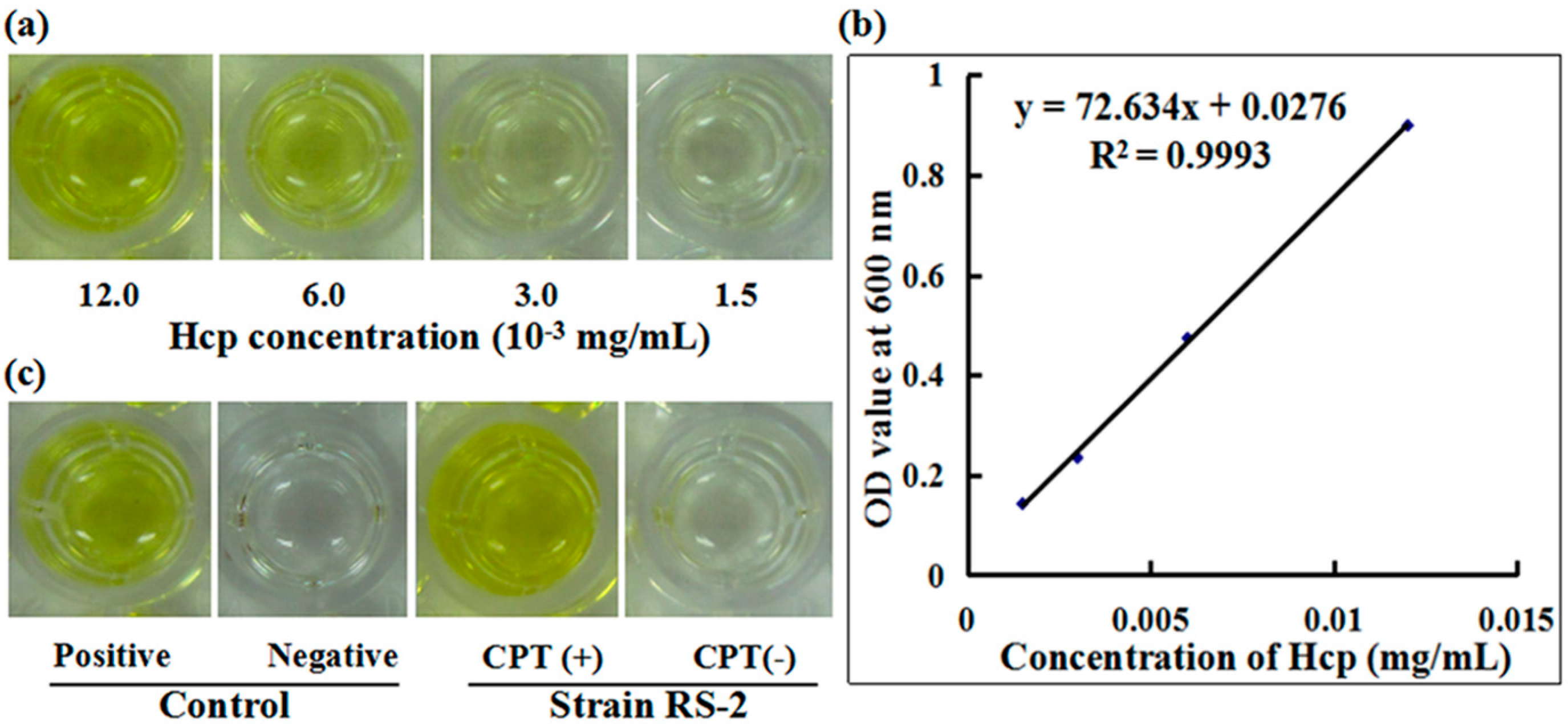
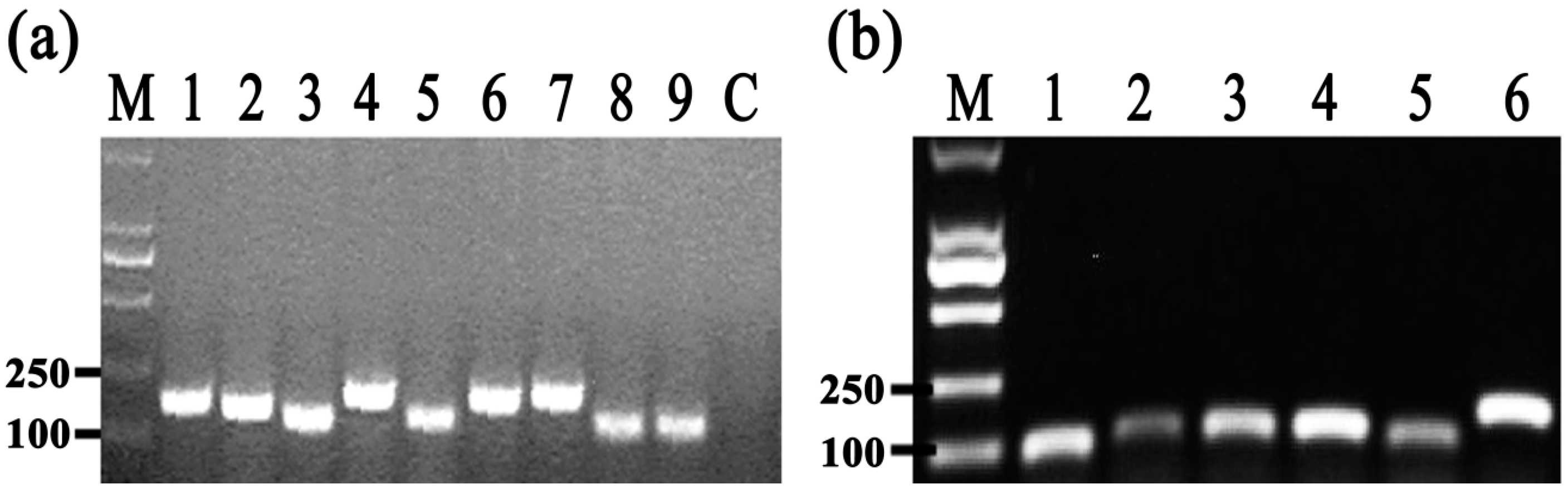
2.7. Secretion of Effector Protein Hcp
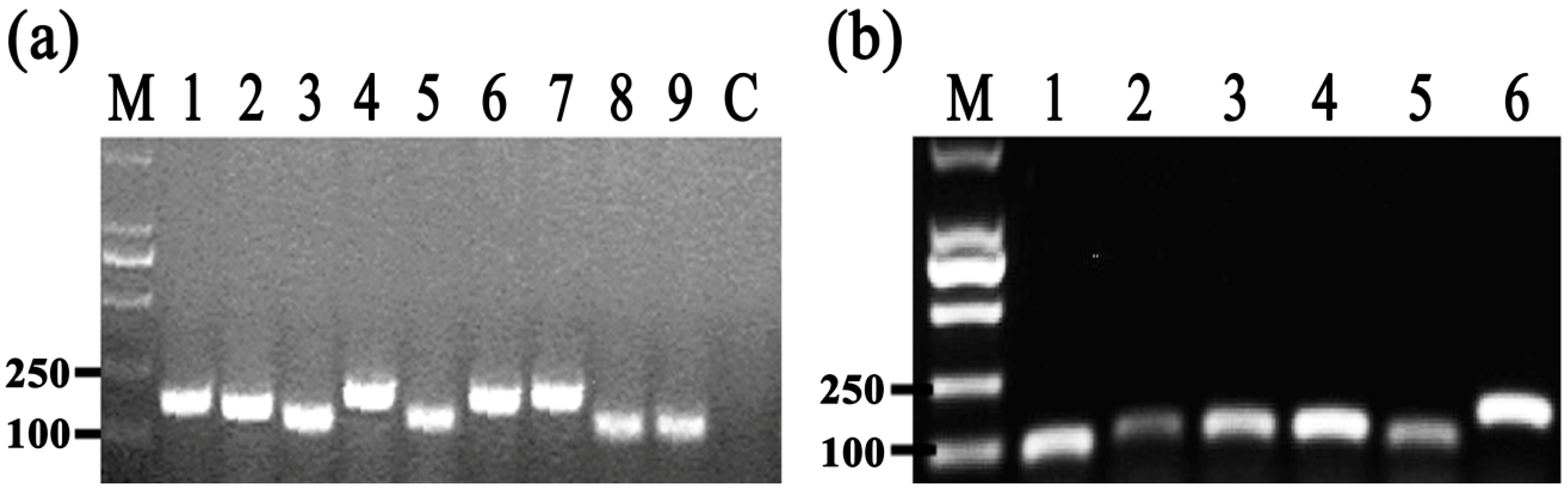
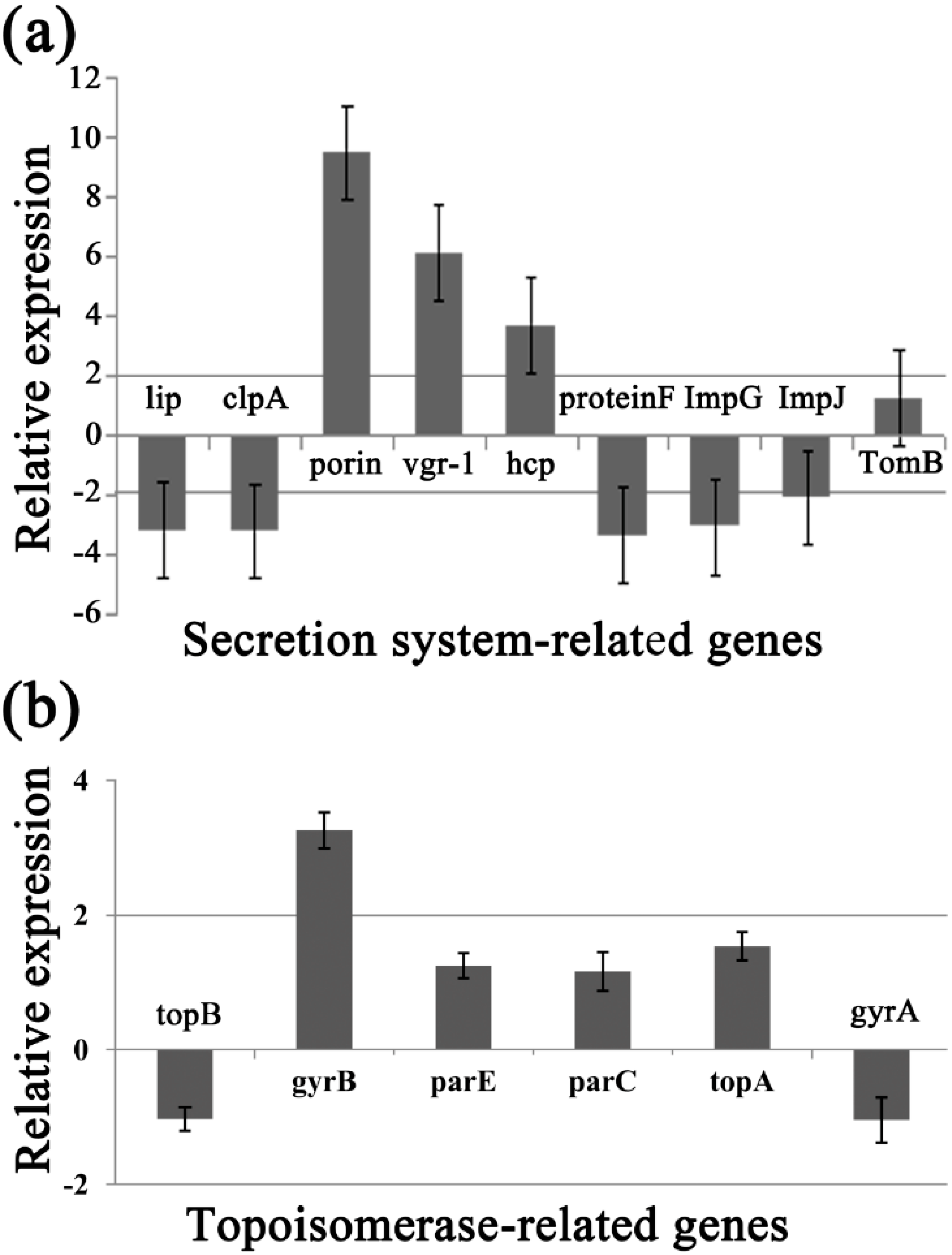
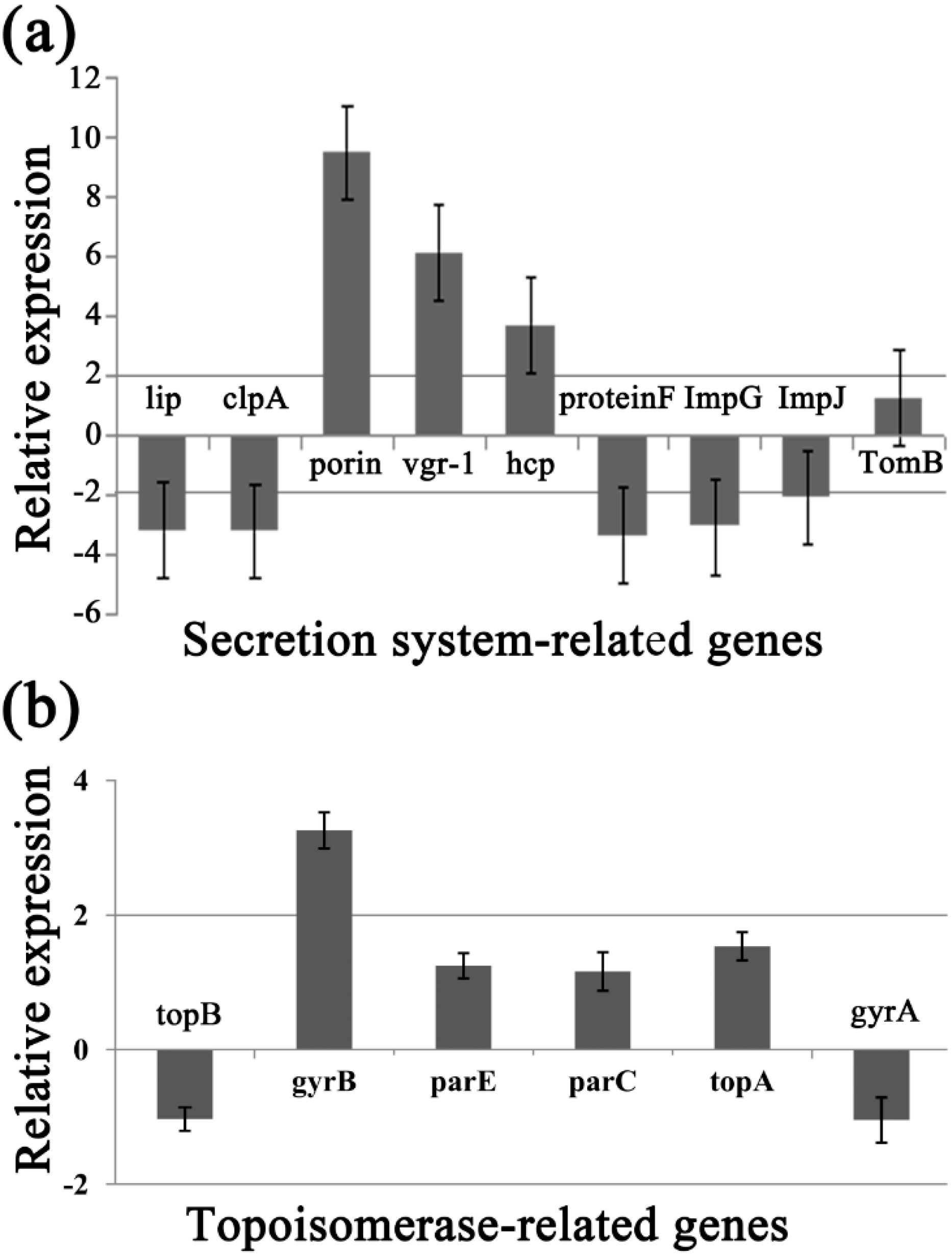
2.8. Differential Gene Expression
3. Experimental Section
3.1. Materials
3.2. In Vitro Antibacterial Activity of CPT
3.3. Swarming Motility
3.4. Live/Dead Cell Staining
3.5. Transmission Electron Microscopy (TEM)
3.6. Measurement of the Secreted Hcp by ELISA
3.7. RNA Isolation and Real-Time Quantitative PCR
3.8. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, S.; Zhang, Z.; Cain, A.; Wang, B.; Long, M.; Taylor, J. Antifungal activity of camptothecin, trifolin, and hyperoside isolated from Camptotheca acuminata. J. Agric. Food Chem. 2005, 53, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Ulukan, H.; Swaan, P.W. Camptothecins: A review of their chemotherapeutic potential. Drugs 2002, 62, 2039–2057. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Chen, S.W.; Yang, L.M.; Wang, R.R.; Pang, W.; Zheng, Y.T. The anti-HIV actions of 7- and 10-substituted camptothecins. Molecules 2010, 15, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, Y.; Wang, P.; Tong, S.M.; Ma, L.J. Antifungal activity of camptothecin on Rhizoctonia solani, Sphaerotheca fuliginea and Pseudoperonospora cubensis. J. Zhejiang For. Coll. 2008, 6, 681–684. [Google Scholar]
- Lee, J.H.; Lee, J.M.; Kim, J.K.; Ahn, S.K.; Lee, S.J.; Kim, M.Y.; Jew, S.S.; Park, J.G.; Hong, C.I. Antitumor activity of 7-[2-(N-isopropylamino) ethyl]-(20S)-camptothecin, CKD602, as a potent DNA topoisomerase I inhibitor. Arch. Pharm. Res. 1998, 21, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Hsiang, Y.H.; Liu, L.F. Identification of mammalian DNA topoisomerase I as an intracellular target of the anticancer drug camptothecin. Cancer Res. 1988, 48, 1722–1726. [Google Scholar] [PubMed]
- Sinha, B.K. Topoisomerase inhibitors. Drugs 1995, 49, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Pommier, Y.; Leo, E.; Zhang, H.; Marchand, C. DNA topoisomerases and their poisoning by anticancer and antibacterial drugs. Chem. Biol. 2010, 17, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.L.; Zhang, G.Q.; Liu, H.; Lou, M.M.; Tian, W.X.; Li, B.; Zhou, X.P.; Zhu, B.; Jin, G.L. Genome sequence of the rice-pathogenic bacterium Acidovorax avenae subsp. avenae RS-1. J. Bacteriol. 2011, 193, 5013–5014. [Google Scholar] [CrossRef] [PubMed]
- Kakar, K.; Nawaz, Z.; Cui, Z.; Almoneafy, A.; Zhu, B.; Xie, G.L. Characterizing the mode of action of Brevibacillus laterosporus B4 for control of bacterial brown strip of rice caused by A. avenae subsp. avenae RS-1. World J. Microbiol. Biotechnol. 2014, 30, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Liu, B.P.; Yu, R.R.; Tao, Z.Y.; Wang, Y.L.; Xie, G.L.; Li, H.; Sun, G.C. Bacterial brown stripe of rice in soil-less culture system caused by Acidovorax avenae subsp. avenae in China. J. Gen. Plant Pathol. 2011, 77, 64–67. [Google Scholar] [CrossRef]
- Yang, C.L.; Li, B.; Ge, M.Y.; Zhou, K.L.; Wang, Y.L.; Luo, J.; Ibrahim, M.; Xie, G.L.; Sun, G.C. Inhibitory effect and mode of action of chitosan solution against rice bacterial brown stripe pathogen Acidovorax avenae subsp. avenae RS-1. Carbohydr. Res. 2014, 391, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.Q.; Jin, G.; Li, B.; Kakar, K.U.; Ojaghian, M.R.; Wang, Y.L.; Xie, G.L.; Sun, G.C. Gene expression of type VI secretion system associated with environmental survival in Acidovorax avenae subsp. avenae by principle component analysis. Int. J. Mol. Sci. 2015, 16, 22008–22026. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tian, W.X.; Ibrahim, M.; Li, B.; Zhang, G.Q.; Zhu, B.; Xie, G.L. Characterization of PilP, a gene required for twitching motility, pathogenicity, and biofilm formation of Acidovorax avenae subsp. avenae RS-1. Eur. J. Plant Pathol. 2012, 134, 551–560. [Google Scholar] [CrossRef]
- Ibrahim, M.; Shi, Y.; Qiu, H.; Li, B.; Jabeen, A.; Li, L.; Liu, H.; Kube, M.; Xie, G.L.; Wang, Y.L. Differential expression of in vivo and in vitro protein profile of outer membrane of Acidovorax avenae subsp. avenae. PLoS ONE 2012, 7, e49657. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, L.; Ibrahim, M.; Ge, M.Y.; Wang, Y.L.; Mannan, S.; Asif, M.; Sun, G.C. Membrane protein profiling of Acidovorax avenae subsp. avenae under various growth conditions. Arch. Microbiol. 2015, 197, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yang, C.L.; Ge, M.Y.; Ibrahim, M.; Li, B.; Zhao, W.J.; Chen, G.Y.; Zhu, B.; Xie, G.L. Regulatory role of tetR gene in a novel gene cluster of Acidovorax avenae subsp. avenae RS-1 under oxidative stress. Front. Microbiol. 2014, 5, 547. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Ibrahim, M.; Ge, M.Y.; Cui, Z.Q.; Sun, G.C.; Xu, F.; Kube, M. Transcriptome analysis of Acidovorax avenae subsp. avenae cultivated in vivo and co-culture with Burkholderia seminalis. Sci. Rep. 2014, 4, 5698. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.Y.; Li, B.; Wang, L.; Tao, Z.Y.; Mao, S.F.; Wang, Y.L.; Xie, G.L.; Sun, G.C. Differentiation in MALDI-TOF MS and FTIR spectra between two pathovars of Xanthomonas oryzae. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 133, 730–734. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Kim, H.; Hwang, C.; Schaad, N. Detection of Acidovorax avenae ssp. avenae in Rice Seeds Using BIO-PCR. J. Phytopathol. 2004, 152, 667–676. [Google Scholar] [CrossRef]
- Shakya, D.; Chung, H. Detection of Pseudomonas avenae in rice seed. Seed Sci. Technol. 1983, 11, 583–587. [Google Scholar]
- Li, B.; Ge, M.Y.; Zhang, Y.; Wang, L.; Ibrahim, M.; Wang, Y.L.; Sun, G.C.; Chen, G.Y. New insights into virulence mechanisms of rice pathogen Acidovorax avenae subsp. avenae strain RS-1 following exposure to ß-lactam antibiotics. Sci. Rep. 2016, 6, 22241. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.F.; Luo, J.Y.; Wu, Z.Y.; Lei, P.; Ge, M.Y.; Li, B.; Wang, Y.L.; Zhang, L.Q.; Sun, G.C. Physiological and morphological changes in Botrytis cinerea induced by the plant alkaloid camptothecin. Trop. Plant. Pathol. 2015, 40, 260–264. [Google Scholar] [CrossRef]
- Connelly, M.B.; Young, G.M.; Sloma, A. Extracellular proteolytic activity plays a central role in swarming motility in Bacillus subtilis. J. Bacteriol. 2004, 186, 4159–4167. [Google Scholar] [CrossRef] [PubMed]
- Lorence, A.; Nessler, C.L. Camptothecin, over four decades of surprising findings. Phytochemistry 2004, 65, 2735–2749. [Google Scholar] [CrossRef] [PubMed]
- Pagán, R.; Mackey, B. Relationship between membrane damage and cell death in pressure-treated Escherichia coli cells: Differences between exponential- and stationary-phase cells and variation among strains. Appl. Environ. Microbiol. 2000, 66, 2829–2834. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.T.; Basler, M.; Mekalanos, J.J. Type 6 secretion system-mediated immunity to type 4 secretion system-mediated horizontal gene transfer. Science 2013, 342, 250–253. [Google Scholar] [CrossRef] [PubMed]
- Achouak, W.; Heulin, T.; Pages, J.M. Multiple facets of bacterial porins. FEMS Microbiol. Lett. 2001, 199, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jani, A.J.; Cotter, P.A. Type VI secretion: Not just for pathogenesis anymore. Cell Host Microbe 2010, 8, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Blondel, C.J.; Jiménez, J.C.; Contreras, I.; Santiviago, C.A. Comparative genomic analysis uncovers 3 novel loci encoding type six secretion systems differentially distributed in Salmonella serotypes. BMC Genom. 2009, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Weber, B.; Hasic, M.; Chen, C.; Wai, S.N.; Milton, D.L. Type VI secretion modulates quorum sensing and stress response in Vibrio anguillarum. Environ. Microbiol. 2009, 11, 3018–3028. [Google Scholar] [CrossRef] [PubMed]
- Aschtgen, M.S.; Bernard, C.S.; De Bentzmann, S.; Lloubès, R.; Cascales, E. SciN is an outer membrane lipoprotein required for type VI secretion in enteroaggregative Escherichia coli. J. Bacteriol. 2008, 190, 7523–7531. [Google Scholar] [CrossRef] [PubMed]
- Gerth, U.; Krüger, E.; Derré, I.; Msadek, T.; Hecker, M. Stress induction of the Bacillus subtilis clpP gene encoding a homologue of the proteolytic component of the Clp protease and the involvement of ClpP and ClpX in stress tolerance. Mol. Microbiol. 1998, 28, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.M.; Jedrychowski, M.P.; Wei, J.R.; Pinkham, J.T.; Park, A.S.; O’Brien, K.; Rehren, G.; Schnappinger, D.; Gygi, S.P.; Rubin, E.J. Post-translational regulation via Clp protease is critical for survival of Mycobacterium tuberculosis. PLoS Pathog. 2014, 10, e1003994. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.M.; Unnikrishnan, M.; Rubin, D.; Krishnamoorthy, V.; Kandror, O.; Akopian, T.N.; Goldberg, A.L.; Rubin, E.J. Mycobacterium tuberculosis ClpP1 and ClpP2 function together in protein degradation and are required for viability in vitro and during infection. PLoS Pathog. 2012, 8, e1002511. [Google Scholar] [CrossRef] [PubMed]
- Burtnick, M.N.; Brett, P.J. Burkholderia mallei and Burkholderia pseudomallei cluster 1 type VI secretion system gene expression is negatively regulated by iron and zinc. PLoS ONE 2013, 8, e76767. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.A.; Askora, A.; Kawasaki, T.; Fujie, M.; Yamada, T. The filamentous phage XacF1 causes loss of virulence in Xanthomonas axonopodis pv. citri, the causative agent of citrus canker disease. Front. Microbiol. 2014, 5, 321. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.Q.; Ibrahim, M.; Yang, C.L.; Fang, Y.; Annam, H.; Li, B.; Wang, Y.L.; Xie, G.L.; Sun, G.C. Susceptibility of opportunistic burkholderia glumae to copper surfaces following wet or dry surface contact. Molecules 2014, 19, 9975–9985. [Google Scholar] [CrossRef] [PubMed]
- Lou, M.M.; Zhu, B.; Muhammad, I.; Li, B.; Xie, G.L.; Wang, Y.L.; Li, H.Y.; Sun, G.C. Antibacterial activity and mechanism of action of chitosan solutions against apricot fruit rot pathogen Burkholderia seminalis. Carbohydr. Res. 2011, 346, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Slutzki, M.; Barak, Y.; Reshef, D.; Schueler-Furman, O.; Lamed, R.; Bayer, E.A. Indirect ELISA-based approach for comparative measurement of high-affinity cohesin–dockerin interactions. J. Mol. Recognit. 2012, 25, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yan, L.; Gu, Q.; Ma, Z. The mitogen-activated protein kinase kinase kinase BcOs4 is required for vegetative differentiation and pathogenicity in Botrytis cinerea. Appl. Environ. Microbiol. 2012, 96, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Primer Sequence (5′-3′) | Putative Protein Function (s) | Amplication Size (bp) |
|---|---|---|---|
| 16S rDNA | F-TTGCGGTCCCCTGCTTTCAT | 120 | |
| R-CGGTAACAGGTCTTCGGATGCT | |||
| Secretion system | |||
| clpA | F-CACGATCCAGATCCTCTGCC | ATP-dependent Clp protease | 163 |
| R-GCCCATGTCCAGCGAATAGA | |||
| porin | F-GCACCACCTGGTCCACAACA | Porin Gram-negative type | 192 |
| R-GCGTCGAACATTCCAAGGTG | |||
| protein F | F-CCGCGGGAATTCTCTTCCAT | General secretion pathway protein F T2SF; Type II secretion system (T2SS) reductase | 131 |
| R-CCGATGAGCTTGGCCTTGA | |||
| tomB | F-ATCAACCGCTACCGC | TomB protein interacts with outer membrane receptor proteins that carry out high-affinity binding and energy-dependent uptake of specific substrates into the periplasmic space | 129 |
| R-ATCAAGGTCTGGACCTCGTTGAAGG | |||
| impG | F-GCCACAAGTTCCTTTTGCA | Type VI secretion protein ImpG | 181 |
| R-AAGAACGGCACGAAATCC | |||
| impJ | F-TCCAGGATGCCAACGACA | Type VI secretion protein ImpJ | 181 |
| R-GACCACGGTGGGAATGAA | |||
| lip | F-GCAGTGCGGATGTCCGTACCTT | Type VI secretion lipoprotein | 174 |
| R-TCCTTGCCCACCGTGATGCT | |||
| hcp | F-GACTCCGTGCTGGTGAAT | Type VI secretion system effector | 100 |
| R-TTCCCTTGATGTCGTAGCC | |||
| vgr-1 | F-ATCCGATGGAAAAGAAACTC | RND efflux system outer membrane lipoprotein [Acidovorax citrulli AAC00-1] | 113 |
| R-AATAGATGCCCTCGTGCT | |||
| Topoisomerase | |||
| topB | F-CTGGACAAGTTCGTGAGCAT | DNA topoisomerase III, Burkholderia type | 89 |
| R-AACTCGAAGTTCACCTTGCC | |||
| gyrB | F-CTGCTTCACCAACAACATCC | DNA gyrase subunit B | 99 |
| R-TTCTGCTCGATGTACTTGCC | |||
| parE | F-ATGAACGAACTGGTCCACCT | Topoisomerase IV subunit B | 105 |
| R-GTATTGCCAGGTCTGGGTCT | |||
| parC | F-CAAGATCGAGCAGGAACTCA | Topoisomerase IV subunit A | 106 |
| R-CTCGATCTCCTTGACCATGA | |||
| topA | F-GGGCAGGAACTCTTCCAGTA | DNA topoisomerase IB (poxvirus type) | 83 |
| R-TCGTGCAGGTAGTCGTTCAC | |||
| gyrA | F-GCACGACTACATCCTGTGCT | DNA gyrase subunit A | 148 |
| R-CAGCACCACGTTGATCTTCT | |||
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Q.; Luo, J.; Qiu, W.; Cai, L.; Anjum, S.I.; Li, B.; Hou, M.; Xie, G.; Sun, G. Inhibitory Effect of Camptothecin against Rice Bacterial Brown Stripe Pathogen Acidovorax avenae subsp. avenae RS-2. Molecules 2016, 21, 978. https://doi.org/10.3390/molecules21080978
Dong Q, Luo J, Qiu W, Cai L, Anjum SI, Li B, Hou M, Xie G, Sun G. Inhibitory Effect of Camptothecin against Rice Bacterial Brown Stripe Pathogen Acidovorax avenae subsp. avenae RS-2. Molecules. 2016; 21(8):978. https://doi.org/10.3390/molecules21080978
Chicago/Turabian StyleDong, Qiaolin, Ju Luo, Wen Qiu, Li Cai, Syed Ishtiaq Anjum, Bin Li, Mingsheng Hou, Guanlin Xie, and Guochang Sun. 2016. "Inhibitory Effect of Camptothecin against Rice Bacterial Brown Stripe Pathogen Acidovorax avenae subsp. avenae RS-2" Molecules 21, no. 8: 978. https://doi.org/10.3390/molecules21080978
APA StyleDong, Q., Luo, J., Qiu, W., Cai, L., Anjum, S. I., Li, B., Hou, M., Xie, G., & Sun, G. (2016). Inhibitory Effect of Camptothecin against Rice Bacterial Brown Stripe Pathogen Acidovorax avenae subsp. avenae RS-2. Molecules, 21(8), 978. https://doi.org/10.3390/molecules21080978