
Mechanisms of Body Weight Reduction by Black Tea Polyphenols

{kind=link}
Abstract
:1. Introduction
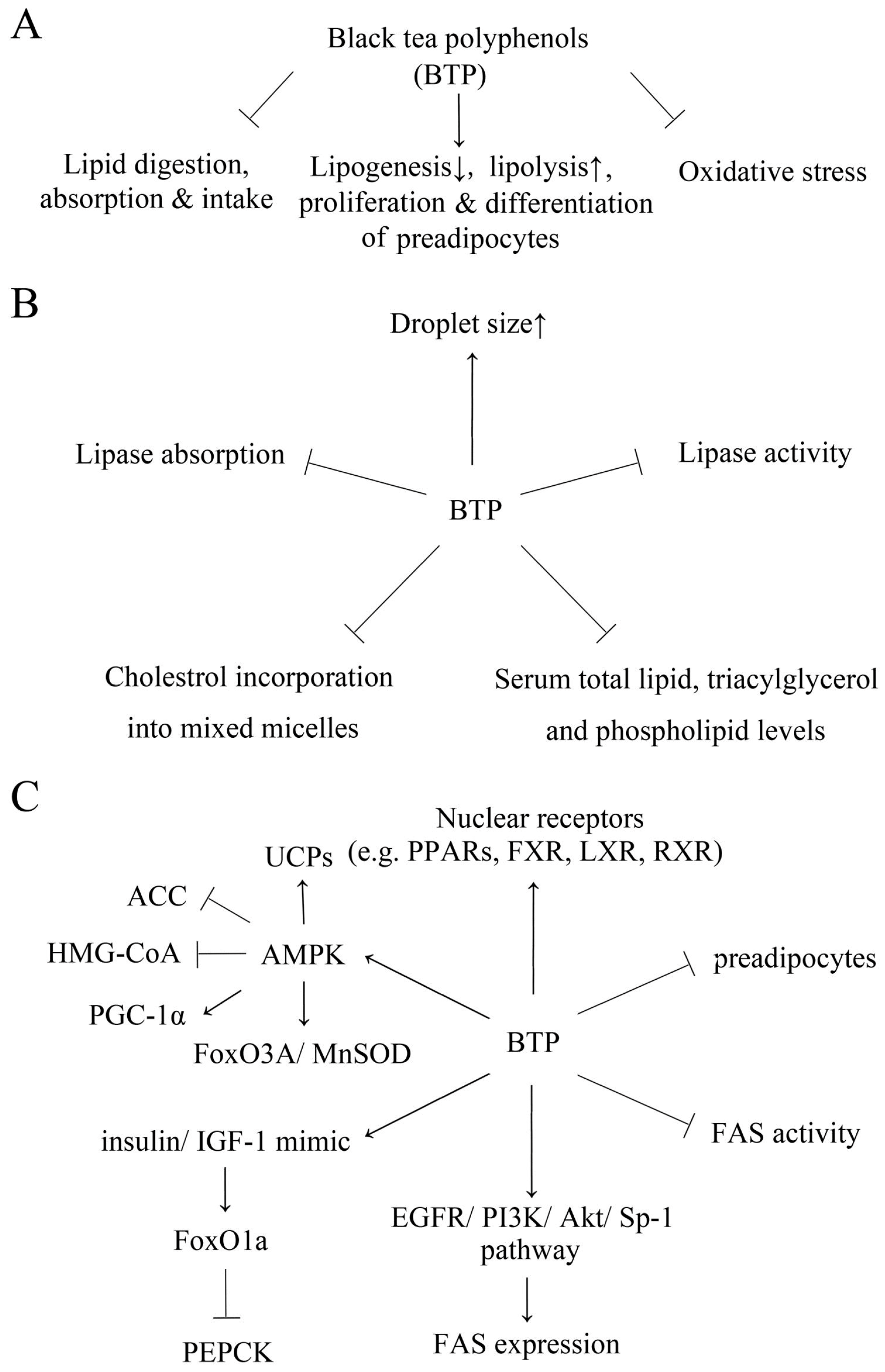
2. The Effect of Black Tea Polyphenols on Lipid Digestion, Absorption and Intake
3. The Effect of Black Tea Polyphenols on Lipid Metabolism and Accumulation
4. The Effect of Black Tea Polyphenols on Saccharide Digestion and Absorption
5. The Anti-Oxidant Effect of Black Tea Polyphenols
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alwan, A. Global Status Report on Noncommunicable Diseases 2010; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M.; Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 2002, 393–403. [Google Scholar]
- Kannel, W.B.; Cupples, L.A.; Ramaswami, R.; Stokes, J.; Kreger, B.E.; Higgins, M. Regional obesity and risk of cardiovascular disease; the framingham study. J. Clin. Epidemiol. 1991, 44, 183–190. [Google Scholar] [CrossRef]
- Marmot, M.; Atinmo, T.; Byers, T.; Chen, J.; Hirohata, T.; Jackson, A.; James, W.; Kolonel, L.; Kumanyika, S.; Leitzmann, C. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; ICR: Washington, DC, USA, 2007. [Google Scholar]
- Bajari, T.M.; Nimpf, J.; Schneider, W.J. Role of leptin in reproduction. Curr. Opin. Lipidol. 2004, 15, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.A.; El-Gharib, N.E. Obesity and clinical riskiness relationship: Therapeutic management by dietary antioxidant supplementation—A review. Appl. Biochem. Biotechnol. 2015, 176, 647–669. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.C.; Chen, H.Y. Antioxidant activity of various tea extracts in relation to their antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Rietveld, A.; Wiseman, S. Antioxidant effects of tea: Evidence from human clinical trials. J. Nutr. 2003, 133, 3285s–3292s. [Google Scholar] [PubMed]
- Fatima, M.; Rizvi, S.I. Anti oxidative effect of black tea theaflavin on erythrocytes subjected to oxidative stress. Natl. Acad. Sci. Lett. 2015, 38, 25–28. [Google Scholar] [CrossRef]
- Wang, Y.C.; Bachrach, U. The specific anti-cancer activity of green tea (−)-epigallocatechin-3-gallate (EGCG). Amino Acids 2002, 22, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Wang, Y.; Zhang, Y.; Wan, X.; Li, J.; Liu, K.; Wang, F.; Liu, K.; Liu, Q.; Yang, C.; et al. Anti-cancer activities of tea epigallocatechin-3-gallate in breast cancer patients under radiotherapy. Curr. Mol. Med. 2012, 12, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Isemura, M.; Saeki, K.; Kimura, T.; Hayakawa, S.; Minami, T.; Sazuka, M. Tea catechins and related polyphenols as anti-cancer agents. Biofactors 2000, 13, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Min, W.K.; Chun, S.; Lee, Y.W.; Park, H.; Lee, D.H.; Lee, Y.K.; Son, J.E. Long-term effects of green tea ingestion on atherosclerotic biological markers in smokers. Clin. Biochem. 2005, 38, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Mandel, S.A.; Amit, T.; Weinreb, O.; Reznichenko, L.; Youdim, M.B.H. Simultaneous manipulation of multiple brain targets by green tea catechins: A potential neuroprotective strategy for Alzheimer and Parkinson diseases. CNS Neurosci. Ther. 2008, 14, 352–365. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, O.; Mandel, S.; Amit, T.; Youdim, M.B.H. Neurological mechanisms of green tea polyphenols in Alzheimer’s and Parkinson’s diseases. J. Nutr. Biochem. 2004, 15, 506–516. [Google Scholar] [CrossRef] [PubMed]
- Anandhan, A.; Tamilselvam, K.; Radhiga, T.; Rao, S.; Essa, M.M.; Manivasagam, T. Theaflavin, a black tea polyphenol, protects nigral dopaminergic neurons against chronic mptp/probenecid induced Parkinson’s disease. Brain Res. 2012, 1433, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Guo, Y.; Liu, R.; Wang, K.; Zhang, M. Black tea polyphenols and polysaccharides improve body composition, increase fecal fatty acid, and regulate fat metabolism in high-fat diet-induced obese rats. Food Funct. 2016, 7, 2469–2478. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ichitani, M.; Suzuki, Y.; Unno, T.; Sugawara, T.; Yamahira, T.; Kato, M.; Takihara, T.; Sagesaka, Y.; Kakuda, T.; et al. Black-tea polyphenols suppress postprandial hypertriacylglycerolemia by suppressing lymphatic transport of dietary fat in rats. J. Agric. Food Chem. 2009, 57, 7131–7136. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Xu, Y.; Mei, X.; Meng, Q.; Gao, Y.; Li, B.; Tu, Y. Antiobesity and lipid lowering effects of theaflavins on high-fat diet induced obese rats. J. Funct. Foods 2013, 5, 1142–1150. [Google Scholar] [CrossRef]
- Ashigai, H.; Taniguchi, Y.; Suzuki, M.; Ikeshima, E.; Kanaya, T.; Zembutsu, K.; Tomita, S.; Miyake, M.; Fukuhara, I. Fecal lipid excretion after consumption of a black tea polyphenol containing beverage. Biol. Pharm. Bull. 2016, 39, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Furne, J.K.; Levitt, M.D. An extract of black, green, and mulberry teas causes malabsorption of carbohydrate but not of triacylglycerol in healthy volunteers. Am. J. Clin. Nutr. 2006, 84, 551–555. [Google Scholar] [PubMed]
- Shishikura, Y.; Khokhar, S.; Murray, B.S. Effects of tea polyphenols on emulsification of olive oil in a small intestine model system. J. Agric. Food Chem. 2006, 54, 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, M.A.; Mulder, T.P.; Molhuizen, H.O. Theaflavins from black tea, especially theaflavin-3-gallate, reduce the incorporation of cholesterol into mixed micelles. J. Agric. Food Chem. 2008, 56, 12031–12036. [Google Scholar] [CrossRef] [PubMed]
- Nakai, M.; Fukui, Y.; Asami, S.; Toyoda-Ono, Y.; Iwashita, T.; Shibata, H.; Mitsunaga, T.; Hashimoto, F.; Kiso, Y. Inhibitory effects of oolong tea polyphenols on pancreatic lipase in vitro. J. Agric. Food Chem. 2005, 53, 4593–4598. [Google Scholar] [CrossRef] [PubMed]
- Hamdaoui, M.H.; Snoussi, C.; Dhaouadi, K.; Fattouch, S.; Ducroc, R.; Le Gall, M.; Bado, A. Tea decoctions prevent body weight gain in rats fed high-fat diet; black tea being more efficient than green tea. J. Nutr. Intermed. Metab. 2016, 6, 33–40. [Google Scholar] [CrossRef]
- Uchiyama, S.; Taniguchi, Y.; Saka, A.; Yoshida, A.; Yajima, H. Prevention of diet-induced obesity by dietary black tea polyphenols extract in vitro and in vivo. Nutrition 2011, 27, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, G.; Nadia, M.; El-Ghffar, E.A.A. Modulatory effects of black v. Green tea aqueous extract on hyperglycaemia, hyperlipidaemia and liver dysfunction in diabetic and obese rat models. Br. J. Nutr. 2009, 102, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, M.; Takemoto, H.; Saijo, R. Theaflavin synthesized in a selective, domino-type, one-pot enzymatic biotransformation method with camellia sinensis cell culture inhibits weight gain and fat accumulation to high-fat diet-induced obese mice. Biol. Pharm. Bull. 2016, 39, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.-T.; Wang, X.; Wu, X.-D.; Tian, W.-X. Keemun black tea extract contains potent fatty acid synthase inhibitors and reduces food intake and body weight of rats via oral administration. J. Enzym. Inhib. Med. Chem. 2005, 20, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Bezzina, R.; Hinch, E.; Lewandowski, P.A.; Cameron-Smith, D.; Mathai, M.L.; Jois, M.; Sinclair, A.J.; Begg, D.P.; Wark, J.D. Green tea, black tea, and epigallocatechin modify body composition, improve glucose tolerance, and differentially alter metabolic gene expression in rats fed a high-fat diet. Nutr. Res. 2009, 29, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Indias, I.; Tinahones, F.J. Impaired adipose tissue expandability and lipogenic capacities as ones of the main causes of metabolic disorders. J. Diabetes Res. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.R.; Anton, S.; Melville, L.; Houston, N.P.; Dayal, S.; McDougall, G.J.; Stewart, D.; Rena, G. Black tea polyphenols mimic insulin/insulin-like growth factor-1 signalling to the longevity factor foxo1a. Aging Cell 2008, 7, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Ramirez, B.; Rotellar, F.; Pastor, C.; Silva, C.; Rodríguez, A.; Gil, M.J.; Cienfuegos, J.A.; Frühbeck, G. Proinflammatory cytokines in obesity: Impact of type 2 diabetes mellitus and gastric bypass. Obes. Surg. 2007, 17, 1464–1474. [Google Scholar] [CrossRef] [PubMed]
- Breslin, W.L.; Johnston, C.A.; Strohacker, K.; Carpenter, K.C.; Davidson, T.R.; Moreno, J.P.; Foreyt, J.P.; McFarlin, B.K. Obese mexican american children have elevated mcp-1, tnf-α, monocyte concentration, and dyslipidemia. Pediatrics 2012, 129, e1180–e1186. [Google Scholar] [CrossRef] [PubMed]
- Higa, J.K.; Liu, W.; Berry, M.J.; Panee, J. Supplement of bamboo extract lowers serum monocyte chemoattractant protein-1 concentration in mice fed a diet containing a high level of saturated fat. Br. J. Nutr. 2011, 106, 1810–1813. [Google Scholar] [CrossRef] [PubMed]
- Bose, T.; Alvarenga, J.C.L.; Tejero, M.E.; Voruganti, V.S.; Proffitt, J.M.; Freeland-Graves, J.H.; Cole, S.A.; Comuzzie, A.G. Association of monocyte chemoattractant protein-1 with adipocyte number, insulin resistance and liver function markers. J. Med. Primatol. 2009, 38, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Lee, S.-H.; Jung, E.S.; Kim, J.-S.; Shim, C.Y.; Ko, Y.-G.; Choi, D.; Jang, Y.; Chung, N.; Ha, J.-W. Visceral adiposity and the severity of coronary artery disease in middle-aged subjects with normal waist circumference and its relation with lipocalin-2 and mcp-1. Atherosclerosis 2010, 213, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Heber, D.; Zhang, Y.; Yang, J.; Ma, J.E.; Henning, S.M.; Li, Z. Green tea, black tea, and oolong tea polyphenols reduce visceral fat and inflammation in mice fed high-fat, high-sucrose obesogenic diets. J. Nutr. 2014, 144, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.-T.; Zhao, J.; Wu, X.-D.; Tian, W.-X. Inhibition of two type of β-ketoacyl reductase by black tea and theaflavins. J. Univ. Chin. Acad. Sci. 2005, 22, 701–706. [Google Scholar]
- Lin, J.K.; Lin-Shiau, S.Y. Mechanisms of hypolipidemic and anti-obesity effects of tea and tea polyphenols. Mol. Nutr. Food Res. 2006, 50, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Daval, M.; Diot-Dupuy, F.; Bazin, R.; Hainault, I.; Viollet, B.; Vaulont, S.; Hajduch, E.; Ferré, P.; Foufelle, F. Anti-lipolytic action of amp-activated protein kinase in rodent adipocytes. J. Biol. Chem. 2005, 280, 25250–25257. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The ampk signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Winder, W.; Hardie, D. Amp-activated protein kinase, a metabolic master switch: Possible roles in type 2 diabetes. Am. J. Physiol.-Endocrinol. Metab. 1999, 277, E1–E10. [Google Scholar]
- Lin, C.-L.; Huang, H.-C.; Lin, J.-K. Theaflavins attenuate hepatic lipid accumulation through activating ampk in human hepg2 cells. J. Lipid Res. 2007, 48, 2334–2343. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Banerjee, S.; Porter, T.D. Green and black tea extracts inhibit hmg-coa reductase and activate amp kinase to decrease cholesterol synthesis in hepatoma cells. J. Nutr. Biochem. 2009, 20, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Kudo, N.; Arai, Y.; Suhara, Y.; Ishii, T.; Nakayama, T.; Osakabe, N. A single oral administration of theaflavins increases energy expenditure and the expression of metabolic genes. PLoS ONE 2015, 10, e0137809. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Wang, L.; Wang, L.; Tanaka, Y.; Zhang, T.; Ashida, H. Oolong, black and pu-erh tea suppresses adiposity in mice via activation of amp-activated protein kinase. Food Funct. 2014, 5, 2420–2429. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.-J.; Lo, C.-Y.; Wang, B.-J.; Chiou, R.Y.-Y.; Lin, S.-M. Theaflavin-3,3′-digallate, a black tea polyphenol, stimulates lipolysis associated with the induction of mitochondrial uncoupling proteins and ampk–foxo3a–mnsod pathway in 3T3-L1 adipocytes. J. Funct. Foods 2015, 17, 271–282. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, M. Biological role of liver x receptors. J. Physiol. Pharmacol. 2008, 59, 31–55. [Google Scholar] [PubMed]
- Claudel, T.; Staels, B.; Kuipers, F. The farnesoid X receptor a molecular link between bile acid and lipid and glucose metabolism. Arterioscler. Thrombos. Vasc. Biol. 2005, 25, 2020–2030. [Google Scholar] [CrossRef] [PubMed]
- Szanto, A.; Narkar, V.; Shen, Q.; Uray, I.; Davies, P.; Nagy, L. Retinoid x receptors: X-ploring their (patho) physiological functions. Cell Death Differ. 2004, 11, S126–S143. [Google Scholar] [CrossRef] [PubMed]
- Wang, K. Enzymatic Synthesis, Purification, Identification and Bioactivity of Theaflavins. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 20 October 2007. [Google Scholar]
- Deng, H.-J.; Li, Y.; Yi, J.; Cao, J. Effects of (−)-epigallocatechin gallate and black tea polyphenols on lipid metabolism related gene expression of HepG2 cells. Acta Nutr. Sin. 2006, 28, 498–501. [Google Scholar]
- Fu, L.; Xi-fu, S. Effect of theaflavins on the differentiation of rabbit bone marrow mesenchymal stem cells into adipocytes. J. Clin. Rehabil. Tissue Eng. Res. 2008, 16, 3061–3064. [Google Scholar]
- Sun, S.-L.; Ling, C.-J.; Liu, J.; Pan, S.-S.; Miao, A.-Q.; Li, J.-X.; Pang, S.; Lai, Z.-X.; Huang, H.-L. Effects of tea polyphenols and theaflavins on proliferation and differentiation of 3T3-L1 preadipocytes. Guangdong Agric. Sci. 2011, 12, 051. [Google Scholar]
- Imamura, M.; Inoguchi, T.; Ikuyama, S.; Taniguchi, S.; Kobayashi, K.; Nakashima, N.; Nawata, H. Adrp stimulates lipid accumulation and lipid droplet formation in murine fibroblasts. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E775–E783. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.; Deng, H.-J.; Cao, J. The comparative study on effects of green tea and black tea polyphenols on genes related to adipocyte differentiation in rats. Acta Nutr. Sin. 2007, 29, 582–587. [Google Scholar]
- Hara, Y.; Honda, M. The inhibition of α-amylase by tea polyphenols. Agric. Biol. Chem. 1990, 54, 1939–1945. [Google Scholar] [CrossRef]
- Kusano, R.; Andou, H.; Fujieda, M.; Tanaka, T.; Matsuo, Y.; Kouno, I. Polymer-like polyphenols of black tea and their lipase and amylase inhibitory activities. Chem. Pharm. Bull. 2008, 56, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Hara, Y. Inhibition of rat small intestinal sucrase and α-glucosidase activities by tea polyphenols. Biosci. Biotechnol. Biochem. 1993, 57, 123–124. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Tanaka, T.; Tamura, S.; Toshima, A.; Tamaya, K.; Miyata, Y.; Tanaka, K.; Matsumoto, K. Α-glucosidase inhibitory profile of catechins and theaflavins. J. Agric. Food Chem. 2007, 55, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Striegel, L.; Kang, B.; Pilkenton, S.J.; Rychlik, M.; Apostolidis, E. Effect of black tea and black tea pomace polyphenols on α-glucosidase and α-amylase inhibition, relevant to type 2 diabetes prevention. Front. Nutr. 2015, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.I.; Apostolidis, E.; Shetty, K. Inhibitory potential of wine and tea against α-amylase and α-glucosidase for management of hyperglycemia linked to type 2 diabetes. J. Food Biochem. 2008, 32, 15–31. [Google Scholar] [CrossRef]
- Koh, L.W.; Wong, L.L.; Loo, Y.Y.; Kasapis, S.; Huang, D. Evaluation of different teas against starch digestibility by mammalian glycosidases. J. Agric. Food Chem. 2009, 58, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Igarashi, M.; Yamada, S.; Takahashi, N.; Watanabe, K. Inhibitory effect of black tea and its combination with acarbose on small intestinal α-glucosidase activity. J. Ethnopharmacol. 2015, 161, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Toyozaki, M.; Osaka, M.; Kondo, K.; Yoshida, M. High fat and high cholesterol diet induces dpp-iv activity in intestinal lymph. J. Oleo Sci. 2013, 62, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, J.; Okuyama, T.; Kyohara, M.; Yoshida, E.; Togashi, Y.; Tajima, K.; Yamazaki, S.; Kaji, M.; Koganei, M.; Sasaki, H. Dpp-4 inhibition improves early mortality, β cell function, and adipose tissue inflammation in db/db mice fed a diet containing sucrose and linoleic acid. Diabetol. Metab. Syndr. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Mardanyan, S.; Sharoyan, S.; Antonyan, A.; Zakaryan, N., IV. Dipeptidyl peptidase iv and adenosine deaminase inhibition by armenian plants and antidiabetic drugs. Int. J. Diabetes Metab. 2011, 19, 69–74. [Google Scholar]
- Yang, M.-H.; Wang, C.-H.; Chen, H.-L. Green, oolong and black tea extracts modulate lipid metabolism in hyperlipidemia rats fed high-sucrose diet. J. Nutr. Biochem. 2001, 12, 14–20. [Google Scholar] [CrossRef]
- Vincent, H.K.; Taylor, A.G. Biomarkers and potential mechanisms of obesity-induced oxidant stress in humans. Int. J. Obes. 2006, 30, 400–418. [Google Scholar] [CrossRef] [PubMed]
- Pihl, E.; Zilmer, K.; Kullisaar, T.; Kairane, C.; Mägi, A.; Zilmer, M. Atherogenic inflammatory and oxidative stress markers in relation to overweight values in male former athletes. Int. J. Obes. 2006, 30, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Chrysohoou, C.; Panagiotakos, D.B.; Pitsavos, C.; Skoumas, I.; Papademetriou, L.; Economou, M.; Stefanadis, C. The implication of obesity on total antioxidant capacity in apparently healthy men and women: The attica study. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Saha, P.; Das, S. Regulation of hazardous exposure by protective exposure: Modulation of phase ii detoxification and lipid peroxidation by camellia sinensis and swertia chirata. Teratog. Carcinog. Mutag. 2003, 23, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-Y.; Li, W.; Xu, Y.; Jin, E.-H.; Tu, Y.-Y. Evaluation of the antioxidant effects of four main theaflavin derivatives through chemiluminescence and DNA damage analyses. J. Zhejiang Univ. Sci. B 2011, 12, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jie, G.; Dong, F.; Xu, Y.; Watanabe, N.; Tu, Y. Radical-scavenging abilities and antioxidant properties of theaflavins and their gallate esters in H2O2-mediated oxidative damage system in the hpf-1 cells. Toxicol. In Vitro 2008, 22, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.K.; Su, Y.; Chen, R.; Zhang, Z.; Huang, Y.; Chen, Z.-Y. Theaflavins in black tea and catechins in green tea are equally effective antioxidants. J. Nutr. 2001, 131, 2248–2251. [Google Scholar] [PubMed]
- Yoshida, H.; Ishikawa, T.; Hosoai, H.; Suzukawa, M.; Ayaori, M.; Hisada, T.; Sawada, S.; Yonemura, A.; Higashi, K.; Ito, T. Inhibitory effect of tea flavonoids on the ability of cells to oxidize low density lipoprotein. Biochem. Pharmacol. 1999, 58, 1695–1703. [Google Scholar] [CrossRef]
- Axling, U.; Olsson, C.; Xu, J.; Fernandez, C.; Larsson, S.; Ström, K.; Ahrné, S.; Holm, C.; Molin, G.; Berger, K. Green tea powder and lactobacillus plantarum affect gut microbiota, lipid metabolism and inflammation in high-fat fed c57bl/6j mice. Nutr. Metab. 2012, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.S.; Touyama, M.; Hisada, T.; Benno, Y. Effects of green tea consumption on human fecal microbiota with special reference to bifidobacterium species. Microbiol. Immunol. 2012, 56, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Kemperman, R.A.; Gross, G.; Mondot, S.; Possemiers, S.; Marzorati, M.; van de Wiele, T.; Doré, J.; Vaughan, E.E. Impact of polyphenols from black tea and red wine/grape juice on a gut model microbiome. Food Res. Int. 2013, 53, 659–669. [Google Scholar] [CrossRef]
- Panickar, K.S. Effects of dietary polyphenols on neuroregulatory factors and pathways that mediate food intake and energy regulation in obesity. Mol. Nutr. Food Res. 2013, 57, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhu, W.; Shen, C.-L.; Gao, W. Green tea polyphenols reduce body weight in rats by modulating obesity-related genes. PLoS ONE 2012, 7, e38332. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, H.; Gao, Y.; Tu, Y. Mechanisms of Body Weight Reduction by Black Tea Polyphenols. Molecules 2016, 21, 1659. https://doi.org/10.3390/molecules21121659
Pan H, Gao Y, Tu Y. Mechanisms of Body Weight Reduction by Black Tea Polyphenols. Molecules. 2016; 21(12):1659. https://doi.org/10.3390/molecules21121659
Chicago/Turabian StylePan, Haibo, Ying Gao, and Youying Tu. 2016. "Mechanisms of Body Weight Reduction by Black Tea Polyphenols" Molecules 21, no. 12: 1659. https://doi.org/10.3390/molecules21121659
APA StylePan, H., Gao, Y., & Tu, Y. (2016). Mechanisms of Body Weight Reduction by Black Tea Polyphenols. Molecules, 21(12), 1659. https://doi.org/10.3390/molecules21121659