
Seaweeds as a Fermentation Substrate: A Challenge for the Food Processing Industry


 , ,
, ,  and
and 
Abstract
1. Introduction
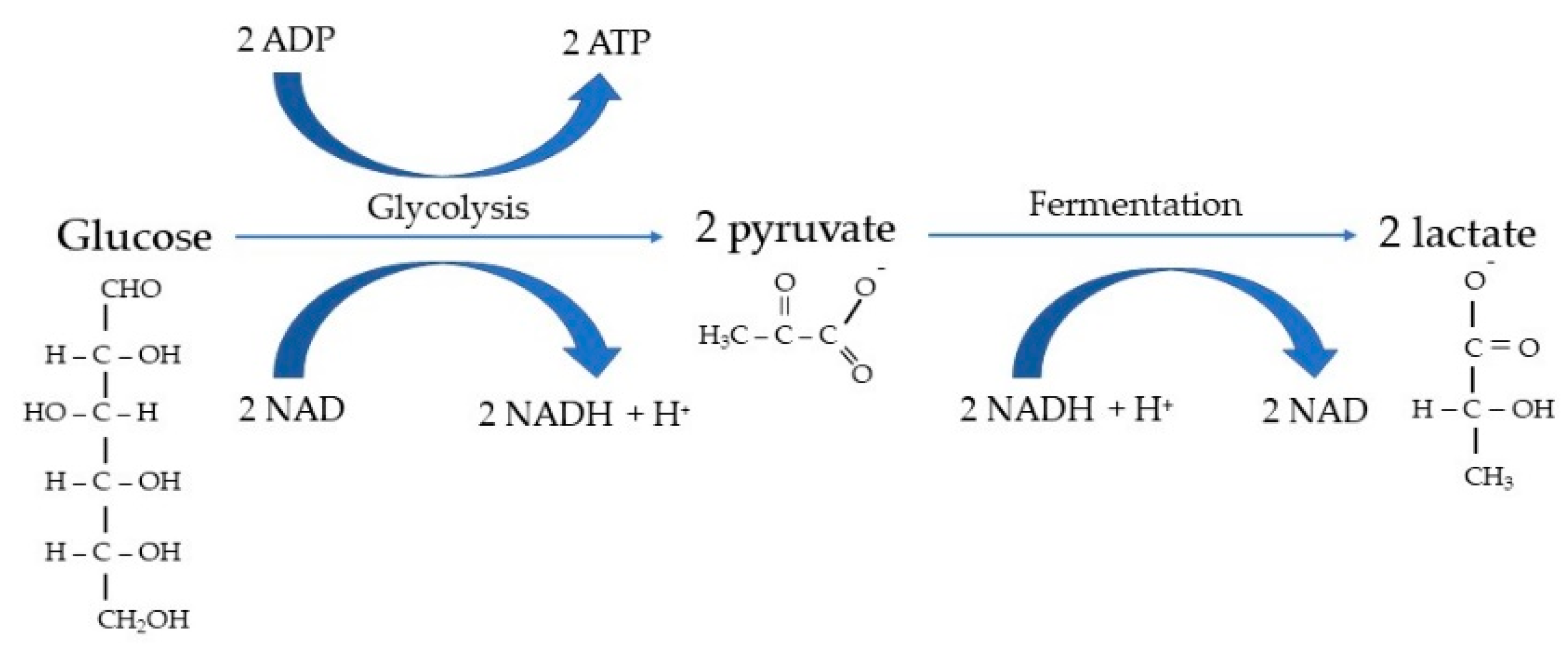
2. Fermentation’s Basic Concepts
Relevant Fermentative Strains—Lactic Acid Bacteria
3. Seaweed as a Fermentation Substrate
3.1. Green Seaweeds
3.2. Red Seaweeds
3.3. Brown Seaweeds
4. Food Industrial Applications of Fermentation Procedures on Algal Biomass
Functional Food Production
5. Challenges to the Fermentation-Based Manufacture of Food Seaweed Products
6. Seaweed Polysaccharides as Prebiotics for Gut Bacteria
6.1. The Biochemical Explanation for the Nutraceutical/Prebiotic Effect
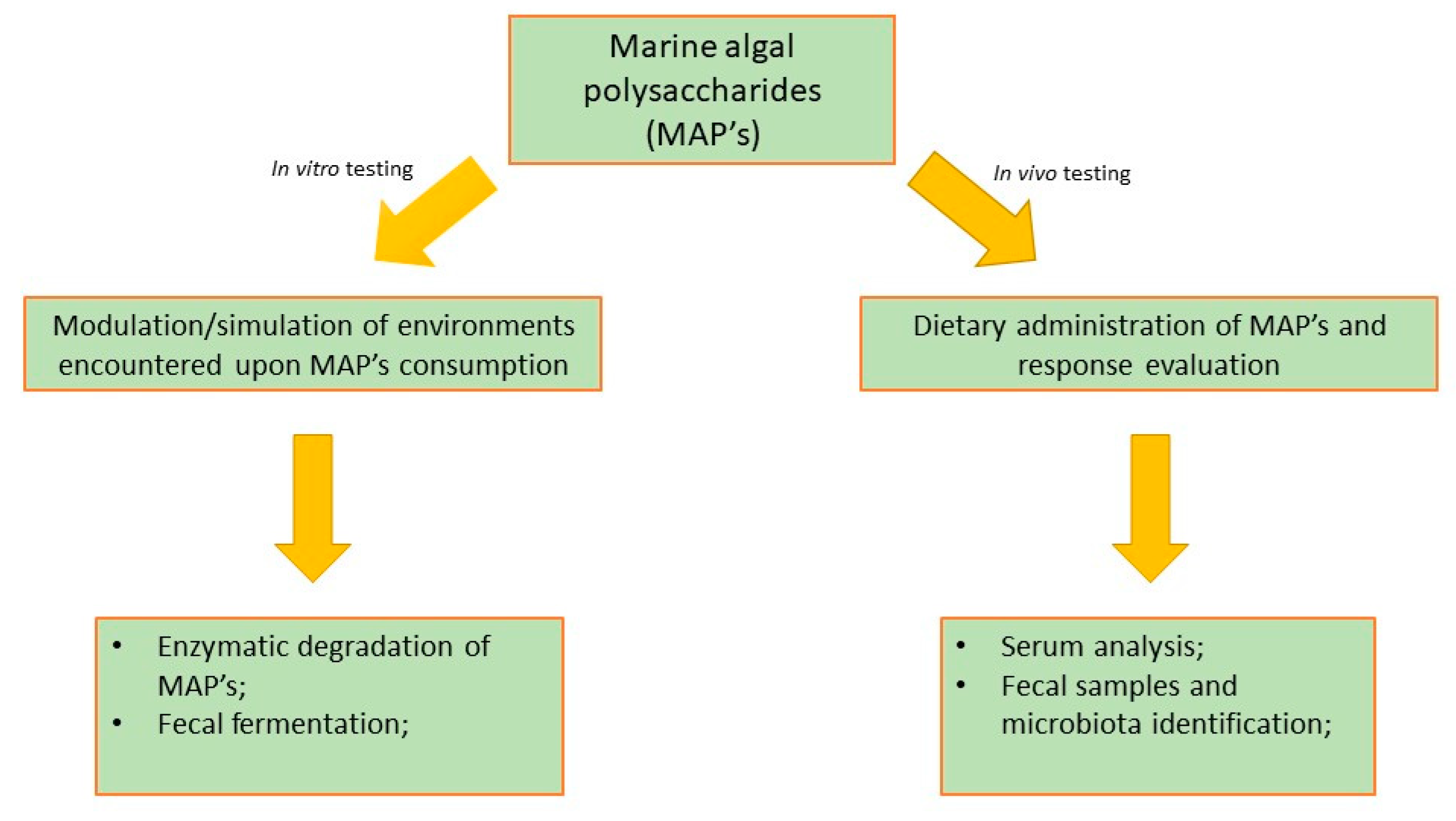
6.2. How Are the Prebiotics Studies Performed?
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pal, A.; Kamthania, M.C.; Kumar, A. Bioactive Compounds and Properties of Seaweeds—A Review. OALib 2014, 1, 1–17. [Google Scholar] [CrossRef]
- Panzella, L.; Napolitano, A. Natural phenol polymers: Recent advances in food and health applications. Antioxidants 2017, 6, 30. [Google Scholar] [CrossRef]
- Ale, M.T.; Meyer, A.S. Fucoidans from brown seaweeds: An update on structures, extraction techniques and use of enzymes as tools for structural elucidation. RSC Adv. 2013, 3, 8131–8141. [Google Scholar] [CrossRef]
- Pereira, L. Therapeutic and Nutritional Uses of Algae; Pereira, L., Ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2018. [Google Scholar]
- Torres, M.D.; Flórez-Fernández, N.; Domínguez, H. Integral utilization of red seaweed for bioactive production. Mar. Drugs 2019, 17, 314. [Google Scholar] [CrossRef]
- Plaza, M.; Herrero, M.; Alejandro Cifuentes, A.; Ibáñez, E. Innovative natural functional ingredients from microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef] [PubMed]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2011, 23, 321–335. [Google Scholar] [CrossRef]
- Cho, M.; Lee, H.S.; Kang, I.J.; Won, M.H.; You, S. Antioxidant properties of extract and fractions from Enteromorpha prolifera, a type of green seaweed. Food Chem. 2011, 127, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Ishwarya, R.; Vaseeharan, B.; Kalyani, S.; Banumathi, B.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Al-anbr, M.N.; Khaled, J.M.; Benelli, G. Facile green synthesis of zinc oxide nanoparticles using Ulva lactuca seaweed extract and evaluation of their photocatalytic, antibiofilm and insecticidal activity. J. Photochem. Photobiol. B Biol. 2018, 178, 249–258. [Google Scholar] [CrossRef]
- Liu, Y.; Morgan, J.B.; Coothankandaswamy, V.; Liu, R.; Jekabsons, M.B.; Mahdi, F.; Nagle, D.G.; Zhou, Y.D. The Caulerpa pigment caulerpin inhibits HIF-1 activation and mitochondrial respiration. J. Nat. Prod. 2009, 72, 2104–2109. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-S.; Athukorala, Y.; Jeon, Y.-J.; Senevirathne, M.; Cho, K.-R.; Kim, S.-H. Antioxidant Activity of Sulfated Polysaccharides Isolated from Sargassum fulvellum. Prev. Nutr. Food Sci. 2007, 12, 65–73. [Google Scholar] [CrossRef]
- Rodrigues, D.; Walton, G.; Sousa, S.; Rocha-Santos, T.A.P.; Duarte, A.C.; Freitas, A.C.; Gomes, A.M.P. In vitro fermentation and prebiotic potential of selected extracts from seaweeds and mushrooms. LWT 2016, 73, 131–139. [Google Scholar] [CrossRef]
- Lin, H.T.V.; Huang, M.Y.; Kao, T.Y.; Lu, W.J.; Lin, H.J.; Pan, C.L. Production of lactic acid from seaweed hydrolysates via lactic acid bacteria fermentation. Fermentation 2020, 6, 37. [Google Scholar] [CrossRef]
- Yadav, H.; Jain, S.; Rastamanesh, R. Fermentation Technology in the Development of Functional Foods for Human Health: Where We Should Head. Ferment. Technol. 2012, 1, 10–13. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented Foods as a Dietary Source of Live Organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Koshio, S. Recent advances in the role of probiotics and prebiotics in carp aquaculture: A review. Aquaculture 2016, 454, 243–251. [Google Scholar] [CrossRef]
- Rakhmanova, A.; Khan, Z.A.; Shah, K. A mini review fermentation and preservation: Role of Lactic Acid Bacteria. MOJ Food Process Technol. 2018, 6, 414–417. [Google Scholar] [CrossRef]
- Felis, G.E.; Pot, B. The family Lactobacillaceae. In Lactic Acid Bacteria; John Wiley & Sons, Ltd.: Chichester, UK, 2014; pp. 245–247. [Google Scholar]
- Ścieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef]
- Narayanan, N.; Roychoudhury, P.K.; Srivastava, A. L (+) lactic acid fermentation and its product polymerization. Electron. J. Biotechnol. 2004, 7, 167–178. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Krishna, B.S.; Nikhilesh, G.S.S.; Tarun, B.; Saibaba, N.K.V.; Gopinadh, R. Industrial production of lactic acid. Int. J. Biotech. Res. 2018, 1, 42–54. [Google Scholar] [CrossRef]
- Wee, Y.J.; Kim, J.N.; Ryu, H.W. Biotechnological production of lactic acid and its recent applications. Food Technol. Biotechnol. 2006, 44, 163–172. [Google Scholar]
- Abd Alsaheb, R.A.; Aladdin, A.; Othman, N.Z.; Abd Malek, R.; Leng, O.M.; Aziz, R.; El Enshasy, H. Lactic acid applications in pharmaceutical and cosmeceutical industries. J. Chem. Pharm. Res. 2015, 7, 729–735. [Google Scholar]
- Bouwstra, J.A.; Ponec, M. The skin barrier in healthy and diseased state. Biochim. Biophys. Acta–Lomembr. 2006, 1758, 2080–2095. [Google Scholar] [CrossRef]
- Södergård, A.; Stolt, M. Properties of polylactic acid fiber based polymers and their correlation with composition. Prog. Polym. Sci. 2002, 27, 1123–1163. [Google Scholar] [CrossRef]
- Vink, E.T.H.; Rábago, K.R.; Glassner, D.A.; Gruber, P.R. Applications of life cycle assessment to NatureWorksTM polylactide (PLA) production. Polym. Degrad. Stab. 2003, 80, 403–419. [Google Scholar] [CrossRef]
- Chen, M.; Sun, Q.; Giovannucci, E.; Mozaffarian, D.; Manson, J.A.E.; Willett, W.C.; Hu, F.B. Dairy consumption and risk of type 2 diabetes: 3 cohorts of US adults and an updated meta-analysis. BMC Med. 2014, 12, 215. [Google Scholar] [CrossRef]
- Eussen, S.J.P.M.; Van Dongen, M.C.J.M.; Wijckmans, N.; Den Biggelaar, L.; Oude Elferink, S.J.W.H.; Singh-Povel, C.M.; Schram, M.T.; Sep, S.J.S.; Van Der Kallen, C.J.; Koster, A.; et al. Consumption of dairy foods in relation to impaired glucose metabolism and type 2 diabetes mellitus: The Maastricht Study. Br. J. Nutr. 2016, 115, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Soedamah-Muthu, S.S.; Masset, G.; Verberne, L.; Geleijnse, J.M.; Brunner, E.J. Consumption of dairy products and associations with incident diabetes, CHD and mortality in the Whitehall II study. Br. J. Nutr. 2013, 109, 718–726. [Google Scholar] [CrossRef]
- Tapsell, L.C. Fermented dairy food and CVD risk. Br. J. Nutr. 2015, 113, S131–S135. [Google Scholar] [CrossRef]
- Lorea Baroja, M.; Kirjavainen, P.V.; Hekmat, S.; Reid, G. Anti-inflammatory effects of probiotic yogurt in inflammatory bowel disease patients. Clin. Exp. Immunol. 2007, 149, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, A.; Bažec, K.; Rodolfi, L.; Biondi, N.; Zlatić, E.; Jamnik, P.; Tredici, M.R. Lactic Acid Fermentation of Arthrospira platensis (Spirulina) in a Vegetal Soybean Drink for Developing New Functional Lactose-Free Beverages. Front. Microbiol. 2020, 11, 2680. [Google Scholar] [CrossRef]
- Adsul, M.G.; Varma, A.J.; Gokhale, D.V. Lactic acid production from waste sugarcane bagasse derived cellulose. Green Chem. 2007, 9, 58–62. [Google Scholar] [CrossRef]
- Ahmad, A.; Banat, F.; Taher, H. A review on the lactic acid fermentation from low-cost renewable materials: Recent developments and challenges. Environ. Technol. Innov. 2020, 20, 101138. [Google Scholar] [CrossRef]
- Caplice, E. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef]
- Bruhn, A.; Brynning, G.; Johansen, A.; Lindegaard, M.S.; Sveigaard, H.H.; Aarup, B.; Fonager, L.; Andersen, L.L.; Rasmussen, M.B.; Larsen, M.M.; et al. Fermentation of sugar kelp (Saccharina latissima)—effects on sensory properties, and content of minerals and metals. J. Appl. Phycol. 2019, 31, 3175–3187. [Google Scholar] [CrossRef]
- Shobharani, P.; Halami, P.M.; Sachindra, N.M. Potential of marine lactic acid bacteria to ferment Sargassum sp. for enhanced anticoagulant and antioxidant properties. J. Appl. Microbiol. 2013, 114, 96–107. [Google Scholar] [CrossRef]
- John, R.P.; Anisha, G.S.; Nampoothiri, K.M.; Pandey, A. Direct lactic acid fermentation: Focus on simultaneous saccharification and lactic acid production. Biotechnol. Adv. 2009, 27, 145–152. [Google Scholar] [CrossRef]
- Mora-Villalobos, J.A.; Montero-Zamora, J.; Barboza, N.; Rojas-Garbanzo, C.; Usaga, J.; Redondo-Solano, M.; Schroedter, L.; Olszewska-Widdrat, A.; López-Gómez, J.P. Multi-product lactic acid bacteria fermentations: A review. Fermentation 2020, 6, 23. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Prebiotics: Concept, definition, criteria, methodologies, and products. In Handbook of Prebiotics; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–504. [Google Scholar]
- Martelli, F.; Favari, C.; Mena, P.; Guazzetti, S.; Ricci, A.; Del Rio, D.; Lazzi, C.; Neviani, E.; Bernini, V. Antimicrobial and fermentation potential of himanthalia elongata in food applications. Microorganisms 2020, 8, 248. [Google Scholar] [CrossRef]
- Uchida, M.; Miyoshi, T. Algal fermentation–he seed for a new fermentation industry of foods and related products. Jpn. Agric. Res. Q. 2013, 47, 53–63. [Google Scholar] [CrossRef]
- Cai, J.; Lovatelli, A.; Gamarro, E.G.; Geehan, J.; Lucente, D.; Mair, G.; Miao, W.; Reantaso, M.; Roubach, R.; Yuan, X.; et al. Seaweeds and Microalgae: An Overview for Unlocking Their Potential in Global Aquaculture Development; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Bajury, D.M.; Rawi, M.H.; Sazali, I.H.; Abdullah, A.; Sarbini, S.R. Prebiotic evaluation of red seaweed (Kappaphycus alvarezii) using in vitro colon model. Int. J. Food Sci. Nutr. 2017, 68, 821–828. [Google Scholar] [CrossRef]
- Fleurence, J. The enzymatic degradation of algal cell walls: A useful approach for improving protein accessibility? J. Appl. Phycol. 1999, 11, 313–314. [Google Scholar] [CrossRef]
- Hesseltine, C.W.; Wang, H.L. Traditional fermented foods. Biotechnol. Bioeng. 1967, 9, 275–288. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine Bioactives as Functional Food Ingredients: Potential to Reduce the Incidence of Chronic Diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef]
- Marrion, O.; Schwertz, A.; Fleurence, J.; Guéant, J.L.; Villaume, C. Improvement of the digestibility of the proteins of the red alga Palmaria palmata by physical processes and fermentation. Nahrung/Food 2003, 47, 339–344. [Google Scholar] [CrossRef]
- Suraiya, S.; Kim, J.H.; Tak, J.Y.; Siddique, M.P.; Young, C.J.; Kim, J.K.; Kong, I.S. Influences of fermentation parameters on lovastatin production by Monascus purpureus using Saccharina japonica as solid fermented substrate. LWT–Food Sci. Technol. 2018, 92, 1–9. [Google Scholar] [CrossRef]
- Wu, S.-C.; Wang, F.-J.; Pan, C.-L. Growth and Survival of Lactic Acid Bacteria during the Fermentation and Storage of Seaweed Oligosaccharides Solution. J. Mar. Sci. Technol. 2007, 15, 104–114. [Google Scholar] [CrossRef]
- Ramnani, P.; Chitarrari, R.; Tuohy, K.; Grant, J.; Hotchkiss, S.; Philp, K.; Campbell, R.; Gill, C.; Rowland, I. Invitro fermentation and prebiotic potential of novel low molecular weight polysaccharides derived from agar and alginate seaweeds. Anaerobe 2012, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Sun, Y.; Hu, L.; Liu, S.; Yu, H.; Xing, R.; Li, F.; Wang, X.; Li, P. In vitro prebiotic effects of seaweed polysaccharides. Chin. J. Oceanol. Limnol. 2017, 36, 926–932. [Google Scholar] [CrossRef]
- Ko, S.J.; Kim, J.; Han, G.; Kim, S.K.; Kim, H.G.; Yeo, I.; Ryu, B.; Park, J.W. Laminaria japonica combined with probiotics improves intestinal microbiota: A randomized clinical trial. J. Med. Food 2014, 17, 76–82. [Google Scholar] [CrossRef]
- Olsson, J.; Toth, G.B.; Oerbekke, A.; Cvijetinovic, S.; Wahlström, N.; Harrysson, H.; Steinhagen, S.; Kinnby, A.; White, J.; Edlund, U.; et al. Cultivation conditions affect the monosaccharide composition in Ulva fenestrata. J. Appl. Phycol. 2020, 32, 3255–3263. [Google Scholar] [CrossRef]
- Pacheco, D.; Cotas, J.; Rocha, C.P.; Araújo, G.S.; Figueirinha, A.; Gonçalves, A.M.M.; Bahcevandziev, K.; Pereira, L. Seaweeds’ carbohydrate polymers as plant growth promoters. Carbohydr. Polym. Technol. Appl. 2021, 2, 100097. [Google Scholar] [CrossRef]
- Pacheco, D.; Araújo, G.S.; Cotas, J.; Gaspar, R.; Neto, J.M.; Pereira, L. Invasive Seaweeds in the Iberian Peninsula: A Contribution for Food Supply. Mar. Drugs 2020, 18, 560. [Google Scholar] [CrossRef]
- Sharma, S.; Neves, L.; Funderud, J.; Mydland, L.T.; Øverland, M.; Horn, S.J. Seasonal and depth variations in the chemical composition of cultivated Saccharina latissima. Algal Res. 2018, 32, 107–112. [Google Scholar] [CrossRef]
- Ohno, M.; Largo, D.B.; Ikumoto, T. Growth rate, carrageenan yield and gel properties of culturedkappa-carrageenan producing red alga Kappaphycus alvarezzi (Doty) Doty in the subtropical waters of Shikoku, Japan. J. Appl. Phycol. 1994, 6, 1–5. [Google Scholar] [CrossRef]
- Ra, C.H.; Nguyen, T.H.; Jeong, G.-T.; Kim, S.-K. Evaluation of hyper thermal acid hydrolysis of Kappaphycus alvarezii for enhanced bioethanol production. Bioresour. Technol. 2016, 209, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Mutripah, S.; Meinita, M.D.N.; Kang, J.-Y.; Jeong, G.-T.; Susanto, A.; Prabowo, R.E.; Hong, Y.-K. Bioethanol production from the hydrolysate of Palmaria palmata using sulfuric acid and fermentation with brewer’s yeast. J. Appl. Phycol. 2014, 26, 687–693. [Google Scholar] [CrossRef]
- Schiener, P.; Zhao, S.; Theodoridou, K.; Carey, M.; Mooney-McAuley, K.; Greenwell, C. The nutritional aspects of biorefined Saccharina latissima, Ascophyllum nodosum and Palmaria palmata. Biomass Convers. Biorefinery 2017, 7, 221–235. [Google Scholar] [CrossRef]
- Villanueva, R.D.; Sousa, A.M.M.; Gonçalves, M.P.; Nilsson, M.; Hilliou, L. Production and properties of agar from the invasive marine alga, Gracilaria vermiculophylla (Gracilariales, Rhodophyta). J. Appl. Phycol. 2010, 22, 211–220. [Google Scholar] [CrossRef]
- Parada, J.L.; De Caire, G.Z.; De Mulé, M.C.Z.; De Cano, M.M.S. Lactic acid bacteria growth promoters from Spirulina platensis. Int. J. Food Microbiol. 1998, 45, 225–228. [Google Scholar] [CrossRef]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C.d.; Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [Google Scholar] [CrossRef]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health benefits of lactic acid bacteria (Lab) fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.A.; Lim, S.-R.; Kim, Y.; Park, J.M. Potentials of macroalgae as feedstocks for biorefinery. Bioresour. Technol. 2013, 135, 182–190. [Google Scholar] [CrossRef]
- Tan, I.S.; Lee, K.T. Enzymatic hydrolysis and fermentation of seaweed solid wastes for bioethanol production: An optimization study. Energy 2014, 78, 53–62. [Google Scholar] [CrossRef]
- Cascais, M.; Monteiro, P.; Pacheco, D.; Cotas, J.; Pereira, L.; Marques, J.C.; Gonçalves, A.M.M. Effects of Heat Treatment Processes: Health Benefits and Risks to the Consumer. Appl. Sci. 2021, 11, 8740. [Google Scholar] [CrossRef]
- Regal, A.L.; Alves, V.; Gomes, R.; Matos, J.; Bandarra, N.M.; Afonso, C.; Cardoso, C. Drying process, storage conditions, and time alter the biochemical composition and bioactivity of the anti-greenhouse seaweed Asparagopsis taxiformis. Eur. Food Res. Technol. 2020, 246, 781–793. [Google Scholar] [CrossRef]
- Gupta, S.; Cox, S.; Abu-Ghannam, N. Effect of different drying temperatures on the moisture and phytochemical constituents of edible Irish brown seaweed. LWT–Food Sci. Technol. 2011, 44, 1266–1272. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; Mcsorley, E.M.; Ross, R.P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed]
- Fabris, M.; Abbriano, R.M.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiuparambil, U.; Ray, P.; et al. Emerging Technologies in Algal Biotechnology: Toward the Establishment of a Sustainable, Algae-Based Bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Fievez, V.; Mao, S.; He, W.; Zhu, W. Dose and time response of ruminally infused algae on rumen fermentation characteristics, biohydrogenation and Butyrivibrio group bacteria in goats. J. Anim. Sci. Biotechnol. 2016, 7, 22. [Google Scholar] [CrossRef]
- Harun, R.; Yip, J.W.S.; Thiruvenkadam, S.; Ghani, W.A.W.A.K.; Cherrington, T.; Danquah, M.K. Algal biomass conversion to bioethanol–A step-by-step assessment. Biotechnol. J. 2014, 9, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Satokari, R. Modulation of Gut Microbiota for Health by Current and Next-Generation Probiotics. Nutrients 2019, 11, 1921. [Google Scholar] [CrossRef]
- Vidhya Hindu, S.; Chandrasekaran, N.; Mukherjee, A.; Thomas, J. A review on the impact of seaweed polysaccharide on the growth of probiotic bacteria and its application in aquaculture. Aquac. Int. 2019, 27, 227–238. [Google Scholar] [CrossRef]
- Kang, Y.M.; Lee, B.J.; Kim, J.I.; Nam, B.H.; Cha, J.Y.; Kim, Y.M.; Ahn, C.B.; Choi, J.S.; Choi, I.S.; Je, J.Y. Antioxidant effects of fermented sea tangle (Laminaria japonica) by Lactobacillus brevis BJ20 in individuals with high level of γ-GT: A randomized, double-blind, and placebo-controlled clinical study. Food Chem. Toxicol. 2012, 50, 1166–1169. [Google Scholar] [CrossRef] [PubMed]
- De Felice, F.G.; Velasco, P.T.; Lambert, M.P.; Viola, K.; Fernandez, S.J.; Ferreira, S.T.; Klein, W.L. Aβ oligomers induce neuronal oxidative stress through an N-methyl-D-aspartate receptor-dependent mechanism that is blocked by the Alzheimer drug memantine. J. Biol. Chem. 2007, 282, 11590–11601. [Google Scholar] [CrossRef]
- Vaisi-Raygani, A.; Rahimi, Z.; Zahraie, M.; Noroozian, M.; Pourmotabbed, A. Association between enzymatic and non-enzymatic antioxidant defense with Alzheimer Disease. Acta Med. Iran. 2007, 45, 271–276. [Google Scholar]
- Mapp, P.I.; Grootveld, M.C.; Blake, D.R. Hypoxia, oxidative stress and rheumatoid arthritis. Br. Med. Bull. 1995, 51, 419–436. [Google Scholar] [CrossRef]
- Smita, M.; Naidu, K.; Suryakar, A.N.; Swami, S.C.; Katkam, R.V.; Kumbar, K.M. Oxidative stress and antioxidant status in cervical cancer patients. Indian J. Clin. Biochem. 2007, 22, 140–144. [Google Scholar] [CrossRef]
- Wu, J.T.; Wu, L.L. Chronic Systemic Inflammation Leading Eventually to Myocardial Infarction, Stroke, COPD, Renal Failure and Cancer is Induced by Multiple Risk Factors. J. Biomed. Lab. Sci. 2007, 19, 1. [Google Scholar]
- Mayr, M.; Zhang, J.; Greene, A.S.; Gutterman, D.; Perloff, J.; Ping, P. Proteomics-based development of biomarkers in cardiovascular disease: Mechanistic, clinical, and therapeutic insights. Mol. Cell. Proteom. 2006, 5, 1853–1864. [Google Scholar] [CrossRef]
- Akhileshwar, V.; Patel, S.P.; Katyare, S.S. Diabetic cardiomyopathy and reactive oxygen species (ROS) related parameters in male and female rats: A comparative study. Indian J. Clin. Biochem. 2007, 22, 84–90. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Babu, P.A.; Suneetha, G.; Boddepalli, R.; Lakshmi, V.V.; Rani, T.S.; RamBabu, Y.; Srinivas, K. A database of 389 medicinal plants for diabetes. Bioinformation 2006, 1, 130–131. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yuan, H.; Zheng, J.C.; Liu, P.; Zhang, S.F.; Xu, J.Y.; Bai, L.M. Pathogenesis of Parkinson’s disease: Oxidative stress, environmental impact factors and inflammatory processes. Neurosci. Bull. 2007, 23, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Chilton, S.N.; Burton, J.P.; Reid, G.; Reid, G. Inclusion of fermented foods in food guides around the world. Nutrients 2015, 7, 390–404. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.H.; Jung, S.J.; Chae, S.W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef]
- Wilburn, J.R.; Ryan, E.P. Fermented Foods in Health Promotion and Disease Prevention: An Overview; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128023099. [Google Scholar]
- EFSA. Panel on Dietetic Products Scientific Opinion on the substantiation of health claims related to dietary fibre (ID 744, 745, 746, 748, 749, 753, 803, 810, 855, 1415, 1416, 4308, 4330) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2010, 8, 1735. [Google Scholar] [CrossRef]
- EFSA. Panel on Dietetic Products Scientific Opinion on the substantiation of health claims related to the replacement of mixtures of saturated fatty acids (SFAs) as present in foods or diets with mixtures of monounsaturated fatty acids (MUFAs) and/or mixtures of polyunsaturated fatty aci. EFSA J. 2011, 9, 2069. [Google Scholar] [CrossRef]
- Clark, M.J.; Slavin, J.L. The Effect of Fiber on Satiety and Food Intake: A Systematic Review. J. Am. Coll. Nutr. 2013, 32, 200–211. [Google Scholar] [CrossRef]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Aweya, J.J.; Huang, Z.-X.; Kang, Z.-Y.; Bai, Z.-H.; Li, K.-H.; He, X.-T.; Liu, Y.; Chen, X.-Q.; Cheong, K.-L. In vitro fermentation of Gracilaria lemaneiformis sulfated polysaccharides and its agaro-oligosaccharides by human fecal inocula and its impact on microbiota. Carbohydr. Polym. 2020, 234, 115894. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-H.R.; Chen, G.-W.; Pan, C.-L.; Lin, H.-T.V. Production of Ulvan Oligosaccharides with Antioxidant and Angiotensin-Converting Enzyme-Inhibitory Activities by Microbial Enzymatic Hydrolysis. Fermentation 2021, 7, 160. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Y.; Chen, X.-Q.; Aweya, J.J.; Cheong, K.-L. Catabolism of Saccharina japonica polysaccharides and oligosaccharides by human fecal microbiota. LWT 2020, 130, 109635. [Google Scholar] [CrossRef]
- Fu, X.; Cao, C.; Ren, B.; Zhang, B.; Huang, Q.; Li, C. Structural characterization and in vitro fermentation of a novel polysaccharide from Sargassum thunbergii and its impact on gut microbiota. Carbohydr. Polym. 2018, 183, 230–239. [Google Scholar] [CrossRef]
- Zheng, L.X.; Chen, X.Q.; Cheong, K.L. Current trends in marine algae polysaccharides: The digestive tract, microbial catabolism, and prebiotic potential. Int. J. Biol. Macromol. 2020, 151, 344–354. [Google Scholar] [CrossRef]
- Tang, Q.; Jin, G.; Wang, G.; Liu, T.; Liu, X.; Wang, B.; Cao, H. Current Sampling Methods for Gut Microbiota: A Call for More Precise Devices. Front. Cell. Infect. Microbiol. 2020, 10, 151. [Google Scholar] [CrossRef]
- Satoor, S.N.; Patil, D.P.; Kristensen, H.D.; Joglekar, M.V.; Shouche, Y.; Hardikar, A.A. Manipulation and Assessment of Gut Microbiome for Metabolic Studies. In Methods in Molecular Biology; Springer Nature: Berlin, Germany, 2014; pp. 449–469. [Google Scholar] [CrossRef]
- Ericsson, A.C.; Franklin, C.L. Manipulating the Gut Microbiota: Methods and Challenges: Figure 1. ILAR J. 2015, 56, 205–217. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Seaweed Species | Strains of Bacteria Involved in Seaweed Fermentation | Reference | |
|---|---|---|---|
| Chlorophyta | Ulva sp. | Lactobacillus brevis, L. plantarum, Lactobacillus casei, and Lactobacillus rhamnosus favour fermentation in non-saline substrate | [46] |
| Monostroma nitidum | Oligosaccharide extracts are potential substrate for Lactobacillus sp. | [55] | |
| Rhodophyta | Kappaphycus alvarezzi | Potential substrate for Bifidobacterium populations | [47] |
| Palmaria palmata (dulse) | Trichoderma pseudokoningii fermentation increase casein digestibility of 65.5% | [52] | |
| Gracilaria vermiculophylla | Potential substrate for L. casei B5201, D. hansenii Y5201 and Candida sp. Y5206 for lactic acid and ethanol production | [46] | |
| Gracilaria sp. | Potential substrate for Lactobacillus acidophilus and Lactobacillus plantarum for lactic acid production | [53] | |
| Gelidium sp. | Agar is a potential substrate for Bifidobacterium populations. An increase of short-chain fatty acids (SCFAs) has been detected | [56] | |
| Grateloupia filicina Eucheuma denticulatum (formerly Eucheuma spinosum) | Extracted polysaccharides are potential substrate for Bifidobacterium populations | [57] | |
| Phaeophyceae | Undaria pinnatifida | Potential substrate for LAB for lactic acid and ethanol production (thallus) | [46] |
| Saccharina japonica | Fermentation with Monascus purpureus and Monascus kaoliang showed an increase in phenolic, flavonoid, anti-diabetic and antioxidant effects | [22] | |
| Angiosperm | Lonicera japonica | Potential substrate for growth and survival of gut microbial lactic acid bacteria in humans | [58] |
| Species | Monosaccharide (%DW) | Polysaccharide (% DW) | Commercially Exploited | Ref. |
|---|---|---|---|---|
| Ulva sp. (Chlorophyta) | 26.64 | 36–43 | Cultivated; | [59] |
| Undaria pinnatifida (Phaeophyceae) | 12.4 | 8.7 | Cultivation and wild harvest; also, invasive species; | [60,61] |
| Saccharina latissimi (Phaeophyceae) | 35.2 | 17 | Cultivated and wild harvest; | [62] |
| Gracilaria fisheri (Rhodophyta) | 79.31 | 13.33 | Cultivated; | [62] |
| Kappaphycus alvarezzi (Rhodophyta) | 7 | 58.8 | Cultivated; | [63,64] |
| Palmaria palmata (Rhodophyta) | 11.6 | 35.4 | Cultivated; | [65,66] |
| Gracilaria vermiculophylla (Rhodophyta) | ND | 24–33 | Cultivated and wild harvest. Invasive species; | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, P.; Lomartire, S.; Cotas, J.; Pacheco, D.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweeds as a Fermentation Substrate: A Challenge for the Food Processing Industry. Processes 2021, 9, 1953. https://doi.org/10.3390/pr9111953
Monteiro P, Lomartire S, Cotas J, Pacheco D, Marques JC, Pereira L, Gonçalves AMM. Seaweeds as a Fermentation Substrate: A Challenge for the Food Processing Industry. Processes. 2021; 9(11):1953. https://doi.org/10.3390/pr9111953
Chicago/Turabian StyleMonteiro, Pedro, Silvia Lomartire, João Cotas, Diana Pacheco, João C. Marques, Leonel Pereira, and Ana M. M. Gonçalves. 2021. "Seaweeds as a Fermentation Substrate: A Challenge for the Food Processing Industry" Processes 9, no. 11: 1953. https://doi.org/10.3390/pr9111953
APA StyleMonteiro, P., Lomartire, S., Cotas, J., Pacheco, D., Marques, J. C., Pereira, L., & Gonçalves, A. M. M. (2021). Seaweeds as a Fermentation Substrate: A Challenge for the Food Processing Industry. Processes, 9(11), 1953. https://doi.org/10.3390/pr9111953