
Immunomodulatory and Antioxidant Effects of Polysaccharides from Gynostemma pentaphyllum Makino in Immunosuppressed Mice

Abstract
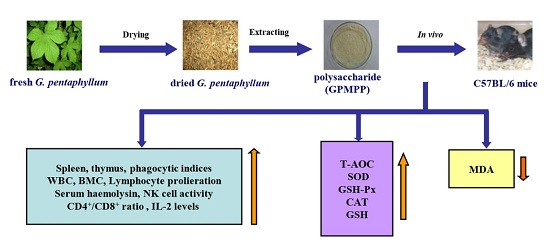
:
1. Introduction
2. Results and Discussions
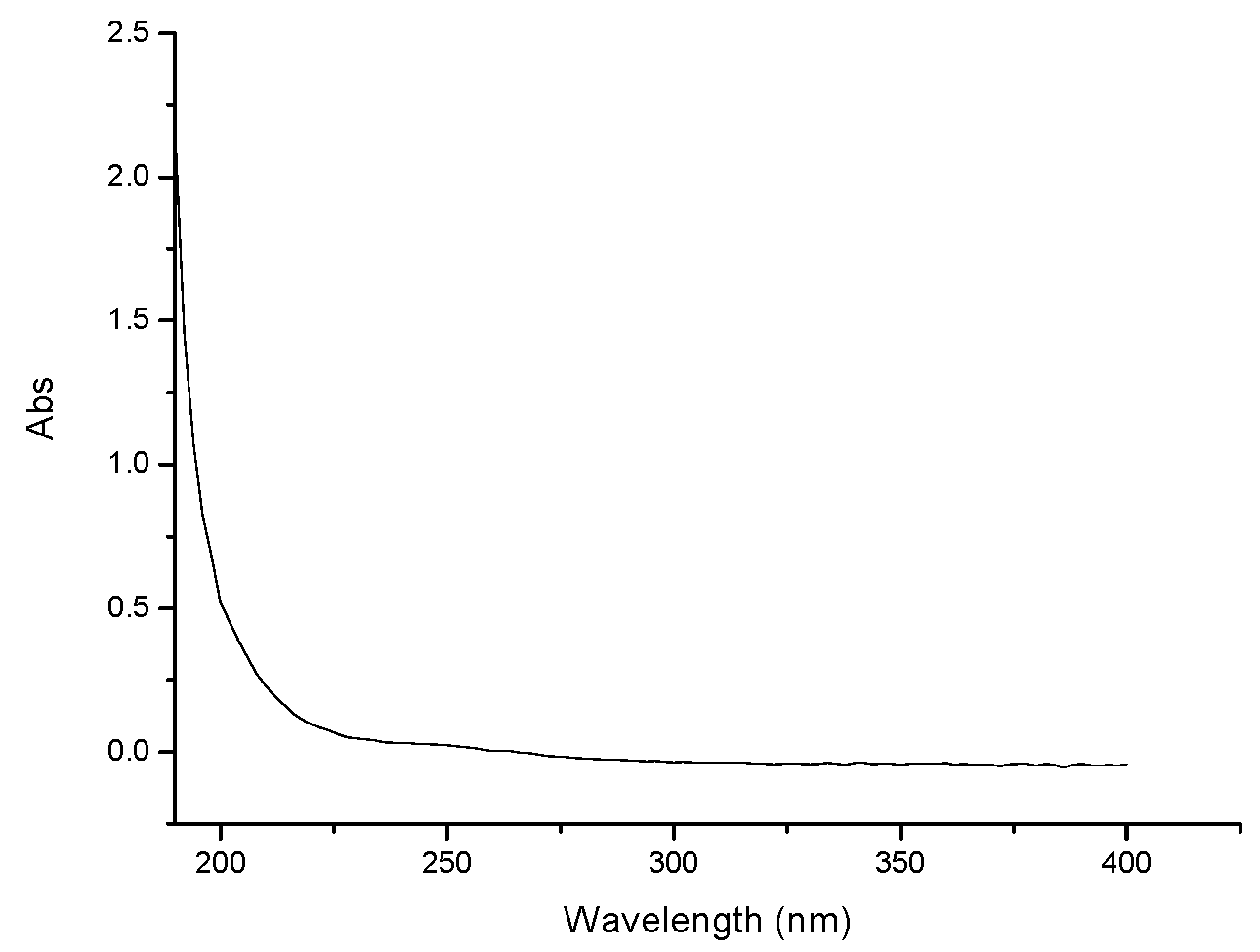
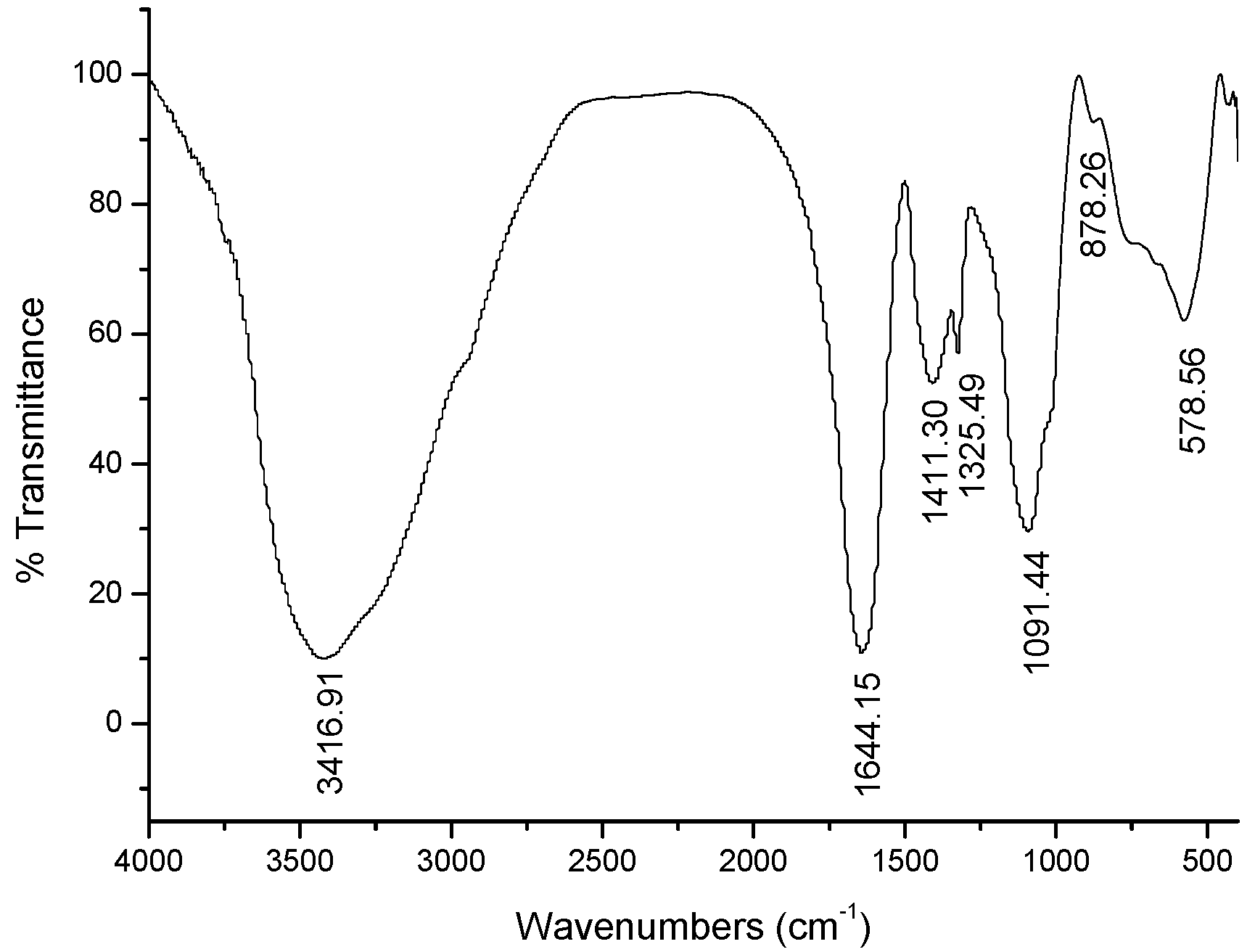
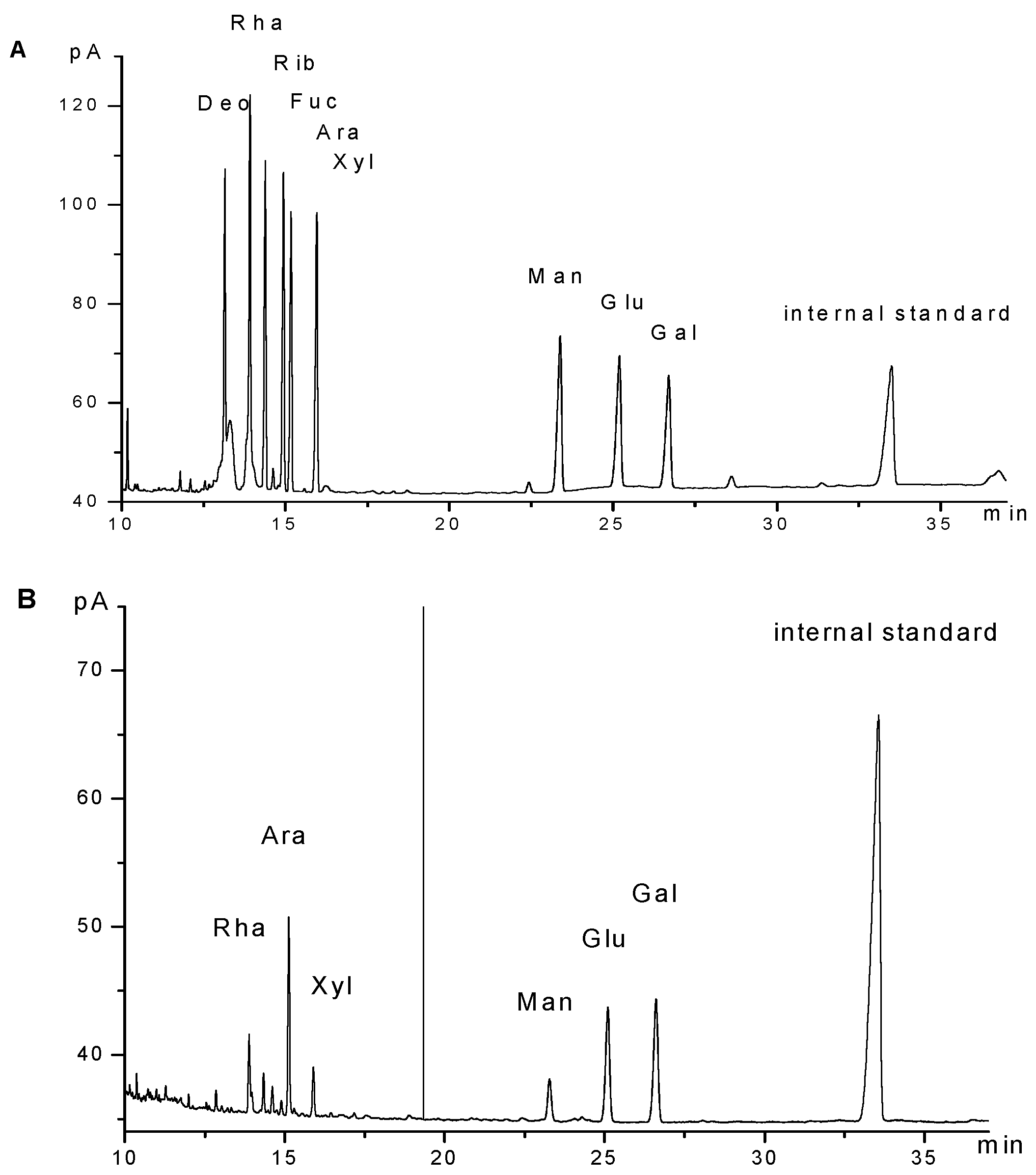
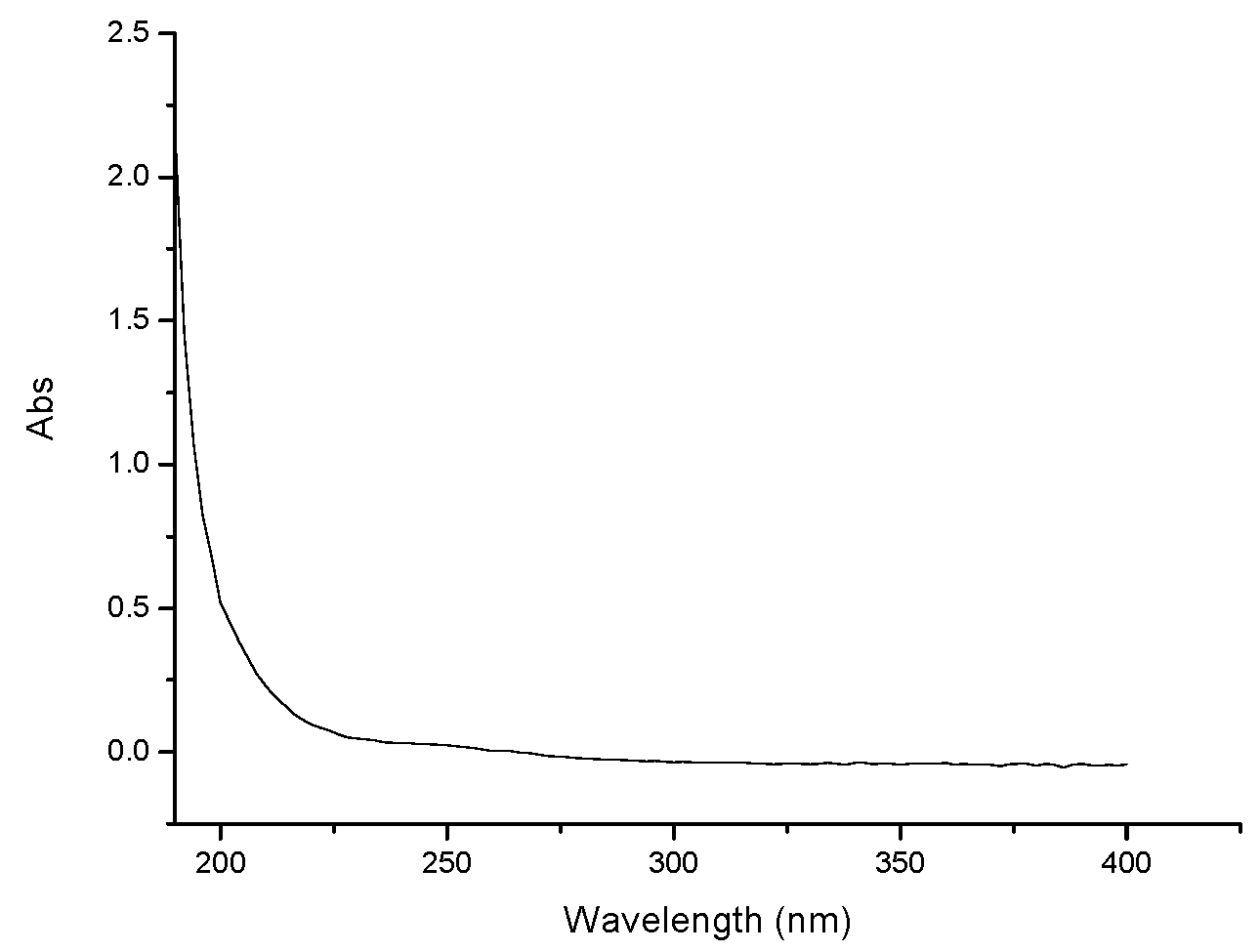
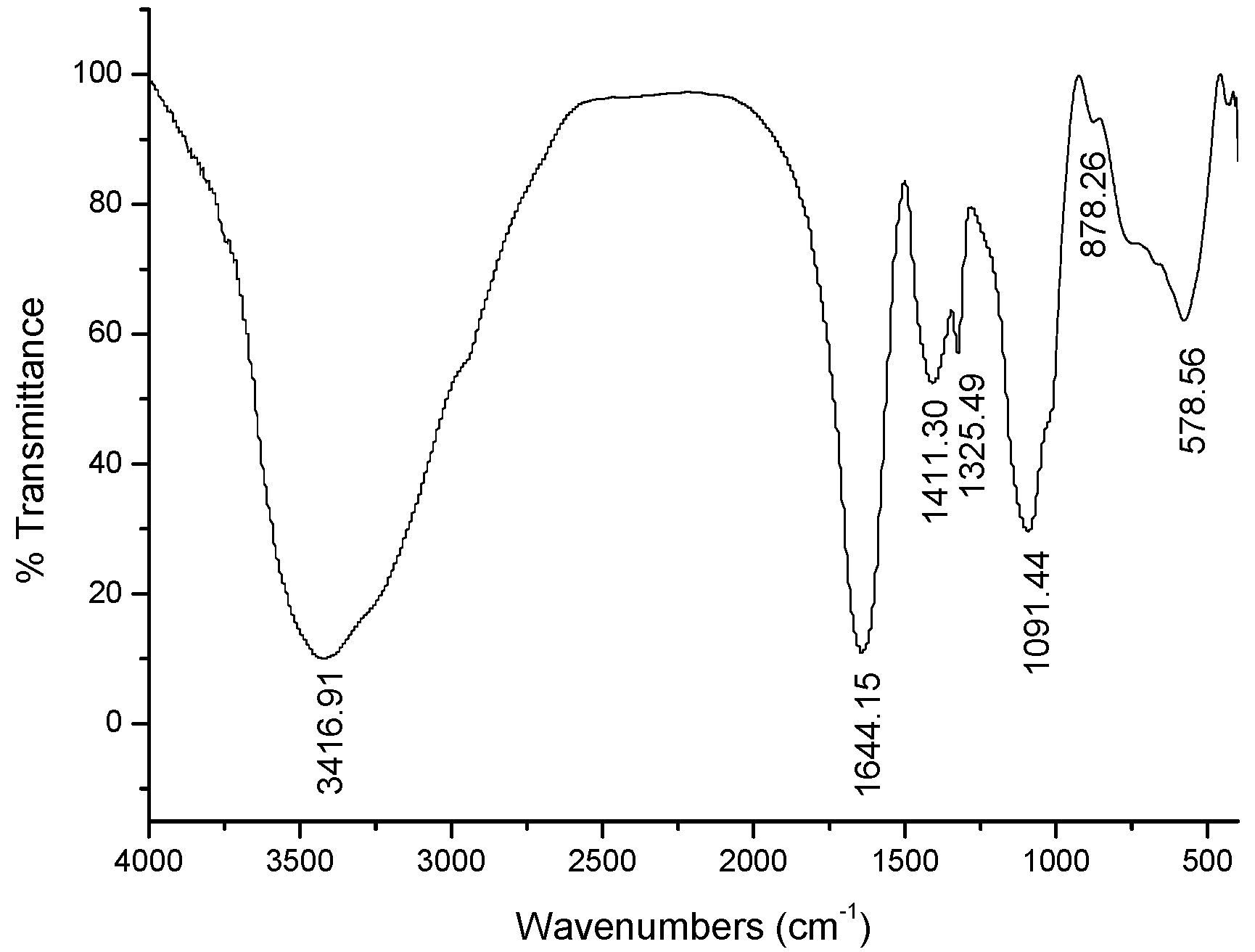
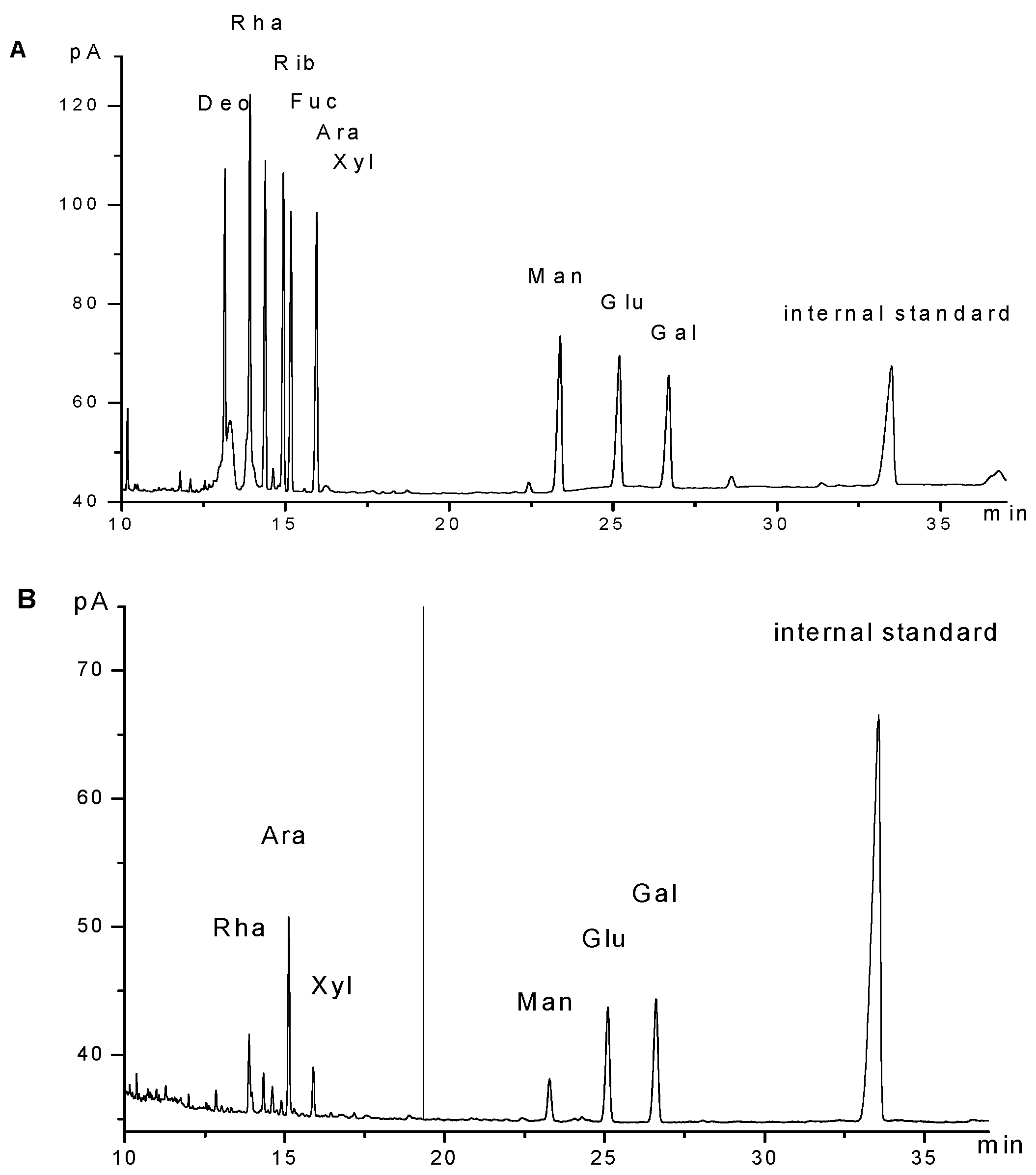
2.1. Characterization of GPMPP
2.2. Effects of GPMPP on Thymus and Spleen Indices in Cy-Immunosuppressed Mice
2.3. Effects of GPMPP on Macrophage Phagocytosis in Cy-Immunosuppressed Mice
2.4. Effects of GPMPP on Leukocytes and Bone Marrow Cells in Cy-Immunosuppressed Mice
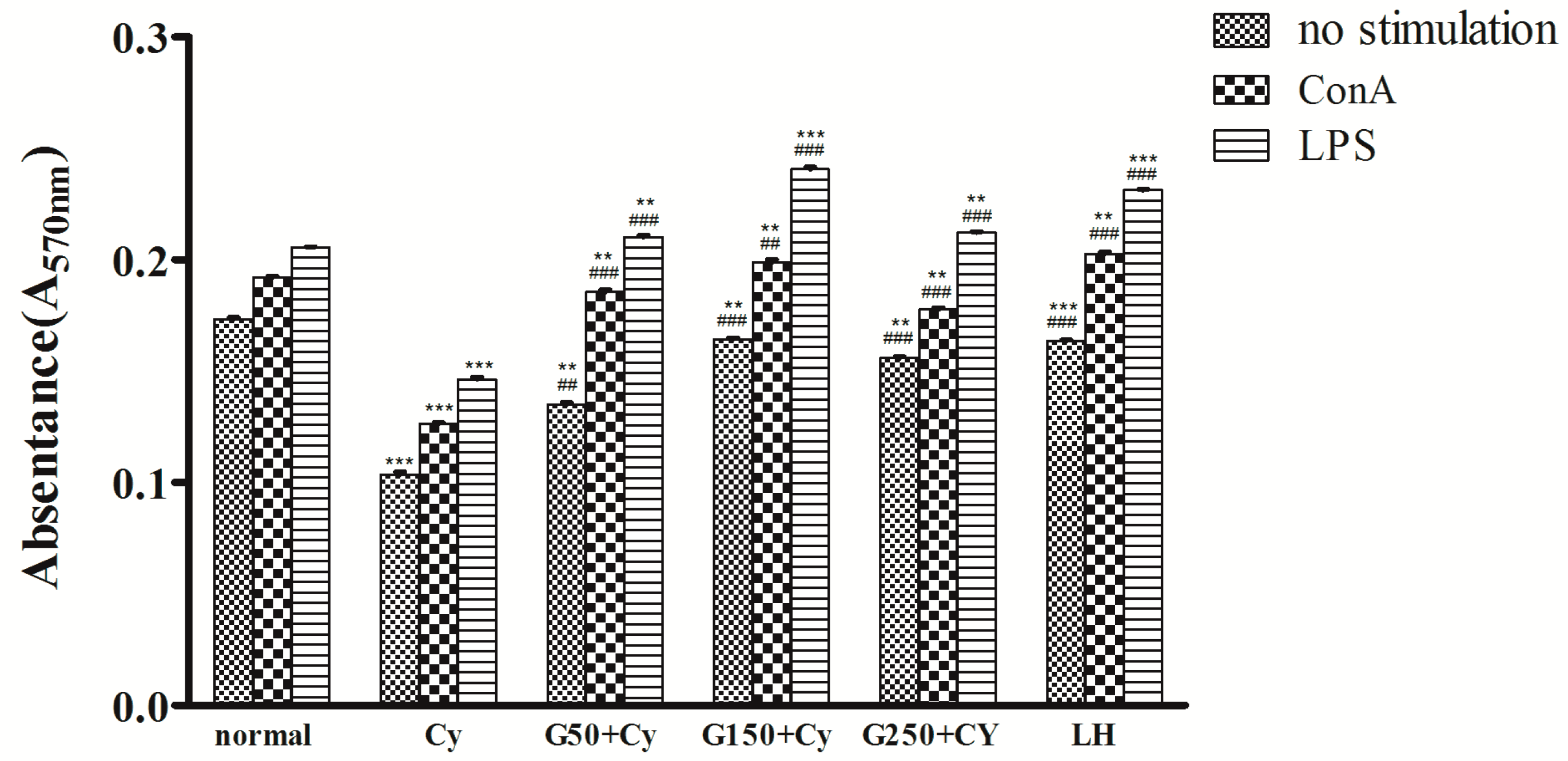
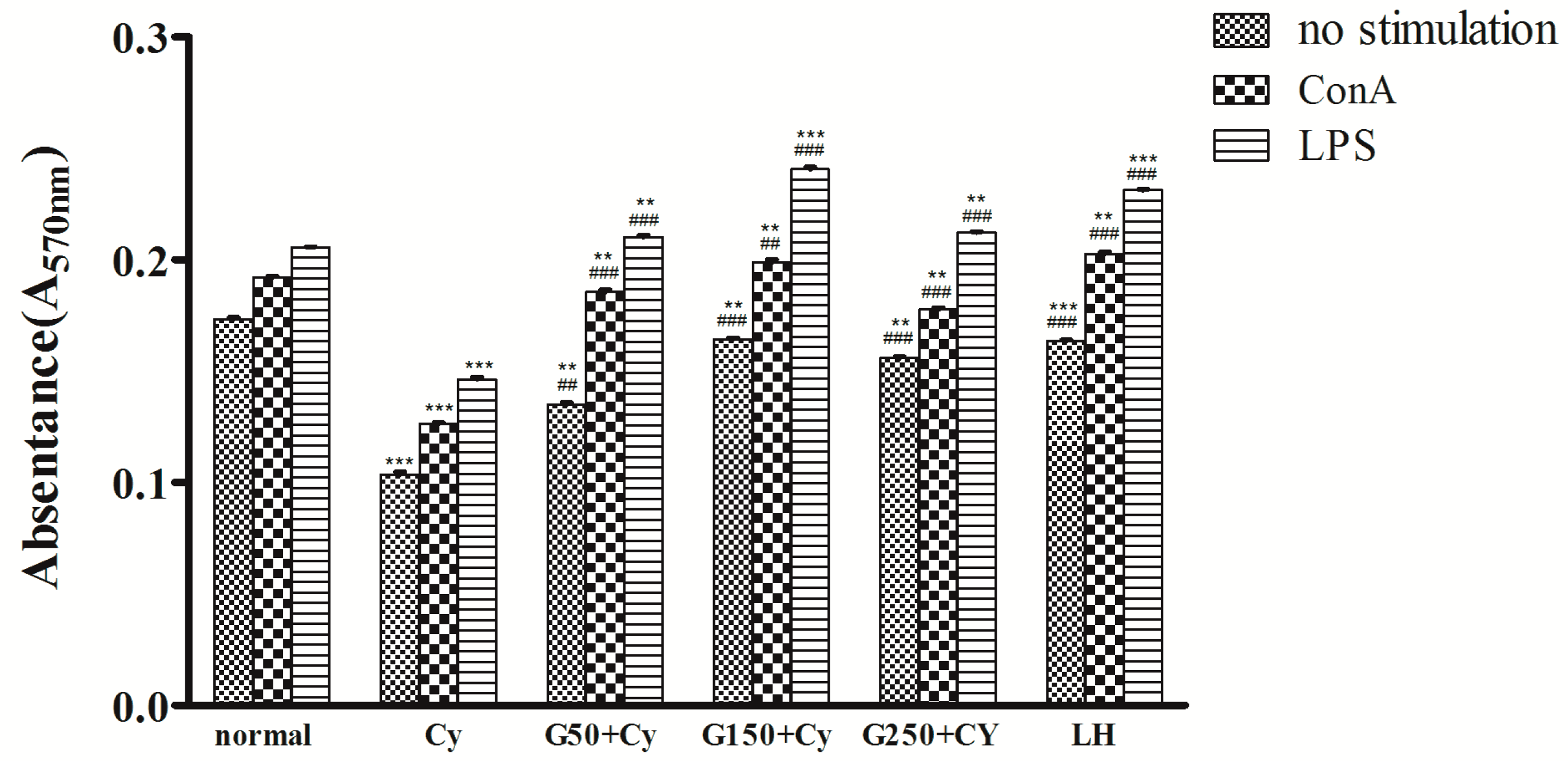
2.5. Effects of GPMPP on Lymphocyte Proliferation in Cy-Immunosuppressed Mice
2.6. Effects of GPMPP on Serum Haemolysin in Cy-Immunosuppressed Mice
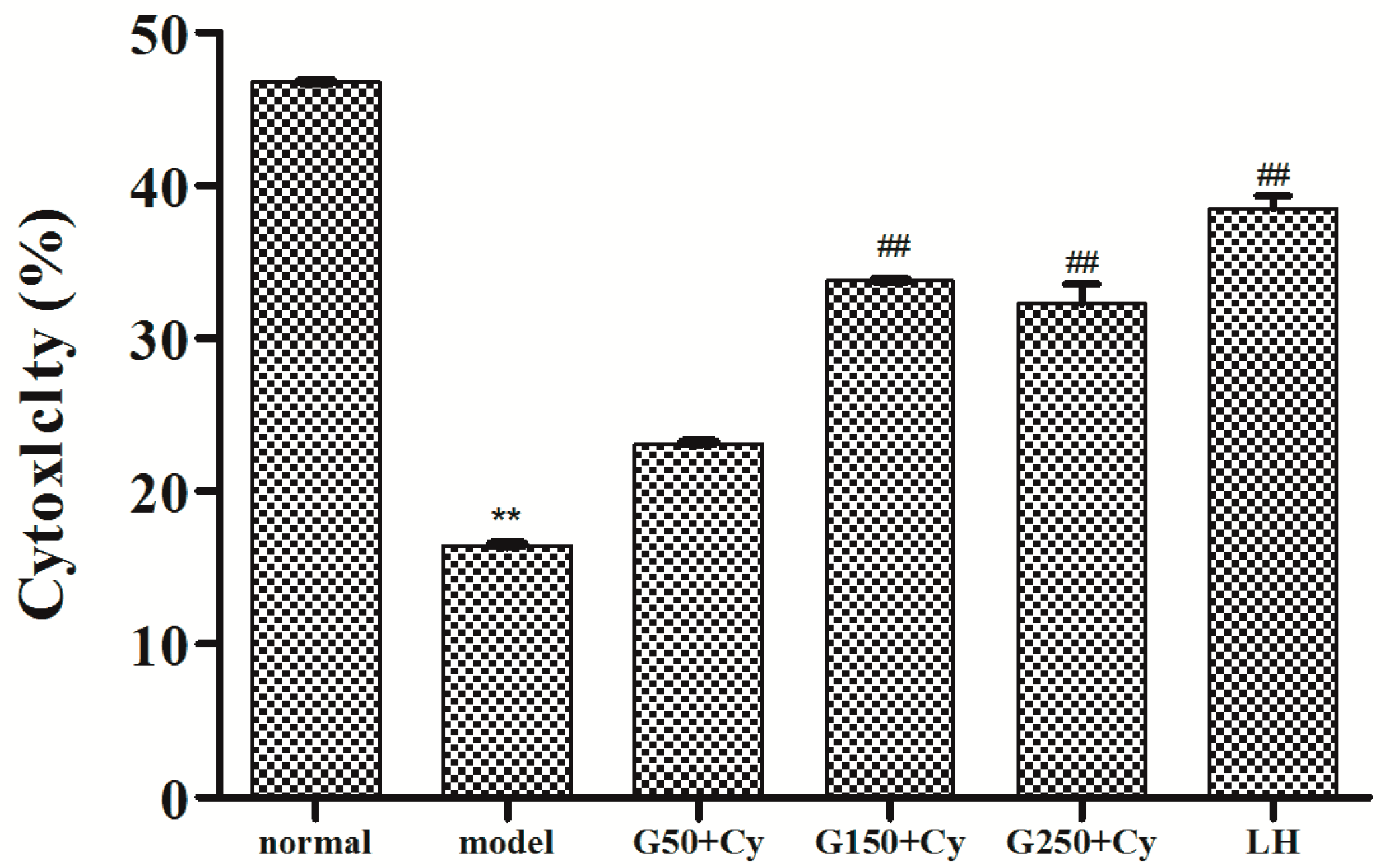
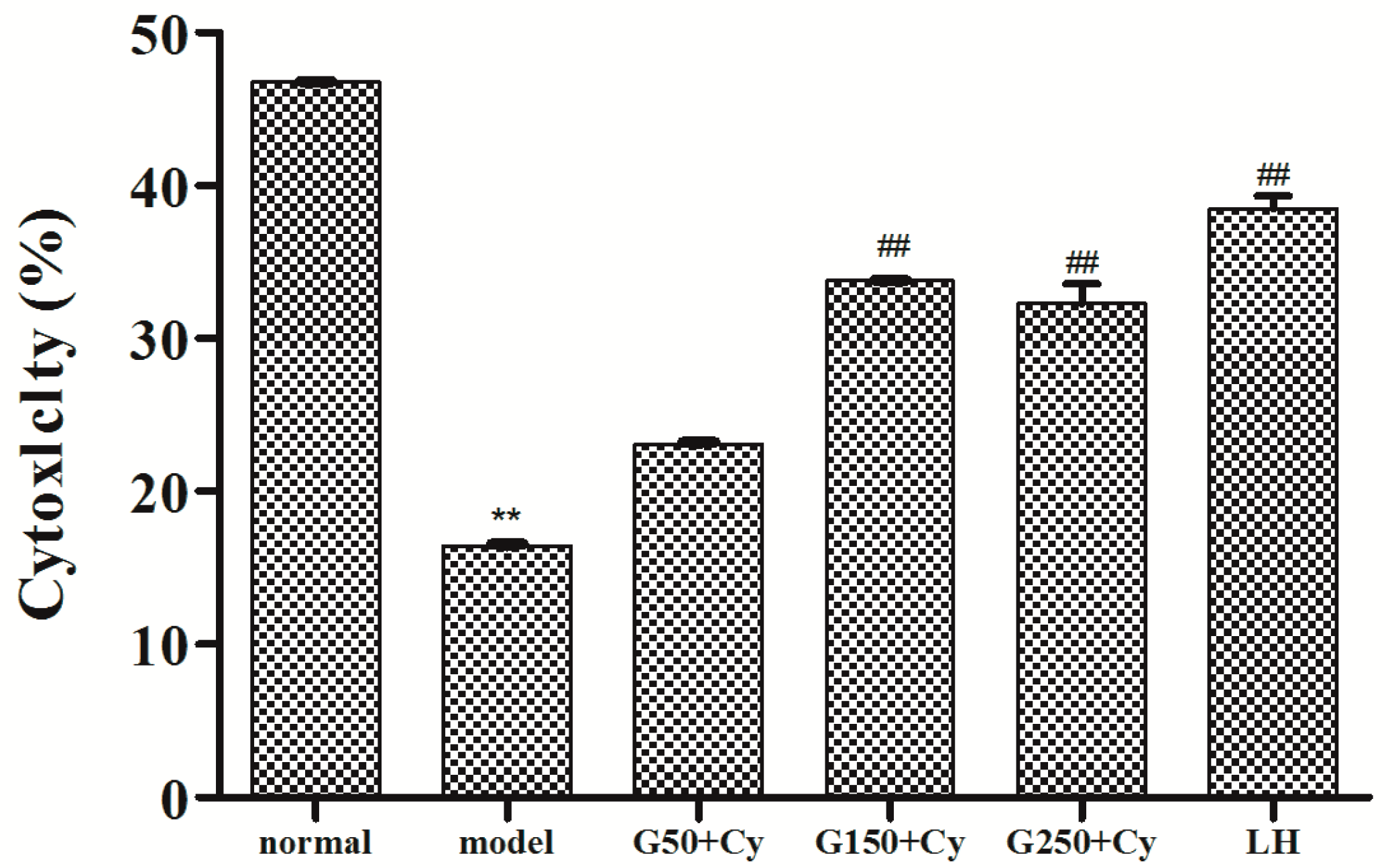
2.7. Effects of GPMPP on Natural Killer Cell Activity in Cy-immunosuppressed Mice
2.8. Effects of GPMPP on T-Lymphocyte Phenotyping in Cy-Immunosuppressed Mice
2.9. Effects of GPMPP on IL-2 Expression in Cy-Immunosuppressed Mice
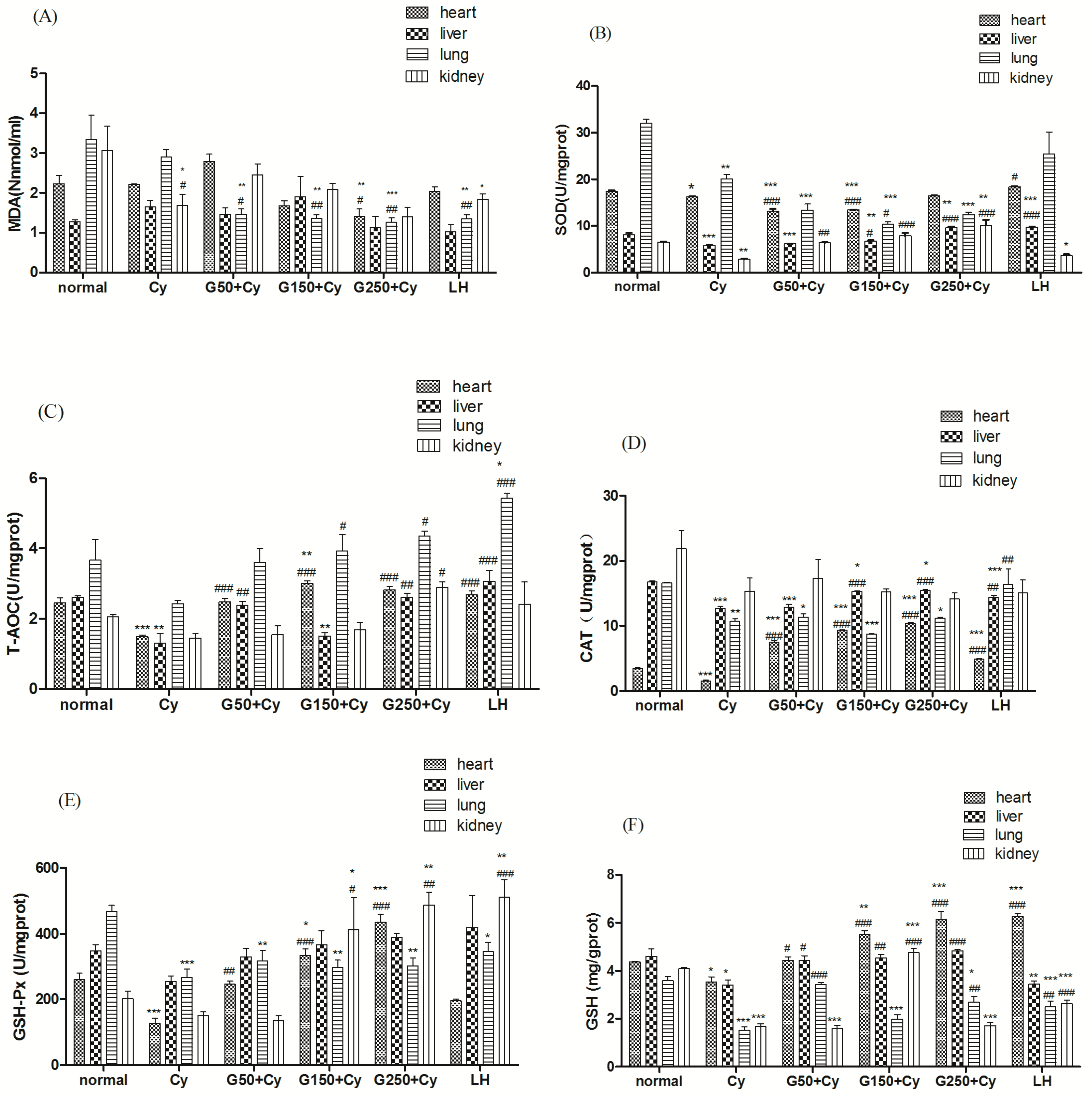
2.10. Effects of GPMPP on Activities of Antioxidant Enzymes in Cy-Immunosuppressed Mice
3. Experimental Section
3.1. Materials
3.2. Preparation of the Crude Polysaccharides from G. pentaphyllum Makino
3.3. Characterization of GPMPP
3.4. Animals and Experimental Design
3.5. Macrophage Phagocytosis Assay
3.6. Measurements of Leukocytes and Bone Marrow Cells
3.7. Splenocyte Proliferation Assay
3.8. Serum Haemolysin Assay
3.9. Natural Killer Cell Activity Assay
3.10. T-Lymphocyte Phenotyping Analysis
3.11. IL-2 Expression Assay
3.12. Biochemical Assay
3.13. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lobo, V.; Phatak, A.; Chandra, N. Free radicals and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Lü, J.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and molecular mechanisms of antioxidants: Experimental approaches and model systems. J. Cell. Mod. Med. 2010, 14, 840–860. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.P.; Chandra, A.; Mahdi, F.; Ray, A.; Sharma, P. Reconvene and reconnect the antioxidant hypothesis in Human health and disease. Ind. J. Clin. Biochem. 2010, 25, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Victor, V.M.; Rocha, M.; de la Fuente, M. Immune cells: Free radicals and antioxidants in sepsis. Int. Immunopharmacol. 2004, 4, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, S.A.; Gordon, L.I. Inflammation and cancer: Role of phagocyte-generated oxidants in carcinogenesis. Blood 1990, 76, 655–663. [Google Scholar] [PubMed]
- De la Fuente, M. Effects of antioxidants on immune system ageing. Eur. J. Clin. Nutr. 2002, 56, S5–S8. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.; Galley, H.F.; Webster, N.R. Oxidative stress and gene expression in sepsis. Br. J. Anaesth. 2003, 90, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Gutteridge, J.M.; Michell, J. Redox imbalance in the critically ill. Br. Med. Bull. 1999, 55, 49–75. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, M.; Victor, V. Antioxidants as modulators of immune function. Immunol. Cell Biol. 2000, 78, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Kong, F.L.; Li, F.E.; He, Z.M.; Jiang, Y.; Hao, R.Y.; Sun, X.; Tong, H.B. Anti-tumor and macrophage activation induced by alkali-extracted polysaccharide from pleurotus ostreatus. Int. J. Biol. Macromol. 2014, 69, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ren, C.J.; Lu, G.B.; Cui, W.Z.; Mu, Z.M.; Gao, H.J.; Wang, Y.W. Purification, characterization and anti-diabetic activity of a polysaccharide from mulberry leaf. Regul. Toxicol. Pharm. 2014, 70, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.G.; Mesquita, J.X.; Aragao, K.S.; Franco, A.X.; Souza, M.H.L.P.; Brito, T.V.; Dias, J.M.; Silva, R.O.; Medeiros, J.V.R.; Oliveira, J.S. Polysaccharides isolated from digenea simplex inhibit inflammatory and nociceptive responses. Carbohydr. Polym. 2014, 108, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Liu, L.N.; Lin, C.W. Isolation, structural characterization and antioxidant activity of a neutral polysaccharide from Sisal waste. Food Hydrocolloid. 2014, 39, 10–18. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.M.; Qi, Z.M.; Wang, S.Y.; Liu, S.X.; Li, X.; Wang, H.J.; Xia, X.C. An overview on natural polysaccharides with antioxidant properties. Curr. Med. Chem. 2013, 20, 2899–2913. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Hu, Y.K.; Shi, S.J.; Jiang, L. Evaluation of antioxidant and immune-enhancing activities of Purslane polysaccharides in gastric cancer rats. Int. J. Biol. Macromol. 2014, 68, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.H.; Huang, H.Q.; Zhao, Y.; Shi, H.M.; Wang, S.K.; Wang, T.T.Y.; Chen, P.; Yu, L.L. Chemical composition and anti-proliferative and anti-inflammatory effects of the leaf and whole-plant samples of diploid and tetraploid Gynostemma pentaphyllum (Thunb.) Makino. Food Chem. 2012, 132, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Razmovski-Naumovski, V.; Huang, T.H.W.; Tran, V.H.; Li, G.; Duke, C.; Roufogalis, B. Chemistry and pharmacology of Gynostemma pentaphyllum. Phytochem. Rev. 2005, 4, 197–219. [Google Scholar] [CrossRef]
- Yan, W.; Niu, Y.G.; Lv, J.L.; Xie, Z.H.; Jin, L.; Yao, W.B.; Gao, X.D.; Yu, L.L. Characterization of a heteropolysaccharide isolated from diploid Gynostemma pentaphyllum Makino. Carbohydr. Polym. 2013, 92, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.G.; Shang, P.P.; Chen, L.; Zhang, H.; Gong, L.; Zhang, X.W.; Yu, W.J.; Xu, Y.H.; Wang, Q.; Yu, L.L. Characterization of a novel alkali-soluble heteropolysaccharide from tetraploid Gynostemma pentaphyllum Makino and its potential anti-inflammatory and antioxidant properties. J. Agric. Food Chem. 2014, 62, 3783–3790. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, X.Y.; Wang, M.Z.; Jiao, L.L. Characterization and antioxidant activities of acidic polysaccharides from Gynostemma pentaphyllum (Thunb.) Markino. Carbohydr. Polym. 2015, 127, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, L.H.; Ren, Y.G.; Gao, Y.L.; Kang, L.; Qiao, Q. Anticancer and immunoregulatory activity of Gynostemma pentaphyllum polysaccharides in H22 tumor-bearing mice. Int. J. Biol. Macromol. 2014, 69, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.Y.; Zhang, Y.; Bai, Y.L.; Xu, C.L.; Niu, W.N.; Qin, C.G. Extraction and antioxidant activities of polysaccharides from Gynostemma pentaphyllum Makino. Asian J. Chem. 2013, 25, 9092–9096. [Google Scholar]
- Xie, J.H.; Xie, M.Y.; Nie, S.P.; Shen, M.Y.; Wang, Y.X.; Li, C. Isolation, chemical composition and antioxidant activities of a water-soluble polysaccharide from Cyclocarya paliurus (Batal.) lljinskaja. Food Chem. 2010, 119, 1626–1632. [Google Scholar] [CrossRef]
- Sun, X.; Liu, N.; Wu, Z.; Feng, Y.; Meng, X. Anti-tumor activity of a polysaccharide from blueberry. Molecules 2015, 20, 3841–3853. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Chen, L.Q.; Zhang, J.; Chen, F.Y. Anti-tumor and immunomodulatory activity of peptide fraction from the larvae of Musca domestica. J. Ethnopharmacol. 2014, 153, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.C.; Hsieh, C.C.; Lin, W.C. Characterization and immunomodulatory activity of rice hull polysaccharides. Carbohydr. Polym. 2015, 124, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.B.; Lv, Y.; Tian, L.M.; Zhao, Y. Composition and systemic immune activity of the polysaccharides from an herbal tea (Lysopus iucidus Trucz). J. Agric. Food Chem. 2010, 58, 6075–6080. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Suciu-Foca, N.; Manavalan, J.S.; Cortesini, R. Generation and function of antigen-specific suppressor and regulatory T cells. Transplant. Immunol. 2003, 11, 235–244. [Google Scholar] [CrossRef]
- Steensberg, A.; Toft, A.D.; Bruunsgaard, H.; Sandmand, M.; Halkjær-Kristensen, J.; Pedersen, B.K. Strenuous exercise decreases the percentage of type 1 T cells in the circulation. J. Appl. Physiol. 2001, 91, 1708–1712. [Google Scholar] [PubMed]
- Wong, K.H.; Lai, C.K.M.; Cheung, P.C.K. Immunomodulatory activities of mushroom sclerotial polysaccharides. Food Hydrocoll. 2011, 25, 150–158. [Google Scholar] [CrossRef]
- Ding, X.; Zhu, F.S.; Gao, S.G. Purification, antitumour and immunomodulatory activity of water-extractable and alkali-extractable polysaccharides from Solanum nigrum L. Food Chem. 2012, 131, 677–684. [Google Scholar] [CrossRef]
- Lovrić, J.; Mesić, M.; Macan, M.; Koprivanac, M.; Kelava, M.; Bradamante, V. Measurement of malondialdehyde (MDA) level in rat plasma after simvastatin treatment using two different analytical methods. Period. Biol. 2008, 110, 63–67. [Google Scholar]
- Carillon, J.; Rouanet, J.M.; Cristol, J.P.; Brion, R. Superoxide dismutase administration, a potential therapy against oxidative stress related diseases: Several routes of supplementation and proposal of an original mechanism of action. Pharm. Res. 2013, 30, 2718–2728. [Google Scholar] [CrossRef] [PubMed]
- Cichoski, A.J.; Rotta, R.B.; Scheuermann, G.; Cunha Junior, A.; Barin, J.S. Investigation of glutathione peroxidase activity in chicken meat under different experimental conditions. Ciênc. Tecnol. Aliment. 2012, 32, 661–667. [Google Scholar] [CrossRef]
- Rees, M.D.; Kennett, E.C.; Whitelock, J.M.; Davies, M.J. Oxidative damage to extracellular matrix and its role in human pathologies. Free Radic. Biol. Med. 2008, 44, 1973–2001. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K. Studies on free radicals, antioxidants and co-factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar] [PubMed]
- Xu, R.; Shang, N.; Li, P. In vitro and in vivo antioxidant activity of exopolysaccharide fractions from Bifidobacterium animalis RH. Anaerobe 2011, 17, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.L.; Gao, Q.; Wen, L.R.; Yang, B.; Tang, J.; You, L.J.; Zhao, M.M. Structural characterization of acid- and alkali-soluble polysaccharides in the fruiting body of Dictyophora indusiata and their immunomodulatory activities. Food Chem. 2012, 132, 739–743. [Google Scholar] [CrossRef]
- Dai, Z.Q.; Su, D.; Zhang, Y.; Sun, Y.; Hu, B.; Ye, H.; Jabbar, S.; Zeng, X.X. Immunomodulatory activity in vitro and in vivo of verbascose from mung beans (Phaseolus aureus). J. Agric. Food Chem. 2014, 62, 10727–10735. [Google Scholar] [CrossRef] [PubMed]
- Chalamaiah, M.; Hemalatha, R.; Jyothirmayi, T.; Diwan, P.V.; Bhaskarachary, K.; Vajreswari, A.; Kumar, R.R.; Kumar, B.D. Chemical composition and immunomodulatory effects of enzymatic protein hydrolysates from common carp (Cyprinus carpio) egg. Nutrition 2015, 31, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not Available.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose (mg/kg BW) | Spleen Index (mg/g) | Thymus Index (mg/g) | Phagocytic Index (α) |
|---|---|---|---|---|
| Normal control | -- | 6.23 ± 0.87 | 1.55 ± 0.07 | 5.212 ± 0.03 |
| Model control (Cy) | 80 | 4.41 ± 0.92 ** | 1.26 ± 0.09 ** | 3.983 ± 0.02 *** |
| GPMPP | 50 + Cy | 5.27 ± 1.69 | 1.28 ± 0.09 | 4.771 ± 0.04 ### |
| 150 + Cy | 6.14 ± 1.24 ## | 1.52 ± 0.18 ## | 5.096 ± 0.02 **### | |
| 250 + Cy | 6.45 ± 1.38 ### | 1.49 ± 0.20 ## | 5.458 ± 0.04 **### | |
| Positive control (LH) | 10 + Cy | 6.63 ± 1.26 | 1.57 ± 0.11 | 5.282 ± 0.04 ### |
| Group | Dose (mg/kg BW) | WBC (106/mL) | BMC (106/mL) | Serum Haemolysin (HC50) |
|---|---|---|---|---|
| Normal control | -- | 5.7 ± 0.8 | 4.1 ± 0.8 | 0.393 ± 0.030 |
| Model control (Cy) | 80 | 3.3 ± 0.7 * | 2.1 ± 0.3 ** | 0.259 ± 0.023 ** |
| GPMPP | 50 + Cy | 4.0 ± 0.8 ## | 2.5 ± 0.4 | 0.291 ± 0.018 # |
| 150 + Cy | 4.5 ± 0.9 ## | 3.6 ± 0.6 ### | 0.399 ± 0.013 ## | |
| 250 + Cy | 4.9 ± 1.0 ## | 3.9 ± 0.3 ## | 0.422 ± 0.010 ## | |
| Positive control (LH) | 10 + Cy | 5.6 ± 0.5 ## | 4.0 ± 0.9 ## | 0.426 ± 0.013 ## |
| Group | Dose (mg/kg BW) | CD4+ (%) | CD8+ (%) | CD4+/CD8+ |
|---|---|---|---|---|
| Normal control | -- | 50.10 ± 3.53 | 34.57 ± 2.69 | 1.45 ± 0.12 |
| Model control (Cy) | 80 | 44.44 ± 3.47 * | 38.68 ± 3.40 | 1.15 ± 0.03 ** |
| GPMPP | 50 + Cy | 45.69 ± 2.67 | 30.19 ± 3.56 ### | 1.38 ± 0.1 # |
| 150 + Cy | 55.07 ± 4.8 *# | 34.97 ± 3.41 ## | 1.58 ± 0.07 *## | |
| 250 + Cy | 51.80 ± 1.49 *# | 38.45 ± 4.45 # | 1.45 ± 0.1 *# | |
| Positive control (LH) | 10 + Cy | 42.58 ± 4.23 | 27.36 ± 5.59 | 1.58 ± 0.17## |
| Group | Dose (mg/kg BW) | IL-2 (pg/mL) | |
|---|---|---|---|
| in Sera | in Spleen | ||
| Normal control | -- | 0.361 ± 0.014 | 0.278 ± 0.012 |
| Model control (Cy) | 80 | 0.334 ± 0.034 | 0.261 ± 0.015 |
| GPMPP | 50 + Cy | 0.427 ± 0.034 *## | 0.383 ± 0.021 ***### |
| 150 + Cy | 0.450 ± 0.018 ***### | 0.376 ± 0.026 ***### | |
| 250 + Cy | 0.561 ± 0.025 ***### | 0.487 ± 0.031 ***### | |
| Positive control (LH) | 10 + Cy | 0.523 ± 0.027 ***### | 0.463 ± 0.017 ***### |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, X.; Chao, Y.; Zhang, Y.; Lu, C.; Xu, C.; Niu, W. Immunomodulatory and Antioxidant Effects of Polysaccharides from Gynostemma pentaphyllum Makino in Immunosuppressed Mice. Molecules 2016, 21, 1085. https://doi.org/10.3390/molecules21081085
Shang X, Chao Y, Zhang Y, Lu C, Xu C, Niu W. Immunomodulatory and Antioxidant Effects of Polysaccharides from Gynostemma pentaphyllum Makino in Immunosuppressed Mice. Molecules. 2016; 21(8):1085. https://doi.org/10.3390/molecules21081085
Chicago/Turabian StyleShang, Xiaoya, Yu Chao, Yuan Zhang, Chengyuan Lu, Chunlan Xu, and Weining Niu. 2016. "Immunomodulatory and Antioxidant Effects of Polysaccharides from Gynostemma pentaphyllum Makino in Immunosuppressed Mice" Molecules 21, no. 8: 1085. https://doi.org/10.3390/molecules21081085
APA StyleShang, X., Chao, Y., Zhang, Y., Lu, C., Xu, C., & Niu, W. (2016). Immunomodulatory and Antioxidant Effects of Polysaccharides from Gynostemma pentaphyllum Makino in Immunosuppressed Mice. Molecules, 21(8), 1085. https://doi.org/10.3390/molecules21081085