
The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis

 , , and
, , and
Abstract
1. Introduction
2. Postnatal Fatty Acid Status in Preterm Infants
3. Fatty Acids of Breastmilk
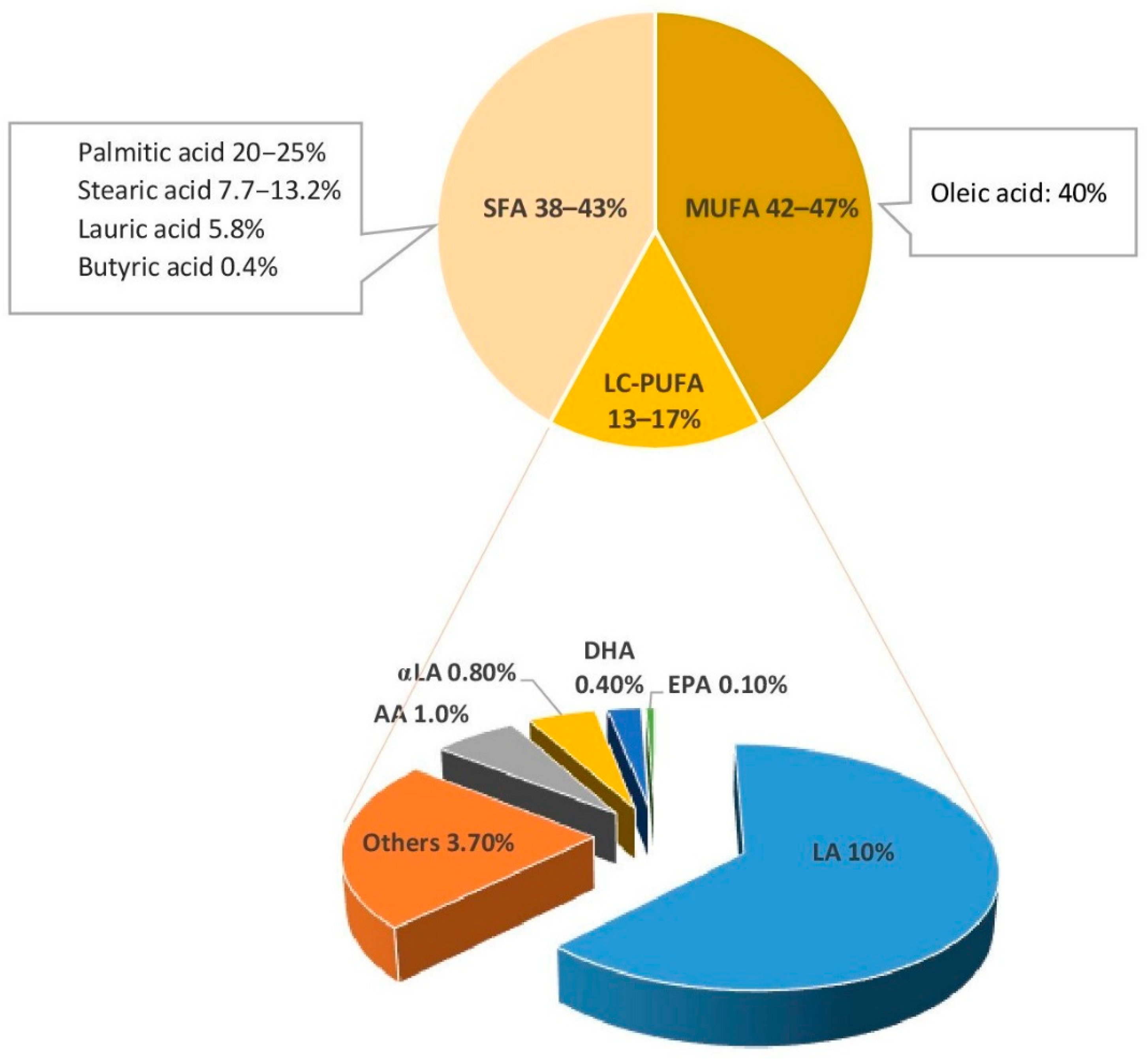
3.1. Fatty Acids Content and Structure
3.2. Role of Fatty Acid Balance in the Infant Diet
4. Fatty Acids in Infant Formula
4.1. Fatty Acids Content in Preterm Infant Formulas and Human Milk Fortifiers
4.2. Source of Fatty Acids in Infant Formula
4.3. n3 and n6 LC-PUFAs Balance in Infant Formula
5. Fatty Acid Digestion and Necrotizing Enterocolitis
5.1. Lipase Activity in Preterm Infants
5.2. Fat Malabsorption and Severity of Necrotizing Enterocolitis
5.3. Medium Chain Triglycerides
5.4. Stereospecific Positioning of Fatty Acids
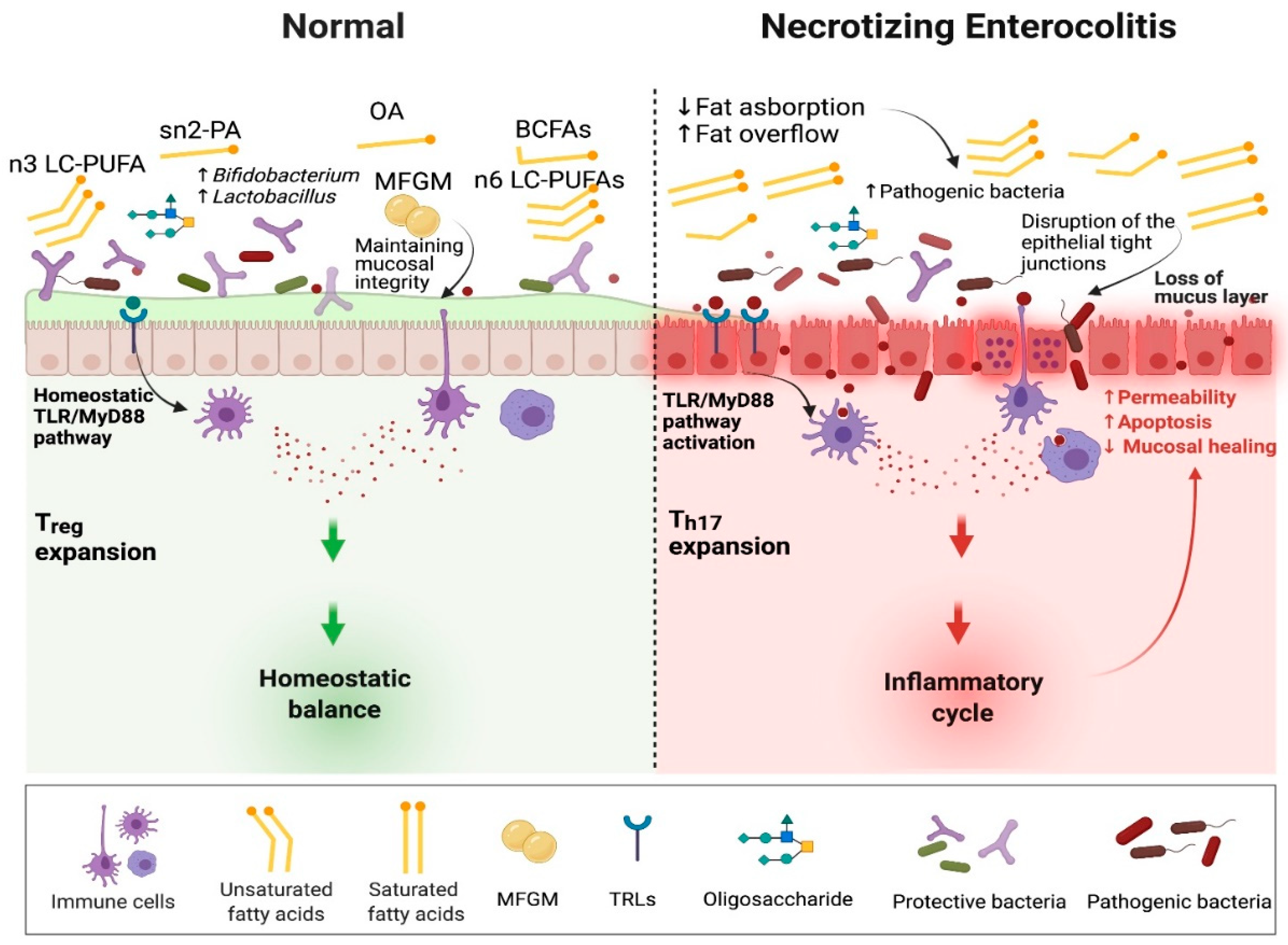
6. Fatty Acids and Pathogenesis of Necrotizing Enterocolitis
6.1. Direct Effect of Fatty Acids on NEC Pathway
6.1.1. TLR4
6.1.2. TLR2
6.1.3. Insulin Signaling Pathway
6.1.4. Platelet Activating Factor (PAF)
6.1.5. Permeability
6.1.6. T Lymphocytes
6.2. Indirect Effect of Fatty Acids on the Pathogenesis of Necrotizing Enterocolitis
6.2.1. Intestinal Microbiome
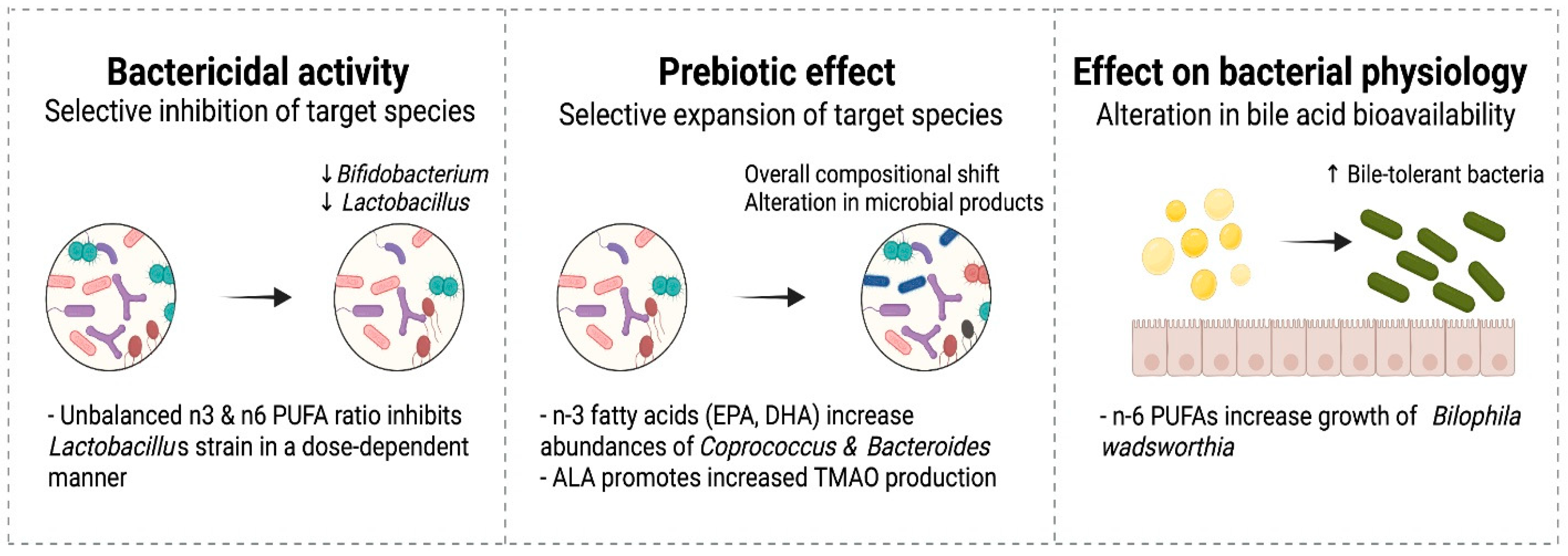
6.2.2. Antibacterial Effects of Fatty Acids and Interaction with Probiotics
6.2.3. Prebiotic Effects of Fatty Acids
6.2.4. Effects of Fatty Acids on Microbial Physiology
6.2.5. High Fat Diet and Intestinal Fat Overflow
6.2.6. Interactions with Other Nutrients
7. Supplementation of LC-PUFAs and Risk of Necrotizing Enterocolitis
7.1. Parenteral Lipids
7.2. Enteral Supplementation
8. Other Fat Supplementation
8.1. Branched-Chain Fatty Acids
8.2. Milk Fat Globule Membrane
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, I.H.; Hall, N.J. Contemporary Outcomes for Infants with Necrotizing Enterocolitis—A Systematic Review. J. Pediatr. 2020, 220, 86–92.e3. [Google Scholar] [CrossRef] [PubMed]
- Neu, J.; Walker, W.A. Medical Progress: Necrotizing Enterocolitis. N. Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef]
- Altobelli, E.; Angeletti, P.M.; Verrotti, A.; Petrocelli, R. The Impact of Human Milk on Necrotizing Enterocolitis: A Systematic Review and Meta-Analysis. Nutrients 2020, 12, 1322. [Google Scholar] [CrossRef]
- Sinclair, T.J.; Ye, C.; Chen, Y.; Zhang, D.; Li, T.; Ling, X.B.; Cohen, H.J.; Shaw, G.M.; Stevenson, D.K.; Chace, D.; et al. Progressive Metabolic Dysfunction and Nutritional Variability Precedes Necrotizing Enterocolitis. Nutrients 2020, 12, 1275. [Google Scholar] [CrossRef] [PubMed]
- Carr, L.E.; Virmani, M.D.; Rosa, F.; Munblit, D.; Matazel, K.S.; Elolimy, A.A.; Yeruva, L. Role of Human Milk Bioactives on Infants’ Gut and Immune Health. Front. Immunol. 2021, 12, 604080. [Google Scholar] [CrossRef] [PubMed]
- Gidrewicz, D.A.; Fenton, T.R. A systematic review and meta-analysis of the nutrient content of preterm and term breast milk. BMC Pediatr. 2014, 14, 216. [Google Scholar] [CrossRef]
- Fenton, T.R.; Kim, J.H. A systematic review and meta-analysis to revise the Fenton growth chart for preterm infants. BMC Pediatr. 2013, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Ling, P.-R.; Blackburn, G.L. Review of infant feeding: Key features of breast milk and infant formula. Nutrients 2016, 8, 279. [Google Scholar] [CrossRef]
- Delplanque, B.; Gibson, R.; Koletzko, B.; Lapillonne, A.; Strandvik, B. Lipid Quality in Infant Nutrition: Current Knowledge and Future Opportunities. J. Pediatr. Gastroenterol. Nutr. 2015, 61, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Himmelsbach, D.S.; Kays, S.E. ATR-Fourier Transform Mid-Infrared Spectroscopy for Determination of trans Fatty Acids in Ground Cereal Products without Oil Extraction. J. Agric. Food Chem. 2007, 55, 4327–4333. [Google Scholar] [CrossRef]
- Lapillonne, A.; Moltu, S.J. Long-Chain Polyunsaturated Fatty Acids and Clinical Outcomes of Preterm Infants. Ann. Nutr. Metab. 2016, 69, 35–44. [Google Scholar] [CrossRef]
- Miles, E.; Childs, C.; Calder, P. Long-Chain Polyunsaturated Fatty Acids (LCPUFAs) and the Developing Immune System: A Narrative Review. Nutrients 2021, 13, 247. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.T.; Carlson, S.E.; Murthy, K.; Frost, B.; Li, S.; Caplan, M. Docosahexaenoic and Arachidonic Acid Levels in Extremely Low Birth Weight Infants with Prolonged Exposure to Intravenous Lipids. J. Pediatr. 2013, 162, 56–61. [Google Scholar] [CrossRef]
- Robinson, D.T.; Caplan, M.; Carlson, S.E.; Yoder, R.; Murthy, K.; Frost, B. Early docosahexaenoic and arachidonic acid supplementation in extremely-low-birth-weight infants. Pediatr. Res. 2016, 80, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; DaSilva, D.A.; Cluette-Brown, J.E.; DiMonda, C.; Hamill, A.; Bhutta, A.Q.; Coronel, E.; Wilschanski, M.; Stephens, A.J.; Driscoll, D.F.; et al. Decreased Postnatal Docosahexaenoic and Arachidonic Acid Blood Levels in Premature Infants are Associated with Neonatal Morbidities. J. Pediatr. 2011, 159, 743–749.e2. [Google Scholar] [CrossRef]
- Frost, B.L.; Jilling, T.; Caplan, M.S. The Importance of Pro-Inflammatory Signaling in Neonatal Necrotizing Enterocolitis. Semin. Perinatol. 2008, 32, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Caplan, M.S.; Jilling, T. The role of polyunsaturated fatty acid supplementation in intestinal inflammation and neonatal necrotizing enterocolitis. Lipids 2001, 36, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Moravec, A.R.; Siv, A.W.; Hobby, C.R.; Lindsay, E.N.; Norbash, L.V.; Shults, D.J.; Symes, S.J.K.; Giles, D.K. Exogenous Polyunsaturated Fatty Acids Impact Membrane Remodeling and Affect Virulence Phenotypes among Pathogenic Vibrio Species. Appl. Environ. Microbiol. 2017, 83, e01415-17. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Thiel, I.; Abiodun, P.O. The fatty acid composition of human milk in Europe and Africa. J. Pediatr. 1992, 120, S62–S70. [Google Scholar] [CrossRef]
- Fidler, N.; Sauerwald, T.U.; Demmelmair, H.; Koletzko, B. Fat Content and Fatty Acid Composition of Fresh, Pasteurized, or Sterilized Human Milk. Adv. Exp. Med. Biol. 2001, 501, 485–495. [Google Scholar] [CrossRef]
- Koletzko, B.; Agostoni, C.; Bergmann, R.; Ritzenthaler, K.; Shamir, R. Physiological aspects of human milk lipids and implications for infant feeding: A workshop report. Acta Paediatr. 2011, 100, 1405–1415. [Google Scholar] [CrossRef]
- Innis, S.M. Dietary Triacylglycerol Structure and Its Role in Infant Nutrition. Adv. Nutr. 2011, 2, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B. Human Milk Lipids. Ann. Nutr. Metab. 2016, 69 (Suppl. 2), 28–40. [Google Scholar] [CrossRef]
- Bobiński, R.; Bobińska, J. Fatty acids of human milk—A review. Int. J. Vitam. Nutr. Res. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.X.; Berger, P.J.; Nold-Petry, C.A.; Nold, M.F. The immunological landscape in necrotising enterocolitis. Expert Rev. Mol. Med. 2016, 18, e12. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 Fatty Acids and Inflammatory Processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [PubMed]
- Drozdowski, L.A.; Clandinin, T.; Thomson, A.B.R. Ontogeny, growth and development of the small intestine: Understanding pediatric gastroenterology. World J. Gastroenterol. 2010, 16, 787–799. [Google Scholar] [PubMed]
- Innis, S.M.; Dai, C.; Wu, X.; Buchan, A.M.J.; Jacobson, K. Perinatal lipid nutrition alters early intestinal development and programs the response to experimental colitis in young adult rats. Am. J. Physiol. Liver Physiol. 2010, 299, G1376–G1385. [Google Scholar] [CrossRef] [PubMed]
- van Greevenbroek, M.M.; Voorhout, W.F.; Erkelens, D.W.; van Meer, G.; de Bruin, T.W. Palmitic acid and linoleic acid metabolism in Caco-2 cells: Different triglyceride synthesis and lipoprotein secretion. J. Lipid Res. 1995, 36, 13–24. [Google Scholar] [CrossRef]
- van Greevenbroek, M.M.; van Meer, G.; Erkelens, D.W.; de Bruin, T.W. Effects of saturated, mono-, and polyunsaturated fatty acids on the secretion of apo B containing lipoproteins by Caco-2 cells. Atherosclerosis 1996, 121, 139–150. [Google Scholar] [CrossRef]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Saturated fat, carbohydrate, and cardiovascular disease. Am. J. Clin. Nutr. 2010, 91, 502–509. [Google Scholar] [CrossRef]
- Gidding, S.S.; Lichtenstein, A.H.; Faith, M.S.; Karpyn, A.; Mennella, J.A.; Popkin, B.; Rowe, J.; van Horn, L.; Whitsel, L. Implementing American Heart Association pediatric and adult nutrition guidelines: A scientific statement from the American Heart Association Nutrition Committee of the Council on Nutrition, Physical Activity and Metabolism, Council on Cardiovascular Disease in the Young, Council on Arteriosclerosis, Thrombosis and Vascular Biology, Council on Cardiovascular Nursing, Council on Epidemiology and Prevention, and Council for High Blood Pressure Research. Circulation 2009, 119, 1161–1175. [Google Scholar]
- Sodhi, C.P.; Fulton, W.B.; Good, M.; Vurma, M.; Das, T.; Lai, C.-S.; Jia, H.; Yamaguchi, Y.; Lu, P.; Prindle, T.; et al. Fat composition in infant formula contributes to the severity of necrotising enterocolitis. Br. J. Nutr. 2018, 120, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Ng, D.H.C.; Klassen, J.R.L.; Embleton, N.D.; McGuire, W. Protein hydrolysate versus standard formula for preterm infants. Cochrane Database Syst. Rev. 2019. [Google Scholar] [CrossRef]
- Walsh, V.; Brown, J.V.E.; Askie, L.M.; Embleton, N.D.; McGuire, W. Nutrient-enriched formula versus standard formula for preterm infants. Cochrane Database Syst. Rev. 2019. [Google Scholar] [CrossRef]
- Qawasmi, A.; Landeros-Weisenberger, A.; Bloch, M.H. Meta-analysis of LCPUFA Supplementation of Infant Formula and Visual Acuity. Pediatrics 2012, 131, e262–e272. [Google Scholar] [CrossRef] [PubMed]
- Willatts, P.; Forsyth, S.; Agostoni, C.; Casaer, P.; Riva, E.; Boehm, G. Effects of long-chain PUFA supplementation in infant formula on cognitive function in later childhood. Am. J. Clin. Nutr. 2013, 98, 536S–542S. [Google Scholar] [CrossRef]
- Mendonça, M.A.; Araújo, W.M.C.; Borgo, L.A.; de Alencar, E.R. Lipid profile of different infant formulas for infants. PLoS ONE 2017, 12, e0177812. [Google Scholar] [CrossRef] [PubMed]
- Fusch, S.; Fusch, G.; Yousuf, E.I.; Rochow, M.; So, H.Y.; Fusch, C.; Rochow, N. Individualized Target Fortification of Breast Milk: Optimizing Macronutrient Content Using Different Fortifiers and Approaches. Front. Nutr. 2021, 8, 652641. [Google Scholar] [CrossRef]
- Robinson, D.T.; Martin, C.R. Fatty acid requirements for the preterm infant. Semin. Fetal Neonatal Med. 2017, 22, 8–14. [Google Scholar] [CrossRef]
- Lapillonne, A.; Jensen, C.L. Reevaluation of the DHA requirement for the premature infant. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Baker, S.; Cleghorn, G.; Neto, U.F.; Gopalan, S.; Hernell, O.; Hock, Q.S.; Jirapinyo, P.; Lonnerdal, B.; Pencharz, P.; et al. Global Standard for the Composition of Infant Formula: Recommendations of an ESPGHAN Coordinated International Expert Group. J. Pediatr. Gastroenterol. Nutr. 2005, 41, 584–599. [Google Scholar] [CrossRef] [PubMed]
- Mazzocchi, A.; D’Oria, V.; De Cosmi, V.; Bettocchi, S.; Milani, G.P.; Silano, M.; Agostoni, C. The Role of Lipids in Human Milk and Infant Formulae. Nutrients 2018, 10, 567. [Google Scholar] [CrossRef]
- López-López, A.; López-Sabater, M.C.; Campoy-Folgoso, C.; Rivero-Urgell, M.; Castellote-Bargalló, A.I. Fatty acid and sn-2 fatty acid composition in human milk from Granada (Spain) and in infant formulas. Eur. J. Clin. Nutr. 2002, 56, 1242–1254. [Google Scholar] [CrossRef]
- Koletzko, B.; Cheah, F.-C.; Domellof, M.; Poindexter, B.; Vain, N.; van Goudoever, J.P. Nutritional Care of Preterm Infants; Karger: Basel, Switzerland, 2021; Volume 122. [Google Scholar]
- Zou, L.; Pande, G.; Akoh, C.C. Infant Formula Fat Analogs and Human Milk Fat: New Focus on Infant Developmental Needs. Annu. Rev. Food Sci. Technol. 2016, 7, 139–165. [Google Scholar] [CrossRef]
- Li, A.; Ha, Y.; Wang, F.; Li, W.; Li, Q. Determination of Thermally Induced trans-Fatty Acids in Soybean Oil by Attenuated Total Reflectance Fourier Transform Infrared Spectroscopy and Gas Chromatography Analysis. J. Agric. Food Chem. 2012, 60, 10709–10713. [Google Scholar] [CrossRef] [PubMed]
- Caplan, M.S.; Russell, T.; Xiao, Y.; Amer, M.; Kaup, S.; Jilling, T. Effect of Polyunsaturated Fatty Acid (PUFA) Supplementation on Intestinal Inflammation and Necrotizing Enterocolitis (NEC) in a Neonatal Rat Model. Pediatr. Res. 2001, 49, 647–652. [Google Scholar] [CrossRef]
- Armand, M. Lipases and lipolysis in the human digestive tract: Where do we stand? Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.N.; Pollnitz, R.P.; Hamilton, J.R.; Chance, G.W. Impaired assimilation of nasojejunal feeds in healthy low-birth-weight newborn infants. J. Pediatr. 1977, 90, 431–434. [Google Scholar] [CrossRef]
- Peterson, J.A.; Hamosh, M.; Scallan, C.D.; Ceriani, R.L.; Henderson, T.R.; Mehta, N.R.; Armand, M.; Hamosh, P. Milk fat globule glycoproteins in human milk and in gastric aspirates of mother’s milk-fed preterm infants. Pediatric Res. 1998, 44, 499–506. [Google Scholar] [CrossRef]
- Hamosh, M.; Bitman, J.; Liao, T.H.; Mehta, N.R.; Buczek, R.J.; Wood, D.L.; Grylack, L.J.; Hamosh, P. Gastric Lipolysis and Fat Absorption in Preterm Infants: Effect of Medium-Chain Triglyceride or Long-Chain Triglyceride-Containing Formulas. Pediatrics 1989, 83, 86–92. [Google Scholar] [CrossRef]
- Liao, T.H.; Hamosh, P.; Hamosh, M. Fat Digestion by Lingual Lipase: Mechanism of Lipolysis in the Stomach and Upper Small Intestine. Pediatr. Res. 1984, 18, 402–409. [Google Scholar] [CrossRef]
- Fink, C.S.; Hamosh, P.; Hamosh, M. Fat Digestion in the Stomach: Stability of Lingual Lipase in the Gastric Environment. Pediatr. Res. 1984, 18, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Brady, M.S.; Rickard, K.A.; Fitzgerald, J.F.; Lemons, J.A. Specialized formulas and feedings for infants with malabsorption or formula intolerance. J. Am. Diet. Assoc. 1986, 86, 191–200. [Google Scholar] [CrossRef]
- Nevin-Folino, N.L.; Loughead, J.L.; Loughead, M.K. Enhanced-Calorie Formulas: Considerations and Options. Neonatal Netw. 2001, 20, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Casper, C.; Carnielli, V.P.; Hascoet, J.-M.; Lapillonne, A.; Maggio, L.; Timdahl, K.; Olsson, B.; Vågerö, M.; Hernell, O. rhBSSL Improves Growth and LCPUFA Absorption in Preterm Infants Fed Formula or Pasteurized Breast Milk. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 61–69. [Google Scholar] [CrossRef]
- Martin, C.R.; Cheesman, A.; Brown, J.; Makda, M.; Kutner, A.J.; DaSilva, D.; Zaman, M.; Freedman, S.D. Factors Determining Optimal Fatty Acid Absorption in Preterm Infants. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 130–136. [Google Scholar] [CrossRef]
- Stauble, C.G.; Helming, M.; Martyn, J.A.; Blobner, M.; Fink, H. Neuromuscular Recovery Is Prolonged After Immobilization or Superimposition of Inflammation With Immobilization Compared to Inflammation Alone: Data From a Preclinical Model. Crit. Care Med. 2016, 44, e1097–e1110. [Google Scholar] [CrossRef]
- Baumgart, S.; Chen, N.M.; Zhang, J.S.; Billadeau, D.D.; Gaisina, I.N.; Kozikowski, A.P.; Singh, S.K.; Fink, D.; Ströbel, P.; Klindt, C.; et al. GSK-3beta Governs Inflammation-Induced NFATc2 Signaling Hubs to Promote Pancreatic Cancer Progression. Mol. Cancer Ther. 2016, 15, 491–502. [Google Scholar] [CrossRef]
- Perretta, L.; Ouldibbat, L.; Hagadorn, J.I.; Brumberg, H.L. High versus low medium chain triglyceride content of formula for promoting short-term growth of preterm infants. Cochrane Database Syst. Rev. 2021, 2021, CD002777. [Google Scholar] [CrossRef]
- Sauerwald, U.C.; Fink, M.M.; Demmelmair, H.; Schoenaich, P.V.; Rauh-Pfeiffer, A.A.; Koletzko, B. Effect of Different Levels of Docosahexaenoic Acid Supply on Fatty Acid Status and Linoleic and α-Linolenic Acid Conversion in Preterm Infants. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 353–363. [Google Scholar] [CrossRef]
- Demmelmair, H.; Koletzko, B. Lipids in human milk. Best Pr. Res. Clin. Endocrinol. Metab. 2018, 32, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Bar-Yoseph, F.; Lifshitz, Y.; Cohen, T.; Malard, P.; Xu, C. SN2-Palmitate Reduces Fatty Acid Excretion in Chinese Formula-fed Infants. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 341–347. [Google Scholar] [CrossRef]
- Béghin, L.; Marchandise, X.; Lien, E.; Bricout, M.; Bernet, J.-P.; Lienhardt, J.-F.; Jeannerot, F.; Menet, V.; Requillart, J.-C.; Marx, J.; et al. Growth, stool consistency and bone mineral content in healthy term infants fed sn-2-palmitate-enriched starter infant formula: A randomized, double-blind, multicentre clinical trial. Clin. Nutr. 2019, 38, 1023–1030. [Google Scholar] [CrossRef] [PubMed]
- Breckenridge, W.C.; Marai, L.; Kuksis, A. Triglyceride structure of human milk fat. Can. J. Biochem. 1969, 47, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Filer, J.L.J.; Mattson, F.H.; Fomon, S.J. Triglyceride Configuration and Fat Absorption by the Human Infant. J. Nutr. 1969, 99, 293–298. [Google Scholar] [CrossRef]
- National High Blood Pressure Education Program Working Group on High Blood Pressure in Pregnancy. Report of the National High Blood Pressure Education Program Working Group on High Blood Pressure in Pregnancy. Am. J. Obstet. Gynecol. 2000, 183, S1–S22. [Google Scholar] [CrossRef]
- Lucas, A.; Quinlan, P.; Abrams, S.; Ryan, S.; Meah, S.; Lucas, P.J. Randomised controlled trial of a synthetic triglyceride milk formula for preterm infants. Arch. Dis. Child.-Fetal Neonatal Ed. 1997, 77, F178–F184. [Google Scholar] [CrossRef] [PubMed]
- Silber, G.H.; Hachey, D.L.; Schanler, R.J.; Garza, C. Manipulation of maternal diet to alter fatty acid composition of human milk intended for premature infants. Am. J. Clin. Nutr. 1988, 47, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, K.; Fewtrell, M.S.; Morley, R.; Abbott, R.; Quinlan, P.T.; Wells, J.C.; Bindels, J.G.; Lucas, A. Double-blind, randomized trial of a synthetic triacylglycerol in formula-fed term infants: Effects on stool biochemistry, stool characteristics, and bone mineralization. Am. J. Clin. Nutr. 1999, 70, 920–927. [Google Scholar] [CrossRef]
- Lu, P.; Bar-Yoseph, F.; Levi, L.; Lifshitz, Y.; Witte-Bouma, J.; De Bruijn, A.C.J.M.; Male, A.M.K.-V.; van Goudoever, J.; Renes, I.B. High Beta-Palmitate Fat Controls the Intestinal Inflammatory Response and Limits Intestinal Damage in Mucin Muc2 Deficient Mice. PLoS ONE 2013, 8, e65878. [Google Scholar] [CrossRef]
- Yaron, S.; Shachar, D.; Abramas, L.; Riskin, A.; Bader, D.; Litmanovitz, I.; Bar-Yoseph, F.; Cohen, T.; Levi, L.; Lifshitz, Y.; et al. Effect of high beta-palmitate content in infant formula on the intestinal microbiota of term infants. J. Pediatric Gastroenterol. Nutr. 2013, 56, 376–381. [Google Scholar] [CrossRef]
- Jiang, T.; Liu, B.; Li, J.; Dong, X.; Lin, M.; Zhang, M.; Zhao, J.; Dai, Y.; Chen, L. Association between sn-2 fatty acid profiles of breast milk and development of the infant intestinal microbiome. Food Funct. 2018, 9, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Tanner, S.M.; Berryhill, T.F.; Ellenburg, J.L.; Jilling, T.; Cleveland, D.S.; Lorenz, R.G.; Martin, C.A. Pathogenesis of necrotizing enterocolitis: Modeling the innate immune response. Am. J. Pathol. 2015, 185, 4–16. [Google Scholar] [CrossRef]
- Lu, P.; Sodhi, C.P.; Hackam, D.J. Toll-like receptor regulation of intestinal development and inflammation in the pathogenesis of necrotizing enterocolitis. Pathophysiology 2014, 21, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.H. Basic and Translational Understandings of Microbial Recognition by Toll-Like Receptors in the Intestine. J. Neurogastroenterol. Motil. 2011, 17, 28–34. [Google Scholar] [CrossRef]
- Neu, J.; Pammi, M. Pathogenesis of NEC: Impact of an altered intestinal microbiome. Semin. Perinatol. 2017, 41, 29–35. [Google Scholar] [CrossRef]
- Torrazza, R.M.; Neu, J. The Altered Gut Microbiome and Necrotizing Enterocolitis. Clin. Perinatol. 2013, 40, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.H.; Kim, J.A.; Lee, J.Y. Mechanisms for the activation of Toll-like receptor 2/4 by saturated fatty acids and inhibition by docosahexaenoic acid. Eur. J. Pharmacol. 2016, 785, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Plakidas, A.; Lee, W.H.; Heikkinen, A.; Chanmugam, P.; Bray, G.; Hwang, D.H. Differential modulation of Toll-like receptors by fatty acids: Preferential inhibition by n3 polyunsaturated fatty acids. J. Lipid Res. 2003, 44, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Sohn, K.H.; Rhee, S.H.; Hwang, D. Saturated fatty acids, but not unsaturated fatty acids, induce the expression of cyclooxygenase-2 mediated through Toll-like receptor 4. J. Biol. Chem. 2001, 276, 16683–16689. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, K.; Takeuchi, O.; Kawai, T.; Sanjo, H.; Ogawa, T.; Takeda, Y.; Takeda, K.; Akira, S. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: Evidence for TLR4 as the Lps gene product. J. Immunol. 1999, 162, 3749–3752. [Google Scholar] [PubMed]
- Lu, J.; Jilling, T.; Li, D.; Caplan, M.S. Polyunsaturated Fatty Acid Supplementation Alters Proinflammatory Gene Expression and Reduces the Incidence of Necrotizing Enterocolitis in a Neonatal Rat Model. Pediatr. Res. 2007, 61, 427–432. [Google Scholar] [CrossRef]
- Lee, J.Y.; Zhao, L.; Youn, H.S.; Weatherill, A.R.; Tapping, R.; Feng, L.; Lee, W.H.; Fitzgerald, K.; Hwang, D.H. Saturated Fatty Acid Activates but Polyunsaturated Fatty Acid Inhibits Toll-like Receptor 2 Dimerized with Toll-like Receptor 6 or 1. J. Biol. Chem. 2004, 279, 16971–16979. [Google Scholar] [CrossRef]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.; Ono-Moore, K.D.; Schneider, D.; Newman, J.; Adams, S.; Hwang, D.H. Saturated fatty acids activate TLR-mediated proinflammatory signaling pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef]
- Pal, D.; Dasgupta, S.; Kundu, R.; Maitra, S.; Das, G.; Mukhopadhyay, S.; Ray, S.; Majumdar, S.S.; Bhattacharya, S. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nat. Med. 2012, 18, 1279–1285. [Google Scholar] [CrossRef]
- Ghanim, H.; Mohanty, P.; Deopurkar, R.; Sia, C.L.; Korzeniewski, K.; Abuaysheh, S.; Chaudhuri, A.; Dandona, P. Acute Modulation of Toll-Like Receptors by Insulin. Diabetes Care 2008, 31, 1827–1831. [Google Scholar] [CrossRef] [PubMed]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vasc. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kim, F.; Pham, M.; Luttrell, I.; Bannerman, D.D.; Tupper, J.; Thaler, J.; Hawn, T.R.; Raines, E.W.; Schwartz, M.W. Toll-Like Receptor-4 Mediates Vascular Inflammation and Insulin Resistance in Diet-Induced Obesity. Circ. Res. 2007, 100, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid–induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef]
- Caplan, M.S.; Sun, X.-M.; Hsueh, W. Hypoxia, PAF, and necrotizing enterocolitis. Lipids 1991, 26, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Frost, B.L.; Caplan, M.S. Necrotizing enterocolitis: Pathophysiology, platelet-activating factor, and probiotics. Semin. Pediatr. Surg. 2013, 22, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Caplan, M.S.; Saraf, A.P.; Li, D.; Adler, L.; Liu, X.; Jilling, T. Platelet-activating factor-induced apoptosis is blocked by Bcl-2 in rat intestinal epithelial cells. Am. J. Physiol. Liver Physiol. 2004, 286, G340–G350. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lu, J.; Caplan, M.S.; Li, D.; Jilling, T. Polyunsaturated fatty acids block platelet-activating factor-induced phosphatidylinositol 3 kinase/Akt-mediated apoptosis in intestinal epithelial cells. Am. J. Physiol. Liver Physiol. 2008, 294, G1181–G1190. [Google Scholar] [CrossRef] [PubMed]
- Durkin, L.; Childs, C.; Calder, P. Omega-3 Polyunsaturated Fatty Acids and the Intestinal Epithelium—A Review. Foods 2021, 10, 199. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Liu, C.; Qin, Q.; Zhang, Y.; Wang, X.; Zhang, J.; Odle, J.; Lin, X.; Hu, C.A.; Liu, Y. EPA and DHA attenuate deoxynivalenol-induced intestinal porcine epithelial cell injury and protect barrier function integrity by inhibiting necroptosis signaling pathway. FASEB J. 2020, 34, 2483–2496. [Google Scholar] [CrossRef]
- Xiao, G.; Tang, L.; Yuan, F.; Zhu, W.; Zhang, S.; Liu, Z.; Geng, Y.; Qiu, X.; Zhang, Y.; Su, L. Eicosapentaenoic Acid Enhances Heat Stress-Impaired Intestinal Epithelial Barrier Function in Caco-2 Cells. PLoS ONE 2013, 8, e73571. [Google Scholar] [CrossRef]
- Beguin, P.; Errachid, A.; Larondelle, Y.; Schneider, Y.-J. Effect of polyunsaturated fatty acids on tight junctions in a model of the human intestinal epithelium under normal and inflammatory conditions. Food Funct. 2013, 4, 923–931. [Google Scholar] [CrossRef]
- Usami, M.; Muraki, K.; Iwamoto, M.; Ohata, A.; Matsushita, E.; Miki, A. Effect of eicosapentaenoic acid (EPA) on tight junction permeability in intestinal monolayer cells. Clin. Nutr. 2001, 20, 351–359. [Google Scholar] [CrossRef]
- Usami, M.; Komurasaki, T.; Hanada, A.; Kinoshita, K.; Ohata, A. Effect of gamma-linolenic acid or docosahexaenoic acid on tight junction permeability in intestinal monolayer cells and their mechanism by protein kinase C activation and/or eicosanoid formation. Nutrition 2003, 19, 150–156. [Google Scholar] [CrossRef]
- Ly, L.H.; Smith, R.; Switzer, K.C.; Chapkin, R.S.; McMurray, D.N. Dietary eicosapentaenoic acid modulates CTLA-4 expression in murine CD4+ T-cells. Prostaglandins Leukot. Essent. Fat. Acids 2006, 74, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Rodriguez-Palmero, M.; Demmelmair, H.; Fidler, N.; Jensen, R.; Sauerwald, T. Physiological aspects of human milk lipids. Early Hum. Dev. 2001, 65, S3–S18. [Google Scholar] [CrossRef]
- Del Prado, M.; Villalpando, S.; Elizondo, A.; Rodriguez, M.; Demmelmair, H.; Koletzko, B. Contribution of dietary and newly formed arachidonic acid to human milk lipids in women eating a low-fat diet. Am. J. Clin. Nutr. 2001, 74, 242–247. [Google Scholar] [CrossRef]
- Rodriguez-Palmero, M.; Koletzko, B.; Kunz, C.; Jensen, R. Nutritional and biochemical properties of human milk: II. Lipids, micronutrients, and bioactive factors. Clin. Perinatol. 1999, 26, 335–359. [Google Scholar] [CrossRef]
- Hong, W.; Xu, D.; Song, X.; Niu, B.; Zhuang, Z.; Lu, Y.; Lei, X.; Ma, R.; Lu, C.; Sun, N.; et al. Vitamin A and retinoic acid accelerate the attenuation of intestinal adaptability upon feeding induced by high-fat diet in mice. J. Nutr. Biochem. 2021, 97, 108803. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Thiel, I.; Springer, S. Lipids in human milk: A model for infant formulae? Eur. J. Clin. Nutr. 1992, 46 (Suppl. 4), S45–S55. [Google Scholar]
- Rakshasbhuvankar, A.A.; Pillow, J.J.; Simmer, K.N.; Patole, S.K. Vitamin A supplementation in very-preterm or very-low-birth-weight infants to prevent morbidity and mortality: A systematic review and meta-analysis of randomized trials. Am. J. Clin. Nutr. 2021, 114, 2084–2096. [Google Scholar] [CrossRef] [PubMed]
- Fundora, J.B.; Guha, P.; Shores, D.R.; Pammi, M.; Maheshwari, A. Intestinal dysbiosis and necrotizing enterocolitis: Assessment for causality using Bradford Hill criteria. Pediatr. Res. 2020, 87, 235–248. [Google Scholar] [CrossRef]
- Mai, V.; Young, C.M.; Ukhanova, M.; Wang, X.; Sun, Y.; Casella, G.; Theriaque, D.; Li, N.; Sharma, R.; Hudak, M.; et al. Fecal Microbiota in Premature Infants Prior to Necrotizing Enterocolitis. PLoS ONE 2011, 6, e20647. [Google Scholar] [CrossRef]
- Morrow, A.L.; Lagomarcino, A.J.; Schibler, K.R.; Taft, D.; Yu, Z.; Wang, B.; Altaye, M.; Wagner, M.; Gevers, D.; Ward, D.V.; et al. Early microbial and metabolomic signatures predict later onset of necrotizing enterocolitis in preterm infants. Microbiome 2013, 1, 13. [Google Scholar] [CrossRef]
- Wang, Y.; Hoenig, J.D.; Malin, K.J.; Qamar, S.; Petrof, E.O.; Sun, J.; Antonopoulos, D.A.; Chang, E.B.; Claud, E.C. 16S rRNA gene-based analysis of fecal microbiota from preterm infants with and without necrotizing enterocolitis. ISME J. 2009, 3, 944–954. [Google Scholar] [CrossRef]
- Moossavi, S.; Atakora, F.; Miliku, K.; Sepehri, S.; Robertson, B.; Duan, Q.L.; Becker, A.B.; Mandhane, P.J.; Turvey, S.; Moraes, T.J.; et al. Integrated Analysis of Human Milk Microbiota With Oligosaccharides and Fatty Acids in the CHILD Cohort. Front. Nutr. 2019, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Fehr, K.; Moossavi, S.; Sbihi, H.; Boutin, R.C.; Bode, L.; Robertson, B.; Yonemitsu, C.; Field, C.J.; Becker, A.B.; Mandhane, P.J.; et al. Breastmilk Feeding Practices Are Associated with the Co-Occurrence of Bacteria in Mothers’ Milk and the Infant Gut: The CHILD Cohort Study. Cell Host Microbe 2020, 28, 285–297.e4. [Google Scholar] [CrossRef] [PubMed]
- Younge, N.; Yang, Q.; Seed, P.C. Enteral High Fat-Polyunsaturated Fatty Acid Blend Alters the Pathogen Composition of the Intestinal Microbiome in Premature Infants with an Enterostomy. J. Pediatr. 2017, 181, 93–101.e6. [Google Scholar] [CrossRef]
- Vijay, A.; Astbury, S.; Le Roy, C.; Spector, T.D.; Valdes, A.M. The prebiotic effects of omega-3 fatty acid supplementation: A six-week randomised intervention trial. Gut Microbes 2021, 13, 1–11. [Google Scholar] [CrossRef]
- Collins, C.T.; Gibson, R.; Anderson, P.; McPhee, A.J.; Sullivan, T.; Gould, J.F.; Ryan, P.; Doyle, L.; Davis, P.G.; McMichael, J.E.; et al. Neurodevelopmental outcomes at 7 years’ corrected age in preterm infants who were fed high-dose docosahexaenoic acid to term equivalent: A follow-up of a randomised controlled trial. BMJ Open 2015, 5, e007314. [Google Scholar] [CrossRef] [PubMed]
- Fava, F.; Gitau, R.; Griffin, B.A.; Gibson, G.R.; Tuohy, K.; Lovegrove, J.A. The type and quantity of dietary fat and carbohydrate alter faecal microbiome and short-chain fatty acid excretion in a metabolic syndrome ‘at-risk’ population. Int. J. Obes. 2012, 37, 216–223. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, F.; Yuan, J.; Li, J.; Jiang, D.; Zhang, J.; Li, H.; Wang, R.; Tang, J.; Huang, T.; et al. Effects of dietary fat on gut microbiota and faecal metabolites, and their relationship with cardiometabolic risk factors: A 6-month randomised controlled-feeding trial. Gut 2019, 68, 1417–1429. [Google Scholar] [CrossRef]
- Chi, C.; Buys, N.; Li, C.; Sun, J.; Yin, C. Effects of prebiotics on sepsis, necrotizing enterocolitis, mortality, feeding intolerance, time to full enteral feeding, length of hospital stay, and stool frequency in preterm infants: A meta-analysis. Eur. J. Clin. Nutr. 2019, 73, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Andersen, A.; Mølbak, L.; Thymann, T.; Michaelsen, K.F.; Lauritzen, L. Dietary long-chain n3 PUFA, gut microbiota and fat mass in early postnatal piglet development—exploring a potential interplay. Prostaglandins Leukot. Essent. Fat. Acids 2011, 85, 345–351. [Google Scholar] [CrossRef]
- Liu, T.; Hougen, H.; Vollmer, A.C.; Hiebert, S.M. Gut bacteria profiles of Mus musculus at the phylum and family levels are influenced by saturation of dietary fatty acids. Anaerobe 2012, 18, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; DeCoffe, D.; Brown, K.; Rajendiran, E.; Estaki, M.; Dai, C.; Yip, A.; Gibson, D.L. Fish Oil Attenuates Omega-6 Polyunsaturated Fatty Acid-Induced Dysbiosis and Infectious Colitis but Impairs LPS Dephosphorylation Activity Causing Sepsis. PLoS ONE 2013, 8, e55468. [Google Scholar] [CrossRef]
- de Wit, N.; Derrien, M.; Bosch-Vermeulen, H.; Oosterink, E.; Keshtkar, S.; Duval, C.; den Bosch, J.; Kleerebezem, M.; Müller, M.; van der Meer, R. Saturated fat stimulates obesity and hepatic steatosis and affects gut microbiota composition by an enhanced overflow of dietary fat to the distal intestine. American journal of physiology. Gastrointest. Liver Physiol. 2012, 303, G589–G599. [Google Scholar] [CrossRef]
- Saravi, S.S.; Bonetti, N.; Camici, G.; Luscher, T.; Beer, J. Distinct dietary alpha-linolenic acid-dependent shifts in the fecal microbiome composition suppresses aging-associated inflammatory responses and thrombus formation. Eur. Heart J. 2020, 41, 3773. [Google Scholar] [CrossRef]
- Tanaka, K.; Hosozawa, M.; Kudo, N.; Yoshikawa, N.; Hisata, K.; Shoji, H.; Shinohara, K.; Shimizu, T. The pilot study: Sphingomyelin-fortified milk has a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev. 2013, 35, 45–52. [Google Scholar] [CrossRef]
- Ghezzal, S.; Postal, B.G.; Quevrain, E.; Brot, L.; Seksik, P.; Leturque, A.; Thenet, S.; Carrière, V. Palmitic acid damages gut epithelium integrity and initiates inflammatory cytokine production. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2020, 1865, 158530. [Google Scholar] [CrossRef]
- Ms, E.Y.H.; Leone, V.A.; Devkota, S.; Wang, Y.; Brady, M.J.; Chang, E.B. Composition of Dietary Fat Source Shapes Gut Microbiota Architecture and Alters Host Inflammatory Mediators in Mouse Adipose Tissue. J. Parenter. Enter. Nutr. 2013, 37, 746–754. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty Acids and Derivatives as Antimicrobial Agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef]
- Kodicek, E.; Worden, A.N. The effect of unsaturated fatty acids on Lactobacillus helveticus and other Gram-positive micro-organisms. Biochem. J. 1945, 39, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Beghetti, I.; Panizza, D.; Lenzi, J.; Gori, D.; Martini, S.; Corvaglia, L.; Aceti, A. Probiotics for Preventing Necrotizing Enterocolitis in Preterm Infants: A Network Meta-Analysis. Nutrients 2021, 13, 192. [Google Scholar] [CrossRef] [PubMed]
- Kankaanpãã, P.E.; Salminen, S.J.; Isolauri, E.; Lee, Y.K. The influence of polyunsaturated fatty acids on probiotic growth and adhesion. FEMS Microbiol. Lett. 2001, 194, 149–153. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.-L.; Rainteau, D.; Bridonneau, C.; Da Costa, G.; Van Hylckama Vlieg, J.E.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 2802. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef]
- Hulzebos, C.V.; Van Zoonen, A.G.J.F.; Hulscher, J.B.F.; Schat, T.E.; Kooi, E.M.W.; Koehorst, M.; Boverhof, R.; Krabbe, P.; Groen, A.K.; Verkade, H.J. Fecal Bile Salts and the Development of Necrotizing Enterocolitis in Preterm Infants. PLoS ONE 2017, 12, e0168633. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Hernell, O. Lipid digestion and absorption in early life: An update. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Hay, W.W.; Thureen, P. Protein for Preterm Infants: How Much is Needed? How Much is Enough? How Much is Too Much? Pediatr. Neonatol. 2010, 51, 198–207. [Google Scholar] [CrossRef]
- da Silva, R.P.; Kelly, K.B.; Lewis, E.D.; Leonard, K.-A.; Goruk, S.; Curtis, J.M.; Vine, D.F.; Proctor, S.D.; Field, C.J.; Jacobs, R.L. Choline deficiency impairs intestinal lipid metabolism in the lactating rat. J. Nutr. Biochem. 2015, 26, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut Flora Metabolism of Phosphatidylcholine Promotes Cardiovascular Disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Seldin, M.M.; Meng, Y.; Qi, H.; Zhu, W.; Wang, Z.; Hazen, S.L.; Lusis, A.J.; Shih, D.M. Trimethylamine N-Oxide Promotes Vascular Inflammation Through Signaling of Mitogen-Activated Protein Kinase and Nuclear Factor-kappaB. J. Am. Heart Assoc. 2016, 5, 1077–1083. [Google Scholar] [CrossRef]
- Valentine, C.J.; Morrow, G.; Fernandez, S.; Gulati, P.; Bartholomew, D.; Long, D.; Welty, S.E.; Morrow, A.L.; Rogers, L.K. Docosahexaenoic Acid and Amino Acid Contents in Pasteurized Donor Milk are Low for Preterm Infants. J. Pediatr. 2010, 157, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Rochow, N.; Fusch, G.; Ali, A.; Bhatia, A.; So, H.Y.; Iskander, R.; Chessell, L.; el Helou, S.; Fusch, C. Individualized target fortification of breast milk with protein, carbohydrates, and fat for preterm infants: A double-blind randomized controlled trial. Clin. Nutr. 2021, 40, 54–63. [Google Scholar] [CrossRef]
- Yap, P.S.X.; Chong, C.W.; Ahmad Kamar, A.; Yap, I.K.S.; Choo, Y.M.; Lai, N.M.; Teh, C.S.J. Neonatal intensive care unit (NICU) exposures exert a sustained influence on the progression of gut microbiota and metabolome in the first year of life. Sci. Rep. 2021, 11, 1353. [Google Scholar] [CrossRef] [PubMed]
- Grimm, H.; Tibell, A.; Norrlind, B.; Blecher, C.; Wilker, S.; Schwemmle, K. Immunoregulation by Parenteral Lipids: Impact of the n3 to n6 Fatty Acid Ratio. J. Parenter. Enter. Nutr. 1994, 18, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Yakah, W.; Singh, P.; Brown, J.; Stoll, B.; Burrin, D.; Premkumar, M.H.; Otu, H.H.; Gu, X.; Dillon, S.T.; Libermann, T.A.; et al. Parenteral lipid emulsions induce unique ileal fatty acid and metabolomic profiles but do not increase the risk of necrotizing enterocolitis in preterm pigs. Am. J. Physiol. Liver Physiol. 2021, 320, G227–G239. [Google Scholar] [CrossRef] [PubMed]
- Bjornvad, C.; Thymann, T.; Deutz, N.; Burrin, D.; Jensen, S.K.; Jensen, B.B.; Mølbak, L.; Boye, M.; Larsson, L.-I.; Schmidt, M.; et al. Enteral feeding induces diet-dependent mucosal dysfunction, bacterial proliferation, and necrotizing enterocolitis in preterm pigs on parenteral nutrition. Am. J. Physiol. Liver Physiol. 2008, 295, G1092–G1103. [Google Scholar] [CrossRef]
- Vegge, A.; Thymann, T.; Lauritzen, L.; Bering, S.B.; Wiinberg, B.; Sangild, P.T. Parenteral lipids and partial enteral nutrition affect hepatic lipid composition but have limited short term effects on formula-induced necrotizing enterocolitis in preterm piglets. Clin. Nutr. 2015, 34, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, V.; Malviya, M.N.; Soll, R. Lipid emulsions for parenterally fed preterm infants. Cochrane Database Syst. Rev. 2018, 4, CD013163. [Google Scholar] [CrossRef]
- Vayalthrikkovil, S.; Rabi, Y.; Amin, H.; Spence, J.-M.; Robertson, H.L.; Lodha, A.; Bashir, R.A. Parenteral Fish-Oil Lipid Emulsions in the Prevention of Severe Retinopathy of Prematurity: A Systematic Review and Meta-Analysis. Am. J. Perinatol. 2016, 34, 705–715. [Google Scholar] [CrossRef]
- Koletzko, B.; Boey, C.C.; Campoy, C.; Carlson, S.E.; Chang, N.; Guillermo-Tuazon, M.A.; Joshi, S.; Prell, C.; Quak, S.H.; Sjarif, D.R.; et al. Current Information and Asian Perspectives on Long-Chain Polyunsaturated Fatty Acids in Pregnancy, Lactation, and Infancy: Systematic Review and Practice Recommendations from an Early Nutrition Academy Workshop. Ann. Nutr. Metab. 2014, 65, 49–80. [Google Scholar] [CrossRef] [PubMed]
- FAO Food and Nutrition. Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation; FAO Food and Nutrition: Geneva, Switzerland, 2008. [Google Scholar]
- Brenna, J.T.; Varamini, B.; Jensen, R.G.; Diersen-Schade, D.A.; Boettcher, J.A.; Arterburn, L.M. Docosahexaenoic and arachidonic acid concentrations in human breast milk worldwide. Am. J. Clin. Nutr. 2007, 85, 1457–1464. [Google Scholar] [CrossRef]
- Baack, M.L.; Norris, A.W.; Yao, J.; Colaizy, T. Long-chain polyunsaturated fatty acid levels in US donor human milk: Meeting the needs of premature infants? J. Perinatol. 2012, 32, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.H.; Harris, W.S. Should there be a target level of docosahexaenoic acid in breast milk? Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 92–96. [Google Scholar] [CrossRef]
- Valentine, C.J.; Morrow, G.; Pennell, M.; Morrow, A.L.; Hodge, A.; Haban-Bartz, A.; Collins, K.; Rogers, L.K. Randomized controlled trial of docosahexaenoic acid supplementation in midwestern U.S. human milk donors. Breastfeed. Med. Off. J. Acad. Breastfeed. Med. 2013, 8, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Berseth, C.; Harris, C.; Wampler, J.; Hoffman, D.; Diersen-Schade, D. Liquid human milk fortifier significantly improves docosahexaenoic and arachidonic acid status in preterm infants. Prostaglandins Leukot. Essent. Fat. Acids 2014, 91, 97–103. [Google Scholar] [CrossRef]
- Bernabe-García, M.; Calder, P.; Villegas-Silva, R.; Rodríguez-Cruz, M.; Chávez-Sánchez, L.; Cruz-Reynoso, L.; Mateos-Sánchez, L.; Lara-Flores, G.; Aguilera-Joaquín, A.; Sánchez-García, L. Efficacy of Docosahexaenoic Acid for the Prevention of Necrotizing Enterocolitis in Preterm Infants: A Randomized Clinical Trial. Nutrients 2021, 13, 648. [Google Scholar] [CrossRef] [PubMed]
- Marc, I.; Piedboeuf, B.; Lacaze-Masmonteil, T.; Fraser, W.; Mâsse, B.; Mohamed, I.; Qureshi, M.; Afifi, J.; Lemyre, B.; Caouette, G.; et al. Effect of Maternal Docosahexaenoic Acid Supplementation on Bronchopulmonary Dysplasia-Free Survival in Breastfed Preterm Infants A Randomized Clinical Trial. Jama-J. Am. Med. Assoc. 2020, 324, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.T.; Makrides, M.; McPhee, A.J.; Sullivan, T.; Davis, P.G.; Thio, M.; Simmer, K.; Rajadurai, V.S.; Travadi, J.; Berry, M.J.; et al. Docosahexaenoic Acid and Bronchopulmonary Dysplasia in Preterm Infants. N. Engl. J. Med. 2017, 376, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- Baack, M.L.; Puumala, S.E.; Messier, S.E.; Pritchett, D.K.; Harris, W.S. Daily Enteral DHA Supplementation Alleviates Deficiency in Premature Infants. Lipids 2016, 51, 423–433. [Google Scholar] [CrossRef]
- Makrides, M.; Gibson, R.A.; McPhee, A.J.; Collins, C.T.; Davis, P.G.; Doyle, L.W.; Simmer, K.; Colditz, P.B.; Morris, S.; Smithers, L.G.; et al. Neurodevelopmental Outcomes of Preterm Infants Fed High-Dose Docosahexaenoic Acid: A Randomized Controlled Trial. Obstet. Gynecol. Surv. 2009, 64, 297–298. [Google Scholar] [CrossRef]
- Henriksen, C.; Haugholt, K.; Lindgren, M.; Aurvåg, A.K.; Rønnestad, A.; Grønn, M.; Solberg, R.; Moen, A.; Nakstad, B.; Berge, R.K.; et al. Improved Cognitive Development Among Preterm Infants Attributable to Early Supplementation of Human Milk With Docosahexaenoic Acid and Arachidonic Acid. Pediatrics 2008, 121, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Groh-Wargo, S.; Jacobs, J.; Auestad, N.; O’Connor, D.L.; Moore, J.J.; Lerner, E. Body Composition in Preterm Infants Who Are Fed Long-Chain Polyunsaturated Fatty Acids: A Prospective, Randomized, Controlled Trial. Pediatr. Res. 2005, 57, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Clandinin, M.T.; Van Aerde, J.E.; Merkel, K.L.; Harris, C.L.; Springer, M.A.; Hansen, J.W.; Diersen-Schade, D.A. Growth and development of preterm infants fed infant formulas containing docosahexaenoic acid and arachidonic acid. J. Pediatr. 2005, 146, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Fewtrell, M.S.; Abbott, R.; Kennedy, K.; Singhal, A.; Morley, R.; Caine, E.; Jamieson, C.; Cockburn, F.; Lucas, A. Randomized, double-blind trial of long-chain polyunsaturated fatty acid supplementation with fish oil and borage oil in preterm infants. J. Pediatr. 2004, 144, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M.; Adamkin, D.H.; Hall, R.T.; Kalhan, S.; Lair, C.; Lim, M.; Stevens, D.C.; Twist, P.F.; Diersen-Schade, D.A.; Harris, C.L.; et al. Docosahexaenoic acid and arachidonic acid enhance growth with no adverse effects in preterm infants fed formula. J. Pediatr. 2002, 140, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Fewtrell, M.S.; Morley, R.; Abbott, R.A.; Singhal, A.; Isaacs, E.B.; Stephenson, T.; MacFadyen, U.; Lucas, A. Double-blind, randomized trial of long-chain polyunsaturated fatty acid supplementation in formula fed to preterm infants. Pediatrics 2002, 110, 73–82. [Google Scholar] [CrossRef]
- O’Connor, D.L.; Hall, R.; Adamkin, D.; Auestad, N.; Castillo, M.; Connor, W.E.; Connor, S.L.; Fitzgerald, K.; Groh-Wargo, S.; Hartmann, E.E.; et al. Growth and Development in Preterm Infants Fed Long-Chain Polyunsaturated Fatty Acids: A Prospective, Randomized Controlled Trial. Pediatrics 2001, 108, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.E.; Montalto, M.B.; Ponder, D.L.; Werkman, S.H.; Korones, S.B. Lower Incidence of Necrotizing Enterocolitis in Infants Fed a Preterm Formula with Egg Phospholipids. Pediatr. Res. 1998, 44, 491–498. [Google Scholar] [CrossRef]
- Carlson, S.E.; Werkman, S.H. A randomized trial of visual attention of preterm infants fed docosahexaenoic acid until two months. Lipids 1996, 31, 85–90. [Google Scholar] [CrossRef]
- Ran-Ressler, R.R.; Devapatla, S.; Lawrence, P.; Brenna, J.T. Branched Chain Fatty Acids Are Constituents of the Normal Healthy Newborn Gastrointestinal Tract. Pediatr. Res. 2008, 64, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, N.; Ray, T. Skin Lipids. 3. Fatty Chains in Skin Lipids. The Use of Vernix Caseosa to Differentiate between Endogenous and Exogenous Components in Human Skin Surface Lipid. J. Am. Oil Chem. Soc. 1965, 42, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Dingess, K.A.; Valentine, C.J.; Ollberding, N.J.; Davidson, B.S.; Woo, J.; Summer, S.; Peng, Y.M.; Guerrero, M.L.; Ruiz-Palacios, G.M.; Ran-Ressler, R.R.; et al. Branched-chain fatty acid composition of human milk and the impact of maternal diet: The Global Exploration of Human Milk (GEHM) Study. Am. J. Clin. Nutr. 2017, 105, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Ran-Ressler, R.R.; Khailova, L.; Arganbright, K.M.; Adkins-Rieck, C.K.; Jouni, Z.E.; Koren, O.; Ley, R.; Brenna, J.T.; Dvorak, B. Branched Chain Fatty Acids Reduce the Incidence of Necrotizing Enterocolitis and Alter Gastrointestinal Microbial Ecology in a Neonatal Rat Model. PLoS ONE 2011, 6, e29032. [Google Scholar] [CrossRef]
- Dewettinck, K.; Rombaut, R.; Thienpont, N.; Le, T.T.; Messens, K.; Van Camp, J. Nutritional and technological aspects of milk fat globule membrane material. Int. Dairy J. 2008, 18, 436–457. [Google Scholar] [CrossRef]
- Snow, D.R.; Ward, R.E.; Olsen, A.; Jimenez-Flores, R.; Hintze, K.J. Membrane-rich milk fat diet provides protection against gastrointestinal leakiness in mice treated with lipopolysaccharide. J. Dairy Sci. 2011, 94, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Bhinder, G.; Allaire, J.M.; Garcia, C.; Lau, J.T.; Chan, J.M.; Ryz, N.R.; Bosman, E.S.; Graef, F.A.; Crowley, S.M.; Celiberto, L.S.; et al. Milk Fat Globule Membrane Supplementation in Formula Modulates the Neonatal Gut Microbiome and Normalizes Intestinal Development. Sci. Rep. 2017, 7, 45274. [Google Scholar] [CrossRef]
- Aziz, M.M.; Ishihara, S.; Mishima, Y.; Oshima, N.; Moriyama, I.; Yuki, T.; Kadowaki, Y.; Rumi, M.A.K.; Amano, Y.; Kinoshita, Y. MFG-E8 Attenuates Intestinal Inflammation in Murine Experimental Colitis by Modulating Osteopontin-Dependent alpha(v)beta(3) Integrin Signaling. J. Immunol. 2009, 182, 7222–7232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wen, J.; Zhou, J.; Cai, W.; Qian, L. Milk Fat Globule Membrane Ameliorates Necrotizing Enterocolitis in Neonatal Rats and Suppresses Lipopolysaccharide-Induced Inflammatory Response in IEC-6 Enterocytes. J. Parenter. Enter. Nutr. 2019, 43, 863–873. [Google Scholar] [CrossRef]
- Zhang, Y.; Brenner, M.; Yang, W.-L.; Wang, P. Recombinant human MFG-E8 ameliorates colon damage in DSS- and TNBS-induced colitis in mice. Lab. Investig. 2015, 95, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Yuasa, H.; Sekiya, M.; Ozeki, S.; Watanabe, J. Evaluation of Milk Fat-Globule Membrane (MFGM) Emulsion for Oral Administration: Absorption of α-Linolenic Acid in Rats and the Effect of Emulsion Droplet Size. Biol. Pharm. Bull. 1994, 17, 756–758. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yu, Z.; Li, Y.; Niu, Y.; Tang, Q.; Wu, J. Milk Fat Globule Membrane Enhances Colonic-Mucus-Barrier Function in a Rat Model of Short-Bowel Syndrome. JPEN. J. Parenter. Enter. Nutr. 2021, 45, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, Y.; Li, Z.; Christensen, B.; Heckmann, A.B.; Stenlund, H.; Lönnerdal, B.; Hernell, O. Feeding Infants Formula With Probiotics or Milk Fat Globule Membrane: A Double-Blind, Randomized Controlled Trial. Front. Pediatr. 2019, 7, 347. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | Model | Fat Intake and Type | Microbiome Changes | Metabolomics Changes |
|---|---|---|---|---|
| Human studies—Infants | ||||
| Younge et al. [115] 2017 | Randomized, controlled trial in preterm infants with enterostomy due NEC or SIP | Fish oil or safflower oil compared to standard of care
| ↓ Proteobacteria ↓ Enterobacteriaceae ↑ Actinobacteria ↑ Enterococcus | - |
| Human studies—Adults | ||||
| Vijay et al. [116] 2021 | 6-week randomized dietary intervention | n3 fatty acid supplementation
| ↑ Coprococcus spp. ↑ Bacteroides spp. ↓ Collinsella spp. ↓ Ruminococcus | ↑ Butyrate, iso-butyrate, isovalerate ↑ Total plasma n3 fatty acids |
| Watson et al. [3] 2018 | 8-week randomized, open-label, cross-over trial with 12-week washout | n3 PUFA supplements 2000 mg EPA and 2000 mg DHA per day in two formulations
| ↑ Bifidobacterium ↑ Roseburia ↑ Lactobacillus ↓ Faecalibacterium | - |
| Kjolbaek et al. [117] 2020 | Cross-over design with two diet periods of 4 weeks with 4-week washout period | Arabinoxylan oligosaccharides and PUFA cross-over Daily PUFA intake of approximately 10% increasing the intake of PUFA including n3 fatty acids and lowering SFA intake. fish oil capsules containing 3.6 g/d n3 PUFA (DHA and EPA) | No change | - |
| Fava et al. [118] 2013 | Randomized trial in adult volunteers with at least two features of metabolic syndrome | Participants followed a 4-week reference diet and then were randomly assigned to the intervention arms for 24 weeks. Reference diet: high SFA diet/high glycemic index Intervention arm:
| ↓ Total bacteria in high MUFA groups ↓ Total bacteria in high SFA compared to baseline ↑ Faecalibacterium prausnitzii in high SFA compared to baseline | ↑ Acetate, propionate, and n-butyrate in high SFA compared to baseline |
| Wan et al. [119] 2019 | 6-month randomized controlled-feeding Trial in healthy young adults with normal BMI | The three isocaloric diets:
| Low-fat diet: ↑ Shannon diversity ↑ Blautia ↑ Faecalibacterium Moderate-fat diet: ↑ Bacteroidetes High-fat diet: ↓ Firmicutes ↑ Bacteroidetes ↓ Faecalibacterium ↑ Bacteroides | - |
| Pig | ||||
| Che et al. [120] 2019 | Piglets with intrauterine growth retardation | Diet contained either flaxseed oil (enriched in n3 PUFAs) compared to soy oil (high in n6) | ↑ Actinobacteria ↑ Melainabacteria ↑ Bifidobacterium ↑ Blautia ↓ Spirochaetes | ↓ Diarrhea ↑ villus height ↑ Ileal Claudin-1 and ZO-1 ↓ Ileal MyD88, NF-κB, TNF-α and IL-10 |
| Anderson et al. [121] 2011 | Piglet | Piglets were grouped into these treatments:
| Fish oil diet: ↑ Proteobacteria ↑ Actinobacteria Sunflower oil diet: ↑ Bacteroides spp. | - |
| Mouse | ||||
| Liu et al. [122] 2012 | Adult mice | Regular rodent chow for 14 days, and then mice received one of the three treatment groups for 10.5 weeks
| ↓ Bacteroidetes-to-Firmicutes in all groups ↓ Bacteroidetes in all groups (more in SFA-rich group) ↓ Porphyromonadaceae in n6 PUFA-rich group ↓ Lachnospiraceae in SFA-rich group | - |
| Ghosh et al. [123] 2013 | Adult mice | Mice were weaned onto two high-fat diets fed for 5 weeks.
| High n6 PUFA: ↑ Enterobacteriaceae ↑ Clostridia spp. High n3 PUFA: ↑ Bifidobacteria ↑ Lactobacillus ↑ Enterococcus faecium | - |
| de Wit et al. [124] 2012 | Adult mice | Standard chow for 3 weeks followed by a low-fat diet based on palm oil for 3 weeks. Then either maintained on the low-fat diet or received high-fat diets for 8 weeks on Palm oil, Olive oil, or Safflower oil | ↑ Firmicutes members bacilli and clostridia ↓ microbiome diversity | ↑ Fecal fat overflow (more in Palm oil diet) ↓ Fat absorption ↑ Intestinal SFA transport |
| Saeedi Saravi et al. [125] 2020 | Old mice | Standard chow until 8–12 weeks of age. One group remained on standard chow and the other mice received modified diets until >18 months of age.
| High αLA diet: ↓ decreased Faith’s phylogenetic richness ↓ Ruminococcaceae ↓ Clostridiaceae ↓ Lachnoclostridium ↑ Bilophila | ↑ Acetate ↓ Trimethylamine N-oxide |
| Marques et al. [126] 2015 | Adult mice | The animals were divided into two groups and received the intervention for 8 weeks.
| ↓ Firmicutes ↓ Bacteroidetes ↓ Desulfovibrionaceae ↓ Peptococcaceae ↑ Porphyromonadaceae | ↑ Acetate ↑ Propionate ↑ Isobutyrate |
| Ghezzal et al. [127] 2020 | Adult mice | Three-month-old male were fed standard chow diet. High fat mice received with palm oil rich in saturated palmitic acid (about 45%) and unsaturated oleic acid (about 35%) | ↓ Clostridium leptum ↓ Akkermansia muciniphila ↑ Bacteroides | ↑ Intestinal permeability |
| Huang et al. [128] 2013 | Adult male mice | Intervention arms included isocaloric high-fat diets, where the dietary fat consisted of:
| ↓ Bacteroides in all groups compared to low fat control ↑ Proteobacteria in milk fat and PUFA groups | - |
| Author & Year | Population | No. of Participants | Supp. Intervention | Supp. Control | n3:n6 Balance | Feeding Type | Start of Intervention | Duration of Supp. | Dose | Primary Outcome | Definition of NEC | Incidence of NEC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bernabe-García et al. [159] 2021 | 1000–1500 g | DHA: 100 Control: 100 | DHA | High-oleic sunflower oil (MUFA) | No | Human milk or enteral formula | 1st feed after birth | 14 days | DHA 75 mg/kg/day | NEC stage ≥ IIa | Modified Bell’s criteria | DHA: 0% Control: 7%; p = 0.007 |
| Marc et al. [160] 2020 | 23–28 weeks gestation | Intervention: 273 Control: 255 | oral capsules of DHA given to mothers | Placebo capsules given to mothers | No | Breast milk | within 72 h of delivery | Until 36 weeks CA | DHA 1.2 g/day of for the intervention group | BPD-free survival | Modified Bell’s criteria | Placebo: 3.0% DHA group: 5.4% p = 0.14 |
| Collins et al. [161] 2017 | <29 weeks gestation | Intervention: 631 Control: 642 | DHA from fish oil | Placebo from soy without DHA | No | Breast milk or formula | within 3 days after their first enteral feeding | Until 36 weeks CA | DHA 60 mg/kg/day | BPD | Proven NEC | Intervention: 8.3% Control: 7.1% p = 0.46 |
| Baack et al. [162] 2016 | 24–34 weeks gestation | Intervention: 31 Control: 29 | DHA liquid | Placebo | No | Breast milk or formula | First week of life | Until discharge or 37 weeks CA | DHA 50 mg/day of for the intervention group | Feasibility and biochemical efficacy | N/A | 0% in both groups |
| Makrides et al. [163] 2009 | <33 weeks gestation | High DHA: 322 Standard DHA: 335 | High DHA: mothers taking tuna oil capsules at ~1% DHA (or high-DHA preterm formula (~1% DHA and 0.6% AA) | Standard DHA: mothers taking soy oil capsules at ~0.3% DHA (or standard preterm formula at ~0.35% DHA and 0.6% AA) | No | For mothers providing breastmilk to their infants | Within 2–4 days of life | Until term CA | High-DHA: six 500-mg tuna oil per day Standard-DHA: six 500-mg soy oil per day | ND at 18 months CA | Not specified | High DHA group: 4.3% Standard DHA group: 2.1% Adjusted p = 0.10 |
| Henriksen et al. [164] 2008 | <1500 g | Intervention: 68 Control: 73 | Soy oil and MCT + AA and DHA as triacylglycerol | Soy oil and MCT without DHA or AA | Yes | Human milk (EBM/DHM) | 1 week after birth | Until discharge (average, 9 weeks) | DHA: 32 mg and AA: 31 mg per 100 mL of human milk | ND at 6 months of age | Modified Bell’s criteria | Control: 2.7% Intervention: 1.5% |
| Groh-Wargo et al. [165] 2005 | 750–1800 g and <33 weeks gestation | Fish/Fungal oil: 20 Egg/Fish oil: 18 Control: 22 | DHA + AA from fish/Fungal oil vs. DHA + AA from egg/fish oil | Regular with non-detected DHA/AA | Yes | Breastmilk or formula | From the first enteral formula feeding | Until 12 months CA | Fish/Fungal 24 kcal: DHA 0.27 g/100 g AA 0.43 g/100 g Egg/Fungal 24 kcal: DHA 0.24 g/100 g AA 0.41 g/100 g | Growth and body composition | Radiographic evidence NEC or surgical NEC | 0% in all groups |
| Clandinin et al. [166] 2005 | ≤35 weeks gestation | Algal-DHA: 112 Fish-DHA: 130 Control: 119 | Algal-DHA with AA from fungal oil vs. fish-DHA with AA from fungal oil | Regular with no DHA or AA | Yes | Formula | Average 30 + 5 to 31.2 wks CA | Until 92 weeks CA | Algal-DHA: 17 mg/100 kcal from algal oil and 34 mg ARA/100 kcal from fungal oil Fish-DHA: 17 mg DHA/100 kcal from fish oil and 34 mg ARA/100 kcal from fungal oil | Growth | Modified Bell’s criteria Stage II or III | Control: 2.5% Algal-DHA: 5.4% Fish-DHA: 5.4% |
| Fewtrell et al. [167] 2004 | ≤2000 g and <35 wks gestation | LC-PUFA: 122 Control: 116 | LC-PUFA | Regular | Unclear | Formula | 14 days | Until 9 months corrected age | AA: 0.4 g/100 g of fat DHA: 0.5 g/100 g of fat γ linolenic acid: 0.9 g/100 g of fat | ND at 18 months | Bell’s criteria or via surgery, or postmortem autopsy | Control: 1.7% LC-PUFA: 4.1% |
| Innis et al. [168] 2002 | VLBW | DHA: 65 DHA + AA: 66 Control: 60 | DHA at 0.34% vs. DHA at 0.33% and + AA at 0.60% | Regular with no DHA or AA | Yes in DHA + AA only | Formula | After reaching an enteral intake of 375 kJ/kg/d | At least 28 days | DHA formula: ~0.15% of energy as DHA DHA + AA formula: 0.14% and 0.27% of energy as DHA and AA, respectively | Growth | Suspected or confirmed | Control: 1.7% DHA: 3.1% DHA + AA: 0% |
| Fewtrell et al. [169] 2002 | <1750 g and preterm | Control: 100 LC-PUFA: 95 Breastfed: 88 | LC-PUFA from vegetable oils, milk fat, evening primrose oil and egg lipids | Preterm infant formula without additional LC-PUFA | Yes | Formula | 5 ± 4 days after birth | Control: 33 ± 17 days LC-PUFA: 31 ± 21 days | AA 0.31 g/100 g of fat DHA 0.17 g/100 g of fat | ND at 18 months | Bell’s criteria or via surgery, or postmortem autopsy | Control: 2% LC-PUFA group: 5.3% p = 0.11 |
| O’Connor et al. [170] 2001 | 750–1800 g | AA + DHA from Fish/Fungal oil: 140 AA + DHA from egg-TG/Fish oil: 143 Control: 144 | AA + DHA (Fish/Fungal oil) vs. AA + DHA (egg-TG/Fish oil | Regular (non-detected DHA/AA) | Yes | Human milk or formula | within 72 h of the first feed | To 12 months CA | AA + DHA (Fish/Fungal oil) -In hospital: DHA 0.27 g/100 g total fatty acids, AA 0.43 g/100 g AA + DHA (Egg-TG/Fish oil) - In hospital: DHA 0.24 g/100 g AA 0.41 g/100 g | growth, visual acuity, and ND | Bell’s criteria or via surgery, or postmortem autopsy | Control: 4% AA + DHA (Fish/Fungal): 4% AA + DHA (Egg-TG/Fish): 3% |
| Carlson et al. [171] 1998 | 725–1375 g & ≤32 wks gestation | DHA + AA: 34 Control: 85 | Egg phospholipid: DHA (0.13%) AA (0.41%) | Regular | Yes | Formula | At a mean of 4.9 days of age | After discharge home | 7-fold more esterified choline, AA, and DHA in experimental formula | NEC stage II or III | Modified Bell’s criteria | DHA + AA: 2.9% Control: 17.6% p < 0.05 |
| Carlson et al. [172] 1996 | 747–1275 g | DHA + EPA: 35 Control: 36 | Marine-oil-supplemented formulas at 0.2% DHA & 0.06% EPA | Standard preterm formula | No | Preterm formula until 2 months CA | 3–5 days of age | Until 48 ± 1 weeks CA | Standard preterm formula contained LA (1.2% of energy) | Visual acuity and growth | Not specified | DHA + EPA: 25.7% Control: 8.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alshaikh, B.N.; Reyes Loredo, A.; Knauff, M.; Momin, S.; Moossavi, S. The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis. Nutrients 2022, 14, 145. https://doi.org/10.3390/nu14010145
Alshaikh BN, Reyes Loredo A, Knauff M, Momin S, Moossavi S. The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis. Nutrients. 2022; 14(1):145. https://doi.org/10.3390/nu14010145
Chicago/Turabian StyleAlshaikh, Belal N., Adriana Reyes Loredo, Megan Knauff, Sarfaraz Momin, and Shirin Moossavi. 2022. "The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis" Nutrients 14, no. 1: 145. https://doi.org/10.3390/nu14010145
APA StyleAlshaikh, B. N., Reyes Loredo, A., Knauff, M., Momin, S., & Moossavi, S. (2022). The Role of Dietary Fats in the Development and Prevention of Necrotizing Enterocolitis. Nutrients, 14(1), 145. https://doi.org/10.3390/nu14010145