Selective Inhibitory Effect of Epigallocatechin-3-gallate on Migration of Vascular Smooth Muscle Cells

 ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
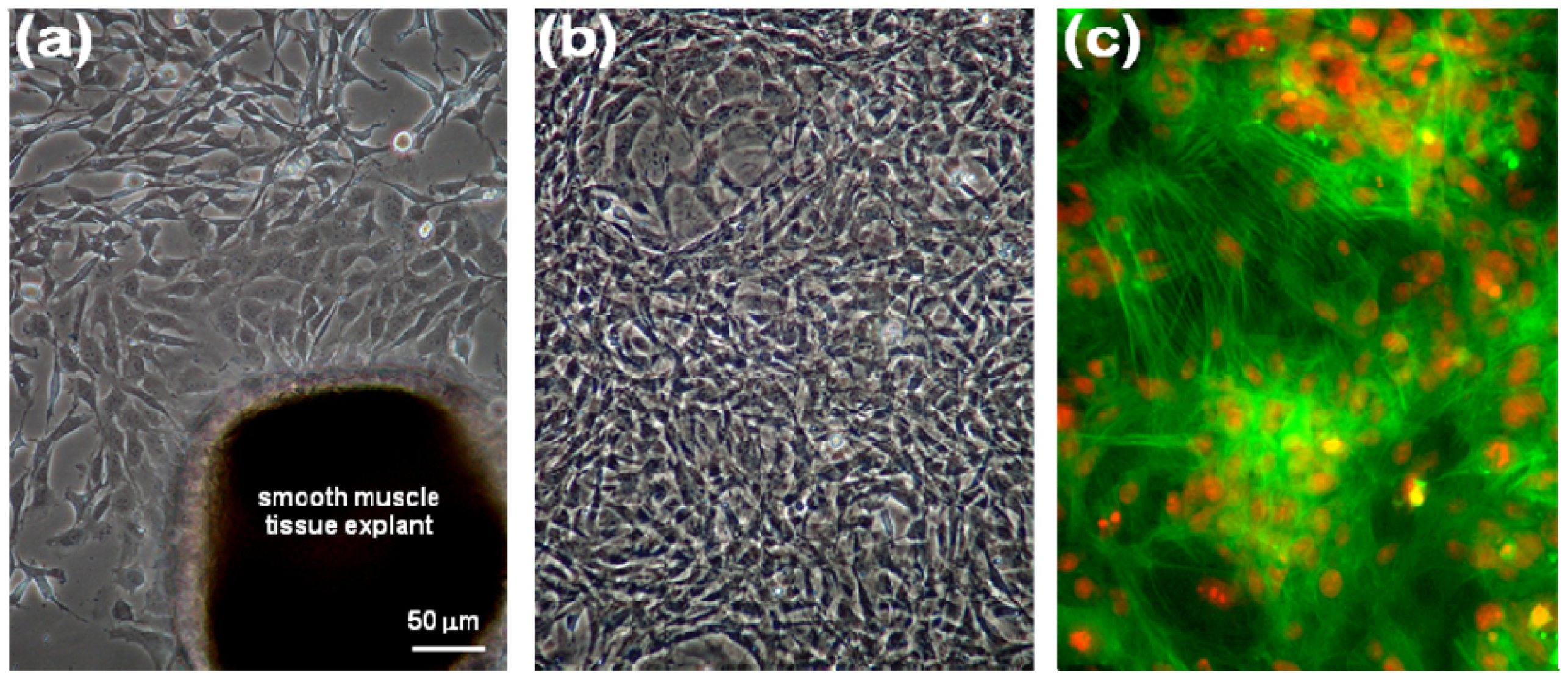
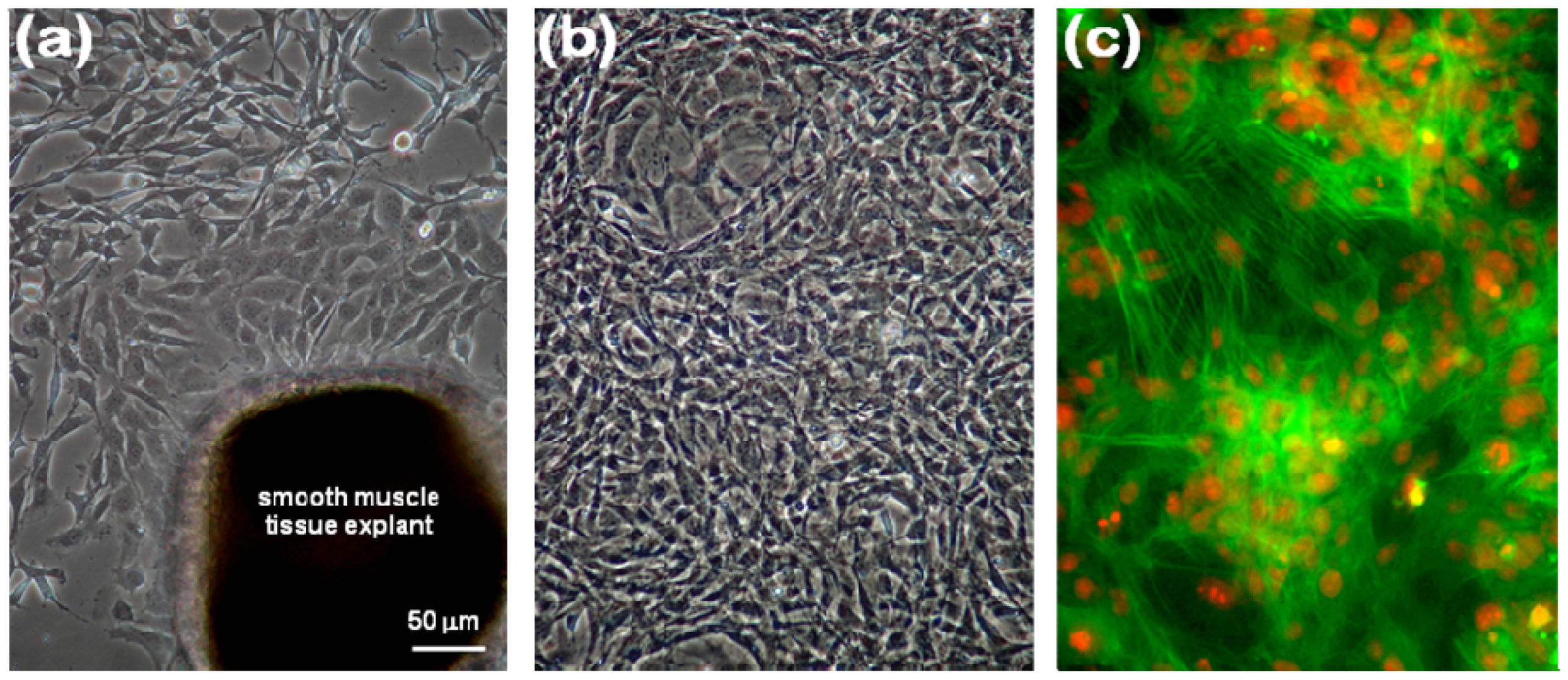
2.1. Immunocytochemical Characterization of VSMCs
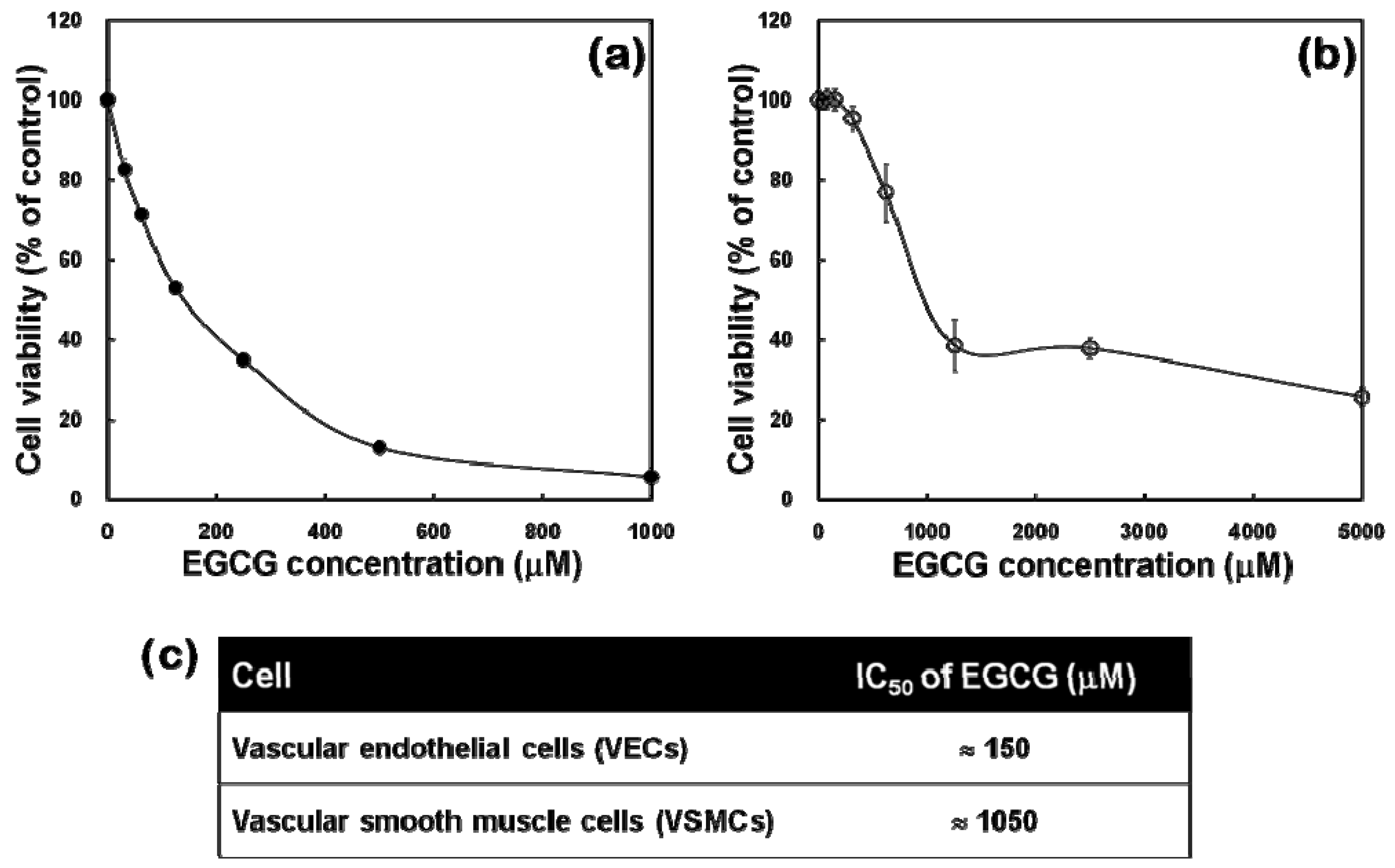
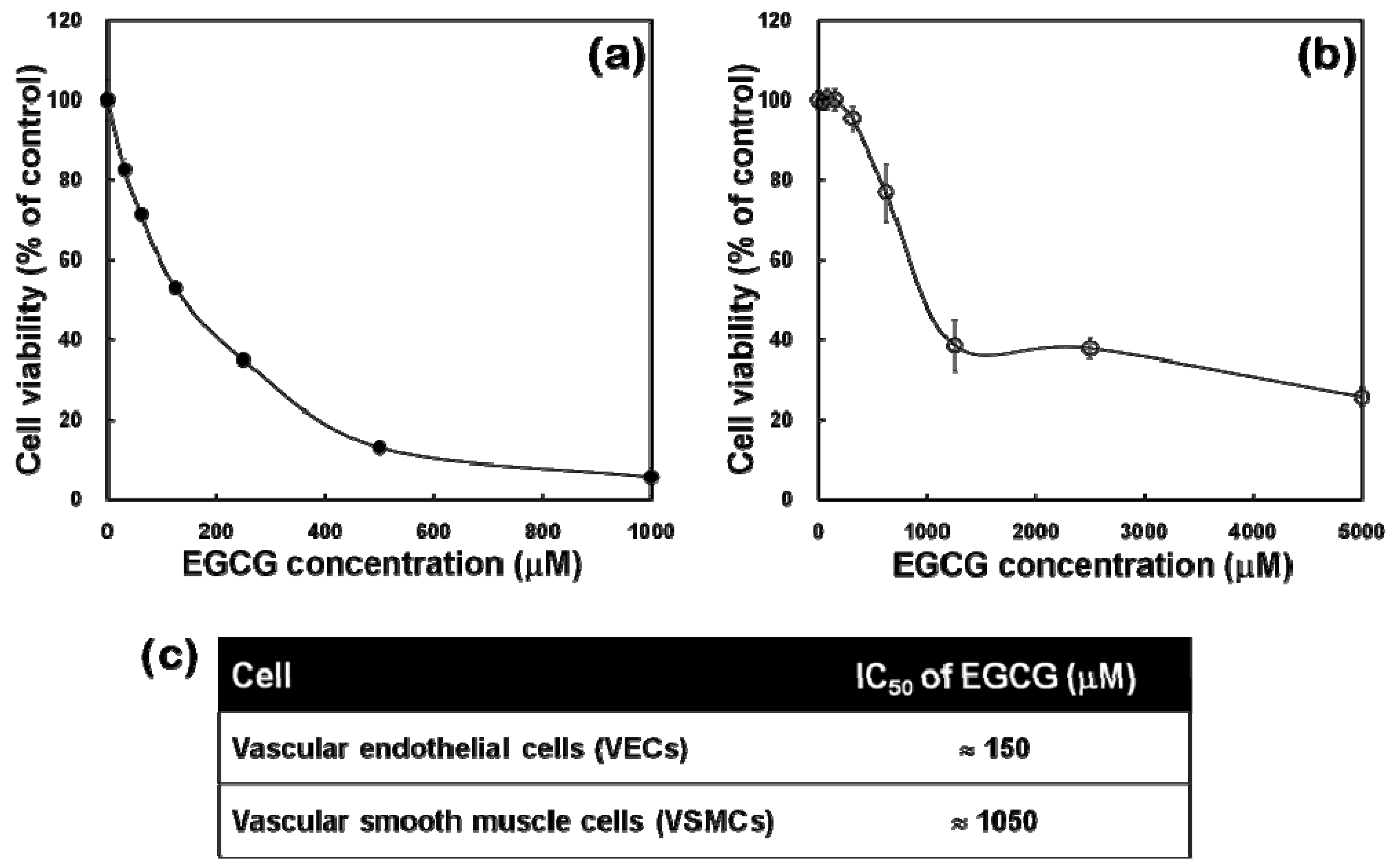
2.2. Cytotoxicity Profiles of EGCG to VECs and VSMCs
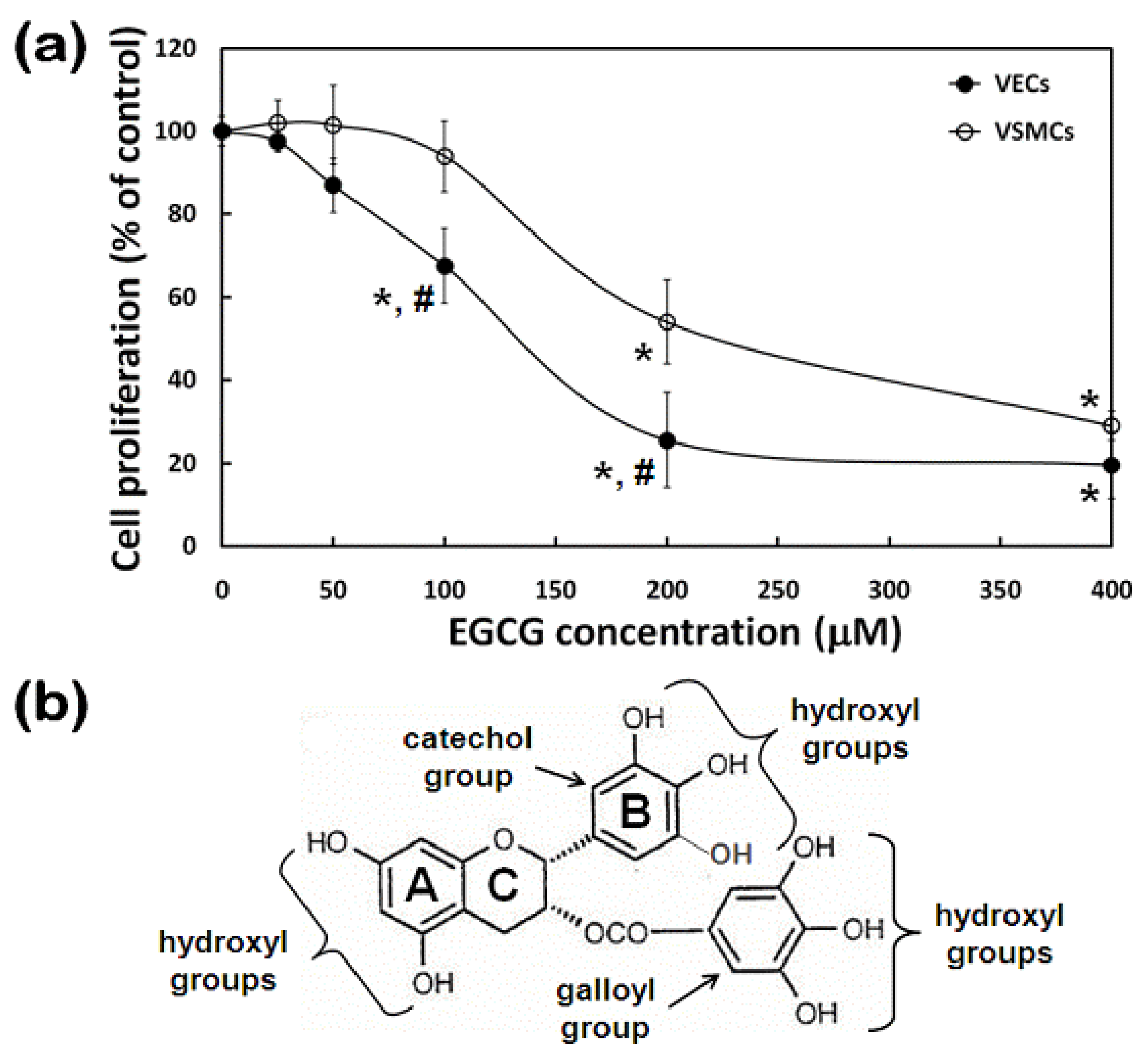
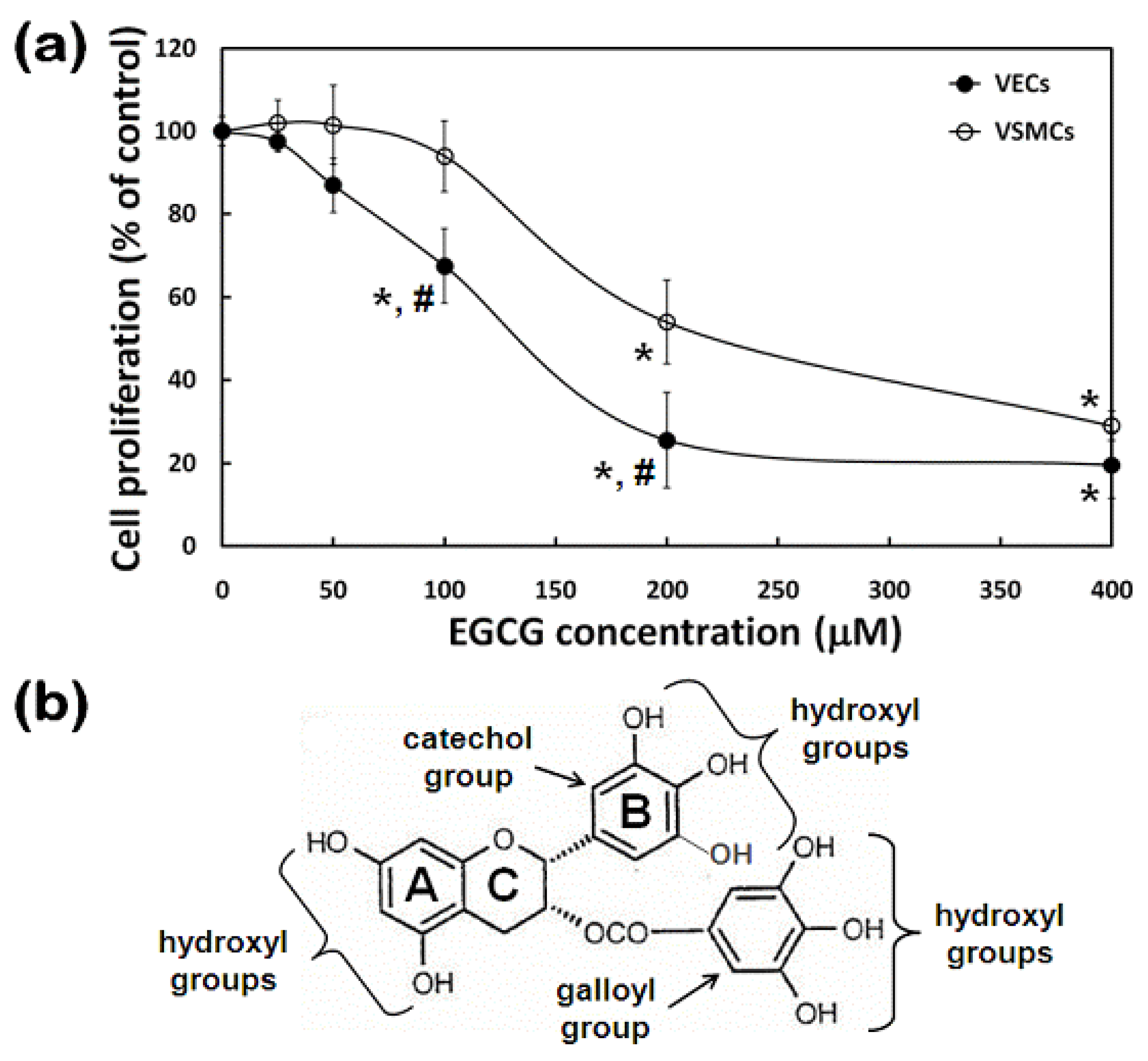
2.3. Effects of EGCG on Proliferation of VECs and VSMCs
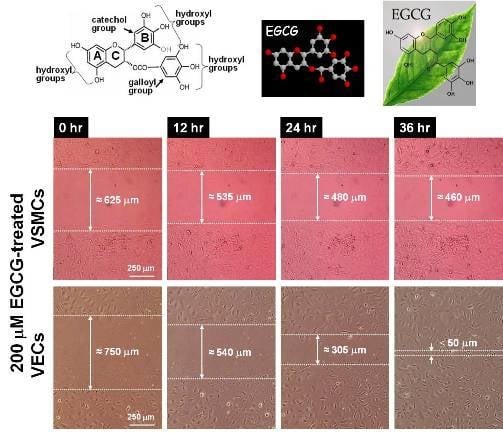
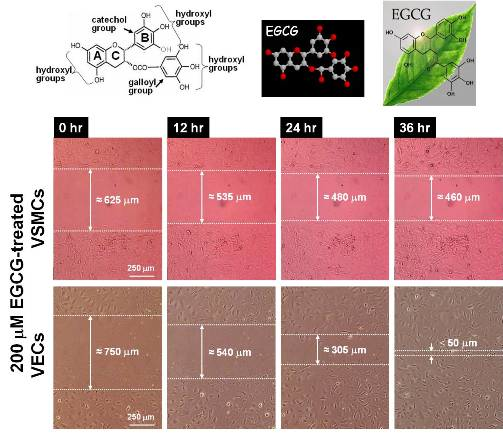
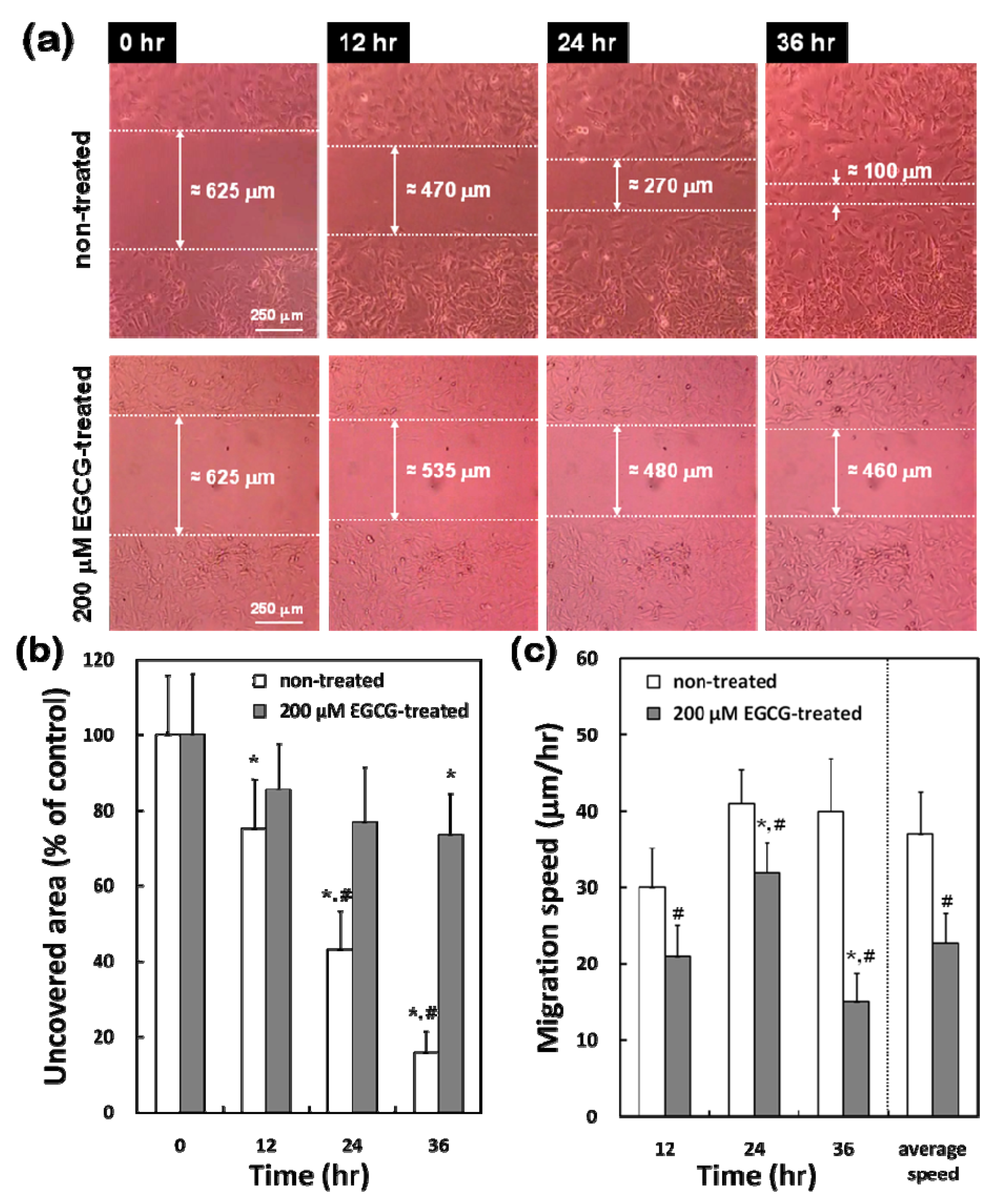
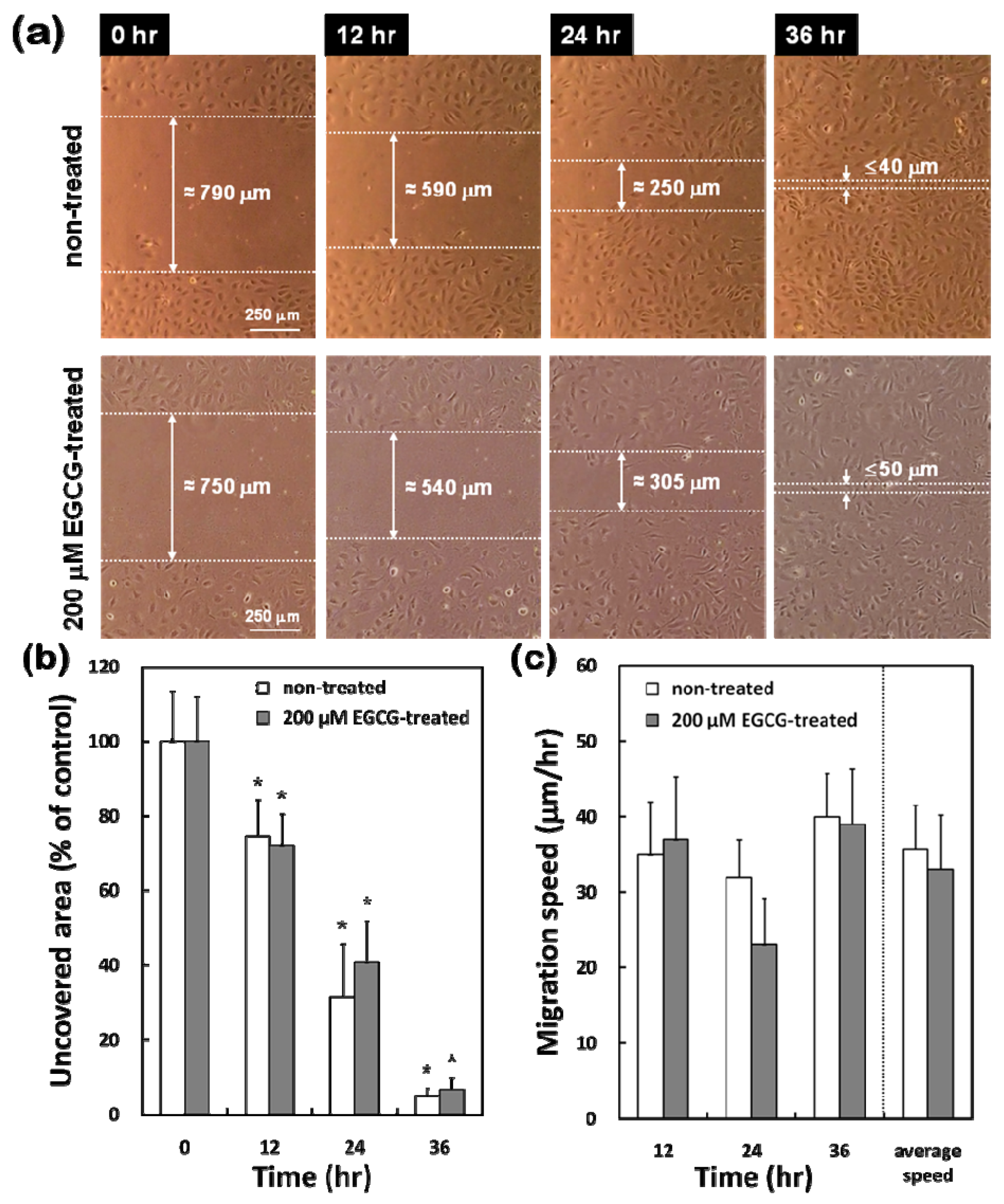
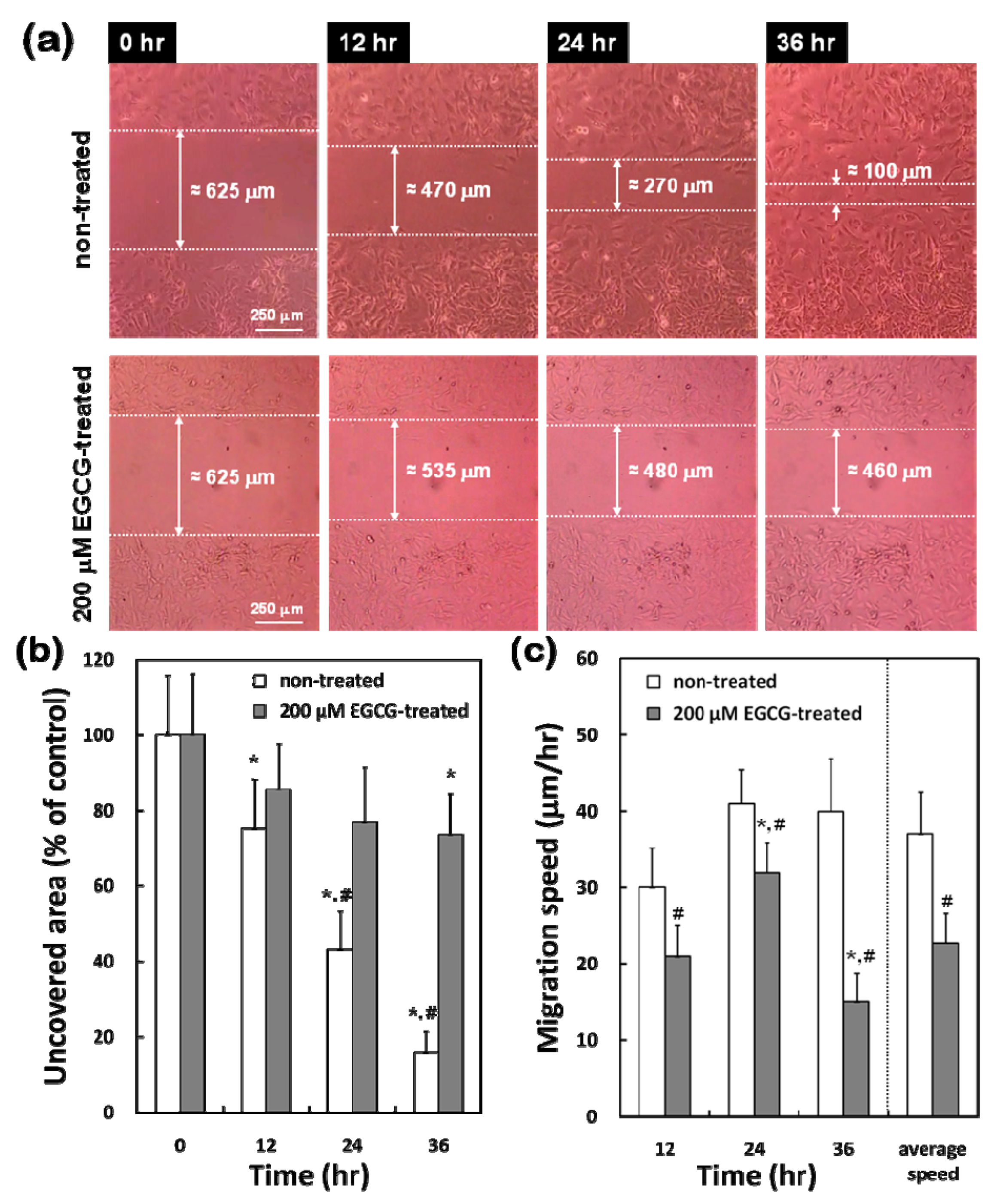
2.4. Differential Effects of EGCG on Migration of VECs vs. VSMCs
3. Experimental
3.1. Cell Cultures
3.2. Morphological and Immunocytochemical Analyses of Primarily Cultured VSMCs
3.3. Cytotoxicity of EGCG to VECs and VSMCs
3.4. EGCG Treatments
3.5. Cell Proliferation Assay
3.6. Cell Migration Assay
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Tsai, S.; Butler, J.; Rafii, S.; Liu, B.; Kent, K.C. The role of progenitor cells in the development of intimal hyperplasia. J. Vasc. Surg. 2009, 49, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, G.H.; Dzau, V.J. The emerging concept of vascular remodeling. N. Engl. J. Med. 1994, 330, 1431–1438. [Google Scholar] [PubMed]
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Lindner, V.; Reidy, M.A. Proliferation of smooth muscle cells after vascular injury is inhibited by an antibody against basic fibroblast growth factor. Proc. Natl. Acad. Sci. USA 1991, 88, 3739–3743. [Google Scholar] [CrossRef] [PubMed]
- Ferns, G.A.; Raines, E.W.; Sprugel, K.H.; Motani, A.S.; Reidy, M.A.; Ross, R. Inhibition of neointimal smooth muscle accumulation after angioplasty by an antibody to PDGF. Science 1991, 253, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Lim, S.; Song, H.; Song, B.W.; Kim, H.J.; Cha, M.J.; Sung, J.M.; Kim, T.W.; Hwang, K.C. Cordycepin inhibits vascular smooth muscle cell proliferation. Eur. J. Pharmacol. 2008, 597, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, D.; Stevens, S.L.; Skillern, C.S.; Wellons, E.D.; Robinson, K.; Matsuura, J.H.; Gannon, B.J. Topical application of β-radiation to reduce intimal hyperplasia after carotid artery balloon injury in rabbit. A possible application for brachytherapy in vascular surgery. Cardiovasc. Radiat. Med. 2002, 3, 16–19. [Google Scholar] [CrossRef]
- O’Connor, D.M.; O’Brien, T. Nitric oxide synthase gene therapy: Progress and prospects. Expert Opin. Biol. Ther. 2009, 9, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Gershlick, A.H. Treating atherosclerosis: Local drug delivery from laboratory studies to clinical trials. Atherosclerosis 2002, 160, 259–271. [Google Scholar] [CrossRef]
- Na, H.-K.; Surh, Y.-J. Intracellular signaling network as a prime chemopreventive target of (–)-epigallocatechin gallate. Mol. Nutr. Food Res. 2006, 50, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Bar-Sela, G.; Epelbaum, R.; Schaffer, M. Curcumin as an anti-cancer agent: Review of the gap between basic and clinical applications. Curr. Med. Chem. 2010, 17, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Wang, Z.; Sarkar, F.H. Regulation of microRNAs by natural agents: An emerging field in chemoprevention and chemotherapy research. Pharm. Res. 2010, 27, 1027–1041. [Google Scholar] [CrossRef] [PubMed]
- Stangl, V.; Lorenz, M.; Stangl, K. The role of tea and tea flavonoids in cardiovascular health. Mol. Nutr. Food Res. 2006, 50, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Stangl, V.; Dreger, H.; Stangl, K.; Lorenz, M. Molecular targets of tea polyphenols in the cardiovascular system. Cardiovasc. Res. 2007, 73, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Jochmann, N.; Baumann, G.; Stangl, V. Green tea and cardiovascular disease: From molecular targets towards human health. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Frei, B.; Higdon, J.V. Antioxidant activity of tea polyphenols in vivo: Evidence from animal studies. J. Nutr. 2003, 133, 3275S–3284S. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Han, D.-W.; Lee, J.J.; Jung, D.Y.; Park, J.-C.; Hyon, S.-H. Development of epigallocatechin gallate-eluting polymeric stent and its physicochemical, biomechanical and biological evaluations. Biomed. Mater. 2009, 4, 044104. [Google Scholar] [CrossRef] [PubMed]
- Han, D.-W.; Hyon, S.-H.; Park, J.-C. Protection and preservation of vascular cells and tissues by green tea polyphenols. In Polyphenols and Health: New and Recent Advances, 1st ed.; Vassallo, N., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2008; pp. 87–111. [Google Scholar]
- Rah, D.K.; Han, D.-W.; Baek, H.S.; Hyon, S.-H.; Park, J.-C. Prevention of reactive oxygen species-induced oxidative stress in human microvascular endothelial cells by green tea polyphenol. Toxicol. Lett. 2005, 155, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Ohga, N.; Hida, K.; Hida, Y.; Muraki, C.; Tsuchiya, K.; Matsuda, K.; Ohiro, Y.; Totsuka, Y.; Shindoh, M. Inhibitory effects of epigallocatechin-3 gallate, a polyphenol in green tea, on tumor-associated endothelial cells and endothelial progenitor cells. Cancer Sci. 2009, 100, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. Atherosclerosis is an inflammatory disease. Am. Heart J. 1999, 138, S419–S420. [Google Scholar] [CrossRef]
- Chyu, K.Y.; Babbidge, S.M.; Zhao, X.; Dandillaya, R.; Rietveld, A.G.; Yano, J.; Dimayuga, P.; Cercek, B.; Shah, P.K. Differential effects of green tea-derived catechin on developing versus established atherosclerosis in apolipoprotein E-null mice. Circulation 2004, 109, 2448–2453. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, C.S.; Sonenshein, G.E. Green tea polyphenol epigallocatechin-3 gallate induces apoptosis of proliferating vascular smooth muscle cells via activation of p53. FASEB J. 2003, 17, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Han, D.-W.; Lim, H.R.; Baek, H.S.; Lee, M.H.; Lee, S.J.; Hyon, S.-H.; Park, J.-C. Inhibitory effects of epigallocatechin-3-O-gallate on serum-stimulated rat aortic smooth muscle cells via nuclear factor-κB down-modulation. Biochem. Biophys. Res. Commun. 2006, 345, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Won, S.M.; Park, Y.H.; Kim, H.J.; Park, K.M.; Lee, W.J. Catechins inhibit angiotensin II-induced vascular smooth muscle cell proliferation via mitogen-activated protein kinase pathway. Exp. Mol. Med. 2006, 38, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, E.; Campos-Toimil, M.; Justiniano-Basaran, H.; Lugnier, C.; Orallo, F. Study of the mechanisms involved in the vasorelaxation induced by (–)-epigallocatechin-3-gallate in rat aorta. Br. J. Pharmacol. 2006, 147, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.A.; Neuhaus, T.; Skach, R.A.; Hescheler, J.; Ahn, H.Y.; Schrör, K.; Ko, Y.; Sachinidis, A. Mechanisms of the inhibitory effects of epigallocatechin-3 gallate on platelet-derived growth factor-BB-induced cell signaling and mitogenesis. FASEB J. 2004, 18, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Galis, Z.S.; Khatri, J.J. Matrix metalloproteinases in vascular remodeling and atherogenesis: The good, the bad, and the ugly. Circ. Res. 2002, 90, 251–262. [Google Scholar] [PubMed]
- Huang, Y.; Mironova, M.; Lopes-Virella, M.F. Oxidized LDL stimulates matrix metalloproteinase-1 expression in human vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2640–2647. [Google Scholar] [CrossRef] [PubMed]
- Sukhova, G.K.; Schönbeck, U.; Rabkin, E.; Schoen, F.J.; Poole, A.R.; Billinghurst, R.C.; Libby, P. Evidence for increased collagenolysis by interstitial collagenases-1 and -3 in vulnerable human atheromatous plaques. Circulation 1999, 99, 2503–2509. [Google Scholar] [CrossRef]
- Cho, H.H.; Han, D.-W.; Matsumura, K.; Tsutsumi, S.; Hyon, S.-H. The behavior of vascular smooth muscle cells and platelets onto epigallocatechin gallate-releasing poly(L-lactide-co-ε-caprolactone) as stent-coating materials. Biomaterials 2008, 29, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Han, D.-W.; Jung, D.Y.; Park, J.-C.; Cho, H.H.; Hyon, S.-H.; Han, D.K. Underlying mechanism for suppression of vascular smooth muscle cells by green tea polyphenol EGCG released from biodegradable polymers for stent application. J. Biomed. Mater. Res. A 2010, 95, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.R.; Baek, H.S.; Lee, M.H.; Woo, Y.I.; Han, D.-W.; Han, M.H.; Baik, H.K.; Choi, W.S.; Park, K.D.; Chung, K.-H.; Park, J.-C. Surface modification for enhancing behaviors of vascular endothelial cells onto polyurethane films by microwave-induced argon plasma. Surf. Coat. Technol. 2008, 202, 5768–5772. [Google Scholar] [CrossRef]
- Park, J.-C.; Park, B.J.; Suh, H.; Park, B.Y.; Rah, D.K. Comparative study on motility of the cultured fetal and neonatal dermal fibroblasts in extracellular matrix. Yonsei Med. J. 2001, 42, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.I.; Park, B.J.; Kim, H.L.; Lee, M.H.; Kim, J.; Yang, Y.I.; Kim, J.K.; Tsubaki, K.; Han, D.-W.; Park, J.-C. The biological activities of (1,3)-(1,6)-β-D-glucan and porous electrospun PLGA membranes containing beta-glucan in human dermal fibroblasts and adipose tissue-derived stem cells. Biomed. Mater. 2010, 5, 044109. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of EGCG are available from the authors. |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Han, D.-W.; Lee, M.H.; Kwon, B.-J.; Kim, H.-L.; Hyon, S.-H.; Park, J.-C. Selective Inhibitory Effect of Epigallocatechin-3-gallate on Migration of Vascular Smooth Muscle Cells. Molecules 2010, 15, 8488-8500. https://doi.org/10.3390/molecules15118488
Han D-W, Lee MH, Kwon B-J, Kim H-L, Hyon S-H, Park J-C. Selective Inhibitory Effect of Epigallocatechin-3-gallate on Migration of Vascular Smooth Muscle Cells. Molecules. 2010; 15(11):8488-8500. https://doi.org/10.3390/molecules15118488
Chicago/Turabian StyleHan, Dong-Wook, Mi Hee Lee, Byeong-Ju Kwon, Hye-Lee Kim, Suong-Hyu Hyon, and Jong-Chul Park. 2010. "Selective Inhibitory Effect of Epigallocatechin-3-gallate on Migration of Vascular Smooth Muscle Cells" Molecules 15, no. 11: 8488-8500. https://doi.org/10.3390/molecules15118488
APA StyleHan, D.-W., Lee, M. H., Kwon, B.-J., Kim, H.-L., Hyon, S.-H., & Park, J.-C. (2010). Selective Inhibitory Effect of Epigallocatechin-3-gallate on Migration of Vascular Smooth Muscle Cells. Molecules, 15(11), 8488-8500. https://doi.org/10.3390/molecules15118488