
Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review

Abstract
1. Introduction
- List the growth factors contained within human milk that are shown to have clinical importance in preventing necrotizing enterocolitis.
- For each growth factor, summarize levels in human milk and describe how levels change over time.
- Describe each growth factor’s biochemical and cellular mechanism for augmenting intestinal health.
- Summarize therapeutic trials of growth factors
- Describe donor milk processing and effects on human milk growth factors.
- Provide insight into the next steps required to establish therapeutic potential for each growth factor.
2. Methods
- exp Milk, Human/ or exp Infant Formula/ or (breast adj2 (milk or feed)).ti,ab,kw. or ((donor or mother*) adj3 milk).ti,ab,kw. (n = 31,935)
- (intestin* adj5 (development or pathology)).ti,ab,kw. (n = 7381)
- growth factor.ti,ab,kw. (n = 328,920)
- exp “Intercellular Signaling Peptides and Proteins”/ (n = 1,078,972)
- 3 or 4 (n = 1,216,365)
- 1 and 5 (n = 1180)
- exp Infant, Premature/ (n = 57,394)
- exp Infant, Premature, Diseases/ or exp Infant, Low Birth Weight/ or exp Infant, Very Low Birth Weight/ (n = 73,576)
- exp Infant, Premature/ or exp Infant, Premature, Diseases/ or exp Infant, Low Birth Weight/ or exp Infant, Very Low Birth Weight/ or (premature or preterm).ti,ab,kw. (n = 239,737)
- 7 or 8 or 9 (n = 239,737)
- 6 and 10 (n = 178)
- limit 11 to english language (n = 174)
- exp Enterocolitis, Necrotizing/ (n = 3683)
- 1 and 5 and 13 (n = 67)
- 12 or 14 (n = 193)
- limit 15 to english language (n = 191)
3. Narrative
3.1. Epidermal Growth Factor (EGF)
3.2. Heparin-Binding EGF-like Growth Factor (HB-EGF)
3.3. Insulin-like Growth Factor (IGF-1 and IGF-2)
3.4. Vascular Endothelial Growth Factor (VEGF)
3.5. Granulocyte Colony Stimulating Growth Factor (G-CSF)
3.6. Donor Milk and Holder Pasteurization’s Effects on Growth Hormone Levels in Human Milk
4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Neu, J.; Walker, W.A. Necrotizing enterocolitis. N. Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Battersby, C.; Santhalingam, T.; Costeloe, K.; Modi, N. Incidence of neonatal necrotising enterocolitis in high-income countries: A systematic review. Arch. Dis. Child. - Fetal Neonatal Ed. 2018, 103, F182–F189. [Google Scholar] [CrossRef] [PubMed]
- Bazacliu, C.; Neu, J. Necrotizing enterocolitis: Long term complications. Curr. Pediatr. Rev. 2019, 15, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Chowning, R.; Radmacher, P.; Lewis, S.L.; Serke, L.; Pettit, N.; Adamkin, D.H. A retrospective analysis of the effect of human milk on prevention of necrotizing enterocolitis and postnatal growth. J. Perinatol. 2016, 36, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.; Carroll, K. An Exclusively Human Milk Diet Reduces Necrotizing Enterocolitis. Breastfeed. Med. 2014, 9, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Al-Nafussi, A.I.; Wright, N.A. The effect of epidermal growth factor (EGF) on cell proliferation of the gastrointestinal mu-cosa in rodents. Virchows Archiv B. 1982, 40, 63. [Google Scholar] [CrossRef]
- Zhang, X.; Bandyopadhyay, S.; Araujo, L.P.; Tong, K.; Flores, J.; Laubitz, D.; Zhao, Y.; Yap, G.; Wang, J.; Zou, Q.; et al. Elevating EGFR-MAPK program by a nonconventional Cdc42 enhances intestinal epithelial survival and regeneration. JCI Insight 2020, 5, 5. [Google Scholar] [CrossRef]
- Basuroy, S.; Seth, A.; Elias, B.; Naren, A.P.; Rao, R. MAPK interacts with occludin and mediates EGF-induced prevention of tight junction disruption by hydrogen peroxide. Biochem. J. 2005, 393, 69–77. [Google Scholar] [CrossRef]
- Knoop, K.A.; Coughlin, P.E.; Floyd, A.N.; Ndao, I.M.; Hall-Moore, C.; Shaikh, N.; Gasparrini, A.J.; Rusconi, B.; Escobedo, M.; Good, M. Maternal activation of the EGFR prevents translocation of gut-residing pathogenic Escherichia coli in a model of late-onset neonatal sepsis. Proc. Natl. Acad. Sci. USA 2020. 117, 7941–7949. [CrossRef]
- Tang, X.; Liu, H.; Yang, S.; Li, Z.; Zhong, J.; Fang, R. Epidermal Growth Factor and Intestinal Barrier Function. Mediat. Inflamm. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Good, M.; Sodhi, C.P.; Egan, C.E.; Afrazi, A.; Jia, H.; Yamaguchi, Y.; Lu, P.; Branca, M.F.; Ma, C.; Prindle, T.; et al. Breast milk protects against the development of necrotizing enterocolitis through inhibition of Toll-like receptor 4 in the intestinal epithelium via activation of the epidermal growth factor receptor. Mucosal Immunol. 2015, 8, 1166–1179. [Google Scholar] [CrossRef]
- Sodhi, C.P.; Neal, M.D.; Siggers, R.; Sho, S.; Ma, C.; Branca, M.F.; Prindle, T., Jr.; Russo, A.M.; Afrazi, A.; Good, M.; et al. Intestinal Epithelial Toll-Like Receptor 4 Regulates Goblet Cell Development and Is Required for Necrotizing Enterocolitis in Mice. Gastroenterology 2012, 143, 708–718. [Google Scholar] [CrossRef]
- Dao, D.T.; Anez-Bustillos, L.; Adam, R.M.; Puder, M.; Bielenberg, D.R. Heparin-Binding Epidermal Growth Factor–Like Growth Factor as a Critical Mediator of Tissue Repair and Regeneration. Am. J. Pathol. 2018, 188, 2446–2456. [Google Scholar] [CrossRef]
- Feng, J.; Besner, G.E. Heparin-binding epidermal growth factor–like growth factor promotes enterocyte migration and prolif-eration in neonatal rats with necrotizing enterocolitis. J. Pediatric Surg. 2007, 42, 214–220. [Google Scholar] [CrossRef]
- Nishi, E.; Prat, A.; Hospital, V.; Elenius, K.; Klagsbrun, M. N-arginine dibasic convertase is a specific receptor for heparin-binding EGF-like growth factor that mediates cell migration. EMBO J. 2001, 20, 3342–3350. [Google Scholar] [CrossRef]
- Xia, G.; Rachfal, A.W.; Martin, A.E.; Besner, G.E. Upregulation of endogenous heparin-binding EGF-like growth factor (HB-EGF) expression after intestinal ischemia/reperfusion injury. J. Investig. Surg. 2003, 16, 57–63. [Google Scholar] [CrossRef]
- Yang, J.; Radulescu, A.; Chen, C.L.; Zhang, H.Y.; James, I.O.; Besner, G.E. Heparin-binding epidermal growth factor-like growth factor improves intestinal barrier function and reduces mortality in a murine model of peritonitis. Surg. 2012, 153, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.A.; Xia, G.; Mehta, V.B.; Glenn, S.; Michalsky, M.; Besner, G.E. Heparin-Binding EGF-Like Growth Factor (HB-EGF) Decreases Oxygen Free Radical Production In Vitro and In Vivo. Antioxidants Redox Signal. 2002, 4, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Mehta, V.B.; Besner, G.E. HB-EGF promotes angiogenesis in endothelial cells via PI3-kinase and MAPK signaling pathways. Growth Factors 2007, 25, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Radulescu, A.; Zorko, N.; Besner, G.E. Heparin-Binding EGF-Like Growth Factor Increases Intestinal Microvascular Blood Flow in Necrotizing Enterocolitis. Gastroenterol. 2009, 137, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Brigstock, D.; Besner, G.E. Heparin-Binding EGF-Like Growth Factor Is a Potent Vasodilator of Terminal Mesenteric Arterioles. J. Surg. Res. 2008, 144, 201. [Google Scholar] [CrossRef]
- Feng, J.; El-Assal, O.N.; Besner, G.E. Heparin-binding epidermal growth factor–like growth factor reduces intestinal apoptosis in neonatal rats with necrotizing enterocolitis. J. Pediatr. Surg. 2006, 41, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Alexander, A.N.; Carey, H.V. Oral IGF-I enhances nutrient and electrolyte absorption in neonatal piglet intestine. Am. J. Physiol. Liver Physiol. 1999, 277, G619–G625. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Liu, G.R.; Li, N.; Yuan, G. Insulin-like growth factor I reduces the occurrence of necrotizing enterocolitis by reducing inflammatory response and protecting intestinal mucosal barrier in neonatal rats model. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 4711–4719. [Google Scholar] [PubMed]
- Lee, O.H.; Bae, S.K.; Bae, M.H.; Lee, Y.M.; Moon, E.J.; Cha, H.J.; Kwon, Y.G.; Kim, K.W. Identification of angiogenic properties of insulin-like growth factor II in in vitro angiogenesis models. Br. J. Cancer 2000, 82, 385–391. [Google Scholar] [CrossRef]
- Shigematsu, S.; Yamauchi, K.; Nakajima, K.; Iijima, S.; Aizawa, T.; Hashizume, K. IGF-1 Regulates Migration and Angiogenesis of Human Endothelial Cells. Endocr. J. 1999, 46, S59–S62. [Google Scholar] [CrossRef] [PubMed]
- Akisu, M.; Durmaz, B.; Koroglu, O.A.; Unlubay, S.; Yalaz, M.; Akin, H.; Ates, U.; Baka, M.; Ozkinay, F.; Cogulu, O.; et al. The effects of IGF-1 and erythropoietin on apoptosis and telomerase activity in necrotizing enterocolitis model. Pediatr. Res. 2021, 90, 559–564. [Google Scholar] [CrossRef]
- Povsic, T.J.; Kohout, T.A.; Lefkowitz, R.J. β-Arrestin1 mediates insulin-like growth factor 1 (IGF-1) activation of phos-phatidylinositol 3-kinase (PI3K) and anti-apoptosis. J. Biol. Chem. 2003, 278, 51334–51339. [Google Scholar] [CrossRef]
- Takeshita, K.; Satoh, M.; Ii, M.; Silver, M.; Limbourg, F.; Mukai, Y.; Rikitake, Y.; Radtke, F.; Gridley, T.; Losordo, D.; et al. Critical Role of Endothelial Notch1 Signaling in Postnatal Angiogenesis. Circ. Res. 2007, 100, 70–78. [Google Scholar] [CrossRef]
- Juul, S.E.; Ledbetter, D.J.; Joyce, A.E.; Dame, C.; Christensen, R.D.; Zhao, Y.; DeMarco, V. Erythropoietin acts as a trophic factor in neonatal rat intestine. Gut 2001, 49, 182–189. [Google Scholar] [CrossRef]
- Shiou, S.R.; Yu, Y.; Chen, S.; Ciancio, M.J.; Petrof, E.O.; Sun, J.; Claud, E.C. Erythropoietin Protects Intestinal Epithelial Barrier Function and Lowers the Incidence of Experimental Neonatal Necrotizing Enterocolitis. J. Biol. Chem. 2011, 286, 12123–12132. [Google Scholar] [CrossRef]
- Claud, E.C.; Savidge, T.; Walker, W.A. Modulation of human intestinal epithelial cell IL-8 secretion by human milk factors. Pediatr Res. 2003, 53, 419–425. [Google Scholar] [CrossRef]
- Canpolat, F.E.; Yurdakök, M.; Özsoy, Ş.; Hazıroğlu, R.; Korkmaz, A. Protective effects of recombinant human granulocyte colony stimulating factor in a rat model of necrotizing enterocolitis. Pediatr. Surg. Int. 2006, 22, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, D.A.; Lunøe, M.; Du, Y.; Christensen, R.D. Granulocyte colony-stimulating factor is present in human milk and its receptor is present in human fetal intestine. Pediatr. 2000, 105, e7. [Google Scholar] [CrossRef]
- Calhoun, D.A.; Maheshwari, A.; Christensen, R.D. Recombinant granulocyte colony-stimulating factor administered enterally to neonates is not absorbed. Pediatr. 2003, 112, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Gersting, J.A.; Christensen, R.D.; Calhoun, D.A. Effects of Enterally Administering Granulocyte Colony-Stimulating Factor to Suckling Mice. Pediatr. Res. 2004, 55, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B.; Fituch, C.C.; Williams, C.S.; Hurst, N.M.; Schanler, R.J. Increased Epidermal Growth Factor Levels in Human Milk of Mothers with Extremely Premature Infants. Pediatr. Res. 2003, 54, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Idota, T. The concentration of epidermal growth factor in Japanese mother’s milk. J. Nutr. Sci. Vitaminol. 1995, 41, 241–251. [Google Scholar] [CrossRef]
- Michalsky, M.; Lara-Marquez, M.L.; Chun, L.; Besner, G. Heparin-binding EGF-like growth factor is present in human amniotic fluid and breast milk. J. Pediatr. Surg. 2002, 37, 1–6. [Google Scholar] [CrossRef]
- Ozgurtas, T.; Aydin, I.; Turan, O.; Koc, E.; Hirfanoglu, I.M.; Acikel, C.H.; Akyol, M.; Erbil, M.K. Vascular endothelial growth factor, basic fibroblast growth factor, insulin-like growth factor-I and platelet-derived growth factor levels in human milk of mothers with term and preterm neonates. Cytokine 2010, 50, 192–194. [Google Scholar] [CrossRef]
- Elmlinger, M.W.; Hochhaus, F.; Loui, A.; Frommer, K.W.; Obladen, M.; Ranke, M.B. Insulin-Like Growth Factors and Binding Proteins in Early Milk from Mothers of Preterm and Term Infants. Horm. Res. Paediatr. 2007, 68, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Siafakas, C.G.; Anatolitou, F.; Fusunyan, R.D.; Walker, W.A.; Sanderson, I.R. Vascular Endothelial Growth Factor (VEGF) Is Present in Human Breast Milk and Its Receptor Is Present on Intestinal Epithelial Cells. Pediatr. Res. 1999, 45, 652–657. [Google Scholar] [CrossRef] [PubMed]
- Kling, P.J.; Sullivan, T.M.; Roberts, R.A.; Philipps, A.F.; Koldovsky, O. Human Milk as a Potential Enteral Source of Erythropoietin. Pediatr. Res. 1998, 43, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Isani, M.; Illingworth, L.; Herman, E.; Schmidt, M.; Barron, L.; Bowling, J.; Elizee, M.; Bai, I.; Gayer, C.; Grishin, A.; et al. Soybean-derived recombinant human epidermal growth factor protects against experimental necrotizing enterocolitis. J. Pediatr. Surg. 2018, 53, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Su, H.H.; Chen, J.C.; Chen, P.T. Production of recombinant human epidermal growth factor in Bacillus subtilis. J. Taiwan Inst. Chem. Eng. 2020, 106, 86–91. [Google Scholar] [CrossRef]
- Hofmann, G.E.; Abramowicz, J.S. Epidermal Growth Factor (Egf) Concentrations in Amniotic Fluid and Maternal Urine During Pregnancy. Acta Obstet. et Gynecol. Scand. 1990, 69, 217–221. [Google Scholar] [CrossRef]
- Varner, M.; Dildy, G.; Hunter, C.; Dudley, D.; Clark, S.; Mitchell, M. Amniotic fluid epidermal growth factor levels in normal and abnormal pregnancies. J. Soc. Gynecol. Investig. JSGI. 1996, 3, 17–19. [Google Scholar] [CrossRef]
- Olsen, P.S.; Poulsen, S.S.; Kirkegaard, P.; Nexø, E. Role of submandibular saliva and epidermal growth factor in gastric cytoprotection. Gastroenterol. 1984, 87, 103–108. [Google Scholar] [CrossRef]
- Untalan, P.B.; Keeney, S.E.; Palkowetz, K.H.; Rivera, A.; Goldman, A. Heat Susceptibility of Interleukin-10 and Other Cytokines in Donor Human Milk. Breastfeed. Med. 2009, 4, 137–144. [Google Scholar] [CrossRef]
- Dembinski, A.B.; Johnson, L.R. Effect of Epidermal Growth Factor on the Development of Rat Gastric Mucosa. Endocrinol. 1985, 116, 90–94. [Google Scholar] [CrossRef]
- Malo, C.; Ménard, D. Influence of Epidermal Growth Factor on the Development of Suckling Mouse Intestinal Mucosa. Gastroenterol. 1982, 83, 28–35. [Google Scholar] [CrossRef]
- Halpern, M.D.; Dominguez, J.A.; Dvorakova, K.; Holubec, H.; Williams, C.S.; Meza, Y.G.; Ruth, M.C.; Dvorak, B. Ileal Cytokine Dysregulation in Experimental Necrotizing Enterocolitis Is Reduced by Epidermal Growth Factor. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Xu, R. Stability and distribution of orally administered epidermal growth factor in neonatal pigs. Life Sci. 1998, 63, 809–820. [Google Scholar] [CrossRef]
- Besner, G.; Higashiyama, S.; Klagsbrun, M. Isolation and characterization of a macrophage-derived heparin-binding growth factor. Cell Regul. 1990, 1, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, B. Milk Epidermal Growth Factor and Gut Protection. J. Pediatr. 2010, 156, S31–S35. [Google Scholar] [CrossRef] [PubMed]
- El-Assal, O.N.; Besner, G.E. HB-EGF enhances restitution after intestinal ischemia/reperfusion via PI3K/Akt and MEK/ERK1/2 activation. Gastroenterology 2005, 129, 609–625. [Google Scholar] [CrossRef]
- Rocourt, D.V.; Mehta, V.B.; Besner, G.E. Heparin-binding EGF-like growth factor decreases inflammatory cytokine expression after intestinal ischemia/reperfusion injury. J. Surg. Res. 2007, 139, 269–273. [Google Scholar] [CrossRef][Green Version]
- Chen, C.L.; Yu, X.; James, I.O.-A.; Zhang, H.Y.; Yang, J.; Radulescu, A.; Zhou, Y.; Besner, G.E. Heparin-binding EGF-like growth factor protects intestinal stem cells from injury in a rat model of necrotizing enterocolitis. Lab. Investig. 2011, 92, 331–344. [Google Scholar] [CrossRef]
- Sara, V.R.; Hall, K. Insulin-like growth factors and their binding proteins. Physiol. Rev. 1990, 70, 591–614. [Google Scholar] [CrossRef]
- He, B.; Zhang, N.; Jia, Y.; Sun, Q.; Zhao, R. Glucocorticoid receptor-mediated insulin-like growth factor-I transcriptional regulation in BeWo trophoblast cells before and after syncytialisation. Steroids 2016, 115, 26–33. [Google Scholar] [CrossRef]
- Lund, P.K.; Moats-Staats, B.M.; Hynes, M.A.; Simmons, J.G.; Jansen, M.; D’Ercole, A.J.; Van Wyk, J.J. Somatomedin-C/insulin-like growth factor-I and insulin-like growth factor-II mRNAs in rat fetal and adult tissues. J. Biol. Chem. 1986, 261, 14539–14544. [Google Scholar] [CrossRef]
- Hellström, A.; Ley, D.; Hansen-Pupp, I.; Hallberg, B.; Ramenghi, L.A.; Löfqvist, C.; Smith, L.E.H.; Hård, A.L. Role of insulinlike growth factor 1 in fetal development and in the early postnatal life of premature infants. Am. J. Perinatol. 2016, 33, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, K.; Itoh, K.; Kuroume, T. Levels of insulin-like growth factor I in full- and preterm human milk in comparison to levels in cow’s milk and in milk formulas. Biol Neonate. 1990, 58, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Milsom, S.R.; Blum, W.F.; Gunn, A. Temporal Changes in Insulin-Like Growth Factors I and II and in Insulin-Like Growth Factor Binding Proteins 1, 2, and 3 in Human Milk. Horm. Res. 2008, 69, 307–311. [Google Scholar] [CrossRef]
- Prosser, C.G. Insulin-like growth factors in milk and mammary gland. J. Mammary Gland. Biol. Neoplasia 1996, 1, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Goelz, R.; Hihn, E.; Hamprecht, K.; Dietz, K.; Jahn, G.; Poets, C.; Elmlinger, M. Effects of Different CMV-Heat-Inactivation-Methods on Growth Factors in Human Breast Milk. Pediatr. Res. 2009, 65, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Van Landeghem, L.; Santoro, M.A.; Mah, A.T.; Krebs, A.E.; Dehmer, J.J.; McNaughton, K.K.; Helmrath, M.A.; Magness, S.T.; Lund, P.K. IGF1 stimulates crypt expansion via differential activation of 2 intestinal stem cell populations. FASEB J. 2015, 29, 2828–2842. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, H.R.; Ohneda, K.; Keku, T.O.; D’Ercole, A.J.; Fuller, C.R.; Williams, K.L.; Lund, P.K. Reduction of spontaneous and irradiation-induced apoptosis in small intestine of IGF-I transgenic mice. Am. J. Physiol. Liver Physiol. 2002, 283, G457–G464. [Google Scholar] [CrossRef]
- Qiu, W.; Leibowitz, B.; Zhang, L.; Yu, J. Growth factors protect intestinal stem cells from radiation-induced apoptosis by suppressing PUMA through the PI3K/AKT/p53 axis. Oncogene 2009, 29, 1622–1632. [Google Scholar] [CrossRef]
- Baregamian, N.; Song, J.; Jeschke, M.G.; Evers, B.M.; Chung, D.H. IGF-1 Protects Intestinal Epithelial Cells From Oxidative Stress-Induced Apoptosis. J. Surg. Res. 2006, 136, 31–37. [Google Scholar] [CrossRef]
- Holgersen, K.; Gao, X.; Narayanan, R.; Gaur, T.; Carey, G.; Barton, N.; Pan, X.; Muk, T.; Thymann, T.; Sangild, P.T. Supplemental Insulin-Like Growth Factor-1 and Necrotizing Enterocolitis in Preterm Pigs. Front. Pediatr. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Corpeleijn, W.E.; van Vliet, I.; de Gast-Bakker, D.-A.H.; van der Schoor, S.R.; Alles, M.S.; Hoijer, M.; Tibboel, D.; van Goudoever, J.B. Effect of Enteral IGF-1 Supplementation on Feeding Tolerance, Growth, and Gut Permeability in Enterally Fed Premature Neonates. J. Pediatr. Gastroenterol. Nutr. 2008, 46, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Hellström, A.; Ley, D.; Hallberg, B.; Löfqvist, C.; Hansen-Pupp, I.; Ramenghi, L.A.; Borg, J.; Smith, L.E.H.; Hard, A.-L. IGF-1 as a Drug for Preterm Infants: A Step-Wise Clinical Development. Curr. Pharm. Des. 2018, 23, 5964–5970. [Google Scholar] [CrossRef] [PubMed]
- Ley, D.; Hallberg, B.; Hansen-Pupp, I.; Dani, C.; Ramenghi, L.A.; Marlow, N.; Beardsall, K.; Bhatti, F.; Dunger, D.; Higginson, J.D.; et al. rhIGF-1/rhIGFBP-3 in Preterm Infants: A Phase 2 Randomized Controlled Trial. J. Pediatr. 2019, 206, 56–65.e8. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Adamis, A.P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef]
- Kobata, R.; Tsukahara, H.; Ohshima, Y.; Ohta, N.; Tokuriki, S.; Tamura, S.; Mayumi, M. High levels of growth factors in human breast milk. Early Hum. Dev. 2008, 84, 67–69. [Google Scholar] [CrossRef]
- Loui, A.; Eilers, E.; Strauss, E.; Pohl-Schickinger, A.; Obladen, M.; Koehne, P. Vascular Endothelial Growth Factor (VEGF) and Soluble VEGF Receptor 1 (sFlt-1) Levels in Early and Mature Human Milk from Mothers of Preterm versus Term Infants. J. Hum. Lact. 2012, 28, 522–528. [Google Scholar] [CrossRef]
- Ozgurtas, T.; Aydin, I.; Turan, O.; Koç, E.; Hirfanoglu, I.M.; Acikel, C.H.; Akyol, M.; Serdar, M.; Erbil, K.M. Soluble Vascular Endothelial Growth Factor Receptor 1 in Human Breast Milk. Horm. Res. Paediatr. 2011, 76, 17–21. [Google Scholar] [CrossRef]
- Oladipupo, S.; Hu, S.; Kovalski, J.; Yao, J.; Santeford, A.; Sohn, R.E.; Shohet, R.; Maslov, K.; Wang, L.; Arbeit, J.M. VEGF is essential for hypoxia-inducible factor-mediated neovascularization but dispensable for endothelial sprouting. Proc. Natl. Acad. Sci. 2011, 108, 13264–13269. [Google Scholar] [CrossRef]
- Shibuya, M.; Claesson-Welsh, L. Signal transduction by VEGF receptors in regulation of angiogenesis and lymphangio-genesis. Exp. Cell Res. 2006, 312, 549–560. [Google Scholar] [CrossRef]
- Stark, A.; Dammann, C.; Nielsen, H.C.; Volpe, M.V. A Pathogenic Relationship of Bronchopulmonary Dysplasia and Retinopathy of Prematurity? A Review of Angiogenic Mediators in Both Diseases. Front. Pediatr. 2018, 6, 125. [Google Scholar] [CrossRef]
- Yan, X.; Managlia, E.; Liu, S.X.; Tan, X.-D.; Wang, X.; Marek, C.; De Plaen, I.G. Lack of VEGFR2 signaling causes maldevel-opment of the intestinal microvasculature and facilitates necrotizing enterocolitis in neonatal mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 2016, 310, G716–G725. [Google Scholar] [CrossRef]
- Yan, X.; Managlia, E.; Tan, X.D.; De Plaen, I.G. Prenatal inflammation impairs intestinal microvascular development through a TNF-dependent mechanism and predisposes newborn mice to necrotizing enterocolitis. Am. J. Physiol. Liver Physiol. 2019, 317, G57–G66. [Google Scholar] [CrossRef]
- Bowker, R.M.; Yan, X.; Managlia, E.; Liu, S.X.; Marek, C.; Tan, X.D.; De Plaen, I.G. Dimethyloxalylglycine preserves the intestinal microvasculature and protects against intestinal injury in a neonatal mouse NEC model: Role of VEGF signaling. Pediatr. Res. 2018, 83, 545–553. [Google Scholar] [CrossRef]
- Juul, S.E.; Yachnis, A.T.; Christensen, R.D. Tissue distribution of erythropoietin and erythropoietin receptor in the developing human fetus. Early Hum. Dev. 1998, 52, 235–249. [Google Scholar] [CrossRef]
- Halvorsen, S.; Finne, P.H. Erythropoietin production in the human fetus and newborn. Ann. N. Y. Acad. Sci. 1968, 149, 576–577. [Google Scholar] [CrossRef] [PubMed]
- Teramo, K.A.; Widness, J.A. Increased fetal plasma and amniotic fluid erythropoietin concentrations: Markers of intrauterine hypoxia. Neonatology 2009, 95, 105–116. [Google Scholar] [CrossRef]
- Juul, S.E.; Zhao, Y.; Dame, J.B.; Du, Y.; Hutson, A.D.; Christensen, R.D. Origin and Fate of Erythropoietin in Human Milk. Pediatr. Res. 2000, 48, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shiou, S.R.; Guo, Y.; Lü, L.; Westerhoff, M.; Sun, J.; Petrof, E.O.; Claud, E.C. Erythropoietin Protects Epithelial Cells from Excessive Autophagy and Apoptosis in Experimental Neonatal Necrotizing Enterocolitis. PLOS ONE 2013, 8, e69620. [Google Scholar] [CrossRef]
- Wang, Y.; Song, J.; Sun, H.; Xu, F.; Li, K.; Nie, C.; Zhang, X.; Peng, X.; Xia, L.; Shen, Z.; et al. Erythropoietin prevents necrotizing enterocolitis in very preterm infants: A randomized controlled trial. J. Transl. Med. 2020, 18, 1–9. [Google Scholar] [CrossRef]
- Han, S.M.; Hong, C.R.; Knell, J.; Edwards, E.M.; Morrow, K.A.; Soll, R.F.; Modi, B.P.; Horbar, J.D.; Jaksic, T. Trends in incidence and outcomes of necrotizing enterocolitis over the last 12 years: A multicenter cohort analysis. J. Pediatr. Surg. 2020, 55, 998–1001. [Google Scholar] [CrossRef]
- Juul, S.E.; Comstock, B.A.; Wadhawan, R.; Mayock, D.E.; Courtney, S.E.; Robinson, T.; Ahmad, K.A.; Bendel-Stenzel, E.; Baserga, M.; LaGamma, E.F.; et al. A Randomized Trial of Erythropoietin for Neuroprotection in Preterm Infants. N. Engl. J. Med. 2020, 382, 233–243. [Google Scholar] [CrossRef]
- Hosseini, M.; Azampour, H.; Raeisi, S.; Behtari, M.; Valizadeh, H.; Saboohi, R. The effects of enteral artificial amniotic fluid-containing erythropoietin on short term outcomes of preterm infants. Turk. J. Pediatr. 2019, 61, 392–398. [Google Scholar] [CrossRef]
- El-Ganzoury, M.M.; Awad, H.A.; El-Farrash, R.A.; El-Gammasy, T.M.; Ismail, E.A.; Mohamed, H.E.; Suliman, S.M. Enteral Granulocyte-Colony Stimulating Factor and Erythropoietin Early in Life Improves Feeding Tolerance in Preterm Infants: A Randomized Controlled Trial. J. Pediatr. 2014, 165, 1140–1145.e1. [Google Scholar] [CrossRef]
- Calhoun, D.A. Granulocyte colony-stimulating factor in preterm and term pregnancy, parturition, and intra-amniotic infection. Obstet. Gynecol. 2001, 97, 229–234. [Google Scholar] [CrossRef]
- Bailie, K.E.M.; Irvine, A.E.; Bridges, J.M.; McClure, B.G. Granulocyte and Granulocyte-Macrophage Colony-Stimulating Factors in Cord and Maternal Serum at Delivery. Pediatr. Res. 1994, 35, 164–168. [Google Scholar] [CrossRef]
- Saito, S.; Kasahara, T.; Kato, Y.; Ishihara, Y.; Ichijo, M. Elevation of amniotic fluid interleukin 6 (IL-6), IL-8 and granulocyte colony stimulating factor (G-CSF) in term and preterm parturition. Cytokine 1993, 5, 81–88. [Google Scholar] [CrossRef]
- Panopoulos, A.D.; Watowich, S.S. Granulocyte colony-stimulating factor: Molecular mechanisms of action during steady state and ‘emergency’ hematopoiesis. Cytokine 2008, 42, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, D.A.; Donnelly, W.H.; Du, Y.; Dame, J.B.; Li, Y.; Christensen, R.D. Distribution of Granulocyte Colony-Stimulating Factor (G-CSF) and G-CSF-Receptor mRNA and Protein in the Human Fetus. Pediatr. Res. 1999, 46, 333–338. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Calhoun, D.A.; Lunøe, M.; Du, Y.; Staba, S.L.; Christensen, R.D. Concentrations of Granulocyte Colony-Stimulating Factor in Human Milk after in Vitro Simulations of Digestion. Pediatr. Res. 1999, 46, 767. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, Y.G.; Duan, X.N.; Liu, Y.H.; Zhao, J.X.; Xu, L.; Ye, J.M. Role of granulocyte colony-stimulating factor in paclitaxel-induced intestinal barrier breakdown and bacterial translocation in rats. Chin. Med J. 2011, 124, 1870–1875. [Google Scholar]
- Klinke, M.; Vincent, D.; Trochimiuk, M.; Appl, B.; Tiemann, B.; Reinshagen, K.; Raluy, L.P.; Boettcher, M. Development of an improved murine model of necrotizing enterocolitis shows the importance of neutrophils in NEC pathogenesis. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Christensen, R.D.; Yoder, B.A.; Baer, V.L.; Snow, G.L.; Butler, A. Early-Onset Neutropenia in Small-for-Gestational-Age Infants. Pediatr. 2015, 136, 136. [Google Scholar] [CrossRef] [PubMed]
- Pantalone, J.M.; Liu, S.; Olaloye, O.O.; Prochaska, E.C.; Yanowitz, T.; Riley, M.M.; Buland, J.R.; Brozanski, B.S.; Good, M.; Konnikova, L. Gestational Age-Specific Complete Blood Count Signatures in Necrotizing Enterocolitis. Front. Pediatr. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Fleit, H.B.; Golightly, M.G.; La Gamma, E.F. In vivo Effect of Recombinant Human Granulocyte Colony-Stimulating Factor on Phagocytic Function and Oxidative Burst Activity in Septic Neutropenic Neonates. Neonatol. 2004, 86, 48–54. [Google Scholar] [CrossRef]
- Chaudhuri, J.; Mitra, S.; Mukhopadhyay, D.; Chakraborty, S.; Chatterjee, S. Granulocyte colony-stimulating factor for preterms with sepsis and neutropenia: A randomized controlled trial. J. Clin. Neonatol. 2012, 1, 202–206. [Google Scholar] [CrossRef]
- Lee, J.A.; Sauer, B.; Tuminski, W.; Cheong, J.; Fitz-Henley, J.; Mayers, M.; Ezuma-Igwe, C.; Arnold, C.; Hornik, C.P.; Clark, R.H.; et al. Effectiveness of Granulocyte Colony-Stimulating Factor in Hospitalized Infants with Neutropenia. Am. J. Perinatol. 2016, 34, 458–464. [Google Scholar] [CrossRef]
- Kuhn, P.; Messer, J.; Paupe, A.; Espagne, S.; Kacet, N.; Mouchnino, G.; Klosowski, S.; Krim, G.; Lescure, S.; Le Bouedec, S.; et al. A Multicenter, Randomized, Placebo-Controlled Trial of Prophylactic Recombinant Granulocyte-Colony Stimulating Factor in Preterm Neonates with Neutropenia. J. Pediatr. 2009, 155, 324–330.e1. [Google Scholar] [CrossRef]
- Canpolat, F.E.; Yurdakök, M.; Korkmaz, A.; Yiğit, Ş.; Tekinalp, G. Enteral granulocyte colony-stimulating factor for the treatment of mild (stage I) necrotizing enterocolitis: A placebo-controlled pilot study. J. Pediatr. Surg. 2006, 41, 1134–1138. [Google Scholar] [CrossRef]
- Committee on Nutrition; Section on Breast Feeding; Committee on Fetus and Newborn. Donor human milk for the high-risk infant: Preparation, safety, and usage options in the United States. Pediatrics 2017, 139, e20163440. [Google Scholar] [CrossRef]
- Meier, P.; Patel, A.; Esquerra-Zwiers, A. Donor human milk update: Evidence, mechanisms, and priorities for research and practice. J. Pediatrics 2017, 180, 15–21. [Google Scholar] [CrossRef]
- Valentine, C.J.; Morrow, G.; Reisinger, A.; Dingess, K.A.; Morrow, A.L.; Rogers, L.K. Lactational Stage of Pasteurized Human Donor Milk Contributes to Nutrient Limitations for Infants. Nutr. 2017, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Peila, C.; Moro, G.E.; Bertino, E.; Cavallarin, L.; Giribaldi, M.; Giuliani, F.; Cresi, F.; Coscia, A. The Effect of Holder Pasteurization on Nutrients and Biologically-Active Components in Donor Human Milk: A Review. Nutr. 2016, 8, 477. [Google Scholar] [CrossRef] [PubMed]
- Ewaschuk, J.B.; Unger, S.; O’Connor, D.; Stone, D.L.; Harvey, S.; Clandinin, M.T.; Field, C. Effect of pasteurization on selected immune components of donated human breast milk. J. Perinatol. 2011, 31, 593–598. [Google Scholar] [CrossRef] [PubMed]
- A Pitino, M.; Unger, S.; Doyen, A.; Pouliot, Y.; Aufreiter, S.; Stone, D.; Kiss, A.; O’Connor, D.L. High Hydrostatic Pressure Processing Better Preserves the Nutrient and Bioactive Compound Composition of Human Donor Milk. J. Nutr. 2019, 149, 497–504. [Google Scholar] [CrossRef]
- Sullivan, S.; Schanler, R.J.; Kim, J.H.; Patel, A.; Trawöger, R.; Kiechl-Kohlendorfer, U.; Chan, G.M.; Blanco, C.L.; Abrams, S.; Cotten, C.M.; et al. An Exclusively Human Milk-Based Diet Is Associated with a Lower Rate of Necrotizing Enterocolitis than a Diet of Human Milk and Bovine Milk-Based Products. J. Pediatr. 2010, 156, 562–567. [Google Scholar] [CrossRef]
- Lucas, A.; Cole, T. Breast milk and neonatal necrotising enterocolitis. Lancet 1990, 336, 1519–1523. [Google Scholar] [CrossRef]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.; Pierro, A. Human breast milk exosomes attenuate intestinal damage. Pediatr. Surg. Int. 2020, 36, 155–163. [Google Scholar] [CrossRef]
- Johnson, T.J.; Berenz, A.; Wicks, J.; Esquerra-Zwiers, A.; Sulo, K.S.; Gross, M.E.; Szotek, J.; Meier, P.; Patel, A.L. The Economic Impact of Donor Milk in the Neonatal Intensive Care Unit. J. Pediatr. 2020, 224, 57–65. [Google Scholar] [CrossRef]
- Boyd, C.A.; Quigley, M.A.; Brocklehurst, P. Donor breast milk versus infant formula for preterm infants: Systematic review and meta-analysis. Arch. Dis. Child. - Fetal Neonatal Ed. 2007, 92, F169–F175. [Google Scholar] [CrossRef] [PubMed]


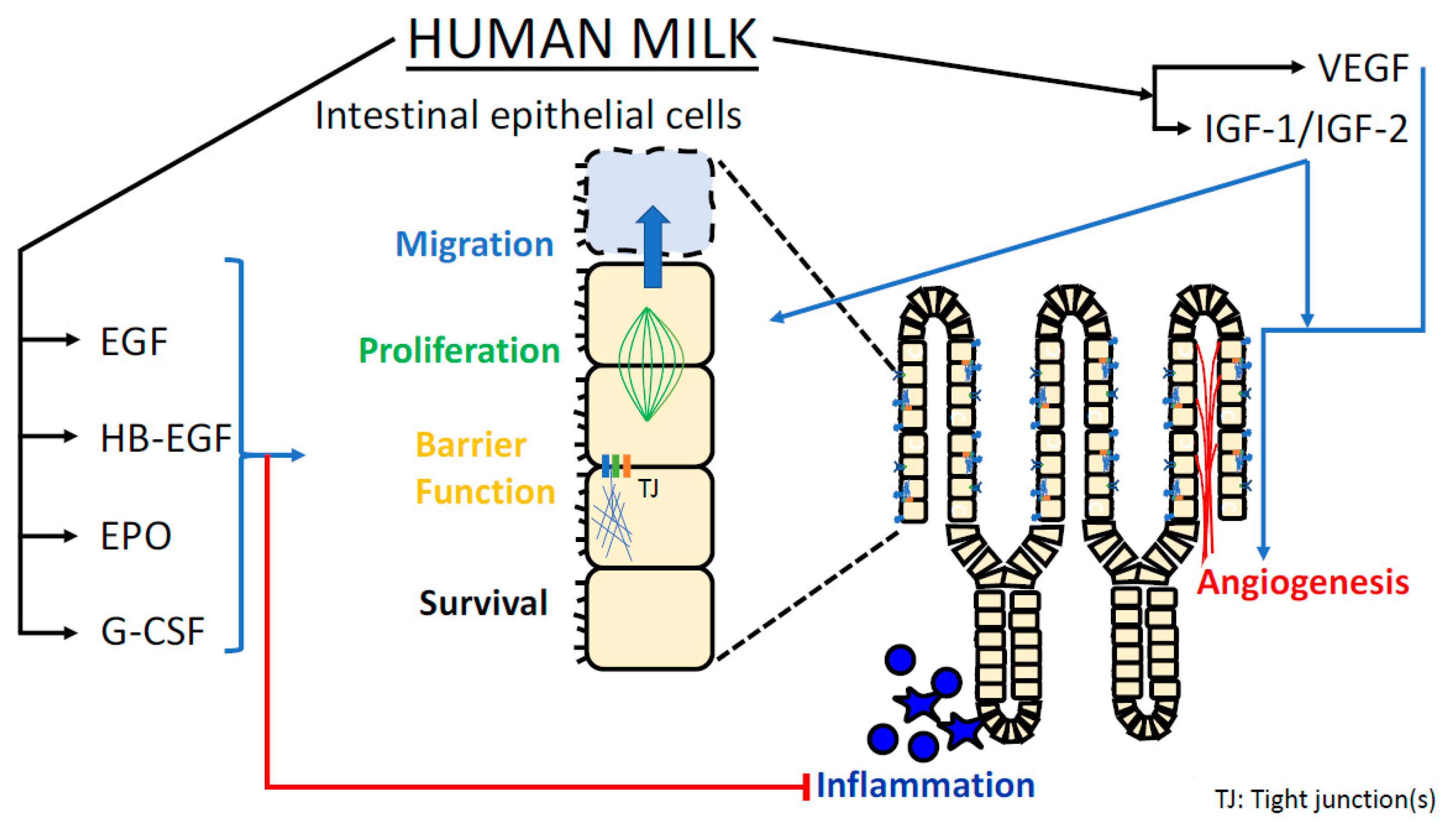{kind=link}
| Hormone | Role in NEC Protection | Stable | |
|---|---|---|---|
| Orally | Holder Pasteurization | ||
| EGF | Increases proliferation of intestinal epithelium [6] Enhances appropriate cellular migration in intestinal healing [7] Optimizes epithelial barrier function [8,9,10] Attenuates TLR4-mediated inflammatory response [11,12] | Yes | Yes |
| HB-EGF | Stimulates intestinal cell proliferation [13,14,15,16] Promotes epithelial barrier function [17] Reduces production of ROS [18] Augments microvascular perfusion (angiogenesis, vasodilation) [19,20,21] Attenuates apoptosis [22] | Yes | Yes |
| IGF-1/ IGF-2 | Promotes epithelial barrier function [23,24] Reduces inflammatory response [24] Promotes mucosal microvascular development [25,26] Attenuates apoptosis [27,28] | Yes | Partially |
| VEGF | Promotes mucosal microvascular development [29] | Not reported | |
| EPO | Trophic effects on intestinal villi [30] Promotes epithelial barrier function [31] Reduces epithelial cell inflammatory response [32] | Yes | Partially |
| G-CSF | Accelerates mucosal healing [33] Trophic effect on intestinal villi [34,35,36] | Yes | Partially |
| Growth Factor | Preterm vs. Term Milk | Statistical Differences between Groups | Early vs. Late Lactation | Statistical Differences between Stages of Lactation |
|---|---|---|---|---|
| EGF | Prem: 16–17 µg/100 mL Pre/Term: 10–11 µg/100 mL [37] | Yes | DOL 3-5: ~15 µg/100 mL DOL 31-60: ~8 µg/100 mL [38] | Yes (among both term and preterm populations) |
| HBEGF | Range: 2×10−3 –2.3×10−2 g/100 mL [39] Gestational age not reported | Not reported | No clear trend | No |
| IGF1 | Term: 0.031 µg/100 mL [40] Prem: 0.419 µg/100 mL [40] At DOL 28 | Yes | Preterm milk significant increase in IGF-1 0.231–0.419 µg/100 mL over DOL 3-28 [40] | Yes |
| IGF2 | Term: 1.2 µg/100 mL [41] Prem: 1.22 µg/100 mL [41] | No | Slight decline over first 3 months [41] | No |
| VEGF | Term: ~8 µg/100 mL Prem: ~3 µg/100 mL [42] Measured within first 7 DOL | Yes | Decline in Term levels from 8 to ~2.5 µg/100 mL. Not reported for premature infants. | Yes |
| EPO | Pre/Term: 11.7 mU/mL [43] Measured in first 4 months of life | No | Increases with duration of lactation, highest mean level 33.8 mU/mL measured over DOL 51–134 [43] | Yes |
| G-CSF | Term: 0.0156 µg/100 mL Prem: 0.0080 µg/100 mL [34] Measured on DOL 0–2 | Yes | Not reported | Not reported |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
York, D.J.; Smazal, A.L.; Robinson, D.T.; De Plaen, I.G. Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review. Nutrients 2021, 13, 3751. https://doi.org/10.3390/nu13113751
York DJ, Smazal AL, Robinson DT, De Plaen IG. Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review. Nutrients. 2021; 13(11):3751. https://doi.org/10.3390/nu13113751
Chicago/Turabian StyleYork, Daniel J., Anne L. Smazal, Daniel T. Robinson, and Isabelle G. De Plaen. 2021. "Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review" Nutrients 13, no. 11: 3751. https://doi.org/10.3390/nu13113751
APA StyleYork, D. J., Smazal, A. L., Robinson, D. T., & De Plaen, I. G. (2021). Human Milk Growth Factors and Their Role in NEC Prevention: A Narrative Review. Nutrients, 13(11), 3751. https://doi.org/10.3390/nu13113751