
Comparing the Fasting and Random-Fed Metabolome Response to an Oral Glucose Tolerance Test in Children and Adolescents: Implications of Sex, Obesity, and Insulin Resistance

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Design
2.2. Laboratory Measurements
2.3. Untargeted Metabolomics
2.4. Statistical Analyses
3. Results
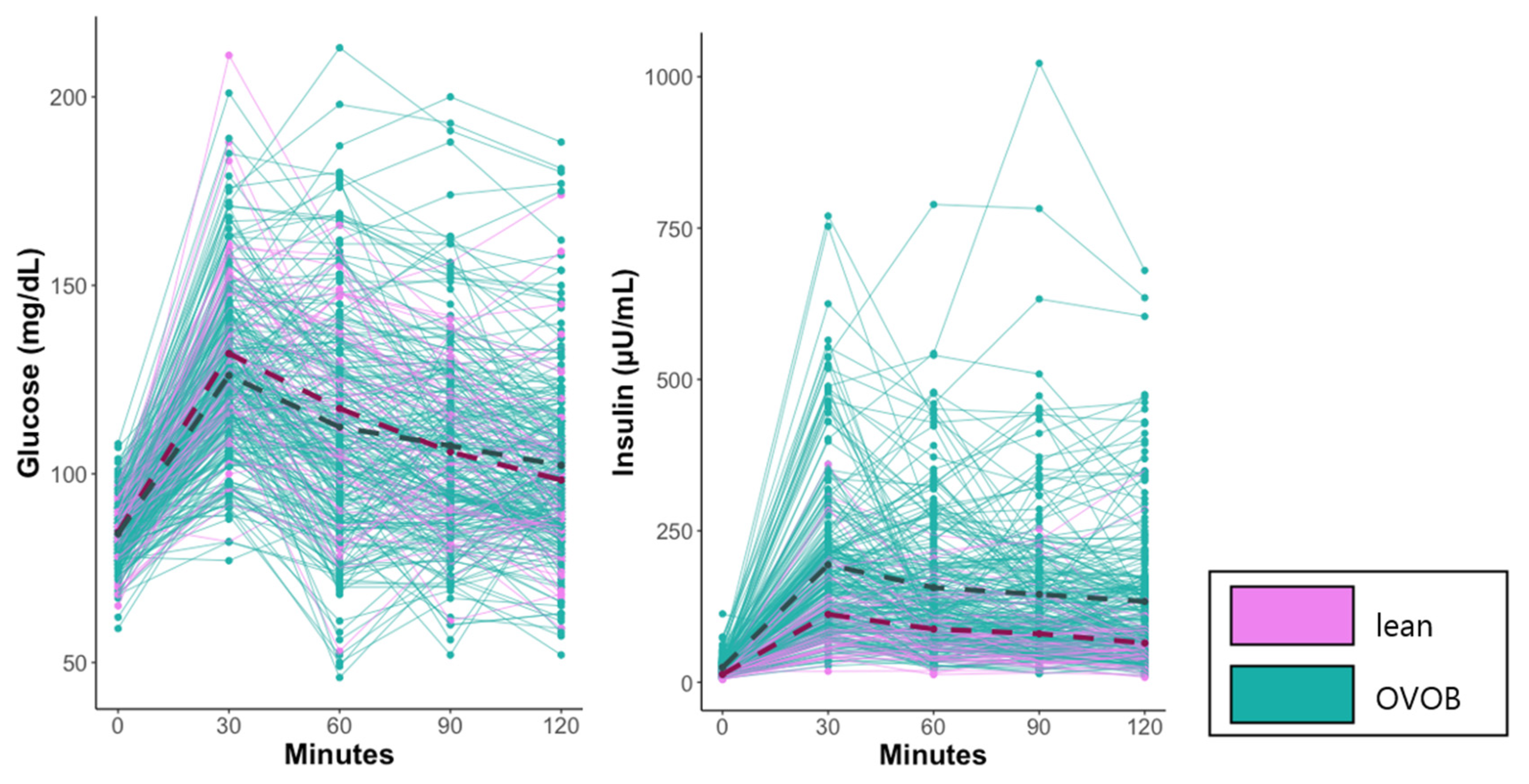
3.1. Participant Characteristics
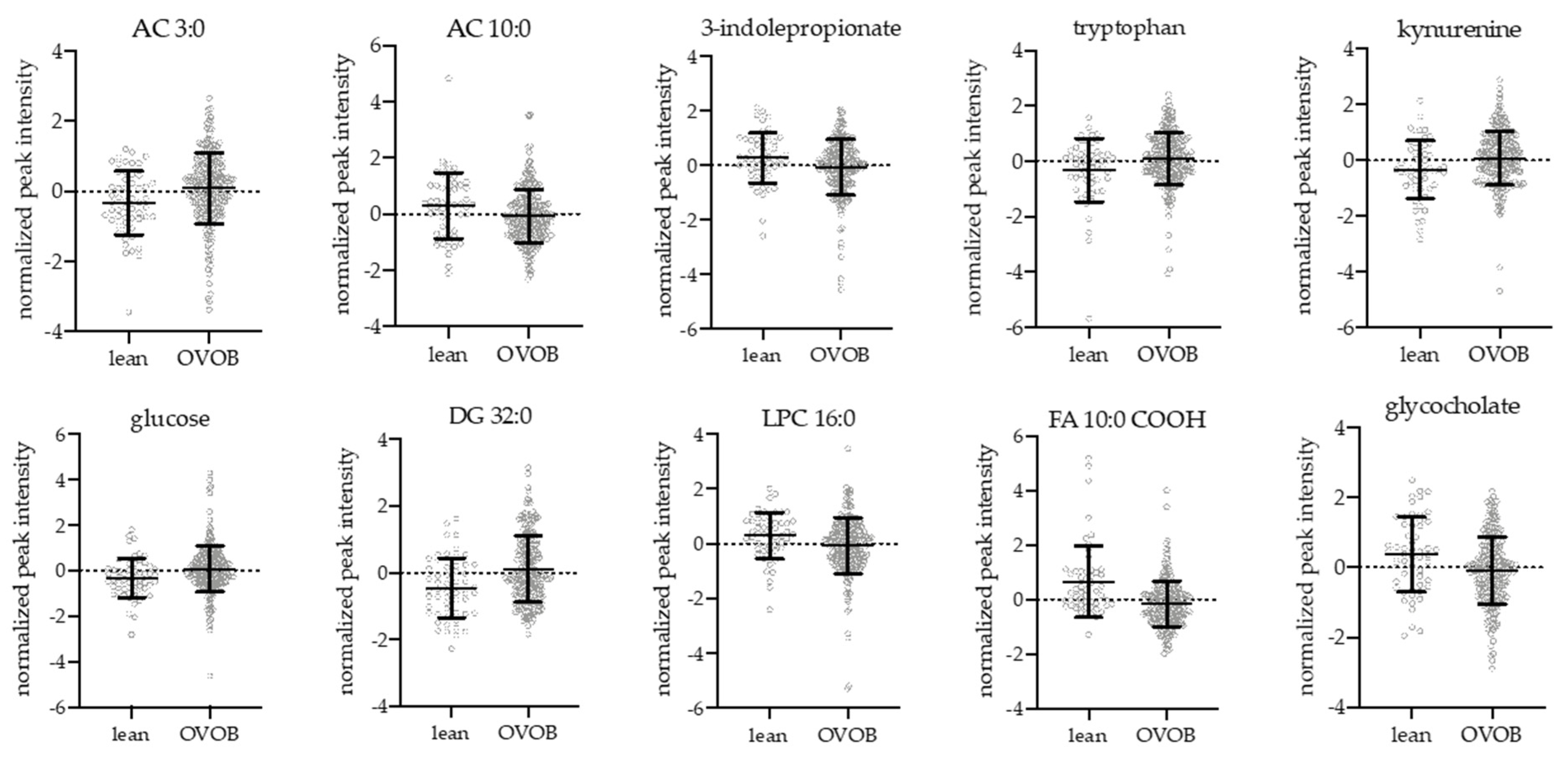
3.2. Influence of Obesity and Sex on the Fasting Metabolome
3.3. Metabolome Response to the OGTT in OVOB and Lean Participants
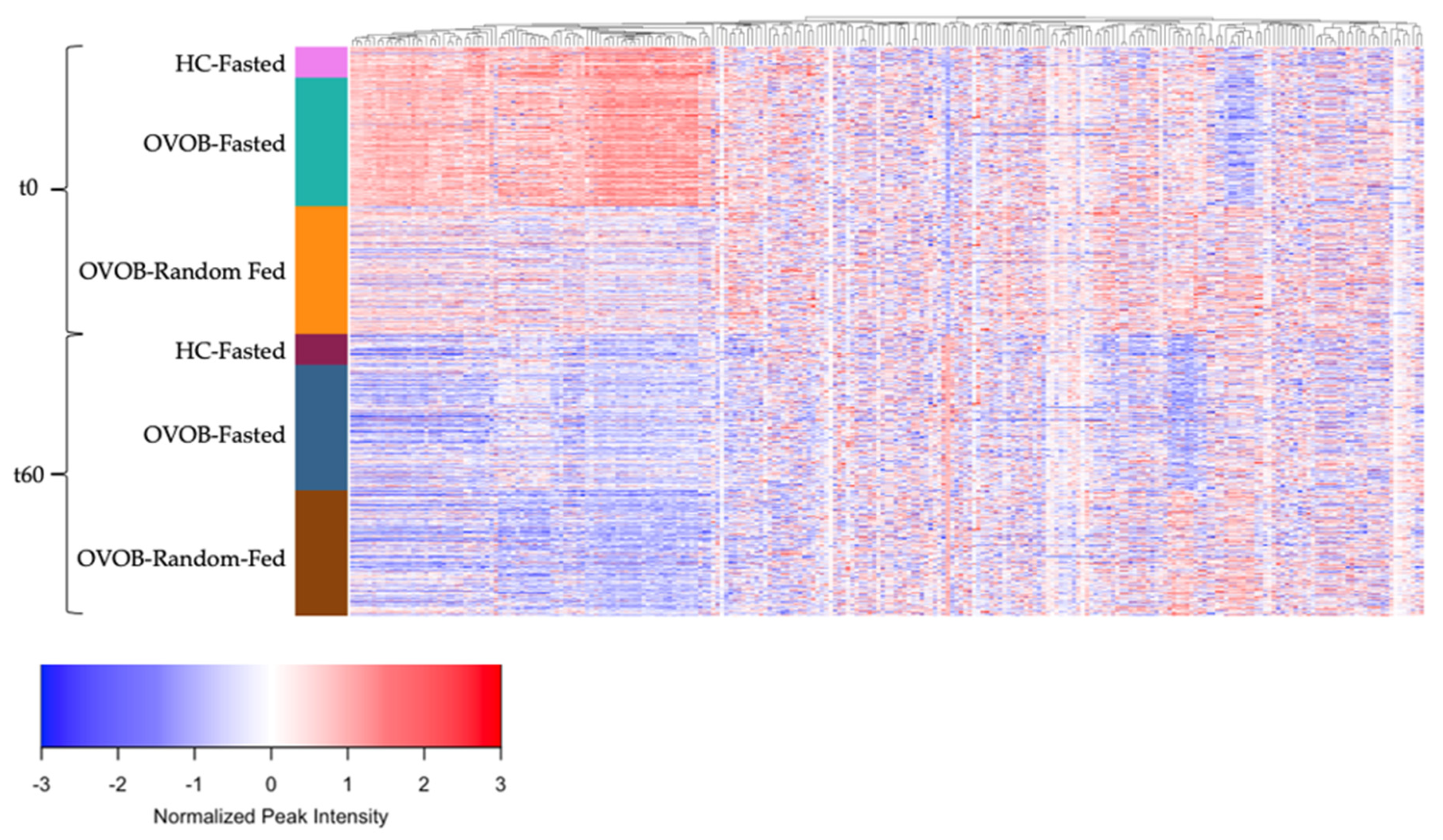
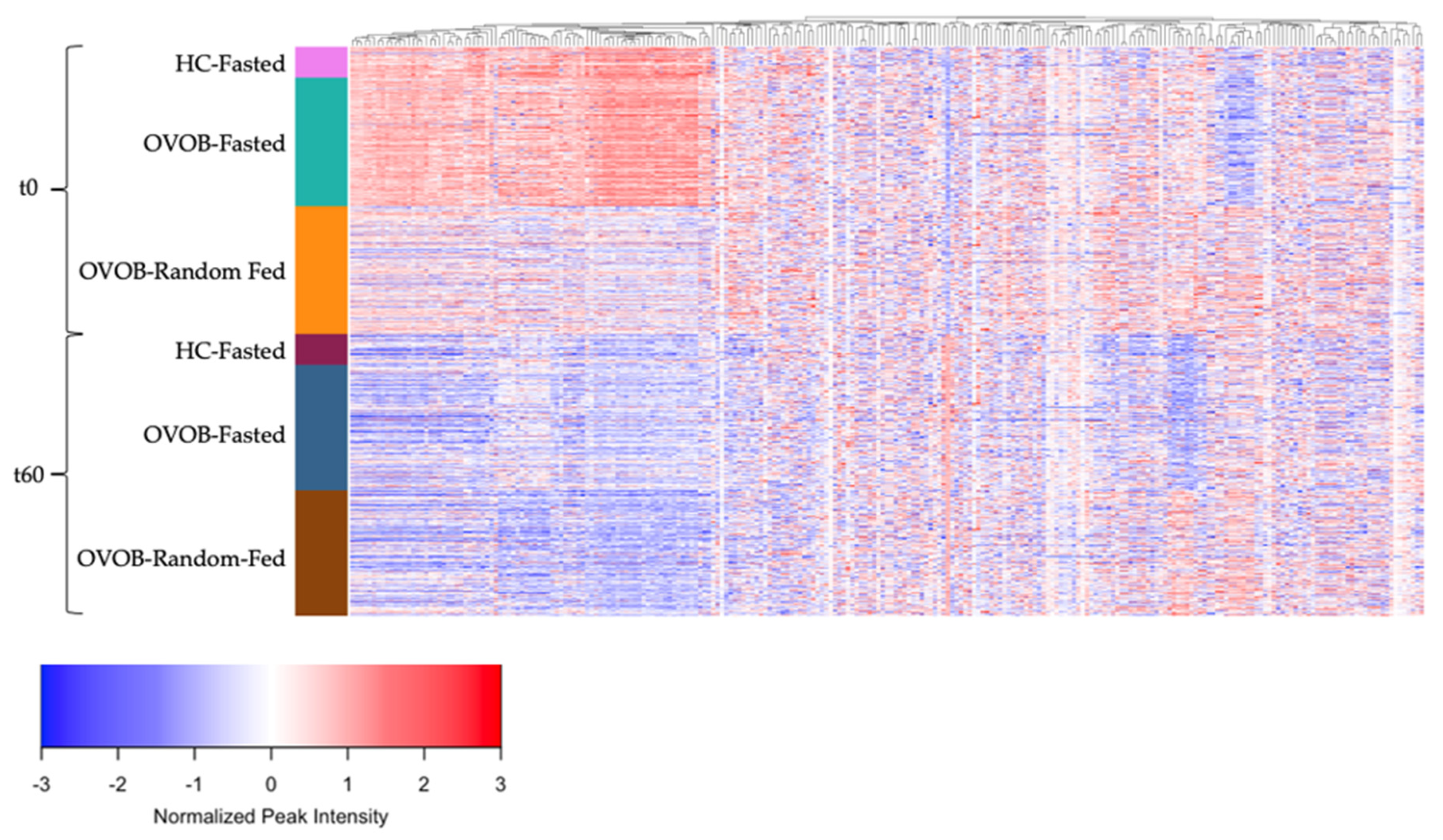
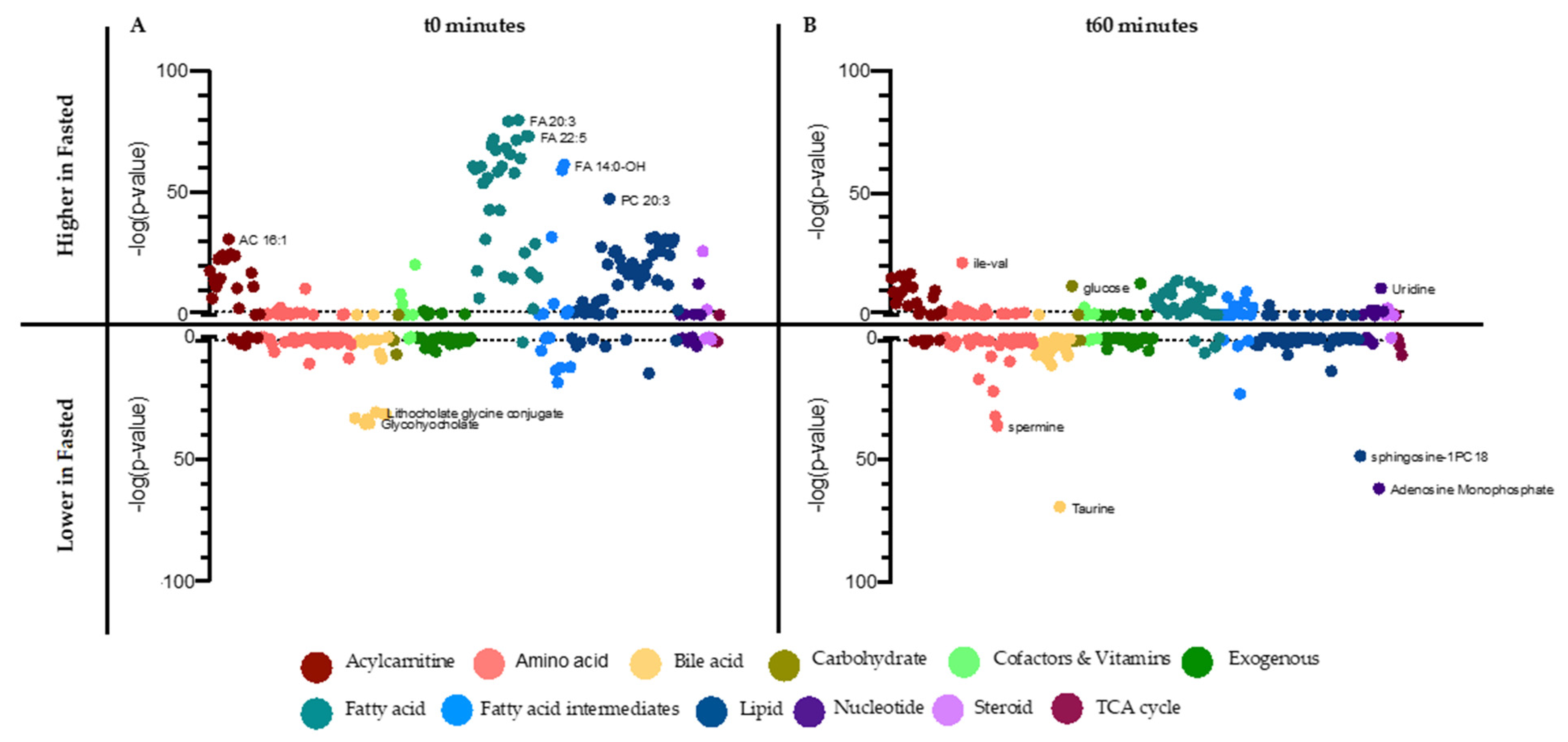
3.4. Metabolome Differences between the Fasted and Random-Fed OGTT Challenge in OVOB
3.5. Sex-Specific Associations of Metabolite Trajectories with Insulin Resistance in Participants with Overweight and Obesity
4. Discussion
4.1. Lipids, Fatty Acids, and Acylcarnitines
4.2. Amino Acids
4.3. Bile Acids
4.4. Conclustions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lascar, N.; Brown, J.; Pattison, H.; Barnett, A.H.; Bailey, C.J.; Bellary, S. Type 2 diabetes in adolescents and young adults. Lancet Diabetes Endocrinol. 2018, 6, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Shaham, O.; Wei, R.; Wang, T.J.; Ricciardi, C.; Lewis, G.D.; Vasan, R.S.; Carr, S.A.; Thadhani, R.; Gerszten, R.E.; Mootha, V.K. Metabolic profiling of the human response to a glucose challenge reveals distinct axes of insulin sensitivity. Mol. Syst. Biol. 2008, 4, 214. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.E.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; Cheng, S.; Rhee, E.P.; Florez, J.C.; Clish, C.B.; Gerszten, R.E.; Wang, T.J. Metabolite profiles during oral glucose challenge. Diabetes 2013, 62, 2689–2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diniz Behn, C.; Jin, E.S.; Bubar, K.; Malloy, C.; Parks, E.J.; Cree-Green, M. Advances in stable isotope tracer methodology part 1: Hepatic metabolism via isotopomer analysis and postprandial lipolysis modeling. J. Investig. Med. 2020, 68, 3–10. [Google Scholar] [CrossRef]
- Fukagawa, N.K.; Minaker, K.L.; Rowe, J.W.; Goodman, M.N.; Matthews, D.E.; Bier, D.M.; Young, V.R. Insulin-mediated reduction of whole body protein breakdown. Dose-response effects on leucine metabolism in postabsorptive men. J. Clin. Investig. 1985, 76, 2306–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twig, G.; Zucker, I.; Afek, A.; Cukierman-Yaffe, T.; Bendor, C.D.; Derazne, E.; Lutski, M.; Shohat, T.; Mosenzon, O.; Tzur, D.; et al. Adolescent Obesity and Early-Onset Type 2 Diabetes. Diabetes Care 2020, 43, 1487–1495. [Google Scholar] [CrossRef]
- LaBarre, J.L.; Peterson, K.E.; Kachman, M.T.; Perng, W.; Tang, L.; Hao, W.; Zhou, L.; Karnovsky, A.; Cantoral, A.; Tellez-Rojo, M.M.; et al. Mitochondrial Nutrient Utilization Underlying the Association Between Metabolites and Insulin Resistance in Adolescents. J. Clin. Endocrinol. Metab. 2020, 105. [Google Scholar] [CrossRef]
- Butte, N.F.; Liu, Y.; Zakeri, I.F.; Mohney, R.P.; Mehta, N.; Voruganti, V.S.; Goring, H.; Cole, S.A.; Comuzzie, A.G. Global metabolomic profiling targeting childhood obesity in the Hispanic population. Am. J. Clin. Nutr. 2015, 102, 256–267. [Google Scholar] [CrossRef]
- Perng, W.; Gillman, M.W.; Fleisch, A.F.; Michalek, R.D.; Watkins, S.M.; Isganaitis, E.; Patti, M.E.; Oken, E. Metabolomic profiles and childhood obesity. Obesity 2014, 22, 2570–2578. [Google Scholar] [CrossRef]
- Geidenstam, N.; Spegel, P.; Mulder, H.; Filipsson, K.; Ridderstrale, M.; Danielsson, A.P. Metabolite profile deviations in an oral glucose tolerance test-a comparison between lean and obese individuals. Obesity 2014, 22, 2388–2395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullner, E.; Rohnisch, H.E.; von Bromssen, C.; Moazzami, A.A. Metabolomics analysis reveals altered metabolites in lean compared with obese adolescents and additional metabolic shifts associated with hyperinsulinaemia and insulin resistance in obese adolescents: A cross-sectional study. Metabolomics 2021, 17, 11. [Google Scholar] [CrossRef]
- Lee, J.M.; Eason, A.; Nelson, C.; Kazzi, N.G.; Cowan, A.E.; Tarini, B.A. Screening practices for identifying type 2 diabetes in adolescents. J. Adolesc. Health 2014, 54, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Gebremariam, A.; Wu, E.L.; LaRose, J.; Gurney, J.G. Evaluation of nonfasting tests to screen for childhood and adolescent dysglycemia. Diabetes Care 2011, 34, 2597–2602. [Google Scholar] [CrossRef] [Green Version]
- International Diabetes Federation Guideline Development Group. Guideline for management of postmeal glucose in diabetes. Diabetes Res. Clin. Pr. 2014, 103, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Carayol, M.; Licaj, I.; Achaintre, D.; Sacerdote, C.; Vineis, P.; Key, T.J.; Onland Moret, N.C.; Scalbert, A.; Rinaldi, S.; Ferrari, P. Reliability of Serum Metabolites over a Two-Year Period: A Targeted Metabolomic Approach in Fasting and Non-Fasting Samples from EPIC. PLoS ONE 2015, 10, e0135437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guasch-Ferre, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasun, P. Role of mitochondria in pathogenesis of type 2 diabetes mellitus. J. Diabetes Metab. Disord. 2020, 19, 2017–2022. [Google Scholar] [CrossRef] [PubMed]
- Grzych, G.; Chávez-Talavera, O.; Descat, A.; Thuillier, D.; Verrijken, A.; Kouach, M.; Legry, V.; Verkindt, H.; Raverdy, V.; Legendre, B.; et al. NASH-related increases in plasma bile acid levels depend on insulin resistance. J. Heptalogy Rep. 2020, 3, 100222. [Google Scholar] [CrossRef]
- Nowak, C.; Hetty, S.; Salihovic, S.; Castillejo-Lopez, C.; Ganna, A.; Cook, N.L.; Broeckling, C.D.; Prenni, J.E.; Shen, X.; Giedraitis, V.; et al. Glucose challenge metabolomics implicates medium-chain acylcarnitines in insulin resistance. Sci. Rep. 2018, 8, 8691. [Google Scholar] [CrossRef]
- Kuczmarski, R.J.; Ogden, C.L.; Grummer-Strawn, L.M.; Flegal, K.M.; Guo, S.S.; Wei, R.; Mei, Z.; Curtin, L.R.; Roche, A.F.; Johnson, C.L. CDC growth charts: United States. Adv. Data 2000, 314, 1–27. [Google Scholar]
- Hayashi, M.; Floyd, J.C., Jr.; Pek, S.; Fajans, S.S. Insulin, proinsulin, glucagon and gastrin in pancreatic tumors and in plasma of patients with organic hyperinsulinism. J. Clin. Endocrinol. Metab. 1977, 44, 681–694. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes, A. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2020. Diabetes Care 2020, 43, S14–S31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. MSEA: A web-based tool to identify biologically meaningful patterns in quantitative metabolomic data. Nucleic Acids Res. 2010, 38, W71–W77. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic. Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.E.; Larson, M.G.; Ghorbani, A.; Cheng, S.; Chen, M.H.; Keyes, M.; Rhee, E.P.; Clish, C.B.; Vasan, R.S.; Gerszten, R.E.; et al. Metabolomic Profiles of Body Mass Index in the Framingham Heart Study Reveal Distinct Cardiometabolic Phenotypes. PLoS ONE 2016, 11, e0148361. [Google Scholar] [CrossRef] [Green Version]
- de Mello, V.D.; Paananen, J.; Lindstrom, J.; Lankinen, M.A.; Shi, L.; Kuusisto, J.; Pihlajamaki, J.; Auriola, S.; Lehtonen, M.; Rolandsson, O.; et al. Indolepropionic acid and novel lipid metabolites are associated with a lower risk of type 2 diabetes in the Finnish Diabetes Prevention Study. Sci. Rep. 2017, 7, 46337. [Google Scholar] [CrossRef]
- Heimerl, S.; Fischer, M.; Baessler, A.; Liebisch, G.; Sigruener, A.; Wallner, S.; Schmitz, G. Alterations of plasma lysophosphatidylcholine species in obesity and weight loss. PLoS ONE 2014, 9, e111348. [Google Scholar] [CrossRef] [Green Version]
- Kubota, K.; Ishizaki, T. Dose-dependent pharmacokinetics of benzoic acid following oral administration of sodium benzoate to humans. Eur. J. Clin. Pharm. 1991, 41, 363–368. [Google Scholar] [CrossRef]
- Owen, O.E.; Reichard, G.A., Jr.; Patel, M.S.; Boden, G. Energy metabolism in feasting and fasting. Adv. Exp. Med. Biol. 1979, 111, 169–188. [Google Scholar] [CrossRef]
- Meessen, E.C.; Bakker, G.J.; Nieuwdorp, M.; Dallinga-Thie, G.M.; Kemper, E.M.; Damink, S.W.O.; Romijn, J.A.; Hartmann, B.; Holst, J.J.; Knop, F.K.; et al. Parenteral nutrition impairs plasma bile acid and gut hormone responses to mixed meal testing in lean healthy men. Clin. Nutr. 2020, 40, 1013–1021. [Google Scholar] [CrossRef]
- de Miranda, J.A.; Almeida, G.G.; Martins, R.I.; Cunha, M.B.; Belo, V.A.; dos Santos, J.E.; Mourao-Junior, C.A.; Lanna, C.M. The role of uric acid in the insulin resistance in children and adolescents with obesity. Rev. Paul Pediatr. 2015, 33, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Peter, A.; Fritsche, J.; Elcnerova, M.; Fritsche, A.; Haring, H.U.; Schleicher, E.D.; Xu, G.; Lehmann, R. Changes of the plasma metabolome during an oral glucose tolerance test: Is there more than glucose to look at? Am. J. Physiol. Endocrinol. Metab. 2009, 296, E384–E393. [Google Scholar] [CrossRef] [PubMed]
- Schooneman, M.G.; Vaz, F.M.; Houten, S.M.; Soeters, M.R. Acylcarnitines: Reflecting or inflicting insulin resistance? Diabetes 2013, 62, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, D.E.; Mandarino, L.J. Fuel selection in human skeletal muscle in insulin resistance: A reexamination. Diabetes 2000, 49, 677–683. [Google Scholar] [CrossRef] [Green Version]
- Dimitriadis, G.; Mitrou, P.; Lambadiari, V.; Maratou, E.; Raptis, S.A. Insulin effects in muscle and adipose tissue. Diabetes Res. Clin. Pr. 2011, 93 (Suppl. 1), S52–S59. [Google Scholar] [CrossRef]
- Mihalik, S.J.; Michaliszyn, S.F.; de las Heras, J.; Bacha, F.; Lee, S.; Chace, D.H.; DeJesus, V.R.; Vockley, J.; Arslanian, S.A. Metabolomic profiling of fatty acid and amino acid metabolism in youth with obesity and type 2 diabetes: Evidence for enhanced mitochondrial oxidation. Diabetes Care 2012, 35, 605–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangipurapu, J.; Stancakova, A.; Smith, U.; Kuusisto, J.; Laakso, M. Nine Amino Acids Are Associated With Decreased Insulin Secretion and Elevated Glucose Levels in a 7.4-Year Follow-up Study of 5,181 Finnish Men. Diabetes 2019, 68, 1353–1358. [Google Scholar] [CrossRef]
- Angelin, B.; Bjorkhem, I.; Einarsson, K.; Ewerth, S. Hepatic uptake of bile acids in man. Fasting and postprandial concentrations of individual bile acids in portal venous and systemic blood serum. J. Clin. Investig. 1982, 70, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Liddle, R.A.; Goldfine, I.D.; Rosen, M.S.; Taplitz, R.A.; Williams, J.A. Cholecystokinin bioactivity in human plasma. Molecular forms, responses to feeding, and relationship to gallbladder contraction. J. Clin. Investig. 1985, 75, 1144–1152. [Google Scholar] [CrossRef] [PubMed]
- Legry, V.; Francque, S.; Haas, J.T.; Verrijken, A.; Caron, S.; Chavez-Talavera, O.; Vallez, E.; Vonghia, L.; Dirinck, E.; Verhaegen, A.; et al. Bile Acid Alterations Are Associated With Insulin Resistance, but Not With NASH, in Obese Subjects. J. Clin. Endocrinol. Metab. 2017, 102, 3783–3794. [Google Scholar] [CrossRef] [PubMed]
- Hannon, T.S.; Janosky, J.; Arslanian, S.A. Longitudinal study of physiologic insulin resistance and metabolic changes of puberty. Pediatr. Res. 2006, 60, 759–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballerini, M.G.; Bergada, I.; Rodriguez, M.E.; Keselman, A.; Bengolea, V.S.; Pipman, V.; Domene, H.M.; Jasper, H.G.; Ropelato, M.G. Insulin level and insulin sensitivity indices among healthy children and adolescents. Arch. Argent. Pediatr. 2016, 114, 329–336. [Google Scholar] [CrossRef]
- Arslanian, S.A.; Heil, B.V.; Becker, D.J.; Drash, A.L. Sexual dimorphism in insulin sensitivity in adolescents with insulin-dependent diabetes mellitus. J. Clin. Endocrinol. Metab. 1991, 72, 920–926. [Google Scholar] [CrossRef] [PubMed]
- Iyer, G.R.; Wigginton, J.; Duren, W.; LaBarre, J.L.; Brandenburg, M.; Burant, C.; Michailidis, G.; Karnovsky, A. Application of Differential Network Enrichment Analysis for Deciphering Metabolic Alterations. Metabolites 2020, 10, 479. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categorical Variables | OVOB | Lean | p-Value 1 |
| n(%) | n(%) | ||
| sex | |||
| Male | 97 (43%) | 26 (47%) | 0.5254 |
| Female | 131 (57%) | 29 (53%) | |
| race | |||
| Asian/Pacific Islander | 4 (2%) | 4 (7%) | 0.1745 |
| African American/Black | 59 (26%) | 12 (22%) | |
| White | 135 (59%) | 32 (58%) | |
| more than one race | 19 (8%) | 6 (11%) | |
| did not wish to report | 11 (5%) | 1 (2%) | |
| ethnicity | |||
| Hispanic | 18 (8%) | 5 (9%) | 0.7707 |
| non-Hispanic | 210 (92%) | 50 (91%) | |
| abnormal 2-hr plasma glucose (≥140 mg/dL) | |||
| Yes | 16 (7%) | 3 (5%) | 0.6776 |
| No | 212 (93%) | 52 (95%) | |
| abnormal fasting plasma glucose (≥100 mg/dL) | |||
| Yes | 8 (4%) | 1 (2%) | 0.5213 |
| No | 220 (96%) | 54 (98%) | |
| ADA prediabetes (FPG≥ 100 mg/dL or 2-hr PG ≥ 140 mg/dL or HbA1c ≥ 5.7%) | |||
| Yes | 35 (15%) | 6 (12%) | 0.4009 |
| No | 193 (85%) | 49 (88%) | |
| Continuous Variables | OVOB | Lean | p-Value 2 |
| Mean (SD) | Mean (SD) | ||
| age (years) | 12.9 (2.5) | 13.0 (2.6) | 0.7301 |
| BMI percentile | 95 (4) | 59 (27) | 7.52−14 |
| HOMA-IR | 5.13 (2.99) | 2.80 (1.21) 3 | 4.53−8 |
| HbA1c | 5.2 (0.3) | 5.1 (0.3) 3 | 0.2082 |
| fast time (hours) | 14.0 (1.3) | 14.1 (1.4) | 0.7606 |
| fasting OGTT response | |||
| glucose (t0) (mg/dL) | 84 (8) | 85 (8) | 0.7475 |
| glucose (t30) (mg/dL) | 126 (22) | 132 (25) | 0.1119 |
| glucose (t60) (mg/dL) | 112 (29) | 117 (26) | 0.2182 |
| glucose (t90) (mg/dL) | 107 (26) | 106 (21) | 0.6263 |
| glucose (t120) (mg/dL) | 102 (24) | 98 (22) | 0.2562 |
| insulin (t0) (µU/mL) | 24 (14) | 13 (5) 3 | 1.55 × 10−8 |
| insulin (t30) (µU/mL) | 194 (132) | 112 (72) | 4.09 × 10−9 |
| insulin (t60) (µU/mL) | 156 (117) | 87 (53) | 7.04 × 10−10 |
| insulin (t90) (µU/mL) | 145 (126) | 80 (56) | 2.69 × 10−8 |
| insulin (t120) (µU/mL) | 133 (117) | 65 (57) | 2.42 × 10−9 |
| AUC glucose | 3121 (2112) | 3294 (1711) | 0.5228 |
| AUC insulin | 17,223 (11,206) | 9648 (5468) | 1.33 × 10−11 |
| Metabolite | Pathway | Fasted t0 | Fasted t60 | Fasted Fold Change | Random-Fed t0 | Random-Fed t60 |
|---|---|---|---|---|---|---|
| AC 12:0 | acylcarnitine | 2.0 ± 0.3 | ||||
| AC 12:1 | acylcarnitine | 2.7 ± 0.5 | ||||
| AC 14:0 | acylcarnitine | 2.3 ± 0.8 | ||||
| AC 16:0 | acylcarnitine | 2.6 ± 0.6 | ||||
| AC 16:1 | acylcarnitine | 1.3 ± 0.7 | ||||
| AC 18:0 | acylcarnitine | 1.6 ± 0.2 | ||||
| AC 5:0-OH | acylcarnitine | 0.9 ± 0.3 | ||||
| AC 5:1 | acylcarnitine | 1.5 ± 1.6 | ||||
| AC 6:0 | acylcarnitine | 1.7 ± 0.6 | ||||
| gamma-glutamyltyrosine | amino acid | 0.8 ± 0.2 | 0.9 ± 0.3 | 0.9 ± 0.3 | ||
| Glu-Phe | amino acid | 0.9 ± 0.2 | ||||
| glutamate | amino acid | 0.7 ± 0.3 | 0.9 ± 0.3 | |||
| indole-3-methyl acetate | amino acid | 0.7 ± 0.3 | ||||
| L-gamma-glutamylisoleucine | amino acid | 0.8 ± 0.3 | ||||
| Leu-Ile | amino acid | 0.7 ± 0.5 | 0.9 ± 0.2 | |||
| leucine+isoleucine | amino acid | 0.7 ± 0.3 | ||||
| N-acetylphenylalanine | amino acid | 0.7 ± 0.3 | ||||
| Phe-Phe | amino acid | −0.7 ± 0.3 | ||||
| Phe-Trp | amino acid | 0.7 ± 0.3 | ||||
| pipecolate | amino acid | −0.6 ± 1.1 | ||||
| proline | amino acid | 0.7 ± 0.3 | ||||
| cholate | bile acid | 0.8 ± 0.3 | ||||
| hyocholate | bile acid | 0.8 ± 0.3 | ||||
| indole-3-lactate | carbohydrate | 0.7 ± 0.3 | ||||
| caffeine | exogenous | 1.7 ± 0.6 | ||||
| FA 18:4 | fatty acid | 0.8 ± 0.3 | ||||
| FA 20:3 | fatty acid | 1.1 ± 0.3 | ||||
| FA 22:1 | fatty acid | 1.9 ± 0.5 | ||||
| 3-hydroxyphenyl-valerate | fatty acid intermediate | 0.9 ± 0.6 | ||||
| DG 32:0 | lipid | 1.2 ± 0.2 | 1.2 ± 0.2 | 2.2 ± 0.4 | 1.1 ± 0.2 | 1.0 ± 0.2 |
| DG 32:1 | lipid | 1.0 ± 0.2 | 1.1 ± 0.2 | 0.9 ± 0.2 | 0.8 ± 0.2 | |
| DG 34:1 | lipid | 1.0 ± 0.2 | 0.9 ± 0.2 | 0.9 ± 0.2 | ||
| DG 34:2 | lipid | 1.1 ± 0.2 | 1.0 ± 0.2 | 0.9 ± 0.2 | 0.8 ± 0.2 | |
| DG 36:2 | lipid | 0.7 ± 0.2 | ||||
| DG 36:3 | lipid | 0.7 ± 0.3 | ||||
| MG 14:0 | lipid | 1.0 ± 0.2 | ||||
| MG 16:0 | lipid | 1.2 ± 0.3 | ||||
| MG 18:1 | lipid | 0.8 ± 0.2 | 0.9 ± 0.2 | 0.7 ± 0.3 | ||
| LPC 16:0 | lipid | 1.1 ± 1.2 | ||||
| LPC 18:2 | lipid | 0.8 ± 1.7 | ||||
| PC 32:1 | lipid | 0.8 ± 0.2 | 1.0 ± 0.4 | |||
| PC 34:3 | lipid | 0.7 ± 0.3 | ||||
| PC 34:4 | lipid | 0.7 ± 0.3 | ||||
| N2,N2-dimethylguanosine | nucleotide | 0.7 ± 0.3 | ||||
| urate | nucleotide | 0.8 ± 0.2 | 1.0 ± 0.2 | 1.0 ± 0.2 | 0.9 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaBarre, J.L.; Hirschfeld, E.; Soni, T.; Kachman, M.; Wigginton, J.; Duren, W.; Fleischman, J.Y.; Karnovsky, A.; Burant, C.F.; Lee, J.M. Comparing the Fasting and Random-Fed Metabolome Response to an Oral Glucose Tolerance Test in Children and Adolescents: Implications of Sex, Obesity, and Insulin Resistance. Nutrients 2021, 13, 3365. https://doi.org/10.3390/nu13103365
LaBarre JL, Hirschfeld E, Soni T, Kachman M, Wigginton J, Duren W, Fleischman JY, Karnovsky A, Burant CF, Lee JM. Comparing the Fasting and Random-Fed Metabolome Response to an Oral Glucose Tolerance Test in Children and Adolescents: Implications of Sex, Obesity, and Insulin Resistance. Nutrients. 2021; 13(10):3365. https://doi.org/10.3390/nu13103365
Chicago/Turabian StyleLaBarre, Jennifer L., Emily Hirschfeld, Tanu Soni, Maureen Kachman, Janis Wigginton, William Duren, Johanna Y. Fleischman, Alla Karnovsky, Charles F. Burant, and Joyce M. Lee. 2021. "Comparing the Fasting and Random-Fed Metabolome Response to an Oral Glucose Tolerance Test in Children and Adolescents: Implications of Sex, Obesity, and Insulin Resistance" Nutrients 13, no. 10: 3365. https://doi.org/10.3390/nu13103365