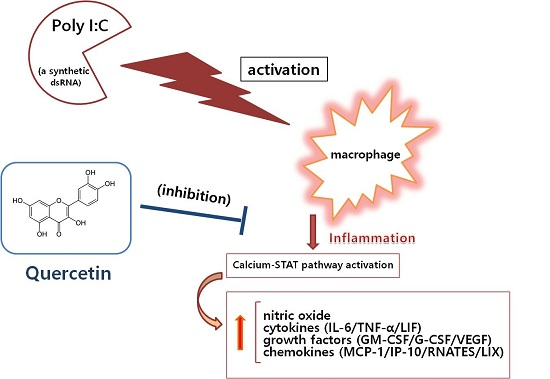
Anti‐Inflammatory Effect of Quercetin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic‐Polycytidylic Acid

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
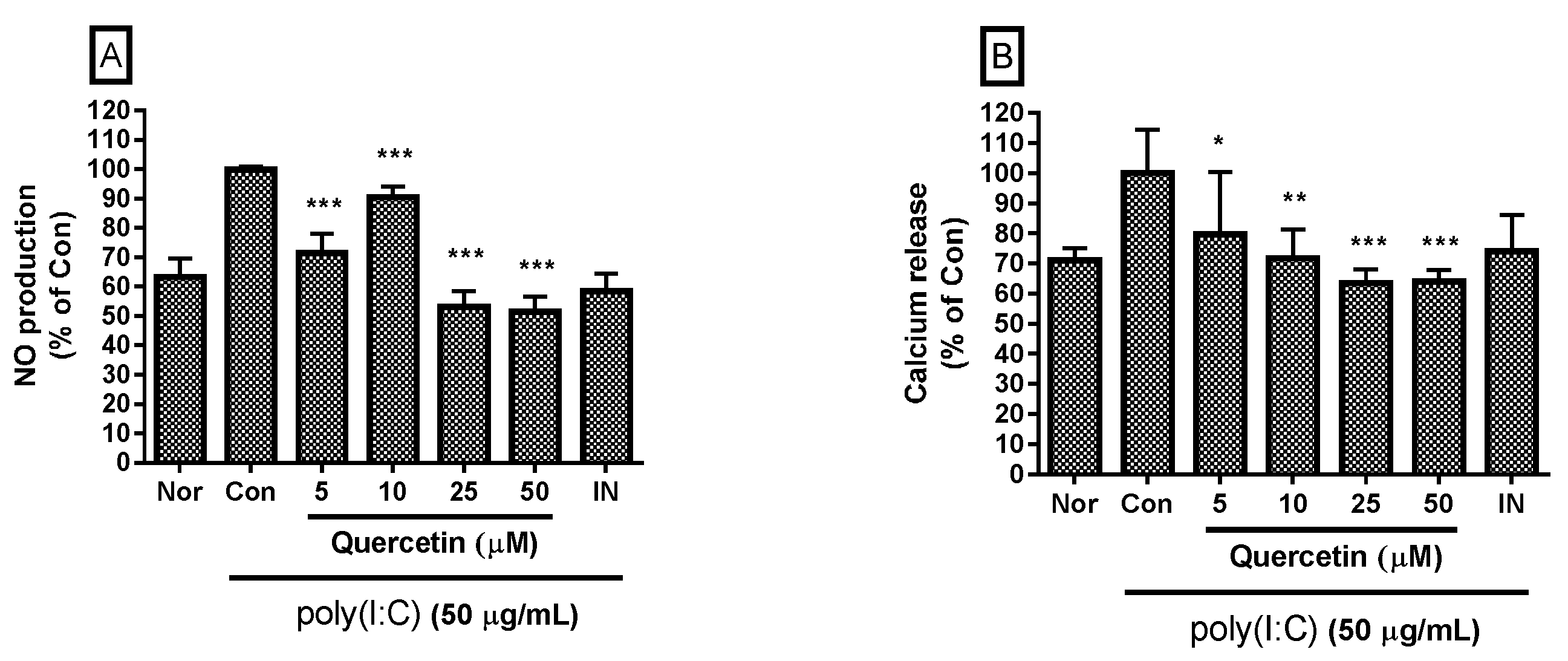
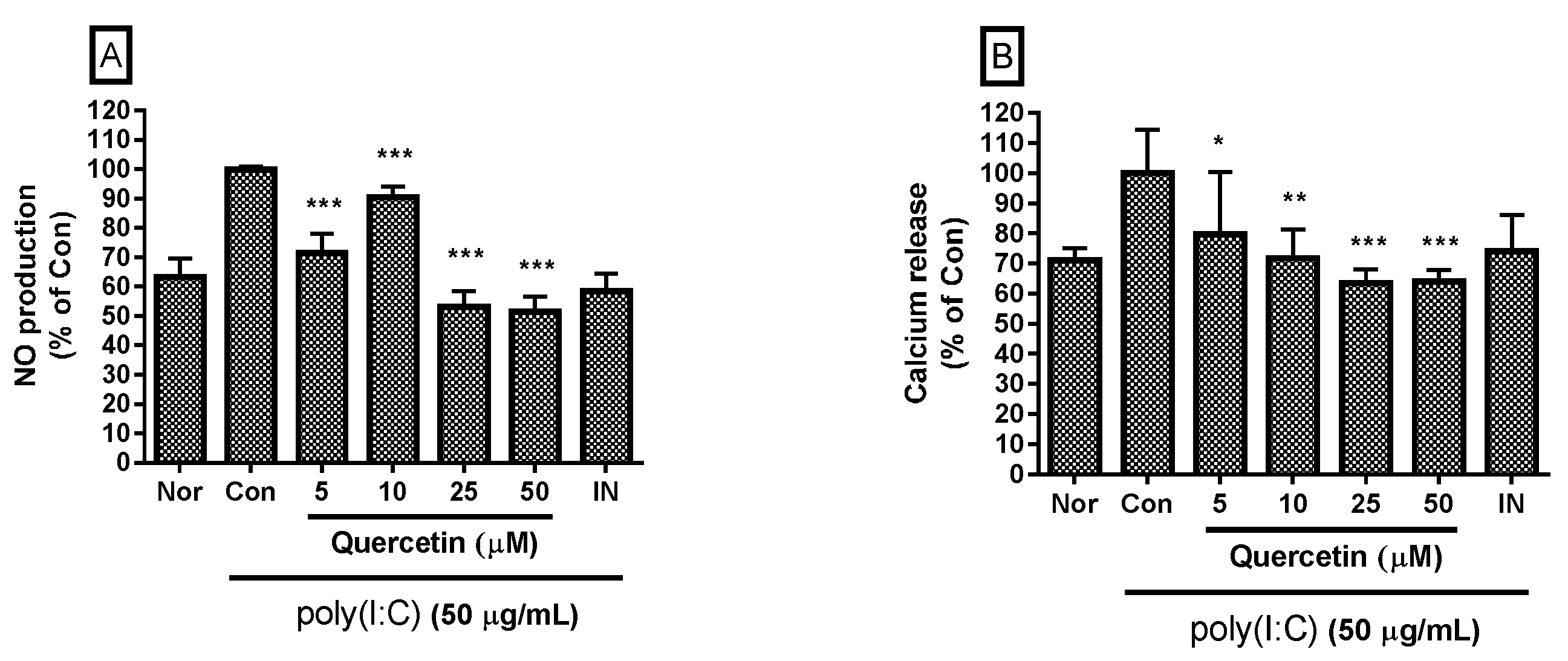
2.1. Effect of Quercetin on NO Production and Intracellular Calcium Release
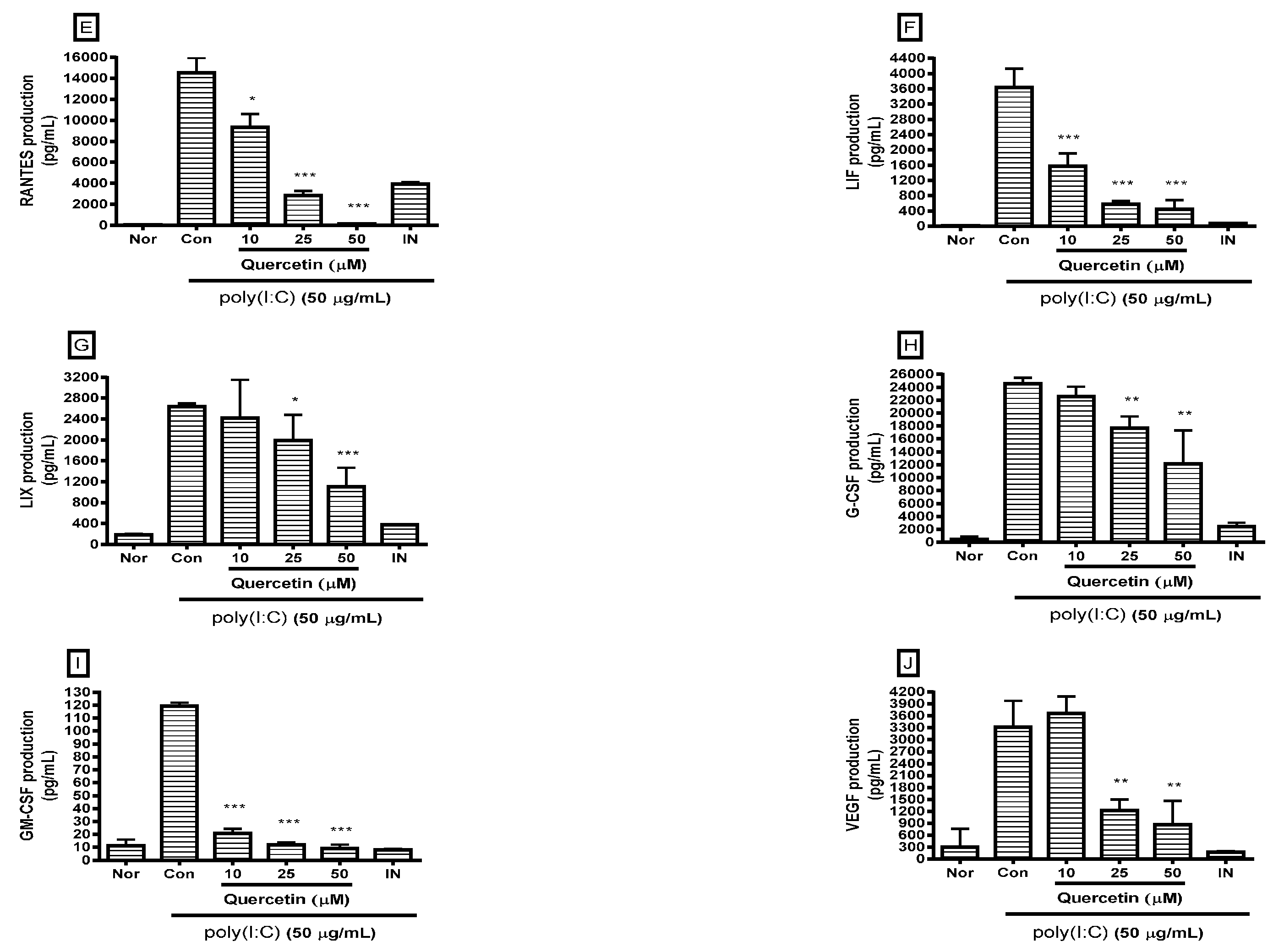
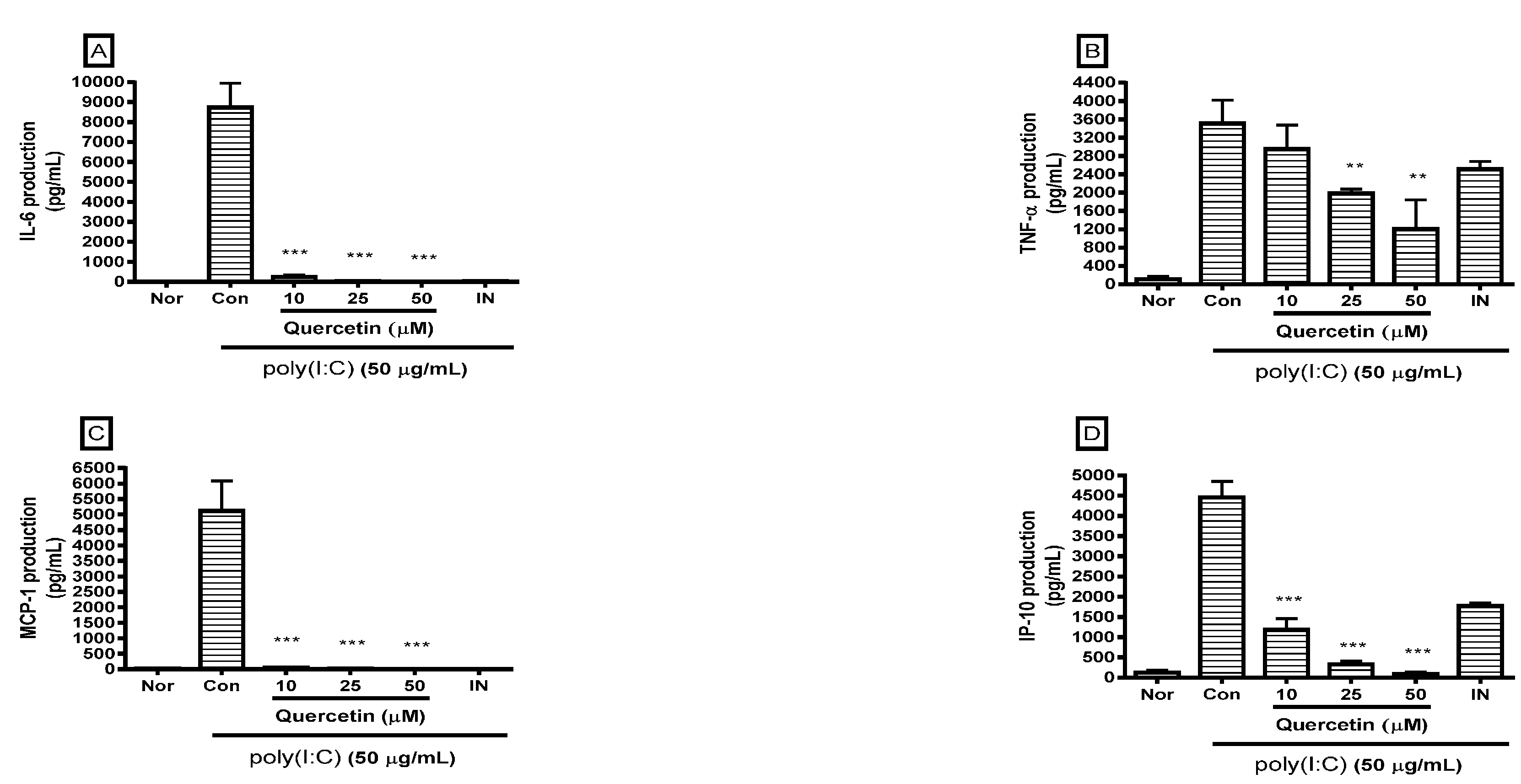
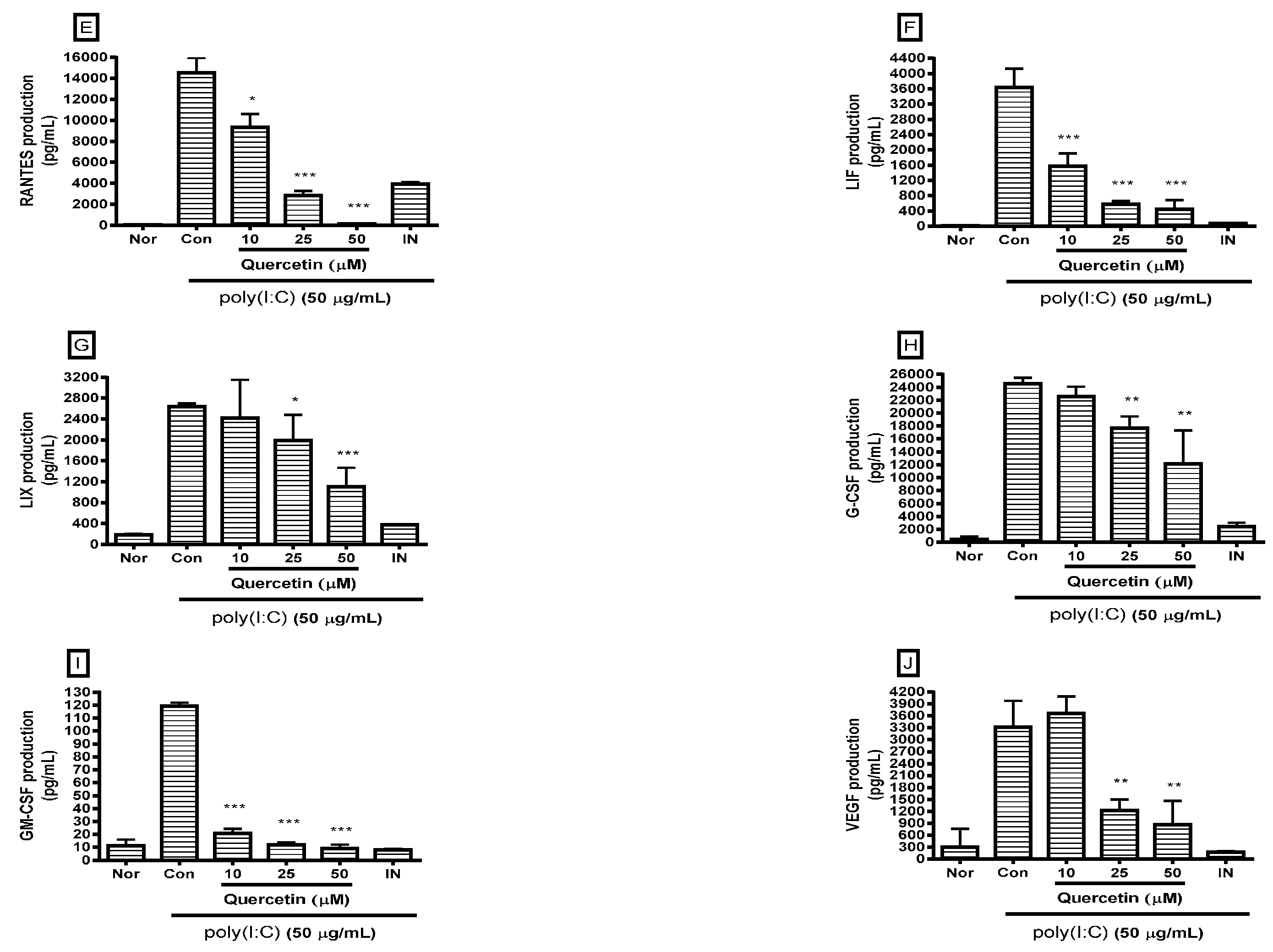
2.2. Effect of Quercetin on Cytokine Production
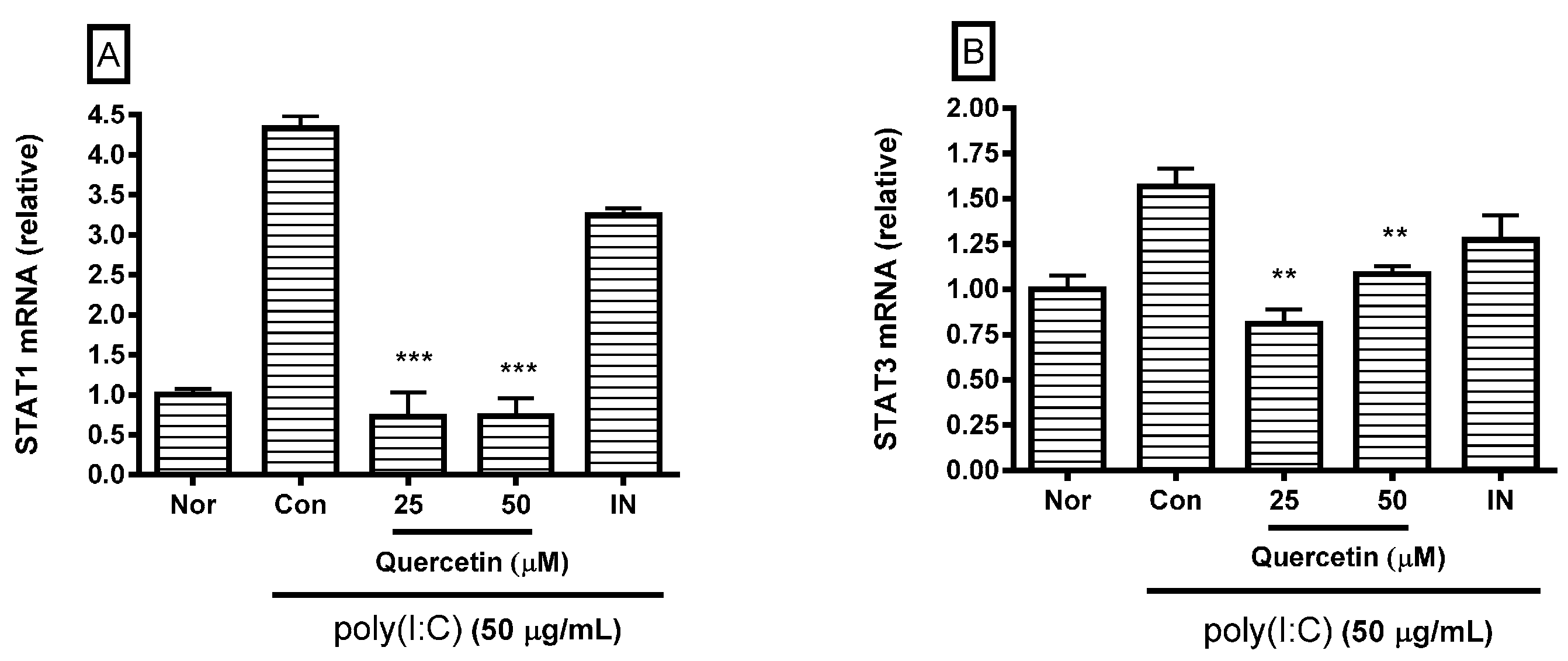
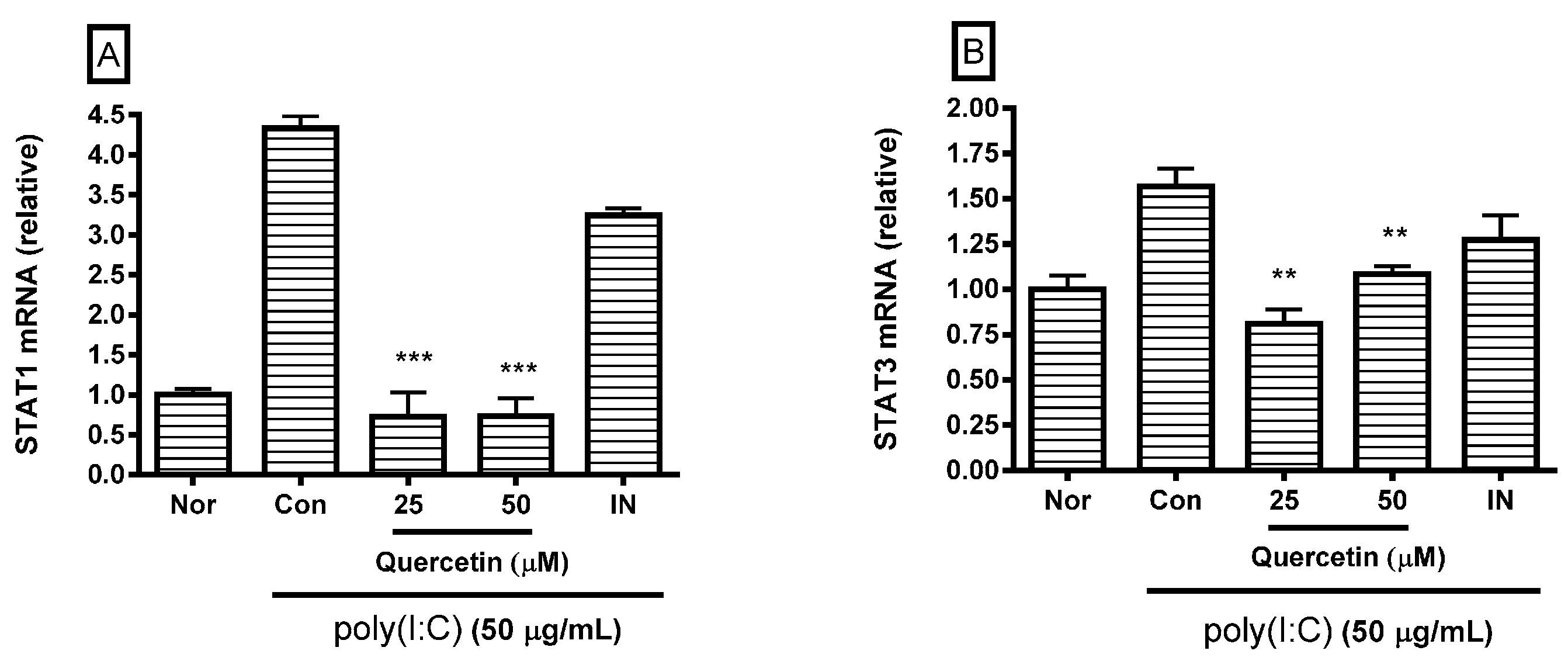
2.3. Effect of Quercetin on STAT1 and STAT3 mRNA Expression
3. Discussion
4. Materials and Methods
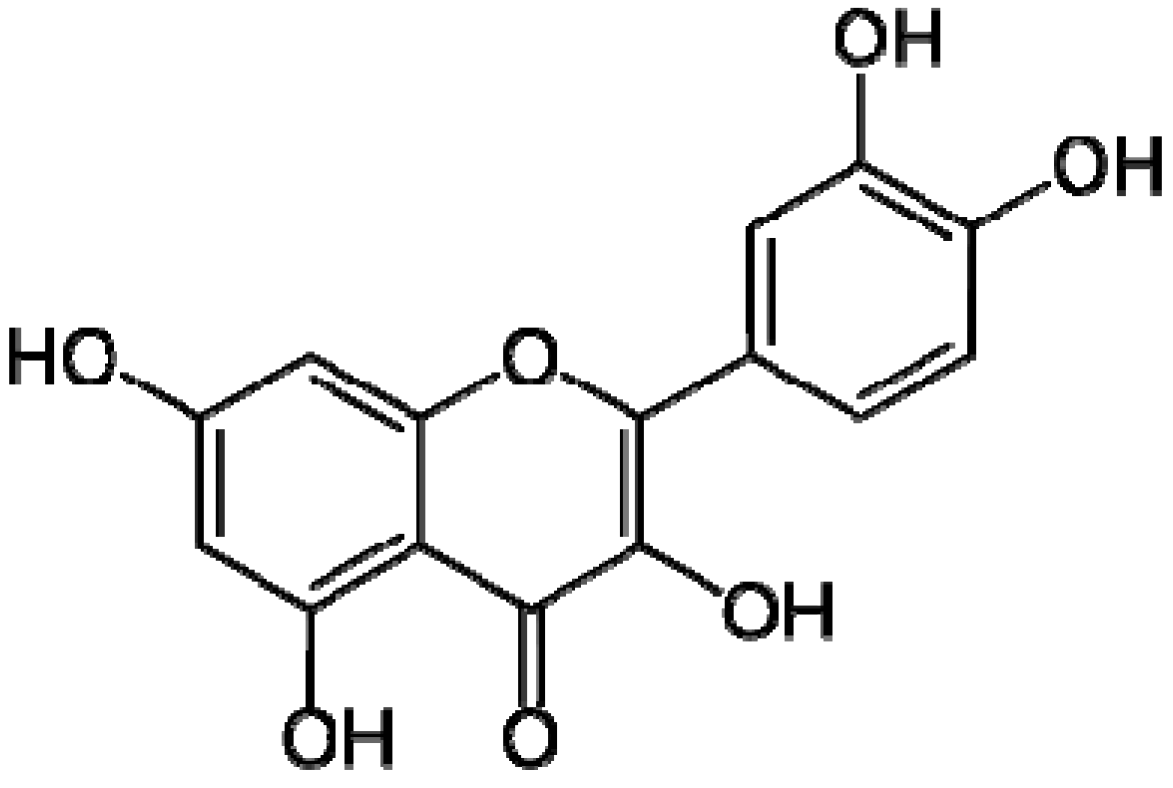
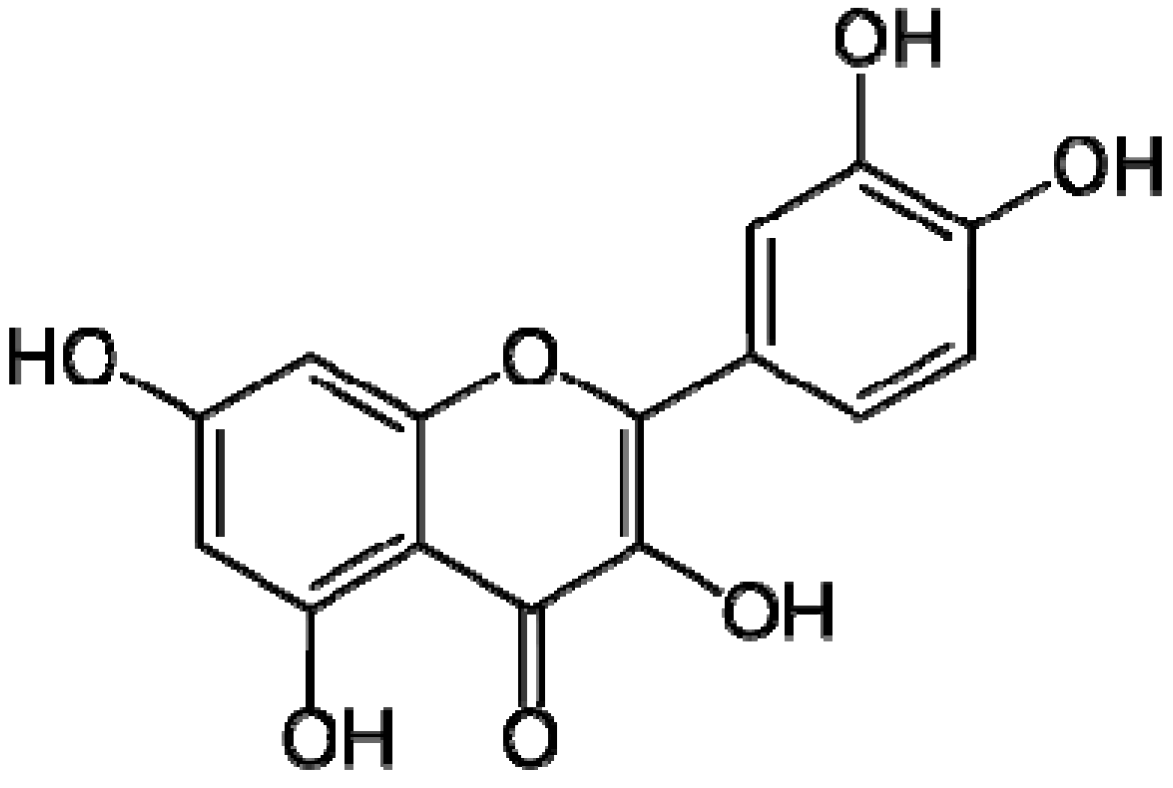
4.1. Materials
4.2. Quantification of NO Production and Intracellular Calcium Release
4.3. Multiplex Bead-Based Cytokine Assay
4.4. RNA Isolation and Real Time RT-PCR Analysis
4.5. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| NO | nitric oxide |
| dsRNA | double-stranded RNA |
| TLR | Toll like receptor |
| poly(I:C) | polyinosinic-polycytidylic acid |
| LPS | lipopolysaccharide |
| IL | interleukin |
| TNF | tumor necrosis factor |
| MCP | monocyte chemotactic protein |
| IP-10 | interferon inducible protein-10 |
| LIX | lipopolysaccharide-induced CXC chemokine |
| LIF | leukemia inhibitory factor |
| G-CSF | granulocyte colony-stimulating factor |
| GM-CSF | granulocyte macrophage colony-stimulating factor |
| VEGF | vascular endothelial growth factor |
| STAT1 | signal transducers and activators of transcription 1 |
| STAT3 | signal transducers and activators of transcription 3 |
| Nor | normal |
| Con | control |
| ER | endoplasmic reticulum |
References
- Bartemes, K.R.; Iijima, K.; Kobayashi, T.; Kephart, G.M.; McKenzie, A.N.; Kita, H. IL-33-Responsive Lineage- CD25+ CD44(Hi) Lymphoid Cells Mediate Innate Type 2 Immunity and Allergic Inflammation in the Lungs. J. Immunol. 2012, 188, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Gilroy, D.W. Eicosanoids and the Endogenous Control of Acute Inflammatory Resolution. Int. J. Biochem. Cell Biol. 2010, 42, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Legatzki, A.; Rösler, B.; von Mutius, E. Microbiome Diversity and Asthma and Allergy Risk. Curr. Allergy Asthma Rep. 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Eo, H.J.; Park, J.H.; Park, G.H.; Lee, M.H.; Lee, J.R.; Koo, J.S.; Jeong, J.B. Anti-Inflammatory and Anti-Cancer Activity of Mulberry (Morus Alba L.) Root Bark. BMC Complement. Altern. Med. 2014, 14, 200. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Ma, A.; Lipsky, P. Cytokines and Autoimmunity. Nat. Rev. Immunol. 2002, 2, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Challa, S.; Moquin, D.; Genga, R.; Ray, T.D.; Guildford, M.; Chan, F.K. Phosphorylation-Driven Assembly of the RIP1-RIP3 Complex Regulates Programmed Necrosis and Virus-Induced Inflammation. Cell 2009, 137, 1112–1123. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Ruksznis, C.; Wang, Y.; Leifer, C.A. Antimicrobial Peptides Inhibit Polyinosinic-Polycytidylic Acid-Induced Immune Responses. J. Immunol. 2011, 187, 5653–5659. [Google Scholar] [CrossRef] [PubMed]
- Arsenault, R.J.; Kogut, M.H.; He, H. Combined CpG and Poly I:C Stimulation of Monocytes Results in Unique Signaling Activation Not Observed with the Individual Ligands. Cell Signal. 2013, 25, 2246–2254. [Google Scholar] [CrossRef] [PubMed]
- Anjaneyulu, M.; Chopra, K. Quercetin, an anti-oxidant Bioflavonoid, Attenuates Diabetic Nephropathy in Rats. Clin. Exp. Pharmacol. Phys. 2004, 31, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.R.; Lin, J.Y. Quercetin Intraperitoneal Administration Ameliorates Lipopolysaccharide-Induced Systemic Inflammation in Mice. Life Sci. 2015, 137, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gardi, C.; Bauerova, K.; Stringa, B.; Kuncirova, V.; Slovak, L.; Ponist, S.; Drafi, F.; Bezakova, L.; Tedesco, I.; Acquaviva, A.; et al. Quercetin Reduced Inflammation and Increased Antioxidant Defense in Rat Adjuvant Arthritis. Arch. Biochem. Biophys. 2015, 583, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Guazelli, C.F.; Fattori, V.; Colombo, B.B.; Georgetti, S.R.; Vicentini, F.T.; Casagrande, R.; Baracat, M.M.; Verri, W.A., Jr. Quercetin-Loaded Microcapsules Ameliorate Experimental Colitis in Mice by Anti-Inflammatory and Antioxidant Mechanisms. J. Nat. Prod. 2013, 76, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Chiow, K.H.; Phoon, M.C.; Putti, T.; Tan, B.K.; Chow, V.T. Evaluation of Antiviral Activities of Houttuynia Cordata Thunb. Extract, Quercetin, Quercetrin and Cinanserin on Murine Coronavirus and Dengue Virus Infection. Asian Pac. J. Trop. Med. 2016, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Li, R.; Li, X.; He, J.; Jiang, S.; Liu, S.; Yang, J. Quercetin as an Antiviral Agent Inhibits Influenza A Virus (IAV) Entry. Viruses 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Gadient, R.A.; Patterson, P.H. Leukemia Inhibitory Factor, Interleukin 6, and Other Cytokines using the GP130 Transducing Receptor: Roles in Inflammation and Injury. Stem Cells 1999, 17, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A. A New Paradigm: Innate Immune Sensing of Viruses Via the Unfolded Protein Response. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Mahima; Ingle, A.M.; Verma, A.K.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Deb, R.; Rajagunalan, S.; Rathore, R.; Dhama, K. Immunomodulators in Day to Day Life: A Review. Pak. J. Biol. Sci. 2013, 16, 826–843. [Google Scholar] [PubMed]
- Cooper, P.R.; Lamb, R.; Day, N.D.; Branigan, P.J.; Kajekar, R.; San Mateo, L.; Hornby, P.J.; Panettieri, R.A., Jr. TLR3 Activation Stimulates Cytokine Secretion without Altering Agonist-Induced Human Small Airway Contraction Or Relaxation. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 297, L530–L537. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Kim, H.J.; Ramirez, M.; Salameh, S.; Ma, X. The Septic Shock-Associated IL-10 –1082 A > G Polymorphism Mediates Allele-Specific Transcription Via Poly(ADP-Ribose) Polymerase 1 in Macrophages Engulfing Apoptotic Cells. J. Immunol. 2010, 184, 3718–3724. [Google Scholar] [CrossRef] [PubMed]
- Cammer, M.; Cox, D. Chemotactic Responses by Macrophages to a Directional Source of a Cytokine Delivered by a Micropipette. Methods Mol. Biol. 2014, 1172, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.A.; Morris, I.R.; Berton, M.T. Phosphatidylinositol 3-Kinase Activation Attenuates the TLR2-Mediated Macrophage Proinflammatory Cytokine Response to Francisella Tularensis Live Vaccine Strain. J. Immunol. 2010, 185, 7562–7572. [Google Scholar] [CrossRef] [PubMed]
- Oates, J.C. The Biology of Reactive Intermediates in Systemic Lupus Erythematosus. Autoimmunity 2010, 43, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Karpuzoglu, E.; Ahmed, S.A. Estrogen Regulation of Nitric Oxide and Inducible Nitric Oxide Synthase (iNOS) in Immune Cells: Implications for Immunity, Autoimmune Diseases, and Apoptosis. Nitric Oxide 2006, 15, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, K.; Su, Z.; Boyanapalli, S.S.; Barman, D.N.; Huang, M.; Chen, L.; Magesh, S.; Hu, L.; Kong, A.T. In Vitro and in Vivo Anti-Inflammatory Effects of a Novel 4, 6-Bis ((E)-4-Hydroxy-3-Methoxystyryl)-1-Phenethylpyrimidine-2 (1H)-Thione. Chem. Res. Toxicol. 2013, 27, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, G.; Kutuzov, M.A.; Ridge, K.M. The Inflammasome in Lung Diseases. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 303, L627–L633. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Murakami, T.; Suzuki, K.; Tamura, H.; Kuwahara-Arai, K.; Iba, T.; Nagaoka, I. Antimicrobial Cathelicidin Peptide LL-37 Inhibits the LPS/ATP-Induced Pyroptosis of Macrophages by Dual Mechanism. PLoS ONE 2014, 9, e85765. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, L.; Tracey, K.J. The “Cytokine Profile”: A Code for Sepsis. Trends Mol. Med. 2005, 11, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Sessler, C.N.; Shepherd, W. New Concepts in Sepsis. Curr. Opin. Crit. Care 2002, 8, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Wareing, M.D.; Lyon, A.B.; Lu, B.; Gerard, C.; Sarawar, S.R. Chemokine Expression during the Development and Resolution of a Pulmonary Leukocyte Response to Influenza A Virus Infection in Mice. J. Leukoc. Biol. 2004, 76, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Timmins, J.M.; Ozcan, L.; Seimon, T.A.; Li, G.; Malagelada, C.; Backs, J.; Backs, T.; Bassel-Duby, R.; Olson, E.N.; Anderson, M.E.; et al. Calcium/calmodulin-Dependent Protein Kinase II Links ER Stress with Fas and Mitochondrial Apoptosis Pathways. J. Clin. Investig. 2009, 119, 2925–2941. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Park, W. Anti-Inflammatory Effect of Myristicin on RAW 264.7 Macrophages Stimulated with Polyinosinic-Polycytidylic Acid. Molecules 2011, 16, 7132–7142. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Park, W.; Yi, D.K. Immunostimulatory Effects of Gold Nanorod and Silica-Coated Gold Nanorod on RAW 264.7 Mouse Macrophages. Toxicol. Lett. 2012, 209, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Lee, Y.; Park, S.K.; Kim, H.; Bae, H.; Kim, H.M.; Ko, S.; Choi, H.Y.; Oh, M.S.; Park, W. Anti-Inflammatory Effects of Scutellaria Baicalensis Water Extract on LPS-Activated RAW 264.7 Macrophages. J. Ethnopharmacol. 2009, 125, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Yuk, S.S.; Lim, E.M.; Lee, J.Y.; Lee, Y.J.; Kim, Y.S.; Lee, T.H.; Park, S.K.; Bae, H.; Kim, H.M.; Ko, S.G.; et al. Antiinflammatory Effects of Epimedium Brevicornum Water Extract on Lipopolysaccharide-Activated RAW264.7 Macrophages. Phytother. Res. 2010, 24, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Park, W. Anti-Inflammatory Effect of Wogonin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic-Polycytidylic Acid. Molecules 2015, 20, 6888–6900. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Quercetin samples are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Park, W. Anti‐Inflammatory Effect of Quercetin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic‐Polycytidylic Acid. Molecules 2016, 21, 450. https://doi.org/10.3390/molecules21040450
Kim Y, Park W. Anti‐Inflammatory Effect of Quercetin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic‐Polycytidylic Acid. Molecules. 2016; 21(4):450. https://doi.org/10.3390/molecules21040450
Chicago/Turabian StyleKim, Young‐Jin, and Wansu Park. 2016. "Anti‐Inflammatory Effect of Quercetin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic‐Polycytidylic Acid" Molecules 21, no. 4: 450. https://doi.org/10.3390/molecules21040450
APA StyleKim, Y., & Park, W. (2016). Anti‐Inflammatory Effect of Quercetin on RAW 264.7 Mouse Macrophages Induced with Polyinosinic‐Polycytidylic Acid. Molecules, 21(4), 450. https://doi.org/10.3390/molecules21040450