The Anabolic Response to Dietary Protein Is Not Limited by the Maximal Stimulation of Protein Synthesis in Healthy Older Adults: A Randomized Crossover Trial

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
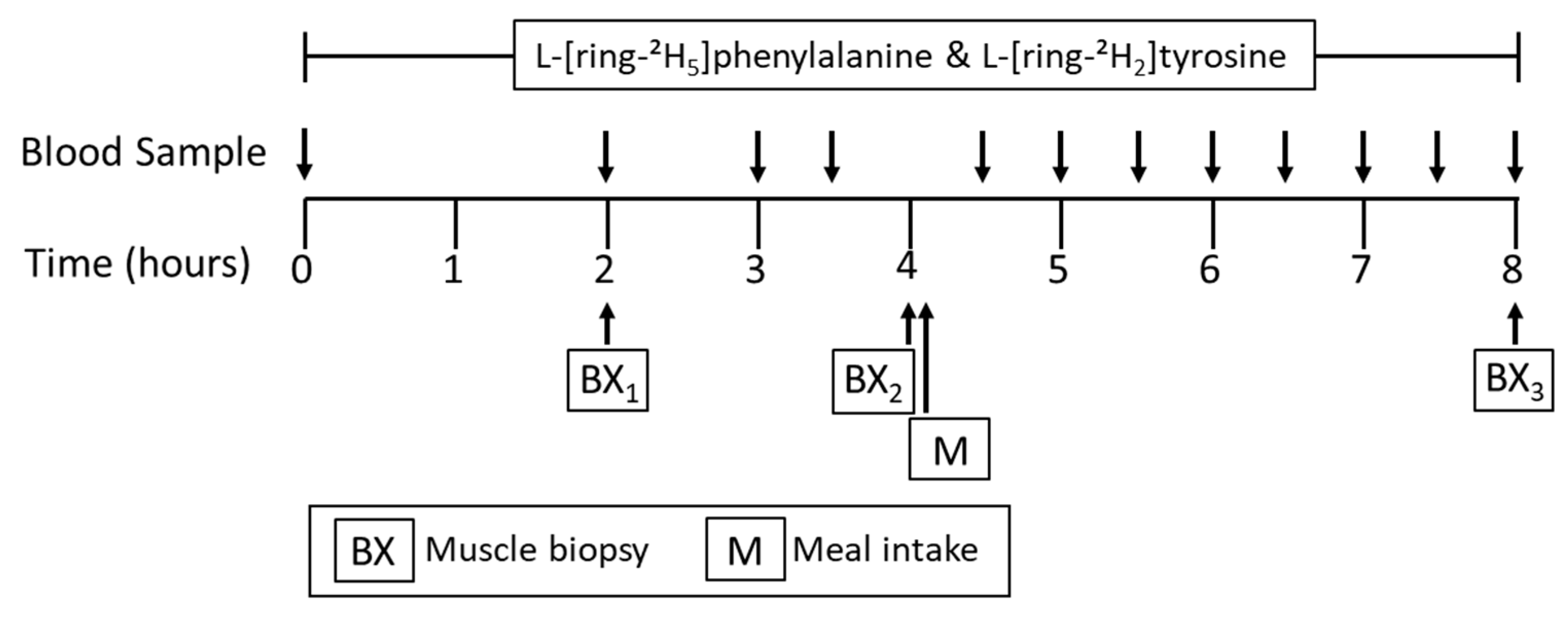
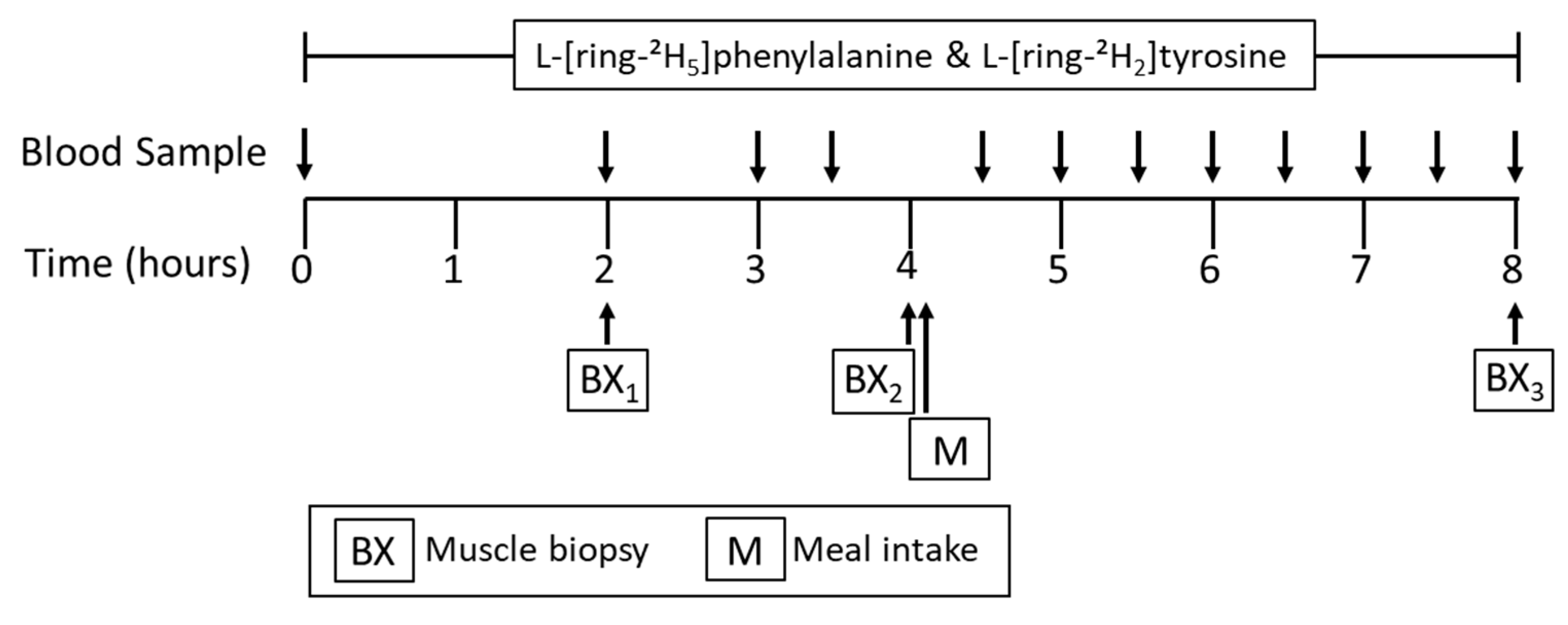
2.2. Experimental Design
2.3. Preparation of Interventional Meals
2.4. Stable Isotope Tracer Infusion Protocol
2.5. Calculations of Protein Kinetics
- Total appearance rate into plasma (Ra) (µmol/kg/min) = F/E
- Fractional Ra of Tyr from Phe (%/100) = ETyr M + 4/EPhe M + 5
- Rate of Phe hydroxylation (HYDROX) (µmol/kg/min) = fractional Ra of Tyr from Phe × Ra TyrMPE
- Rate of protein synthesis (µmol/kg/min) = [(Ra Phe − HYDROX)/0.04]
- Ra Exo = (Protein intake × GITD) × (1 fraction of Ra Phe HYDROX)
- Protein breakdown rate (µmol/kg/min) = [(Ra Phe − FPhe)/0.04 − Ra Exo]
- Net protein balance (µmol/kg/min) = Protein synthesis rate − Protein breakdown rate
- MPS (%/h) = [(EBP2 − EBP1)/(EIC × t)] × 60 × 100
2.6. Analytic Methods
2.7. Statistical Analysis
3. Results
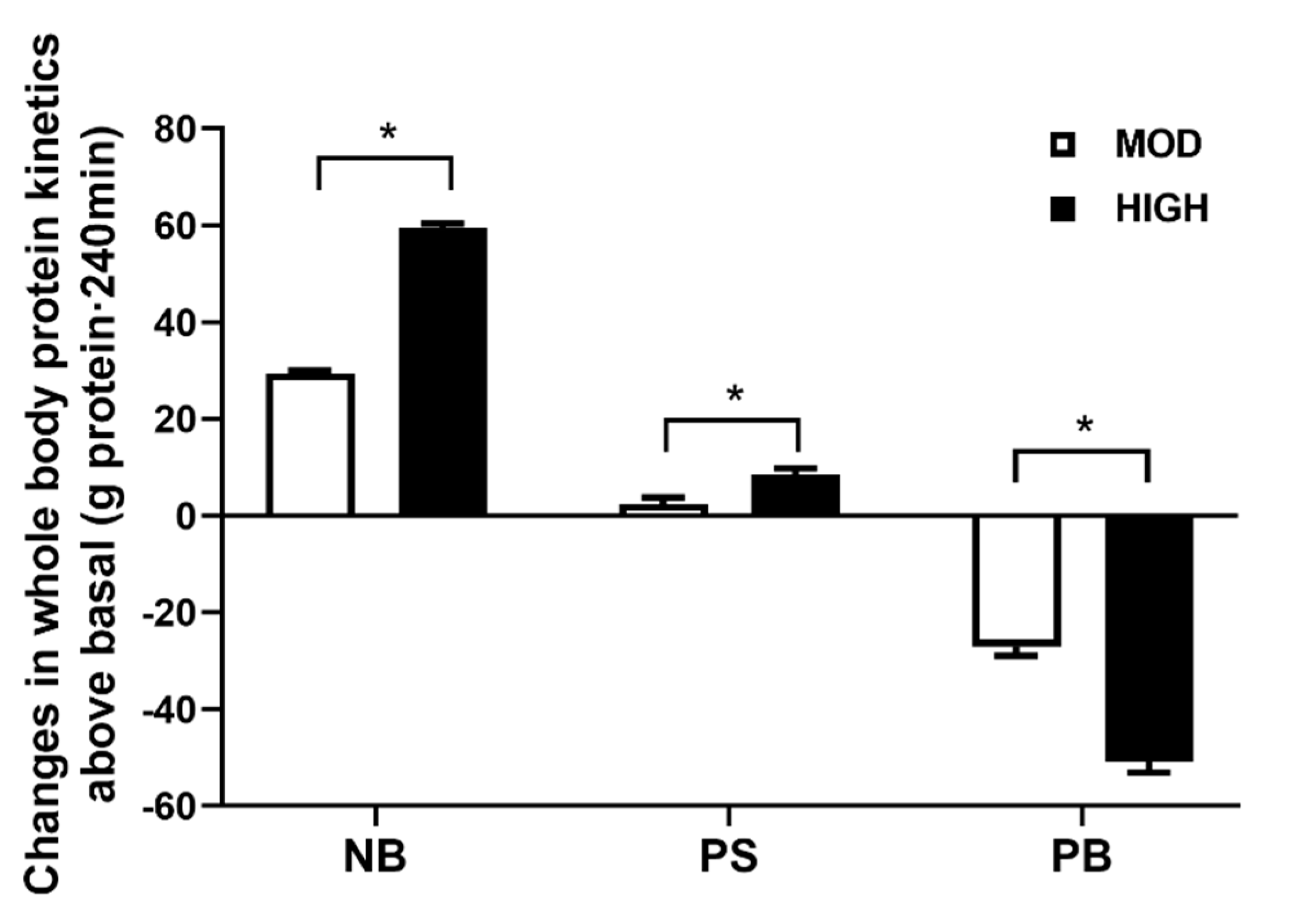
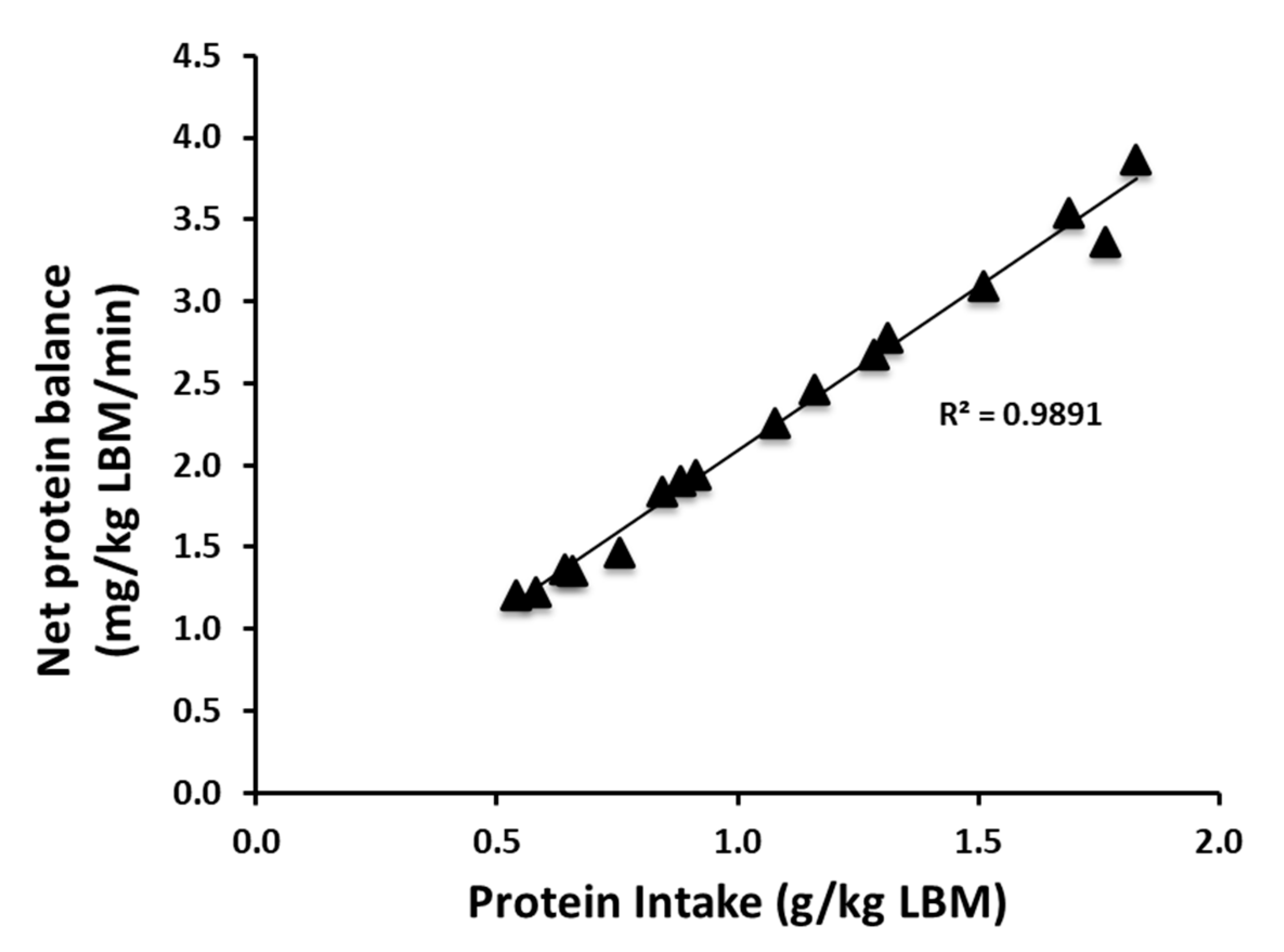
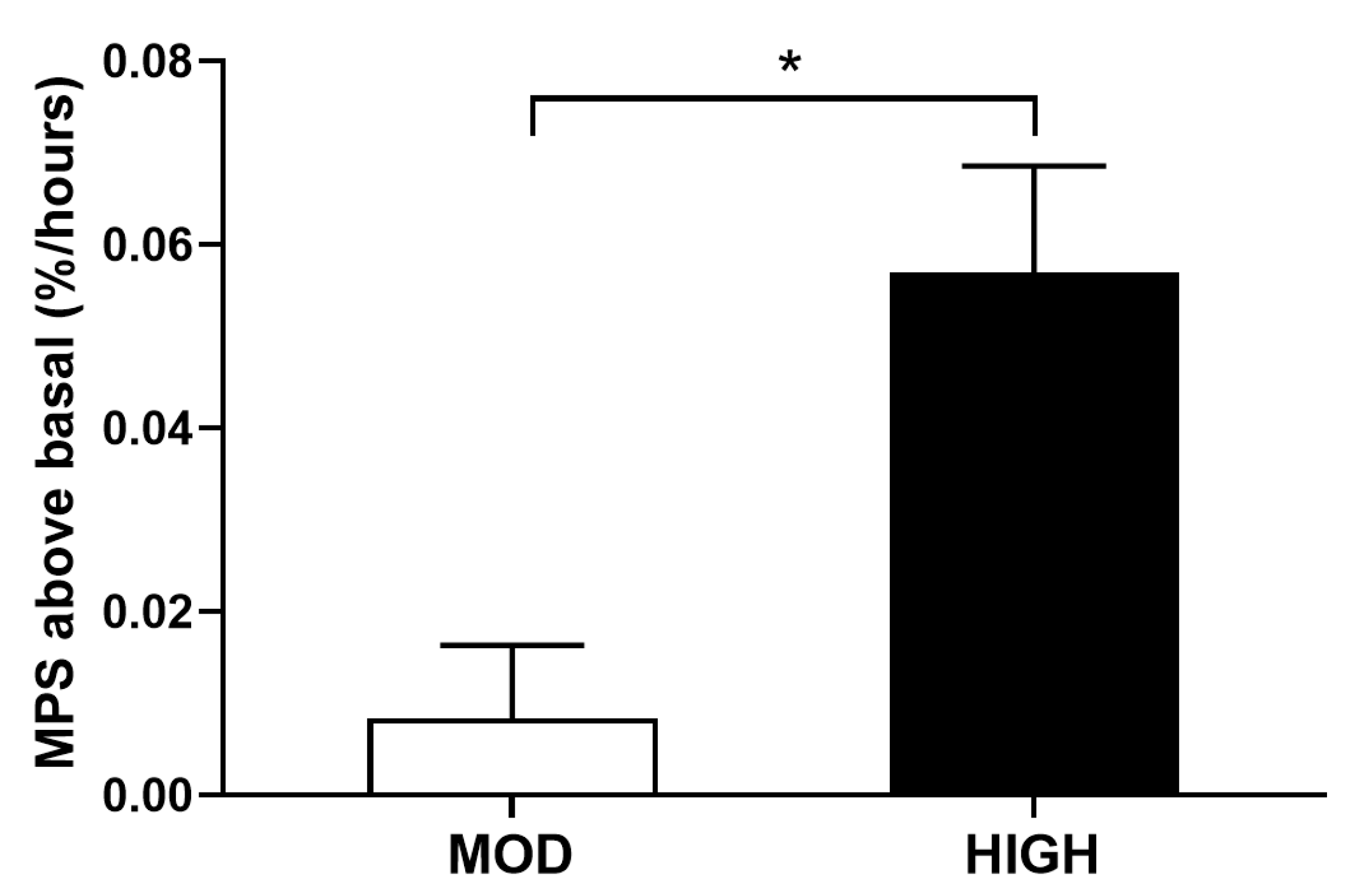
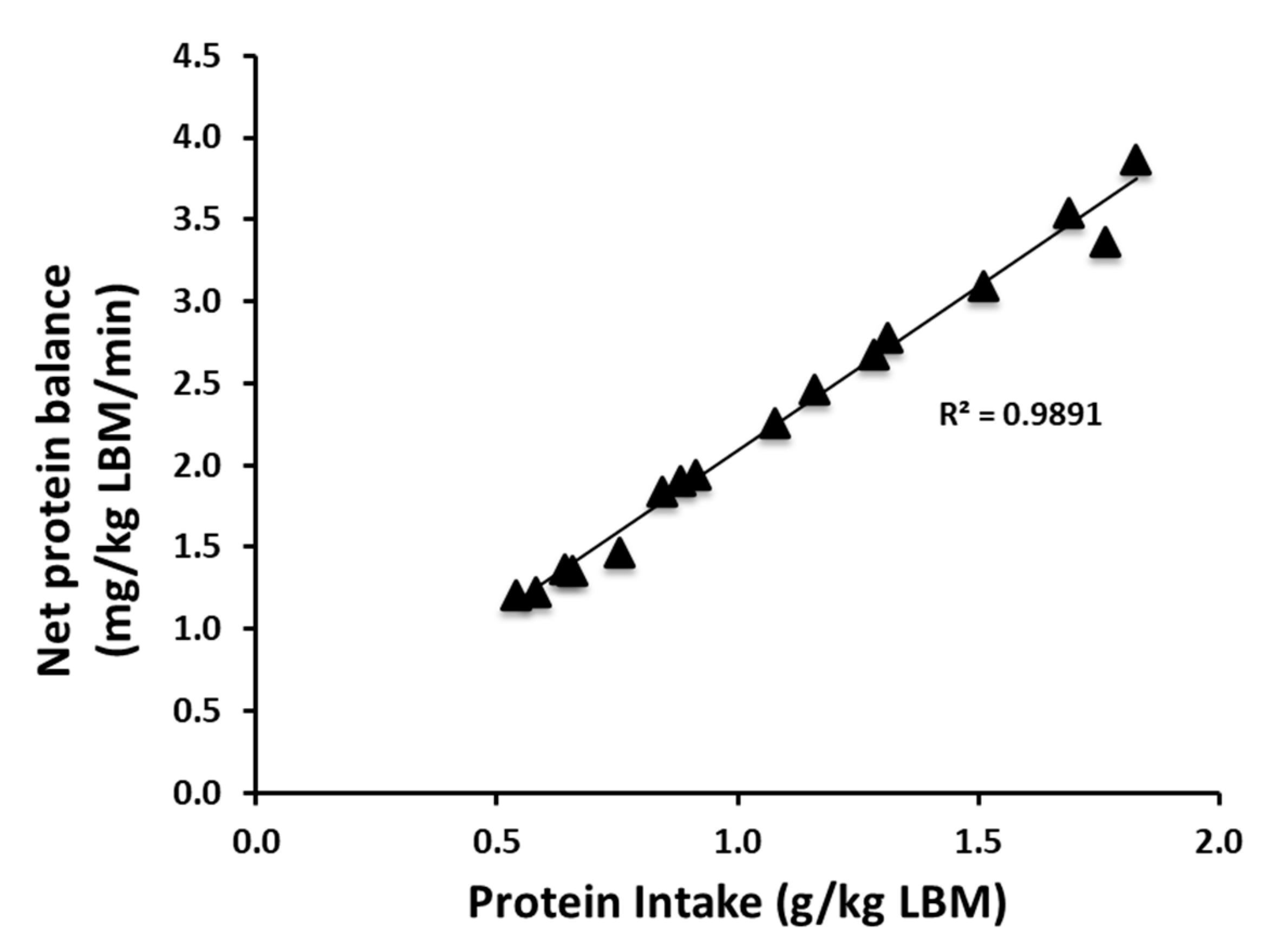
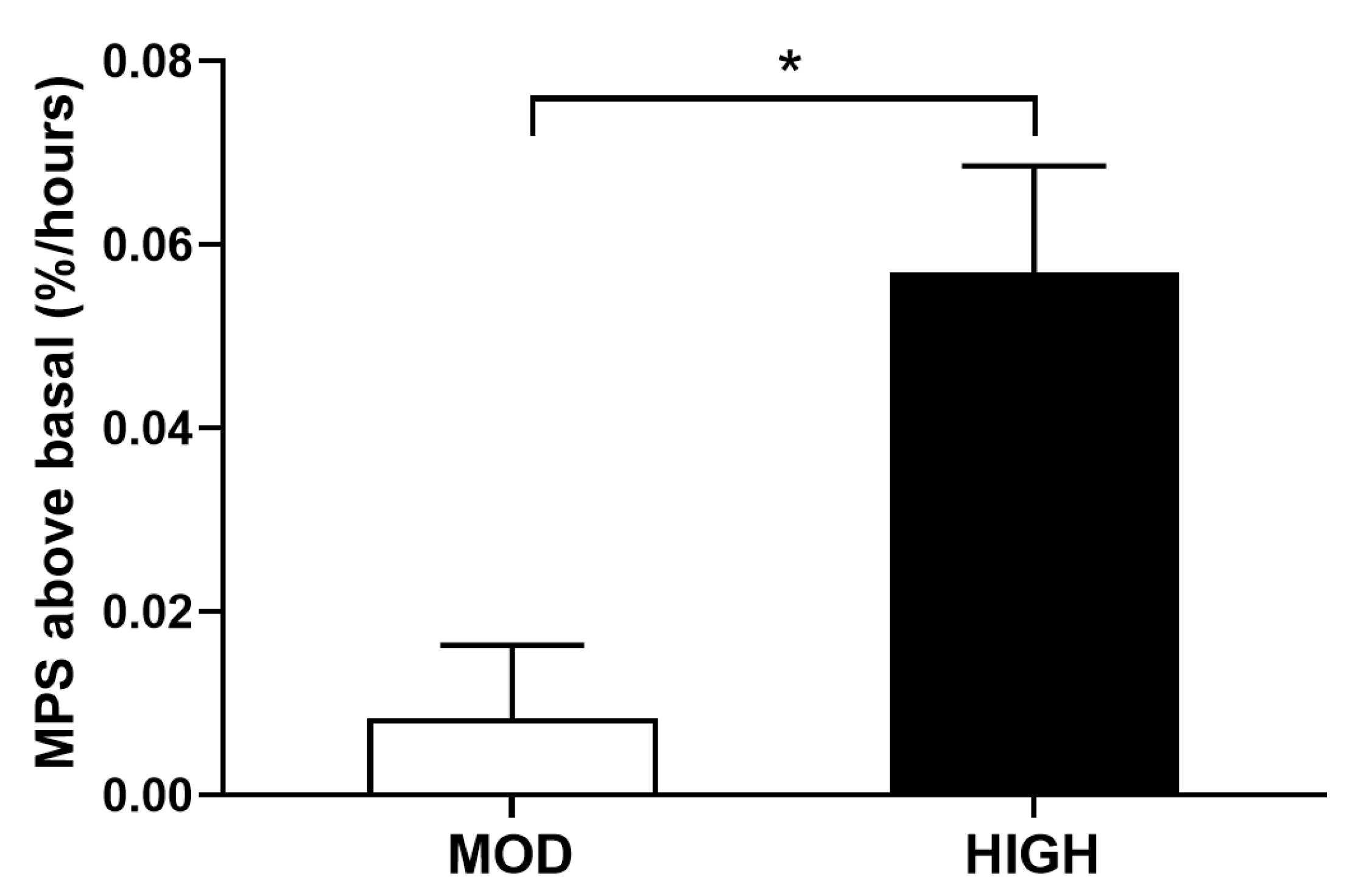
3.1. Protein Kinetics at Whole-Body and Muscle Levels
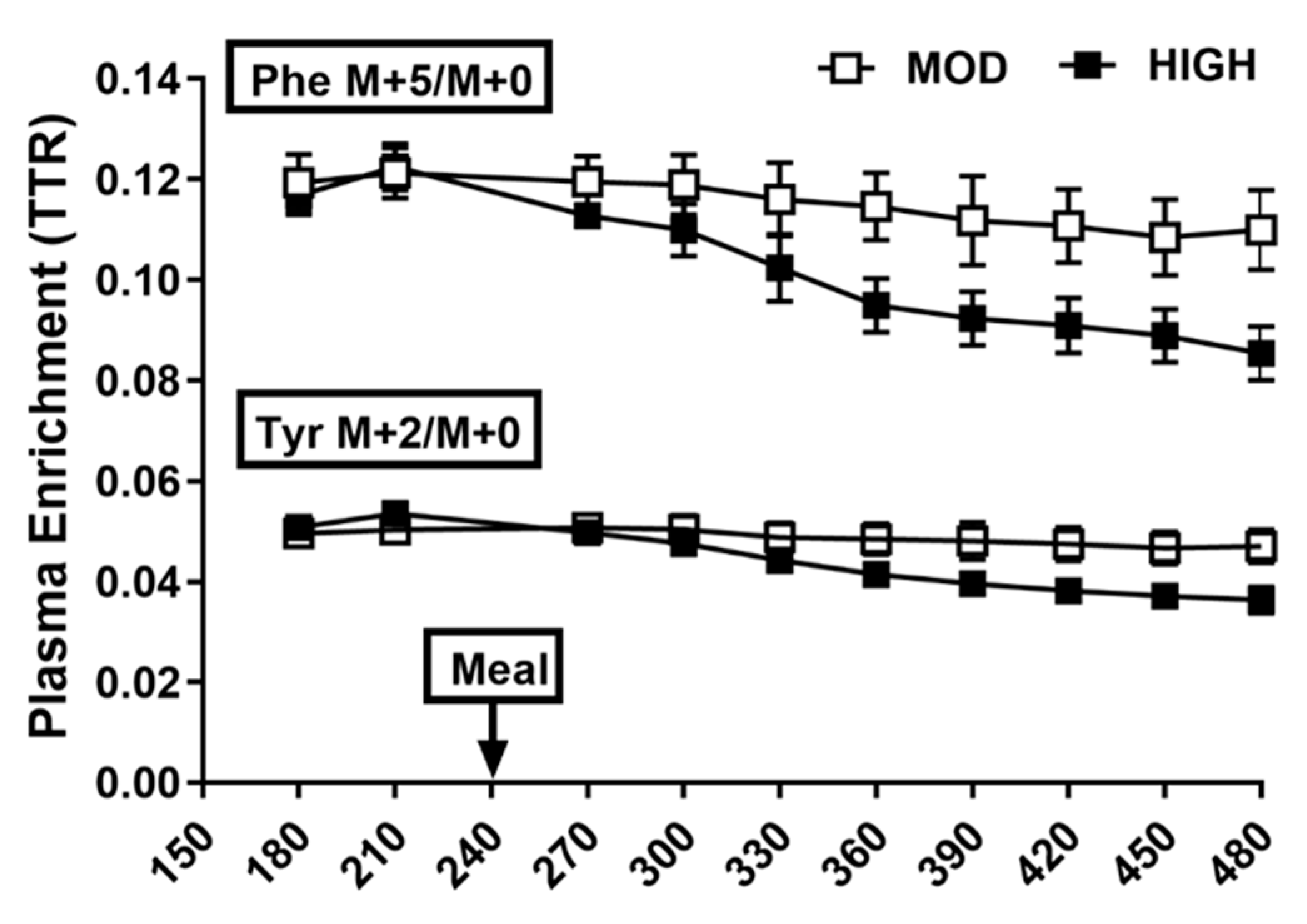
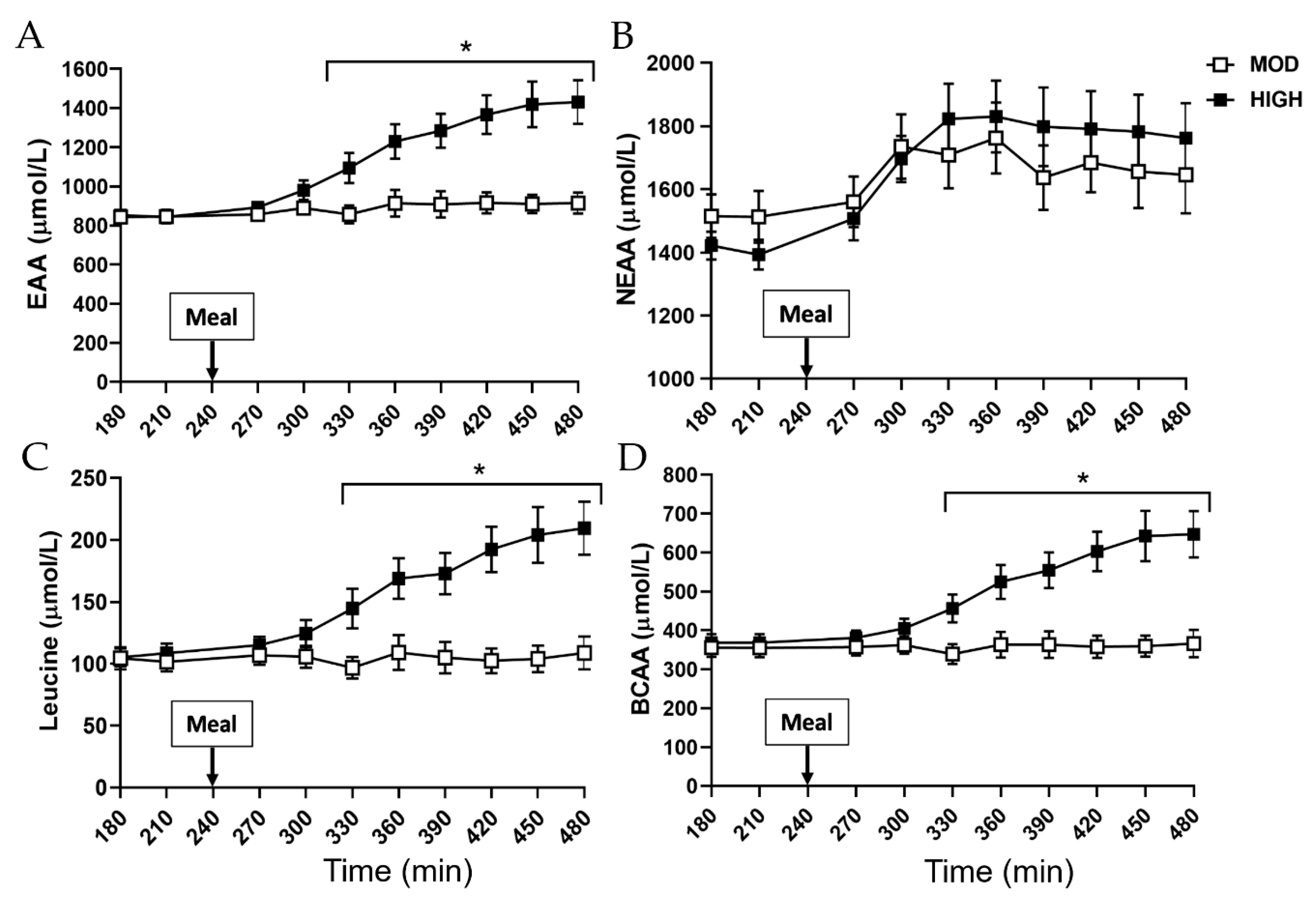
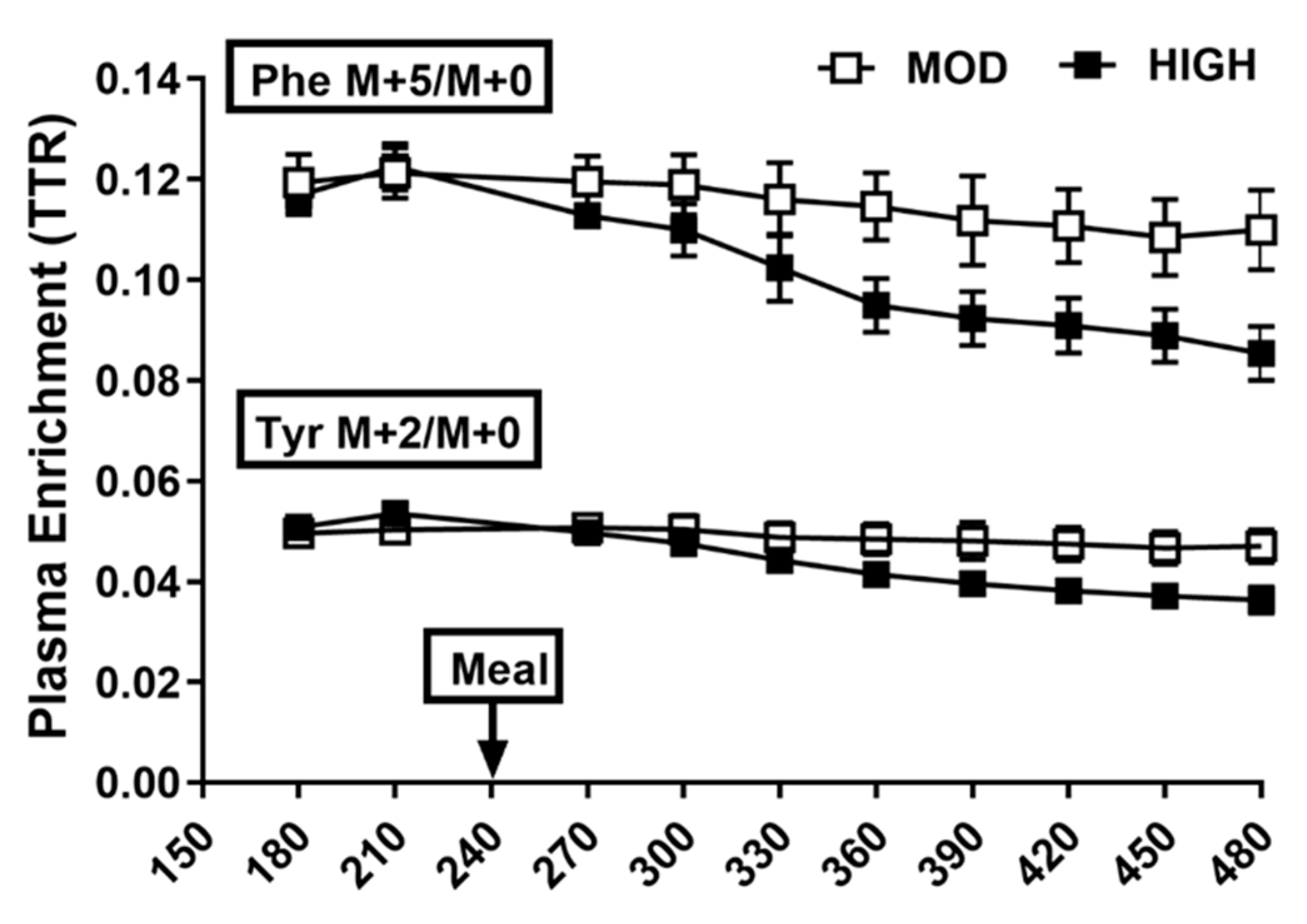
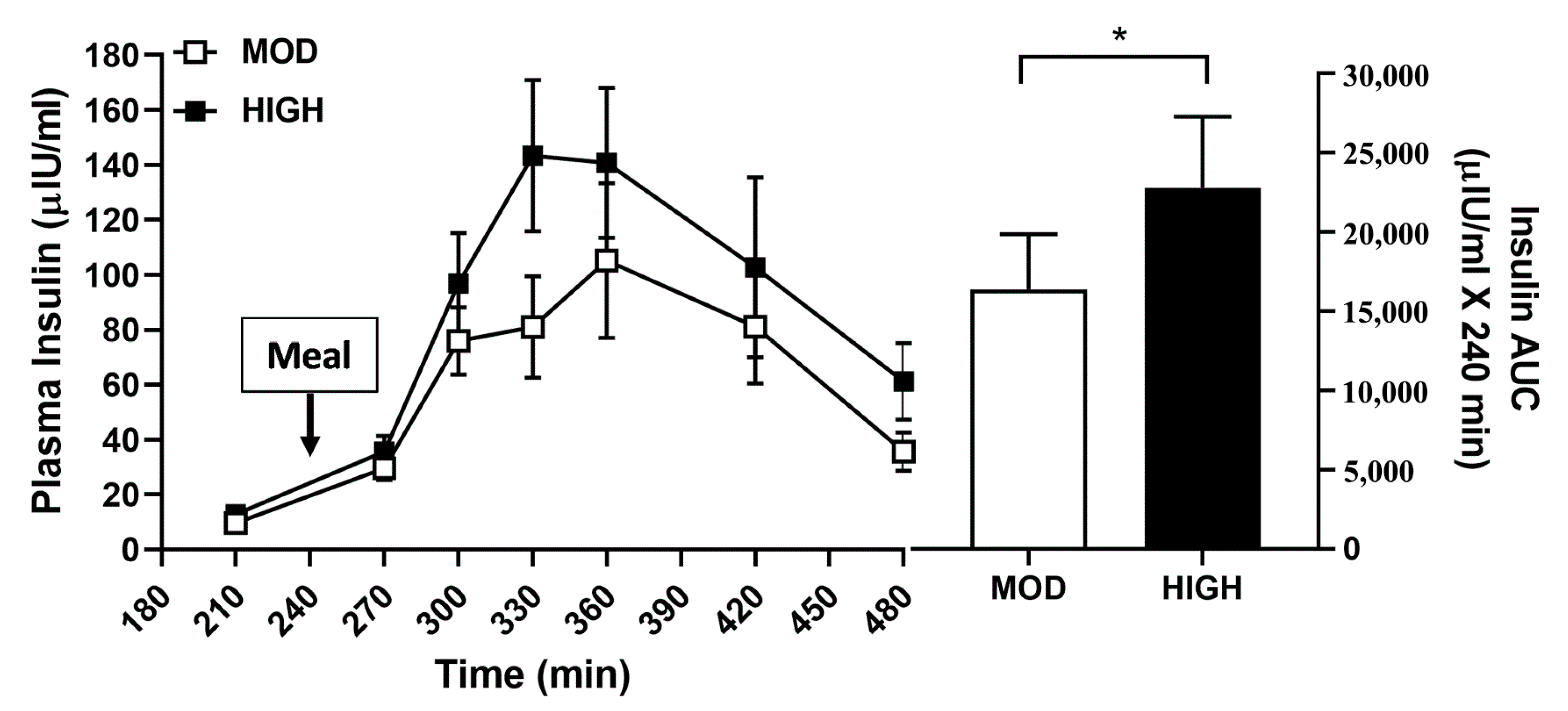
3.2. Plasma Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Evans, W.J. What is sarcopenia? J. Gerontol. A Biol. Sci. Med. Sci. 1995, 50, 5–8. [Google Scholar] [CrossRef]
- Lexell, J. Human aging, muscle mass, and fiber type composition. J. Gerontol. A Biol. Sci. Med. Sci. 1995, 50, 11–16. [Google Scholar] [PubMed]
- Volpi, E.; Kobayashi, H.; Sheffield-Moore, M.; Mittendorfer, B.; Wolfe, R.R. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am. J. Clin. Nutr. 2003, 78, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.R.; Miller, S.L.; Miller, K.B. Optimal protein intake in the elderly. Clin. Nutr. 2008, 27, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Volpi, E.; Mittendorfer, B.; Wolf, S.E.; Wolfe, R.R. Oral amino acids stimulate muscle protein anabolism in the elderly despite higher first-pass splanchnic extraction. Am. J. Physiol. 1999, 277, E513–E520. [Google Scholar] [CrossRef] [PubMed]
- Katsanos, C.S.; Kobayashi, H.; Sheffield-Moore, M.; Aarsland, A.; Wolfe, R.R. Aging is associated with diminished accretion of muscle proteins after the ingestion of a small bolus of essential amino acids. Am. J. Clin. Nutr. 2005, 82, 1065–1073. [Google Scholar] [CrossRef] [Green Version]
- Paddon-Jones, D.; Short, K.R.; Campbell, W.W.; Volpi, E.; Wolfe, R.R. Role of dietary protein in the sarcopenia of aging. Am. J. Clin. Nutr. 2008, 87, 1562S–1566S. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.R.; Churchward-Venne, T.A.; Witard, O.; Breen, L.; Burd, N.A.; Tipton, K.D.; Phillips, S.M. Protein ingestion to stimulate myofibrillar protein synthesis requires greater relative protein intakes in healthy older versus younger men. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.-Y.; Deutz, N.E.P.; Wolfe, R.R. Update on maximal anabolic response to dietary protein. Clin. Nutr. 2017. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Park, S.; Jang, J.; Wolfe, R.R. Understanding muscle protein dynamics: Technical considerations for advancing sarcopenia research. Ann. Geriatr. Med. Res. 2020. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Schutzler, S.; Schrader, A.M.; Spencer, H.J.; Azhar, G.; Wolfe, R.R.; Ferrando, A.A. Protein intake distribution pattern does not affect anabolic response, lean body mass, muscle strength or function over 8 weeks in older adults: A randomized-controlled trial. Clin. Nutr. 2018. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Shin, Y.-A.; Schutzler, S.E.; Azhar, G.; Wolfe, R.R.; Ferrando, A.A. Quality of meal protein determines anabolic response in older adults. Clin. Nutr. 2018. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Schutzler, S.; Schrader, A.; Spencer, H.J.; Azhar, G.; Ferrando, A.A.; Wolfe, R.R. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. Am. J. Physiol. Metab. 2016, 310, E73–E80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symons, T.B.; Sheffield-Moore, M.; Wolfe, R.R.; Paddon-Jones, D. A moderate serving of high-quality protein maximally stimulates skeletal muscle protein synthesis in young and elderly subjects. J. Am. Diet. Assoc. 2009, 109, 1582–1586. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.-Y.; Schutzler, S.; Schrader, A.; Spencer, H.; Kortebein, P.; Deutz, N.E.P.; Wolfe, R.R.; Ferrando, A.A. Quantity of dietary protein intake, but not pattern of intake, affects net protein balance primarily through differences in protein synthesis in older adults. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E21–E28. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-Y.; Suh, S.-H.; Lee, I.-K.; Wolfe, R.R. Applications of stable, nonradioactive isotope tracers in in vivo human metabolic research. Exp. Mol. Med. 2016, 48, e203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, R.R.; Chinkes, D.L. Isotope Tracers in Metabolic Research: Principles and Practice of Kinetic Analysis; Wiley-Liss: New York, NY, USA, 2004; ISBN 9780471462095. [Google Scholar]
- Oberli, M.; Marsset-Baglieri, A.; Airinei, G.; Santé-Lhoutellier, V.; Khodorova, N.; Rémond, D.; Foucault-Simonin, A.; Piedcoq, J.; Tomé, D.; Fromentin, G.; et al. High true ileal digestibility but not postprandial utilization of nitrogen from bovine meat protein in humans is moderately decreased by high-temperature, long-duration cooking. J. Nutr. 2015, 145, 2221–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deglaire, A.; Moughan, P.J. Animal models for determining amino acid digestibility in humans—A review. Br. J. Nutr. 2012, 108, S273–S281. [Google Scholar] [CrossRef] [Green Version]
- Gilani, S.; Tomé, D.; Moughan, P.; Burlingame, B. Report of a Sub-Committee of the 2011 FAO Consultation on “Protein Quality Evaluation in Human Nutrition” on: The Assessment of Amino Acid Digestibility in Foods for Humans and Including a Collation of Published Ileal Amino Acid Digestibility Data for Human Foods. FAO Expert. 2012. Available online: http://www.fao.org/ag/humannutrition/36216-04a2f02ec02eafd4f457dd2c9851b4c45.pdf (accessed on 21 October 2020).
- de Betue, C.T.; Joosten, K.F.; Deutz, N.E.; Vreugdenhil, A.C.; van Waardenburg, D.A. Arginine appearance and nitric oxide synthesis in critically ill infants can be increased with a protein-energy–enriched enteral formula. Am. J. Clin. Nutr. 2013, 98, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Deutz, N.E.; Have, G.A.T.; Soeters, P.B.; Moughan, P.J. Increased intestinal amino-acid retention from the addition of carbohydrates to a meal. Clin. Nutr. 1995, 14, 354–364. [Google Scholar] [CrossRef]
- Burd, N.A.; Gorissen, S.H.; Van Loon, L.J.C. Anabolic resistance of muscle protein synthesis with aging. Exerc. Sport Sci. Rev. 2013, 41, 169–173. [Google Scholar] [CrossRef]
- Pennings, B.; Groen, B.; de Lange, A.; Gijsen, A.P.; Zorenc, A.H.; Senden, J.M.G.; van Loon, L.J.C. Amino acid absorption and subsequent muscle protein accretion following graded intakes of whey protein in elderly men. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E992–E999. [Google Scholar] [CrossRef] [Green Version]
- Bohé, J.; Low, A.; Wolfe, R.R.; Rennie, M.J. Human muscle protein synthesis is modulated by extracellular, not intramuscular amino acid availability: A dose-response study. J. Physiol. 2003, 552, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Volpi, E.; Ferrando, A.A.; Yeckel, C.W.; Tipton, K.D.; Wolfe, R.R. Exogenous amino acids stimulate net muscle protein synthesis in the elderly. J. Clin. Investig. 1998, 101, 2000–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, R.R.; Miller, S.L. Amino acid availability controls muscle protein metabolism. Diabetes. Nutr. Metab. 1999, 12, 322–328. [Google Scholar]
- Boirie, Y.; Gachon, P.; Beaufrère, B. Splanchnic and whole-body leucine kinetics in young and elderly men. Am. J. Clin. Nutr. 1997, 65, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Biolo, G.; Fleming, R.Y.D.; Wolfe, R.R. Physiologic hyperinsulinemia stimulates protein synthesis and enhances transport of selected amino acids in human skeletal muscle. J. Clin. Investig. 1995, 95, 811–819. [Google Scholar] [CrossRef] [Green Version]
- Reitelseder, S.; Dideriksen, K.; Agergaard, J.; Malmgaard-Clausen, N.M.; Bechshoeft, R.L.; Petersen, R.K.; Serena, A.; Mikkelsen, U.R.; Holm, L. Even effect of milk protein and carbohydrate intake but no further effect of heavy resistance exercise on myofibrillar protein synthesis in older men. Eur. J. Nutr. 2018, 58, 583–595. [Google Scholar] [CrossRef]
- Greenhaff, P.L.; Karagounis, L.G.; Peirce, N.; Simpson, E.J.; Hazell, M.; Layfield, R.; Wackerhage, H.; Smith, K.; Atherton, P.; Selby, A.; et al. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am. J. Physiol. Metab. 2008, 295, E595–E604. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects (M/F) | 8 (4/4) |
| Age, year | 69.3 ± 1.8 |
| Weight, kg | 82.9 ± 4.9 |
| BMI, kg/m2 | 27.4 ± 0.9 |
| LBM, kg | 49.9 ± 3.5 |
| Fat mass, % | 34.5 ± 2.3 |
| Run-in Foods on Day 1–2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Protein Levels | Energy Intake, Kcal | Protein | Fat | CHO | ||||||
| g | % | g | % | g | % | |||||
| MOD | 2324 ± 135 | 83.1 ± 4.9 | 14.1 ± 0.3 | 91.0 ± 5.6 | 34.7 ± 0.3 | 301.0 ± 16.7 | 51.2 ± 0.2 | |||
| HIGH | 2328 ± 135 | 83.2 ± 4.8 | 14.1 ± 0.3 | 91.7 ± 5.6 | 34.9 ± 0.2 | 300.5 ± 16.9 | 51.0 ± 0.2 | |||
| Interventional Meals of Metabolic Infusion Study on Day 3 | ||||||||||
| Protein Levels | Meal, Kcal | Beef Protein, g | Protein | Fat | CHO | Nonprotein Energy, % | ||||
| g | % | g | % | g | % | Fat | CHO | |||
| MOD | 1100 | 23.6 | 35.7 | 12.9 | 41.3 | 33.6 | 147.9 | 53.5 | 38.6 | 61.4 |
| HIGH | 1100 | 59 | 70.3 | 25.9 | 36.2 | 30 | 119.6 | 44.1 | 40.5 | 59.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Jang, J.; Choi, M.D.; Shin, Y.-A.; Schutzler, S.; Azhar, G.; Ferrando, A.A.; Wolfe, R.R.; Kim, I.-Y. The Anabolic Response to Dietary Protein Is Not Limited by the Maximal Stimulation of Protein Synthesis in Healthy Older Adults: A Randomized Crossover Trial. Nutrients 2020, 12, 3276. https://doi.org/10.3390/nu12113276
Park S, Jang J, Choi MD, Shin Y-A, Schutzler S, Azhar G, Ferrando AA, Wolfe RR, Kim I-Y. The Anabolic Response to Dietary Protein Is Not Limited by the Maximal Stimulation of Protein Synthesis in Healthy Older Adults: A Randomized Crossover Trial. Nutrients. 2020; 12(11):3276. https://doi.org/10.3390/nu12113276
Chicago/Turabian StylePark, Sanghee, Jiwoong Jang, Myung Dong Choi, Yun-A Shin, Scott Schutzler, Gohar Azhar, Arny A. Ferrando, Robert R. Wolfe, and Il-Young Kim. 2020. "The Anabolic Response to Dietary Protein Is Not Limited by the Maximal Stimulation of Protein Synthesis in Healthy Older Adults: A Randomized Crossover Trial" Nutrients 12, no. 11: 3276. https://doi.org/10.3390/nu12113276